
Induction of Apoptosis and Antitumor Activity of Eel Skin Mucus, Containing Lactose-Binding Molecules, on Human Leukemic K562 Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
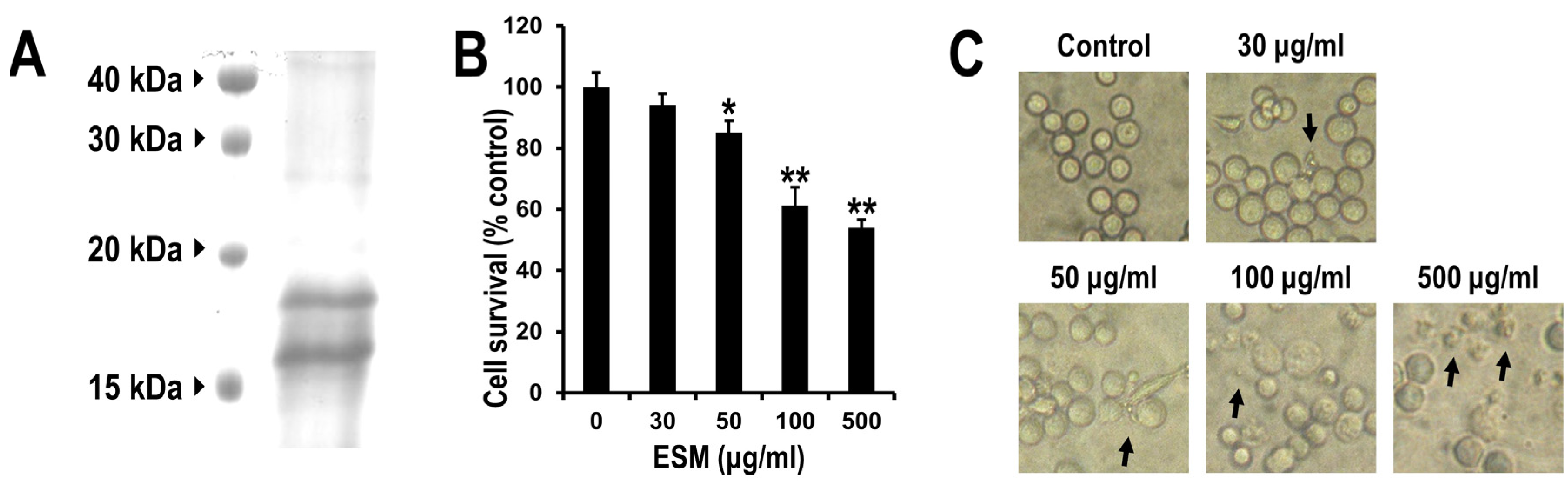
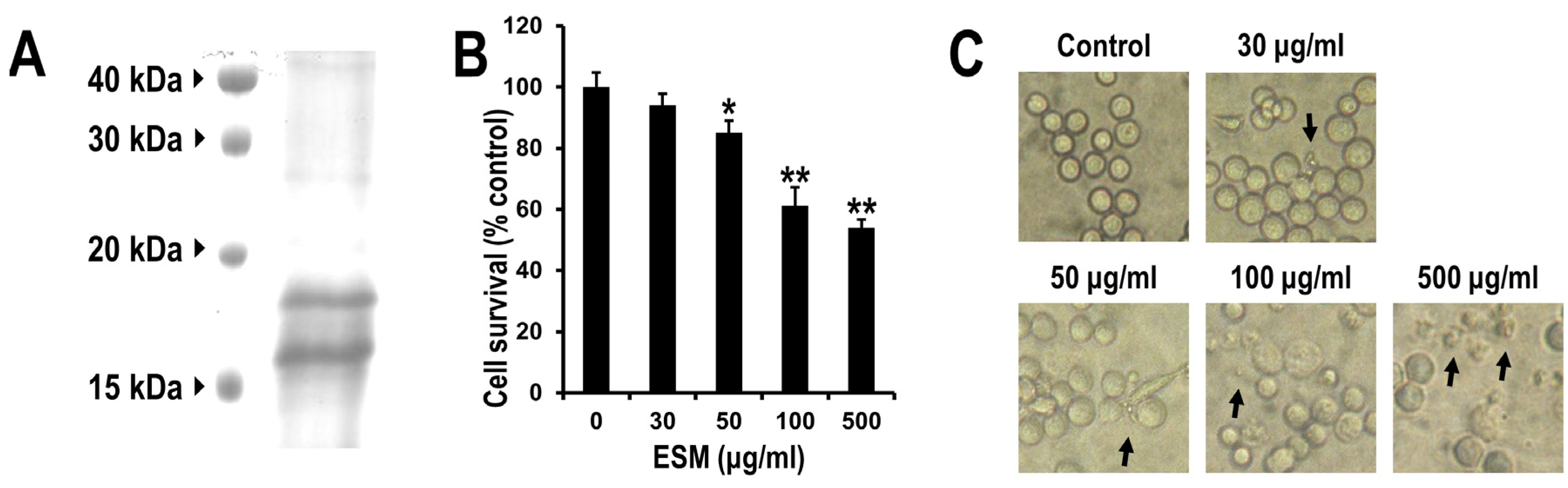
2.1. Reduction of Cell Viability of Leukemic K562 Cells by ESM
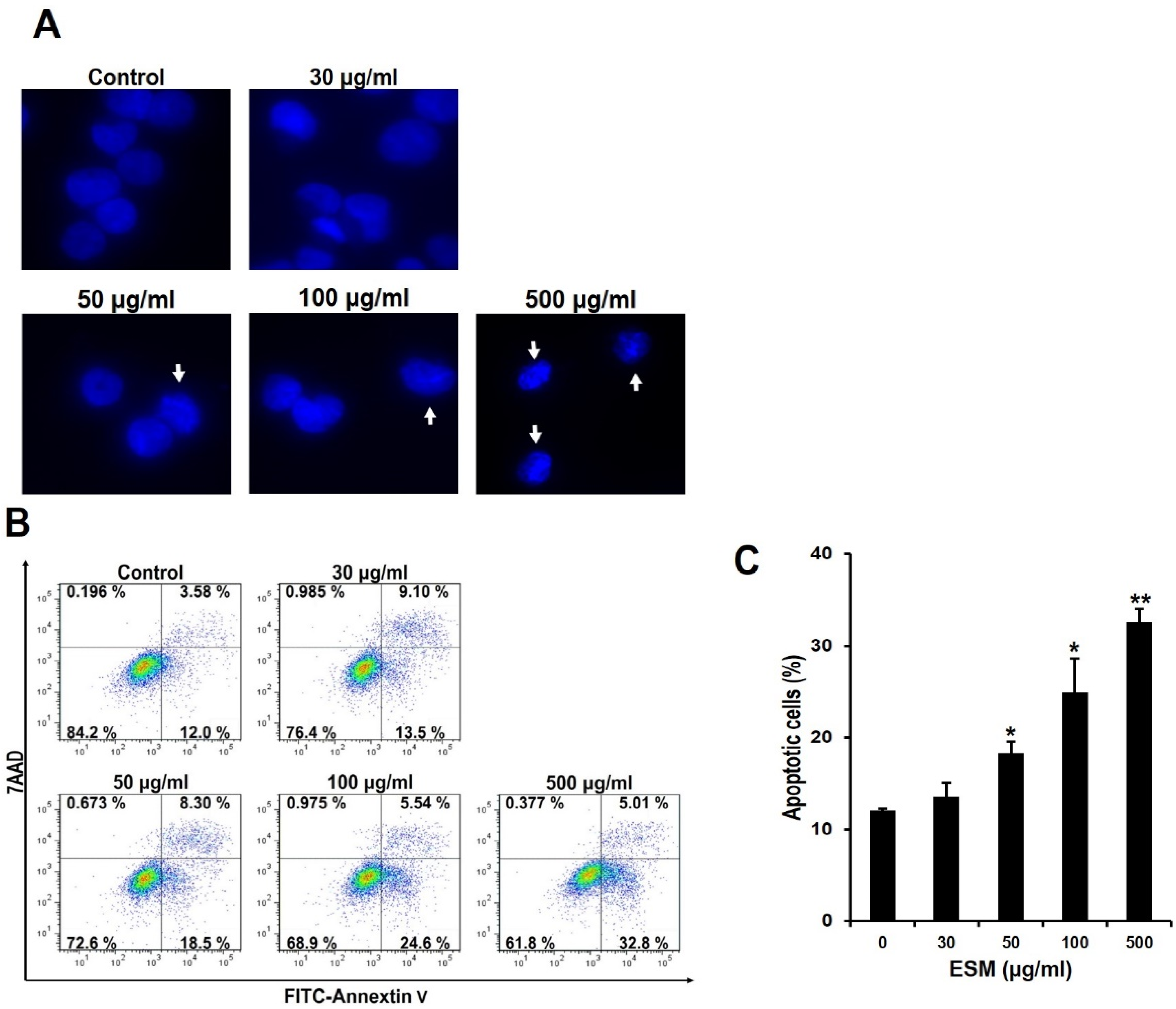
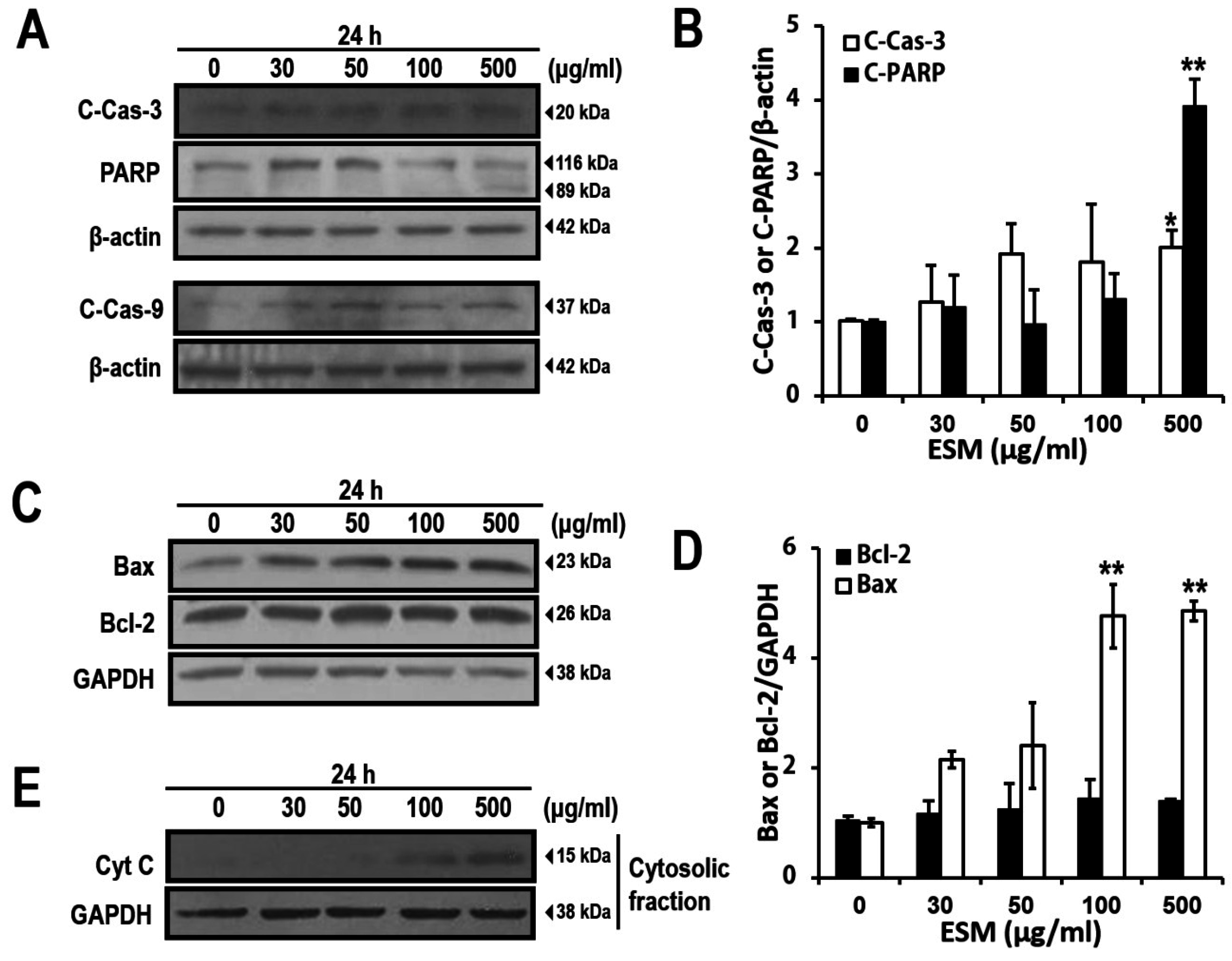
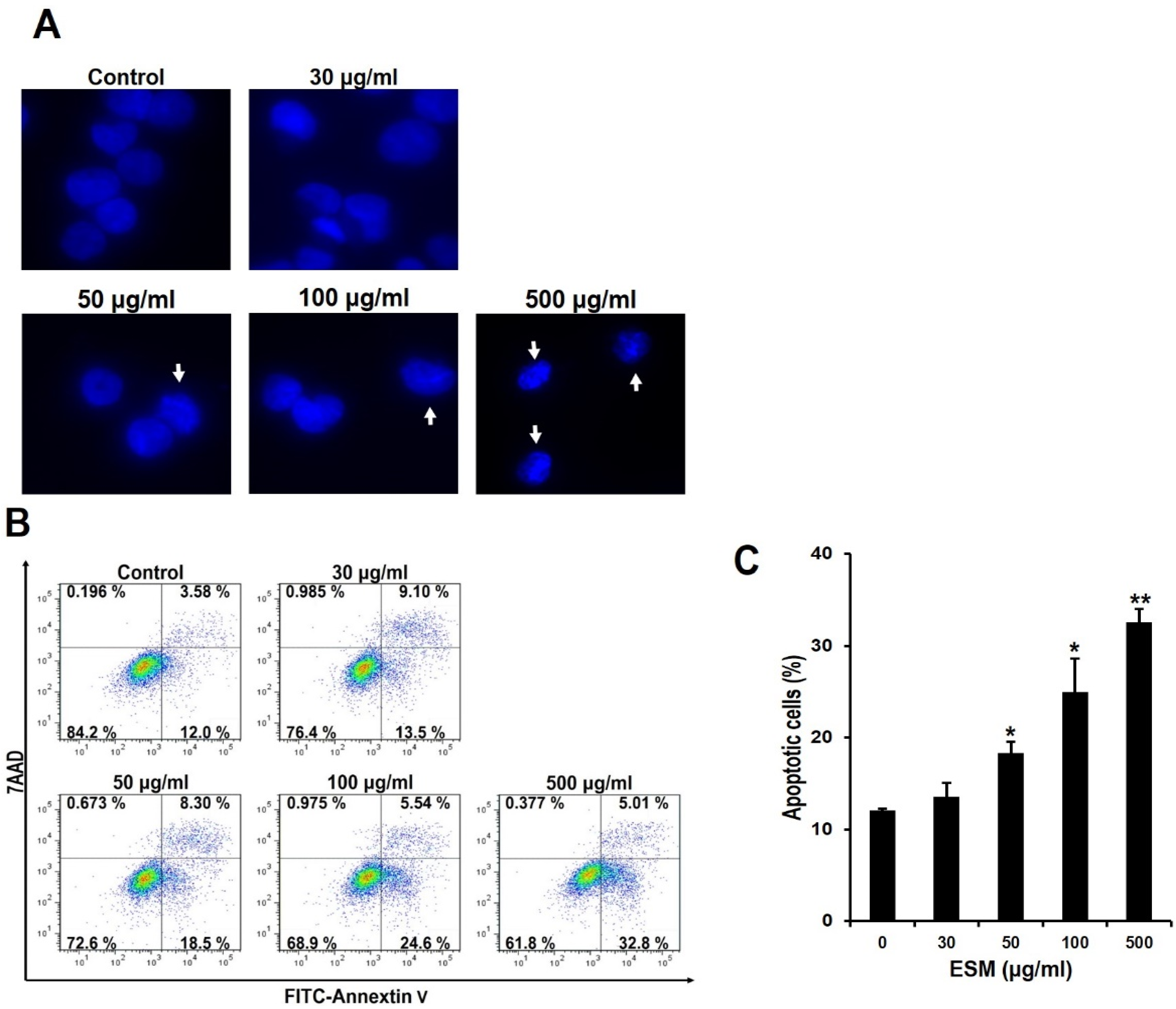
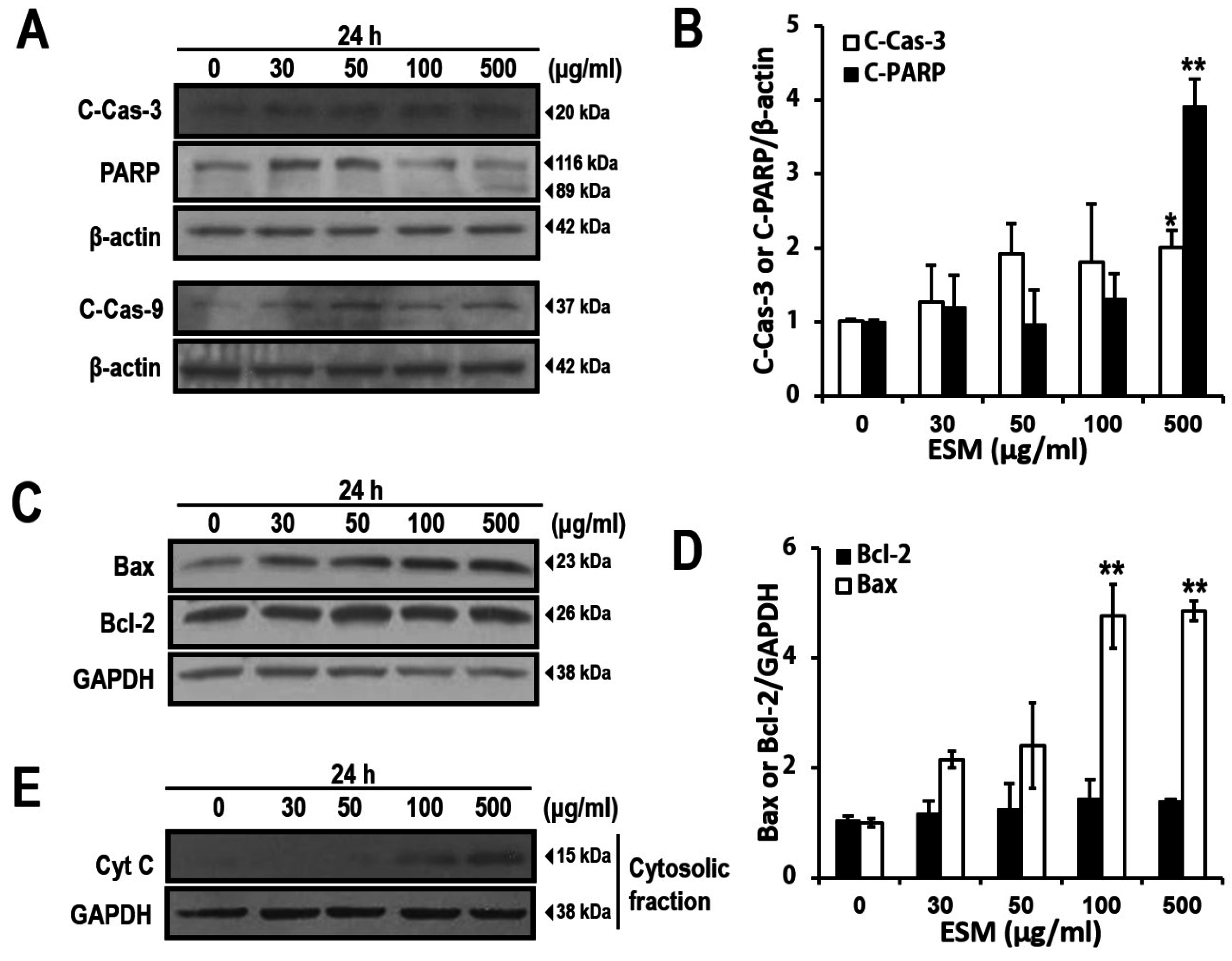
2.2. Induction of Apoptosis by ESM and Analysis of ESM-Mediated Signaling Pathway
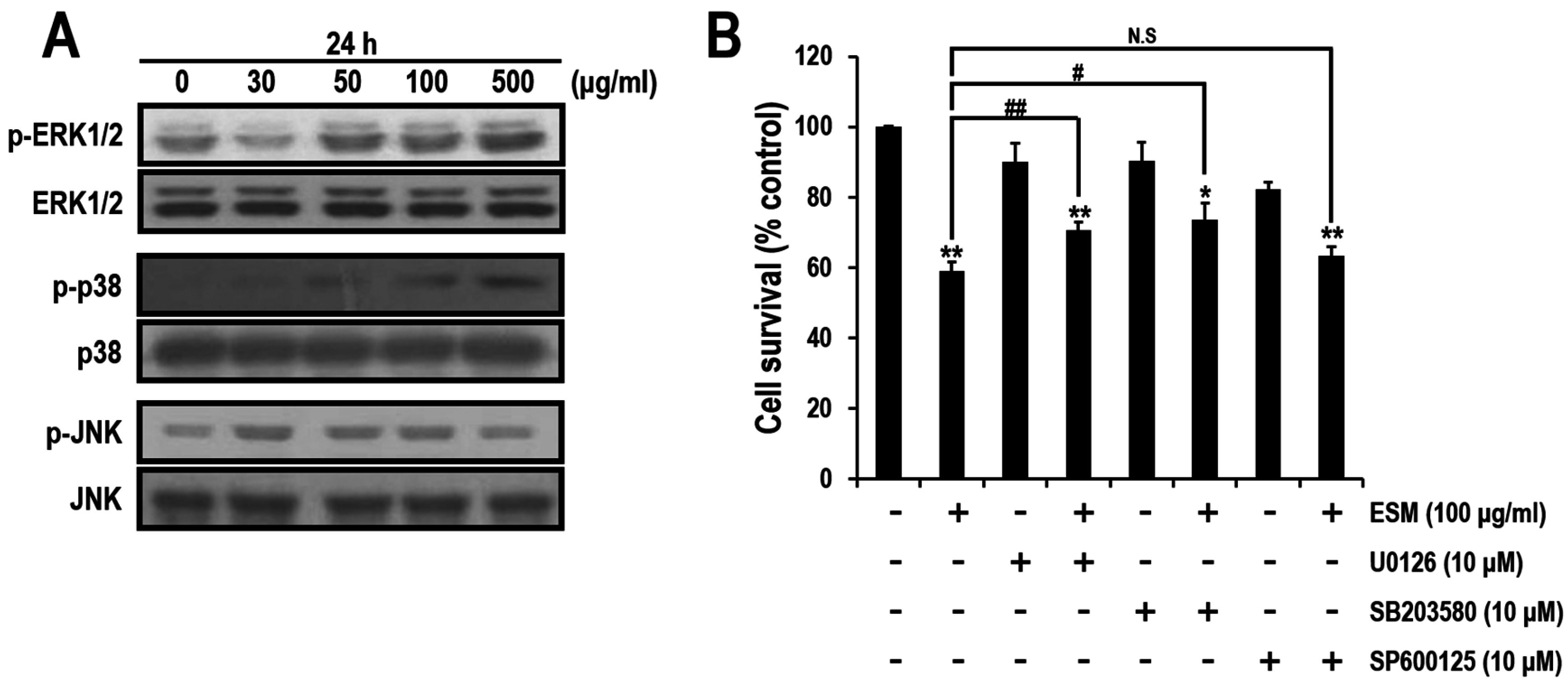
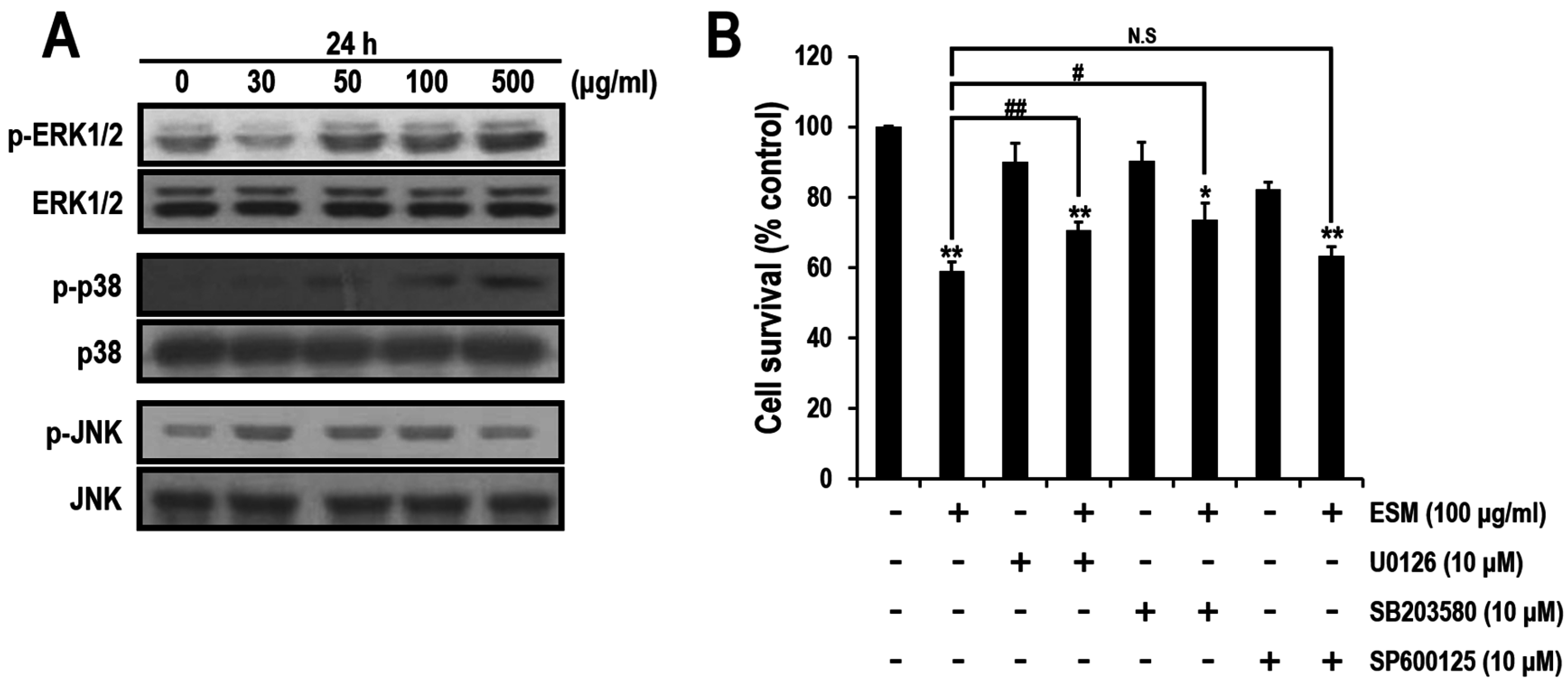
2.3. ERK and p38, But Not JNK Signals, Are Involved in ESM-Mediated Apoptosis
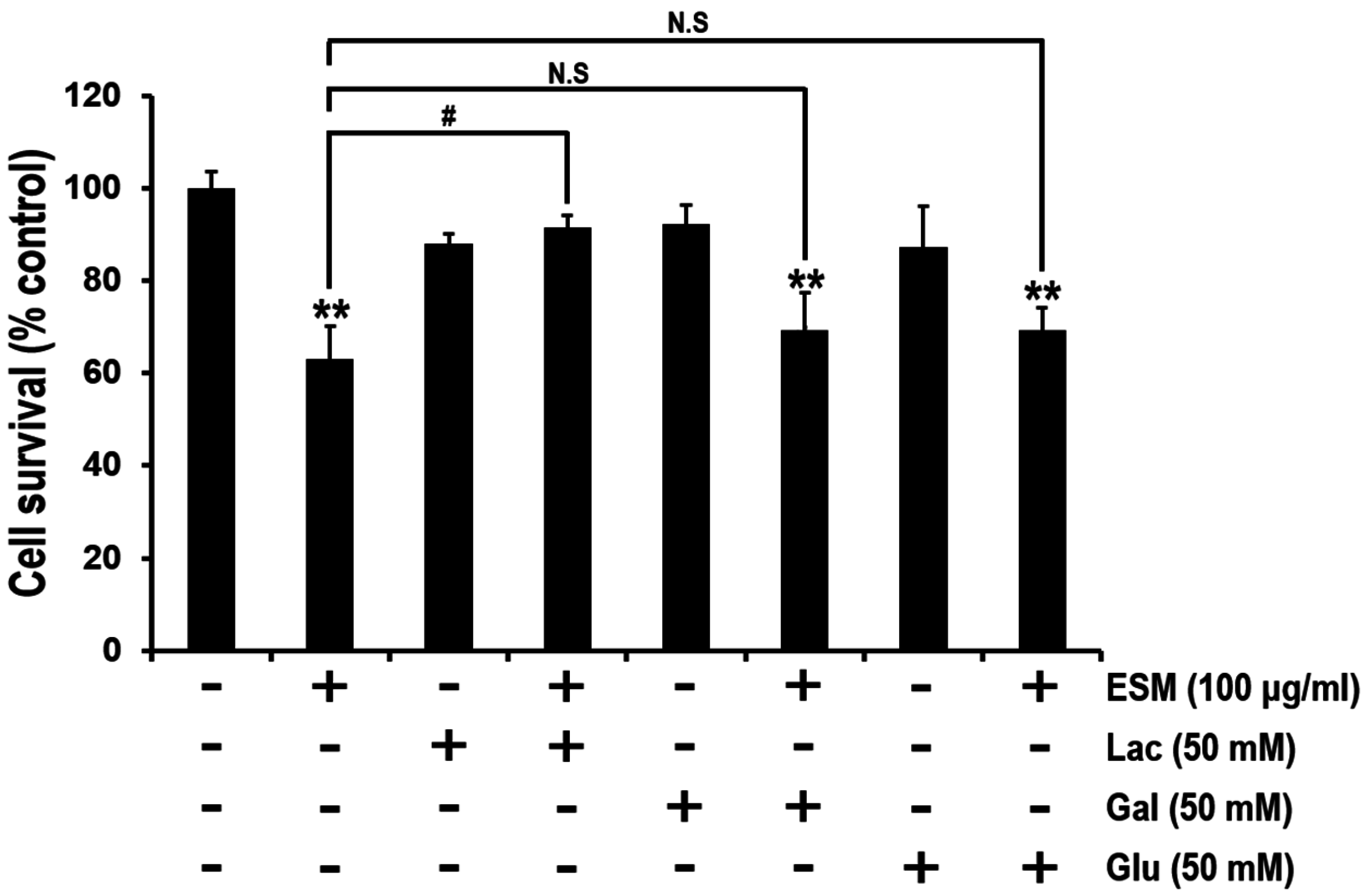
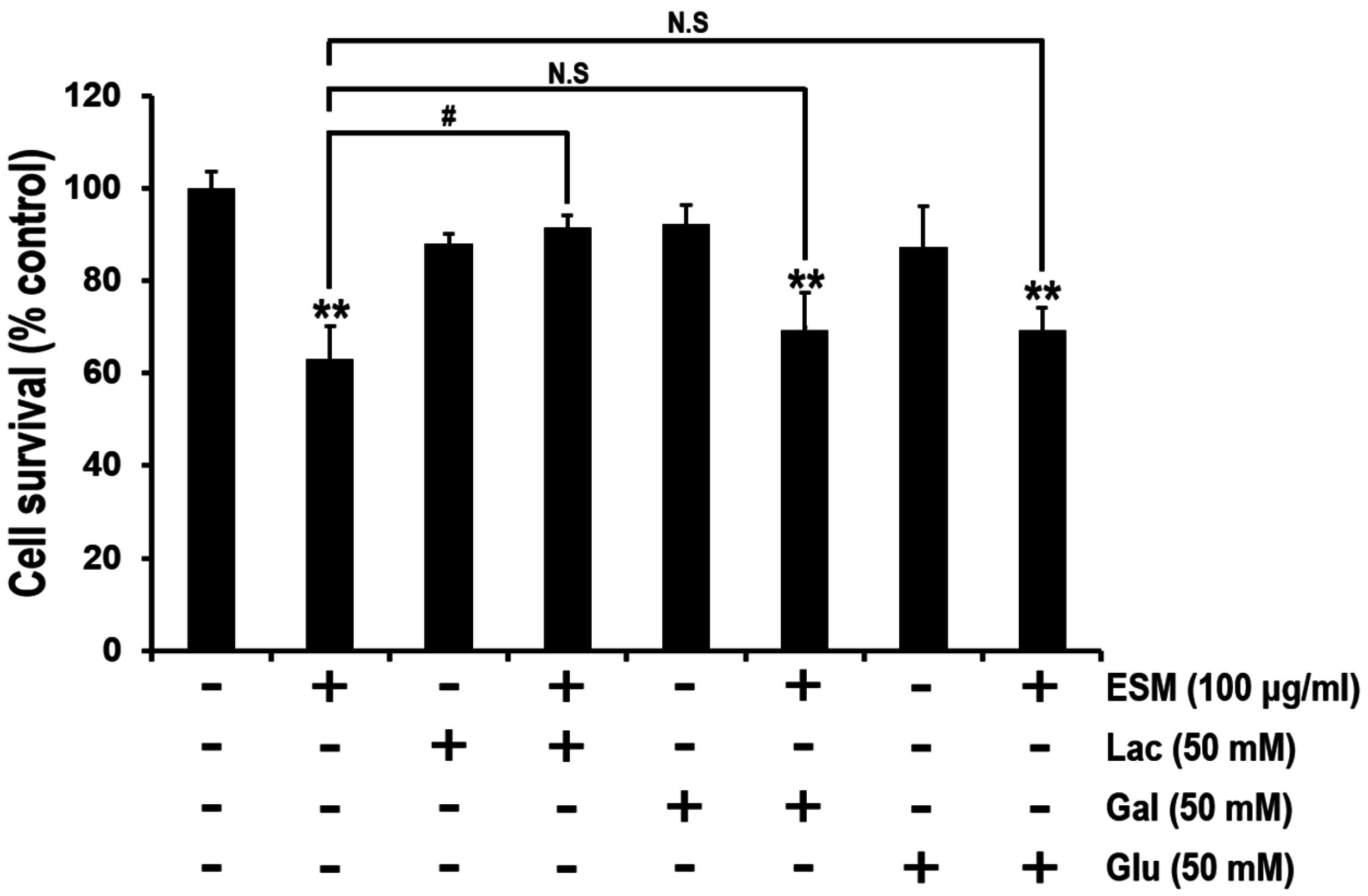
2.4. Lactose Suppresses Reduction of Cell Viability by ESM
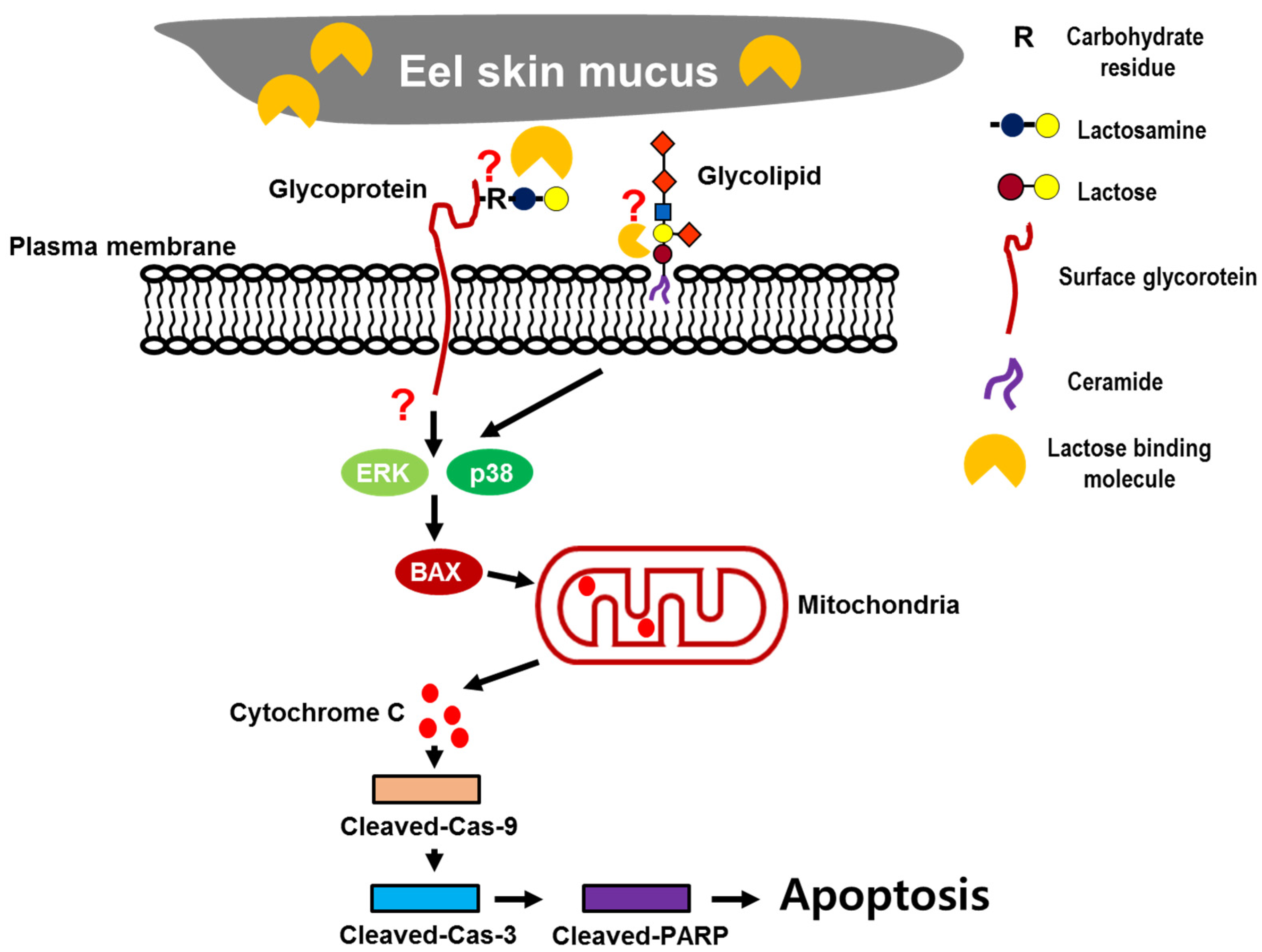
3. Discussion
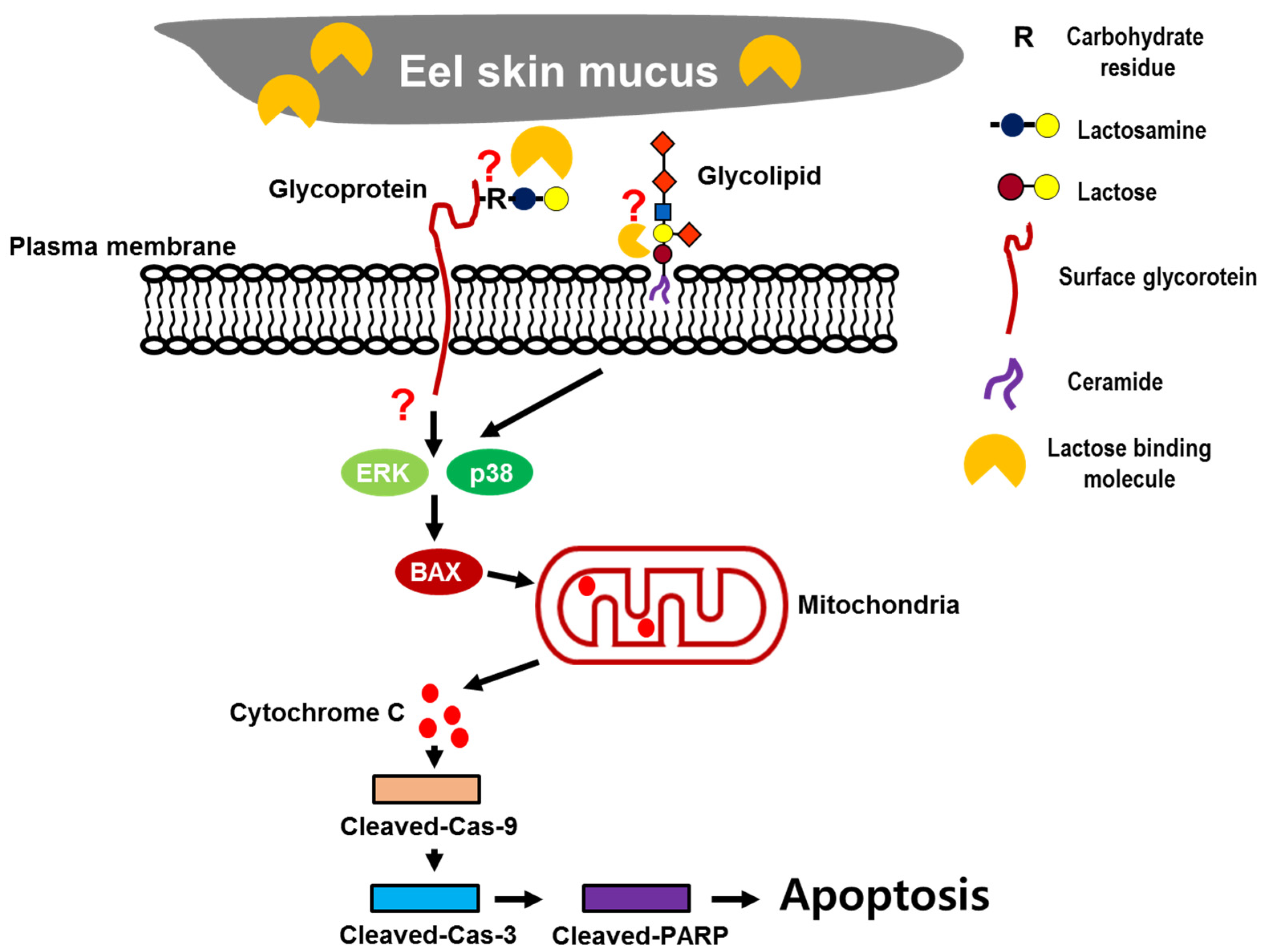
4. Materials and Methods
4.1. Preparation of Eel Skin Mucus
4.2. Cell Culture and Reagents
4.3. MTT Assay and Light Microscopic Analysis
4.4. Immunoblot and Densitometric Analysis
4.5. 4′,6-Diamidino-2-phenylindole (DAPI) Staining
4.6. Annexin V-FITC/7-Amino-Actinomycin (7AAD) Staining and Analysis by Flow Cytometry
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nasr, R.; Bazarbachi, A. Chronic myeloid leukemia: “Archetype” of the impact of targeted therapies. Pathol. Biol. 2012, 60, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, P.; Di Persio, J.F. Therapy options in imatinib failures. Oncologist 2008, 13, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Long, J.S.; Ryan, K.M. New frontiers in promoting tumour cell death: Targeting apoptosis, necropdtosis and autophagy. Oncogene 2012, 31, 5045–5060. [Google Scholar] [CrossRef] [PubMed]
- Jin, U.H.; Song, K.H.; Motomura, M.; Suzuki, I.; Gu, Y.H.; Kang, Y.J.; Moon, T.C.; Kim, C.H. Caffeic acid phenethyl ester induces mitochondria-mediated apoptosis in human myeloid leukemia U937 cells. Mol. Cell. Biochem. 2008, 310, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.Y.; Kang, S.K.; Lee, Y.C.; Choi, H.J.; Lee, Y.S.; Cho, S.Y.; Kim, Y.S.; Ko, J.H.; Kim, C.H. Transcriptional regulation of the human GD3 synthase gene expression in Fas-induced Jurkat T cells: A critical role of transcription factor NF-kappaB in regulated expression. Glycobiology 2006, 16, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.K.; Kim, H.M.; Lee, Y.C.; Kim, C.H. Disialoganglioside (GD3) synthase gene expression suppresses vascular smooth muscle cell responses via the inhibition of ERK1/2 phosphorylation, cell cycle progression, and matrix metalloproteinase-9 expression. J. Biol. Chem. 2004, 279, 33063–33070. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Kim, K.S.; An, H.K.; Kim, C.H.; Moon, H.I.; Lee, Y.C. Dendropanoxide induces autophagy through ERK1/2 activation in MG-63 human osteosarcoma cells and autophagy inhibition enhances dendropanoxide-induced apoptosis. PLoS ONE 2013, 8, e83611. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Guan, R.; Huang, W.; Xu, T. Isolation and identification of a novel inducible antibacterial peptide from the skin mucus of Japanese eel, Anguilla japonica. Protein J. 2011, 30, 413–421. [Google Scholar]
- Suzuki, Y.; Tasumi, S.; Tsutsui, S.; Okamoto, M.; Suetake, H. Molecular diversity of skin mucus lectins in fish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 723–730. [Google Scholar] [CrossRef]
- Odom, E.W.; Vasta, G.R. Characterization of a binary tandem domain F-type lectin from striped bass (Morone saxatilis). J. Biol. Chem. 2006, 281, 1698–1713. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Nakamura, O.; Muramoto, K.; Ogawa, T. Allosteric regulation of the carbohydrate-binding ability of a novel conger eel galectin by d-mannoside. J. Biol. Chem. 2012, 287, 31061–31072. [Google Scholar] [CrossRef] [PubMed]
- Dirks, R.P.; Burgerhout, E.; Brittijn, S.A.; de Wijze, D.L.; Ozupek, H.; Tuinhof-Koelma, N.; Minegishi, Y.; Jong-Raadsen, S.A.; Spaink, H.P.; van den Thillart, G.E. Identification of molecular markers in pectoral fin to predict artificial maturation of female European eels (Anguilla anguilla). Gen. Comp. Endocrinol. 2014, 204, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Tasumi, S.; Ohira, T.; Kawazoe, I.; Suetake, H.; Suzuki, Y.; Aida, K. Primary structure and characteristics of a lectin from skin mucus of the Japanese eel Anguilla japonica. J. Biol. Chem. 2002, 277, 27305–27311. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, S.; Iwamoto, K.; Nakamura, O.; Watanabe, T. Yeast-binding C-type lectin with opsonic activity from conger eel (Conger myriaster) skin mucus. Mol. Immunol. 2007, 44, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Perillo, N.L.; Uittenbogaart, C.H.; Nguyen, J.T.; Baum, L.G. Galectin-1, an endogenous lectin produced by thymic epithelial cells, induces apoptosis of human thymocytes. J. Exp. Med. 1997, 185, 1851–1858. [Google Scholar] [CrossRef] [PubMed]
- Pace, K.E.; Lee, C.; Stewart, P.L.; Baum, L.G. Restricted receptor segregation into membrane microdomains occurs on human T cells during apoptosis induced by galectin-1. J. Immunol. 1999, 163, 3801–3811. [Google Scholar] [PubMed]
- Clausse, N.; van den Brûle, F.; Waltregny, D.; Garnier, F.; Castronovo, V. Galectin-1 expression in prostate tumor-associated capillary endothelial cells is increased by prostate carcinoma cells and modulates heterotypic cell-cell adhesion. Angiogenesis 1999, 3, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Stillman, B.N.; Hsu, D.K.; Pang, M.; Brewer, C.F.; Johnson, P.; Liu, F.T.; Baum, L.G. Galectin-3 and galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J. Immunol. 2006, 176, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Na, M.K.; Suh, S.J.; Li, X.; Kim, G.J.; Chao, G.H.; Jeong, Y.T.; Kim, D.S.; Chang, Y.C.; Murakami, M.; et al. Anti-inflammatory activity of hexane extracts from bones and internal organs of Anguilla japonica suppresses cyclooxygenase-2-dependent prostaglandin D2 generation in mast cells and anaphylaxis in mice. Food Chem. Toxicol. 2013, 57, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.T.; Yang, J.H.; Li, X.; Kim, G.J.; Kim, D.S.; Kim, C.H.; Na, M.K.; Chang, H.W. Ethylacetate extracts of the muscles of Anguilla japonica suppress glucose levels in db/db mice via activation of AMP-activated protein kinase. Food Sci. Biotechnol. 2014, 23, 1695–1701. [Google Scholar] [CrossRef]
- Boldt, S.; Weidle, U.H.; Kolch, W. The role of MAPK pathways in the action of chemotherapeutic drugs. Carcinogenesis 2002, 23, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Su, C.C.; Chen, J.Y.; Din, Z.H.; Su, J.H.; Yang, Z.Y.; Chen, Y.J.; Wang, R.Y.; Wu, Y.J. 13-acetoxysarcocrassolide induces apoptosis on human gastric carcinoma cells through mitochondria-related apoptotic pathways: p38/JNK activation and PI3K/AKT suppression. Mar. Drugs 2014, 12, 5295–5315. [Google Scholar] [CrossRef] [PubMed]
- Mistry, A.C.; Honda, S.; Hirose, S. Structure, properties and enhanced expression of galactose-binding C-type lectins in mucous cells of gills from freshwater Japanese eels (Anguilla japonica). Biochem. J. 2001, 360 Pt 1, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Osonoi, T.; Terauchi, Y. Relationship between urinary sodium excretion and pioglitazone-induced edema. J. Diabetes Investig. 2010, 1, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.T. Purification and properties of lung lectin. Rat lung and human lung beta-galactoside-binding proteins. Biochem. J. 1980, 187, 123–129. [Google Scholar] [PubMed]
- Rabinovich, G.A.; Croci, D.O. Regulatory circuits mediated by lectin-glycan interactions in autoimmunity and cancer. Immunity 2012, 36, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Muramoto, K.; Kamiya, H. The amino-acid sequence of a lectin from conger eel, Conger myriaster, skin mucus. Biochim. Biophys. Acta 1992, 1116, 129–136. [Google Scholar] [CrossRef]
- Muramoto, K.; Kagawa, D.; Sato, T.; Ogawa, T.; Nishida, Y.; Kamiya, H. Functional and structural characterization of multiple galectins from the skin mucus of conger eel, Conger myriaster. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1999, 123, 33–45. [Google Scholar] [CrossRef]
- Tasumi, S.; Yang, W.J.; Usami, T.; Tsutsui, S.; Ohira, T.; Kawazoe, I.; Wilder, M.N.; Aida, K.; Suzuki, Y. Characteristics and primary structure of a galectin in the skin mucus of the Japanese eel, Anguilla japonica. Dev. Comp. Immunol. 2004, 28, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Saitoh, E.; Kimizuka, R.; Yamada, S.; Kato, T. Effect of eel galectin AJL-1 on periodontopathic bacterial biofilm formation and their lipopolysaccharide-mediated inflammatory cytokine induction. Int. J. Antimicrob. Agents 2009, 34, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Bonni, A.; Brunet, A.; West, A.E.; Datta, S.R.; Takasu, M.A.; Greenberg, M.E. Cell survival promoted by the Ras-MAPK signaling pathway by transcription-dependent and -independent mechanisms. Science 1999, 286, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M.W.; Norris, J.L.; Baldwin, A.S. Ras regulation of NF-kappa B and apoptosis. Methods Enzymol. 2001, 333, 73–87. [Google Scholar] [PubMed]
- Cagnol, S.; Chambard, J.C. ERK and cell death: mechanisms of ERK-induced cell death-apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Wu, L.J.; Zuo, H.J.; Tashiro, S.; Onodera, S.; Ikejima, T. Cytochrome c release from oridonin-treated apoptotic A375-S2 cells is dependent on p53 and extracellular signal-regulated kinase activation. J. Pharmacol. Sci. 2004, 96, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Li, D.W.; Liu, J.P.; Mao, Y.W.; Xiang, H.; Wang, J.; Ma, W.Y.; Dong, Z.; Pike, H.M.; Brown, R.E.; Reed, J.C. Calcium-activated RAF/MEK/ERK signaling pathway mediates p53-dependent apoptosis and is abrogated by alpha B-crystallin through inhibition of RAS activation. Mol. Biol. Cell 2005, 16, 4437–4453. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.J.; Kuo, P.L.; Hsu, Y.C.; Huang, Y.F.; Tsai, E.M.; Hsu, Y.L. Arctigenin, a dietary phytoestrogen, induces apoptosis of estrogen receptor-negative breast cancer cells through the ROS/p38 MAPK pathway and epigenetic regulation. Free Radic. Biol. Med. 2014, 67, 159–170. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, C.-H.; Lee, S.-H.; Lee, S.-K.; Ha, S.-H.; Suh, S.-J.; Kwon, K.-M.; Chung, T.-W.; Ha, K.-T.; Chang, Y.-C.; Lee, Y.-C.; et al. Induction of Apoptosis and Antitumor Activity of Eel Skin Mucus, Containing Lactose-Binding Molecules, on Human Leukemic K562 Cells. Mar. Drugs 2015, 13, 3936-3949. https://doi.org/10.3390/md13063936
Kwak C-H, Lee S-H, Lee S-K, Ha S-H, Suh S-J, Kwon K-M, Chung T-W, Ha K-T, Chang Y-C, Lee Y-C, et al. Induction of Apoptosis and Antitumor Activity of Eel Skin Mucus, Containing Lactose-Binding Molecules, on Human Leukemic K562 Cells. Marine Drugs. 2015; 13(6):3936-3949. https://doi.org/10.3390/md13063936
Chicago/Turabian StyleKwak, Choong-Hwan, Sook-Hyun Lee, Sung-Kyun Lee, Sun-Hyung Ha, Seok-Jong Suh, Kyung-Min Kwon, Tae-Wook Chung, Ki-Tae Ha, Young-Chae Chang, Young-Choon Lee, and et al. 2015. "Induction of Apoptosis and Antitumor Activity of Eel Skin Mucus, Containing Lactose-Binding Molecules, on Human Leukemic K562 Cells" Marine Drugs 13, no. 6: 3936-3949. https://doi.org/10.3390/md13063936
APA StyleKwak, C.-H., Lee, S.-H., Lee, S.-K., Ha, S.-H., Suh, S.-J., Kwon, K.-M., Chung, T.-W., Ha, K.-T., Chang, Y.-C., Lee, Y.-C., Kim, D.-S., Chang, H.-W., & Kim, C.-H. (2015). Induction of Apoptosis and Antitumor Activity of Eel Skin Mucus, Containing Lactose-Binding Molecules, on Human Leukemic K562 Cells. Marine Drugs, 13(6), 3936-3949. https://doi.org/10.3390/md13063936