
Tilapia Piscidin 4 (TP4) Stimulates Cell Proliferation and Wound Closure in MRSA-Infected Wounds in Mice


Abstract
:1. Introduction
2. Results
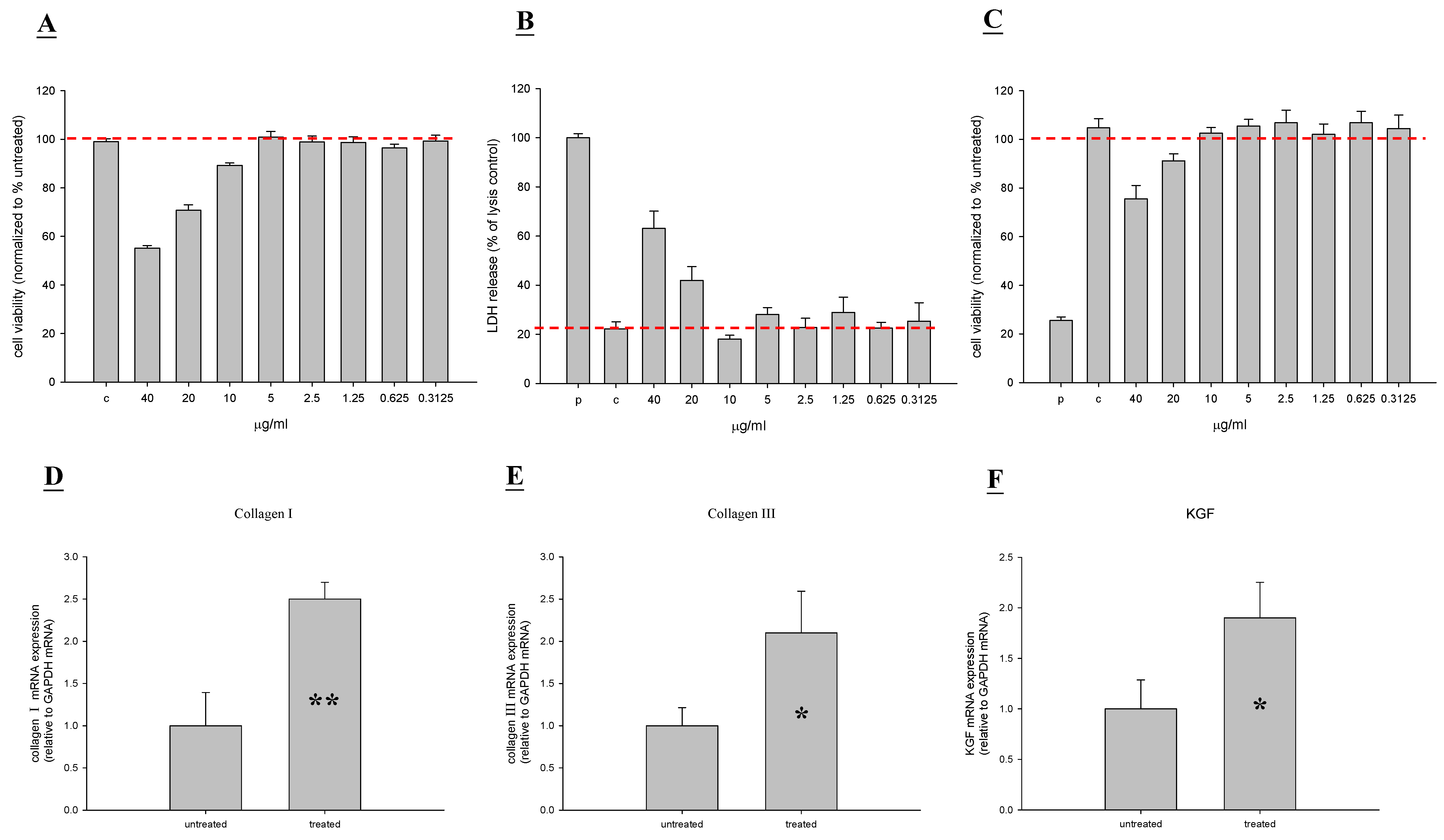
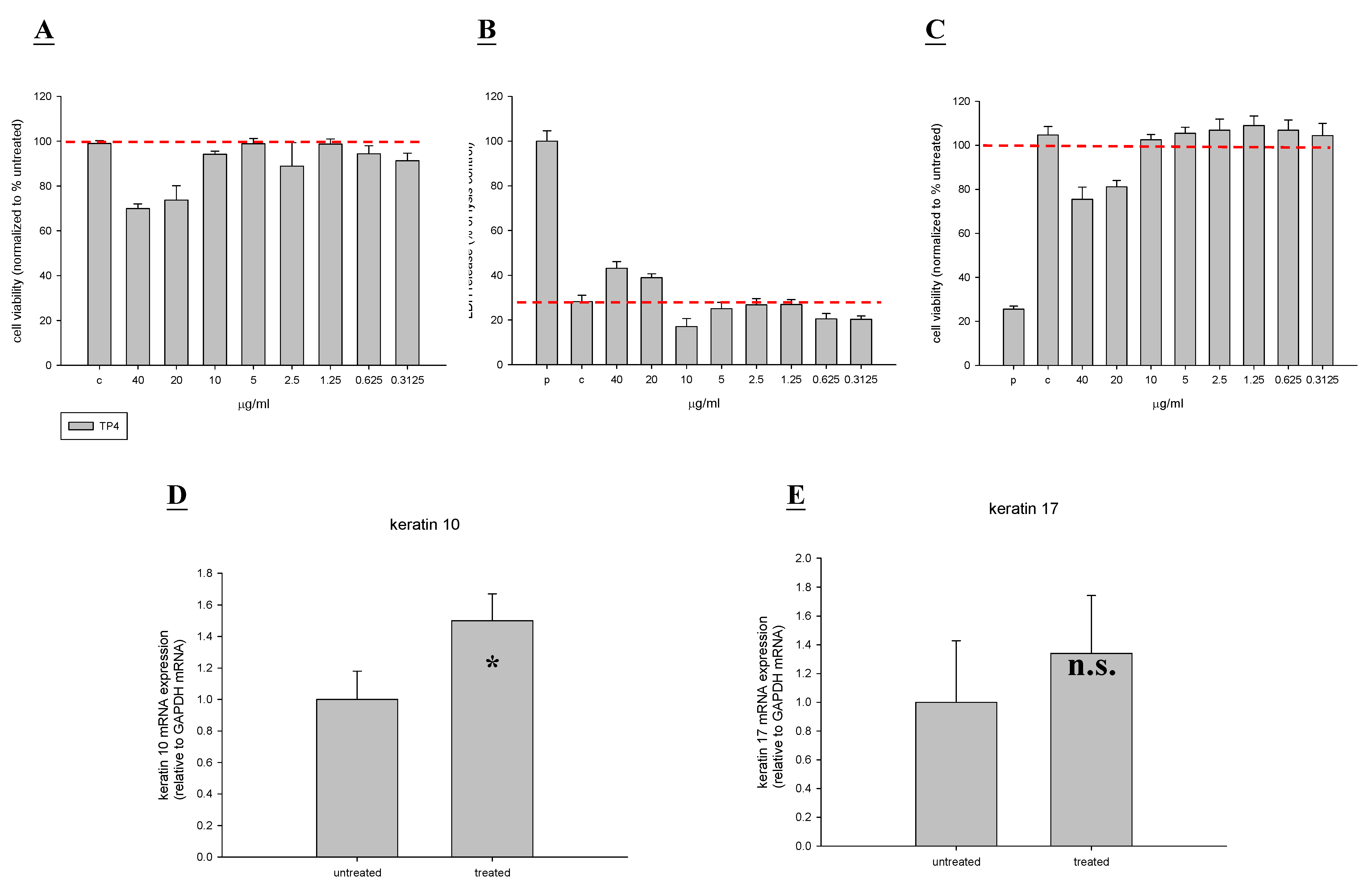
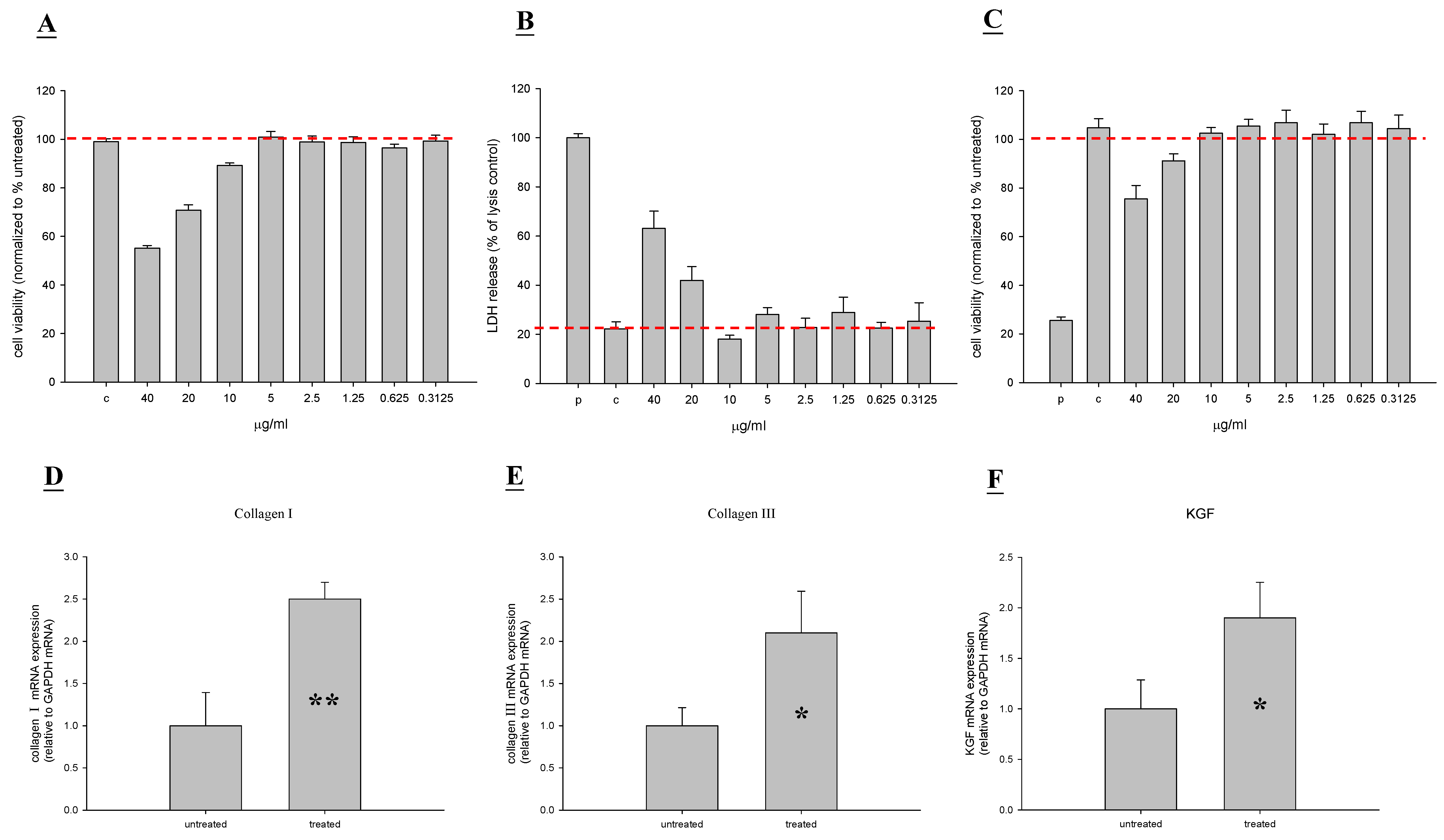
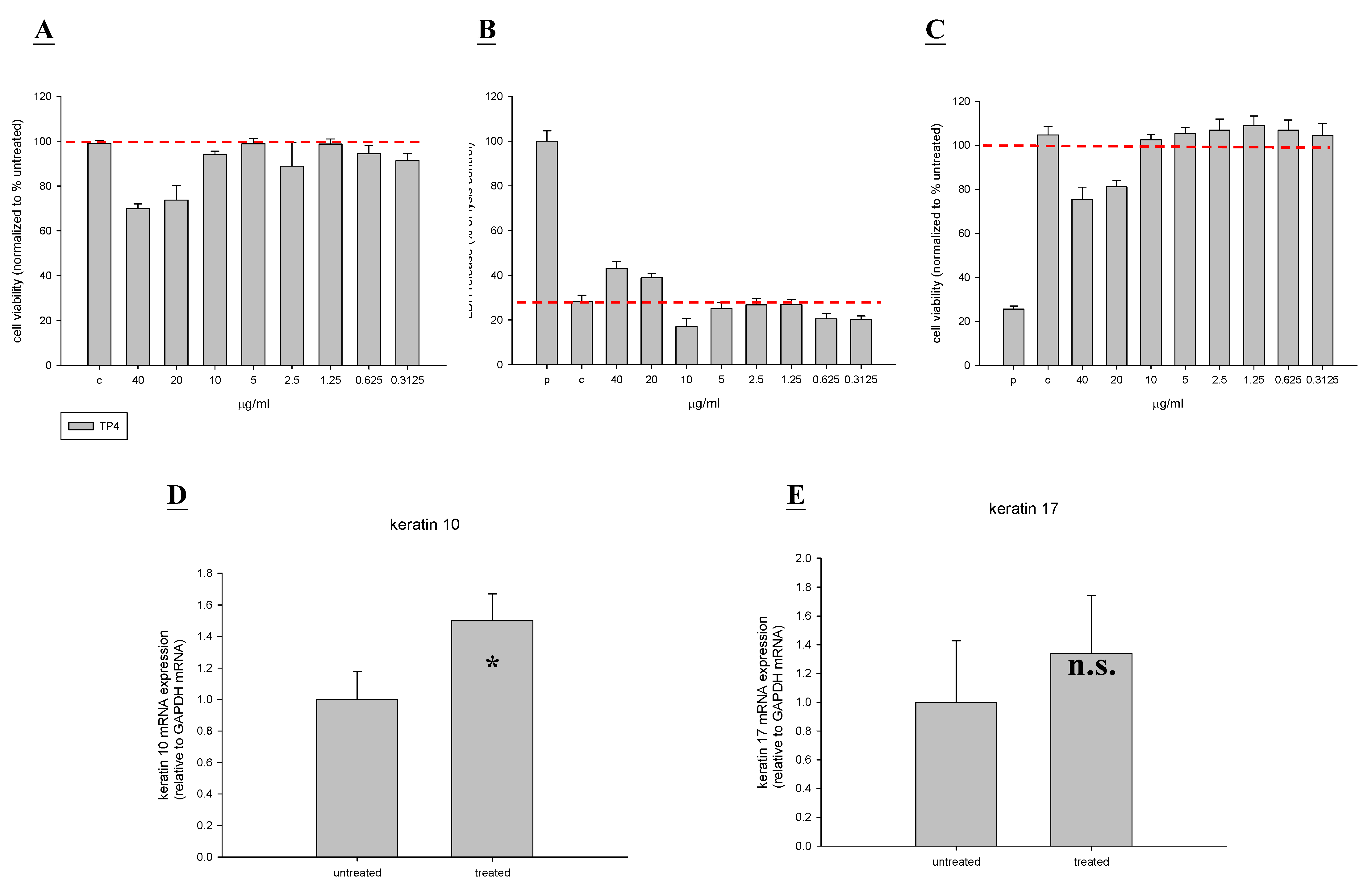
2.1. In Vitro Toxicity and Stimulation of Proliferation by TP4
2.2. Acute Toxic Effects of TP4 in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 10) | TP4 (n = 10) | |||||
|---|---|---|---|---|---|---|
| Time (Day) | 1 | 3 | 6 | 1 | 3 | 6 |
| GOT (U/L) | 42.1 ± 1.3 A | 44.2 ± 2.1 A | 41.6 ± 1.5 A | 102.1 ± 32.1 C | 43.2 ± 1.5 A | 45 ± 6.1 A |
| GPT (U/L) | 45.2 ± 1.1 A | 43.4 ± 4.3 A | 46.1 ± 3.3 A | 81.2 ± 5.2 C | 41.2 ± 4.7 A | 48 ± 3.7 A |
| CRE (mg/dL) | 0.2 ± 0.1 A | 0.6 ± 0.3 A | 0.4 ± 0.2 A | 0.3 ± 0.1 A | 0.5 ± 0.1 A | 0.5 ± 0.1 A |
| BUN (mg/dL) | 13.1 ± 2.1 A | 14 ± 0.3 AB | 16.1 ± 1 A | 14.2 ± 1.4 A | 15.4 ± 1.3 A | 16.2 ± 1 A |
| GLU (mg/dL) | 206.0 ± 11.1 A | 213.1 ± 11.3 A | 208 ± 13.1 A | 238.6 ± 32.1 AB | 209.1 ± 7.2 A | 220.1 ± 23.5 A |
| CPK (U/L) | 101.3 ± 1.7 A | 110.1 ±6.3 A | 104.9 ± 4.2 A | 109 ± 11.1 A | 108.1 ± 3.2 A | 109 ± 25.3 AB |
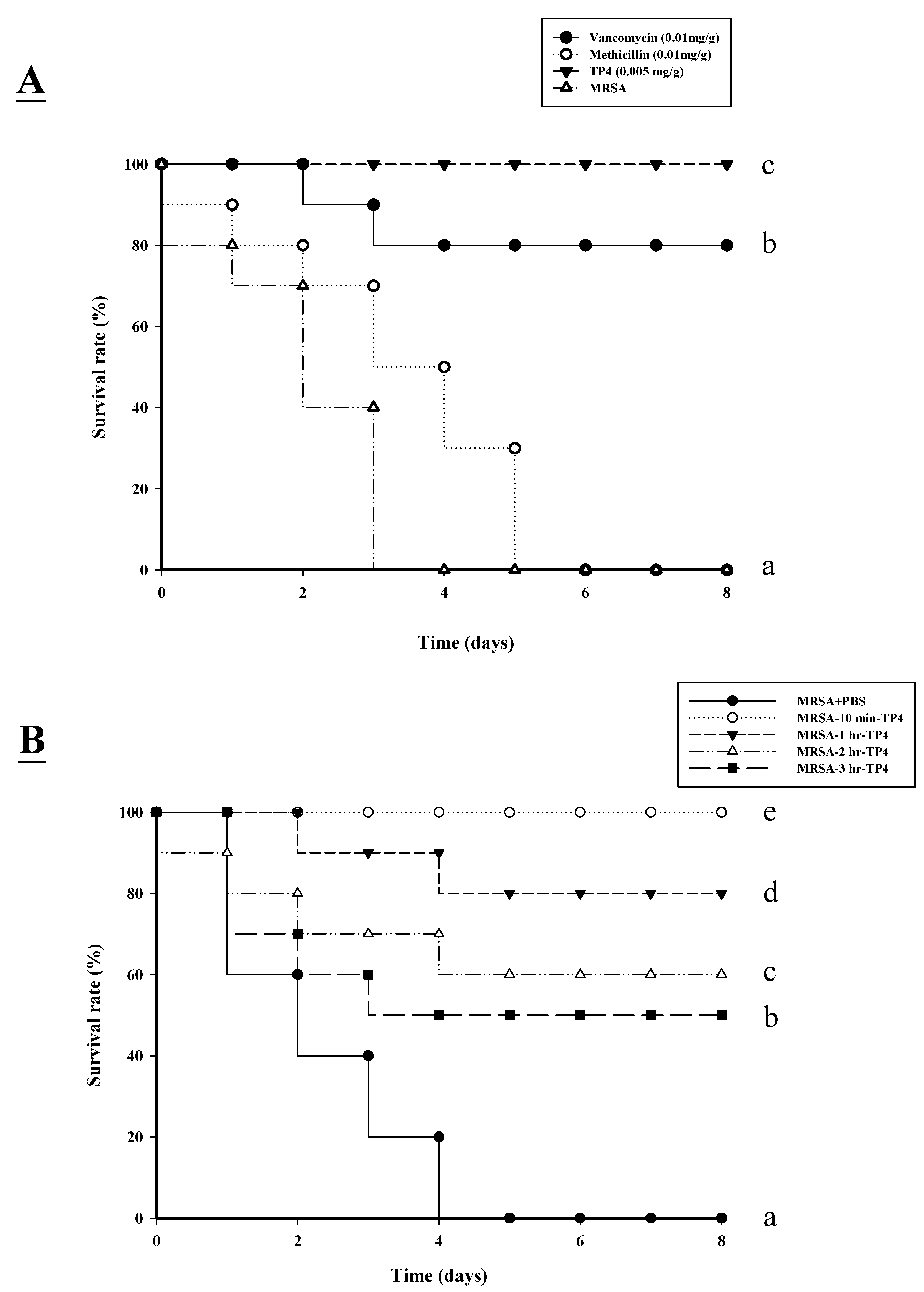
2.3. TP4 Enhances the Survival of Mice Infected with MRSA
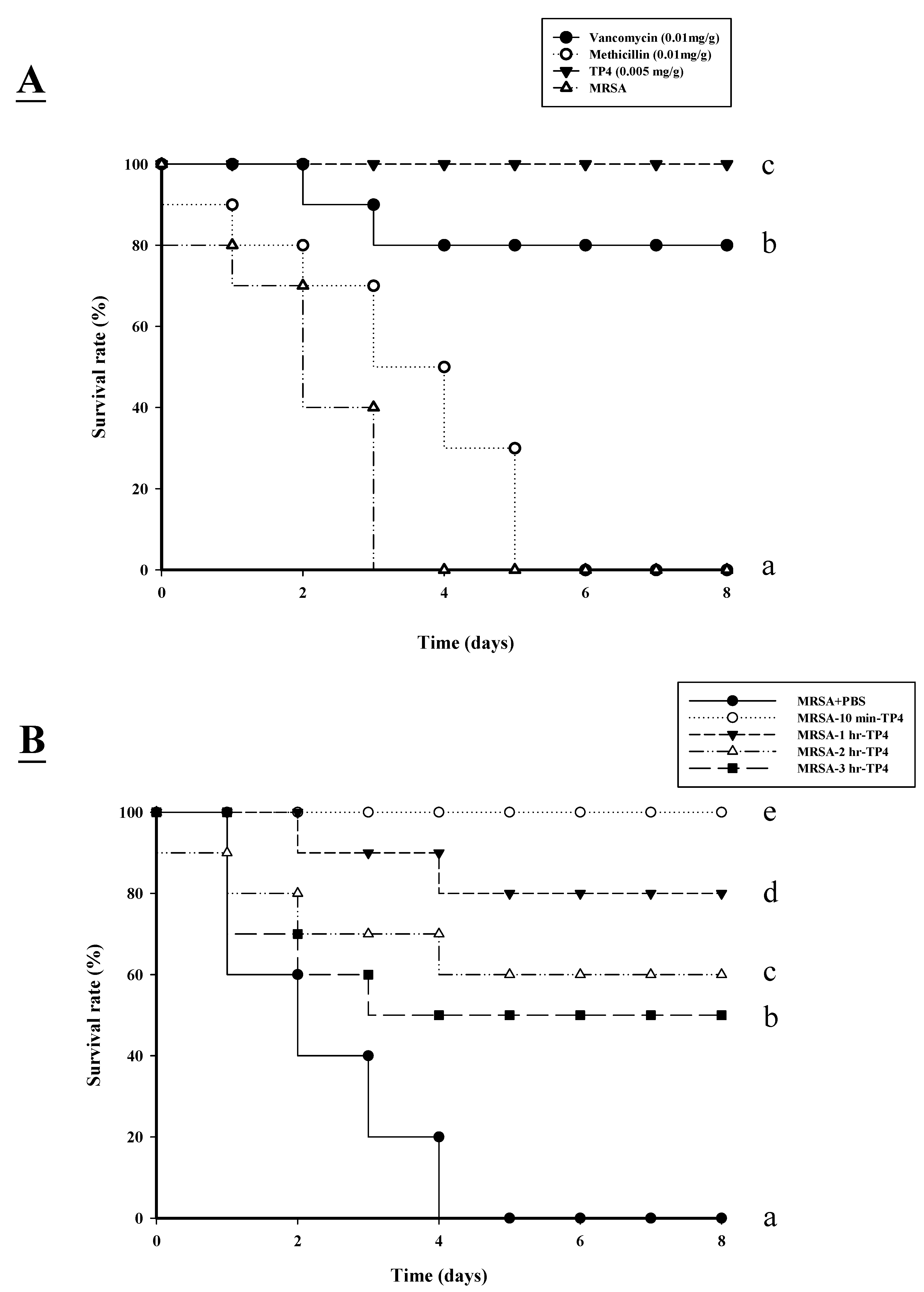
| Strain and Treatment | % lethality | Mean ± SD Count (CFU/mL) | ||||
|---|---|---|---|---|---|---|
| Blood | Peritoneum | Spleen | Liver | Mesenteric Lymph Nodes | ||
| MRSA + PBS | 60 C | 6 × 108 ± 1.9 × 107 C | 2.0 × 1010 ± 1.6 × 109 C | 5.9 × 109 ± 2.0 × 108 C | 2.1 × 108 ± 9.2 × 107 C | 3.2 × 108 ± 1.3 × 107 C |
| MRSA + Methcillin (0.01 mg/g) | 20 B | 4.1 × 107 ± 1.3 × 107 B | 2.3 × 109 ± 2 × 108 B | 1.5 × 108 ± 2.1 × 107 B | 2.7 × 108 ± 3.3 × 106 B | 4.1 × 107 ± 3 × 107 C |
| MRSA + Vancomycin (0.01 mg/g) | 10 AB | 8 × 104 ± 1.1 × 104 B | 1.5 × 106 ± 1 × 105 A | 1.4 × 107 ± 1.1 × 106 B | 1.3 × 107 ± 1.8 × 105 B | 1 × 107 ± 4 × 106 B |
| MRSA + TP4 | 0 A | 0 A | 1 × 106 ± 1 × 106 A | 3 × 106 ± 1.2 × 106 A | 2.4 × 106 ± 1 × 106 A | 6 × 105 ± 1 × 105 A |
| (0.005 mg/g) | ||||||
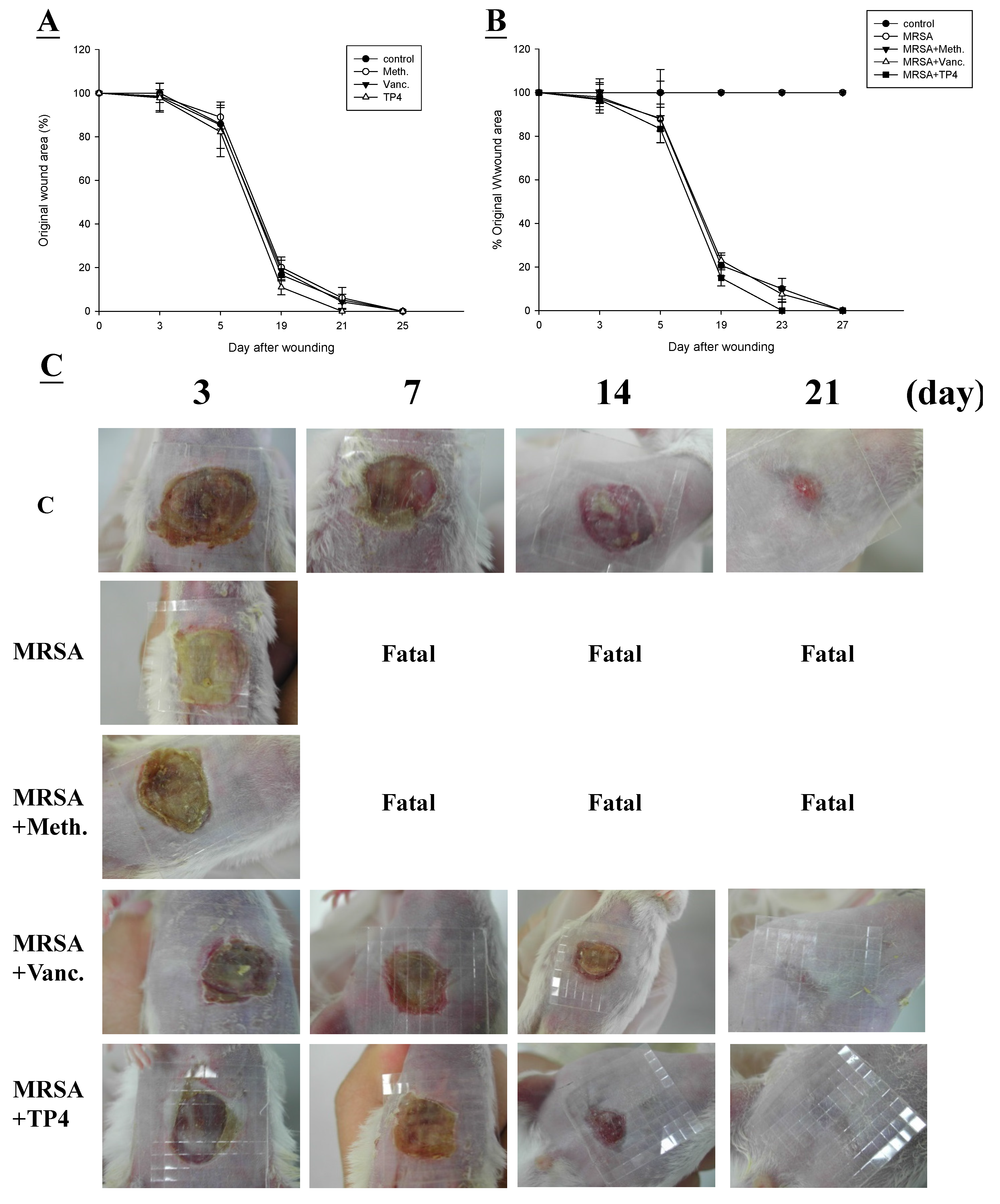
2.4. Efficacy of TP4 on in Vivo Wound Closure
2.5. TP4 Reduced Inflammatory Cytokines
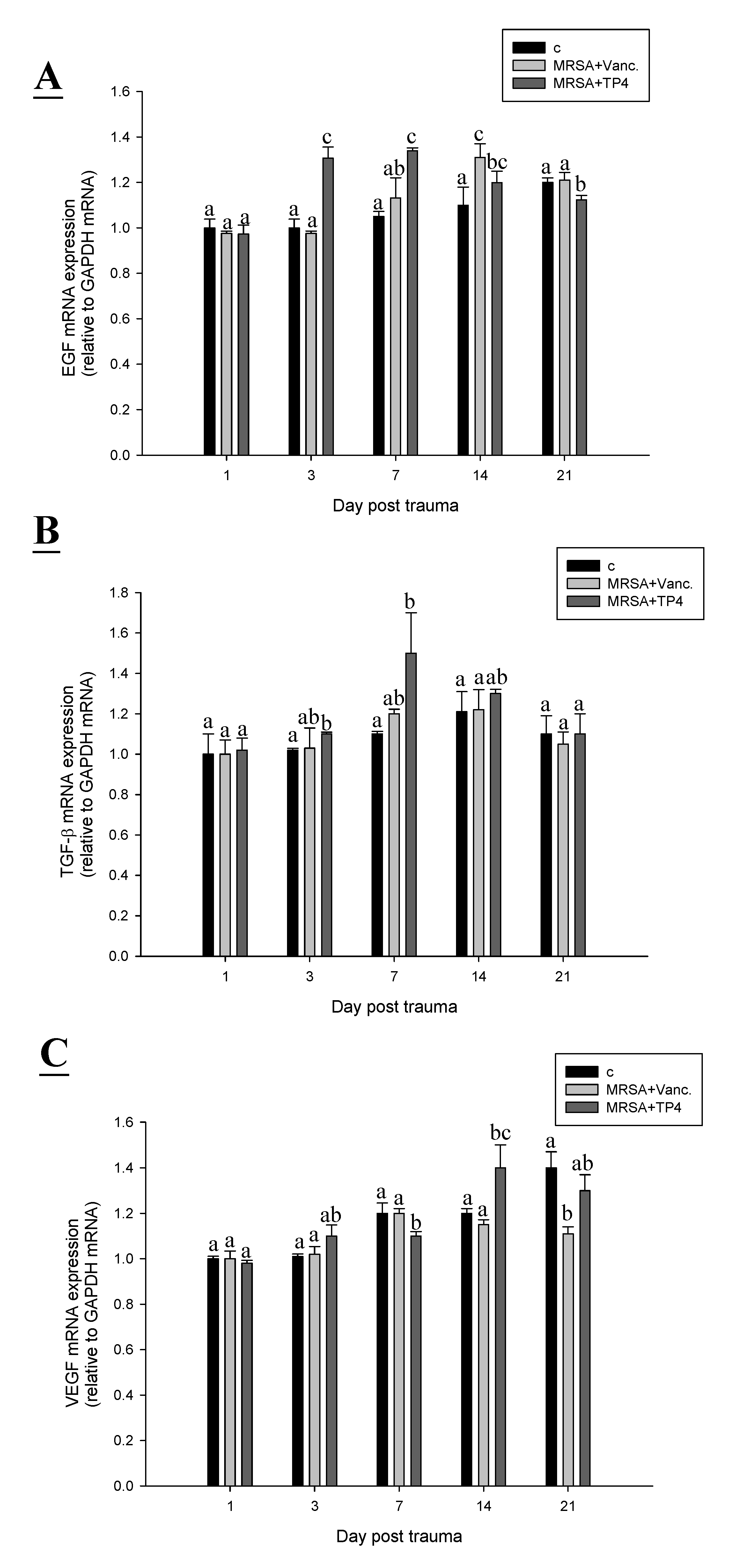
2.6. TP4 Alters Cell Proliferation Gene Expression Profiles in MRSA-Infected Mice
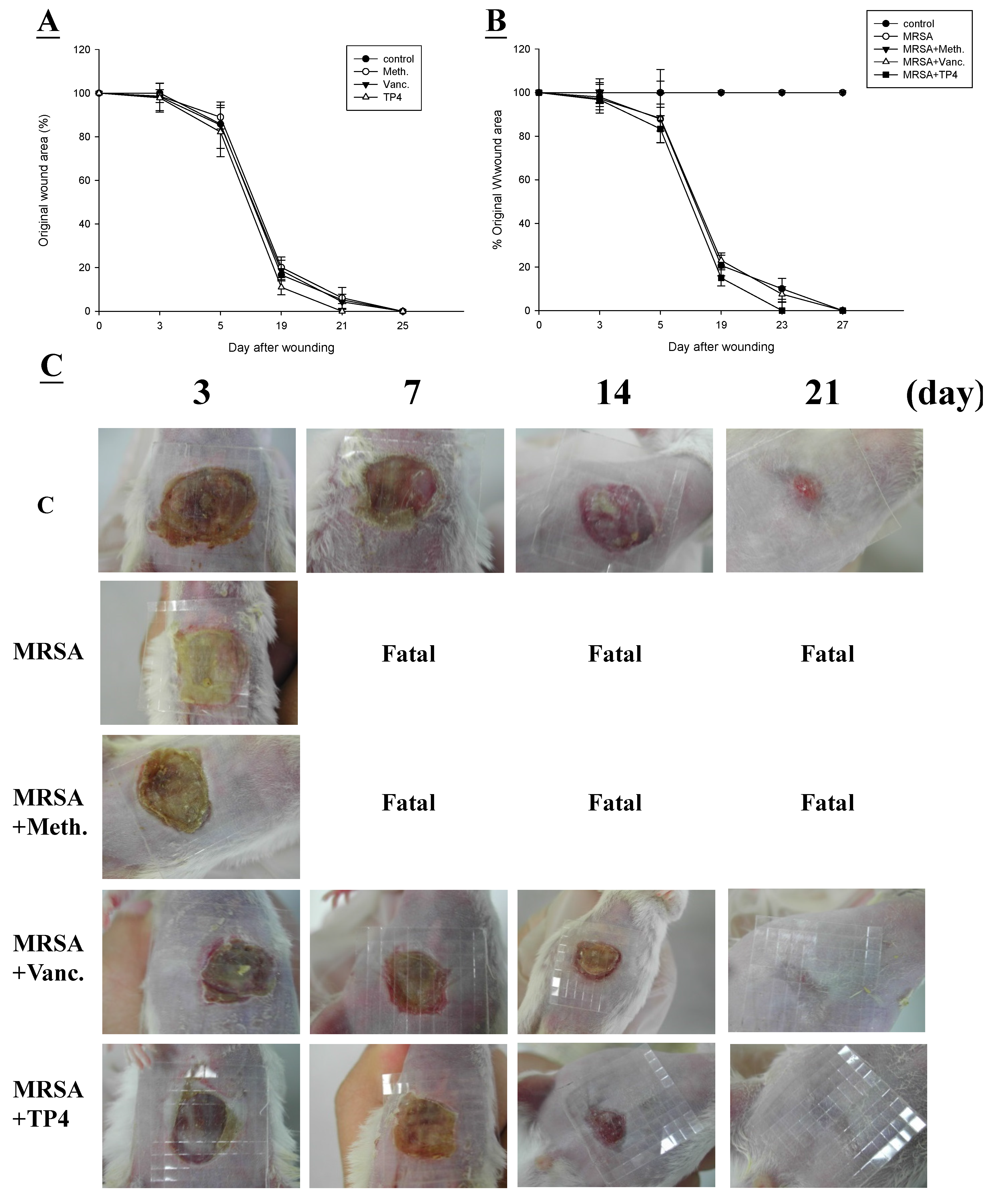
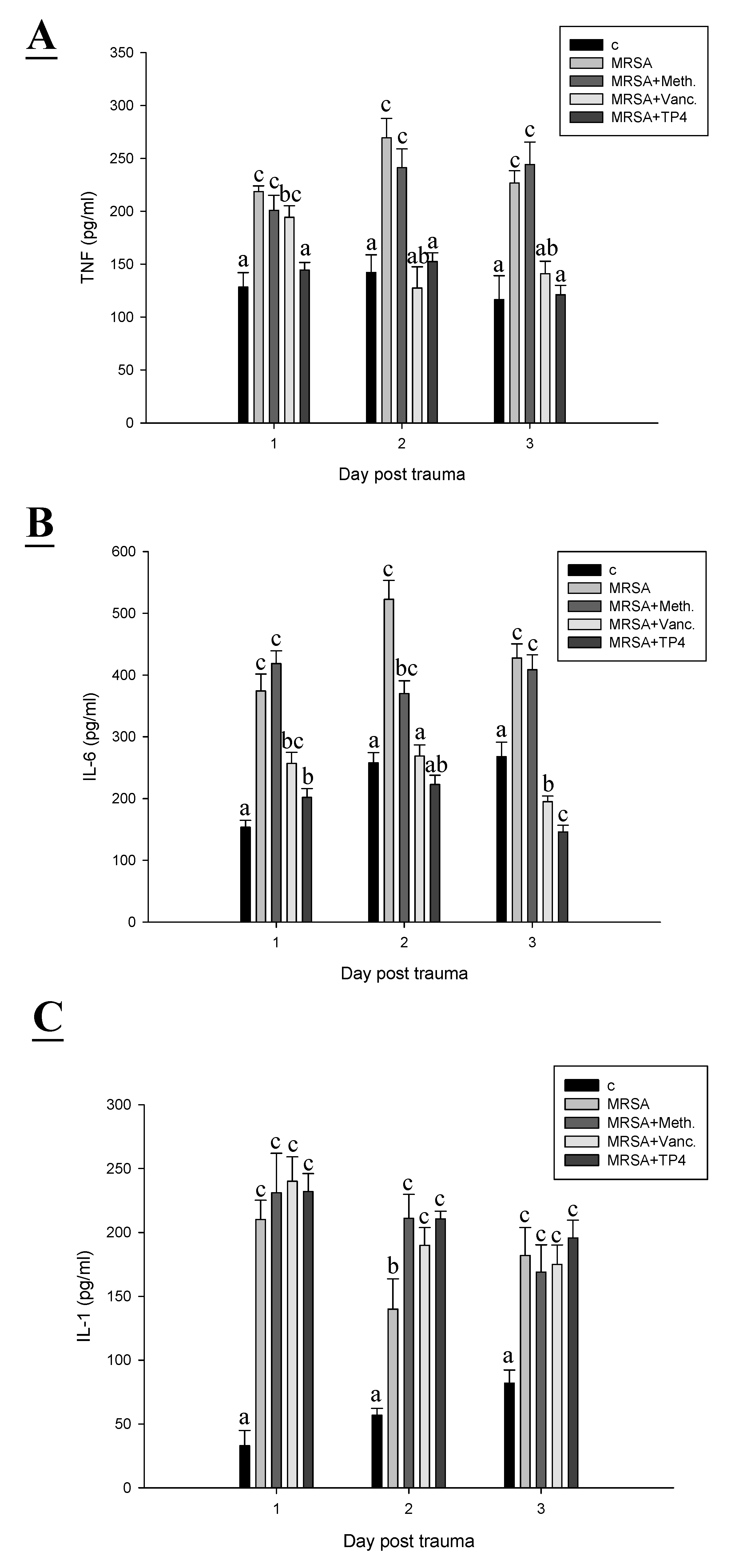
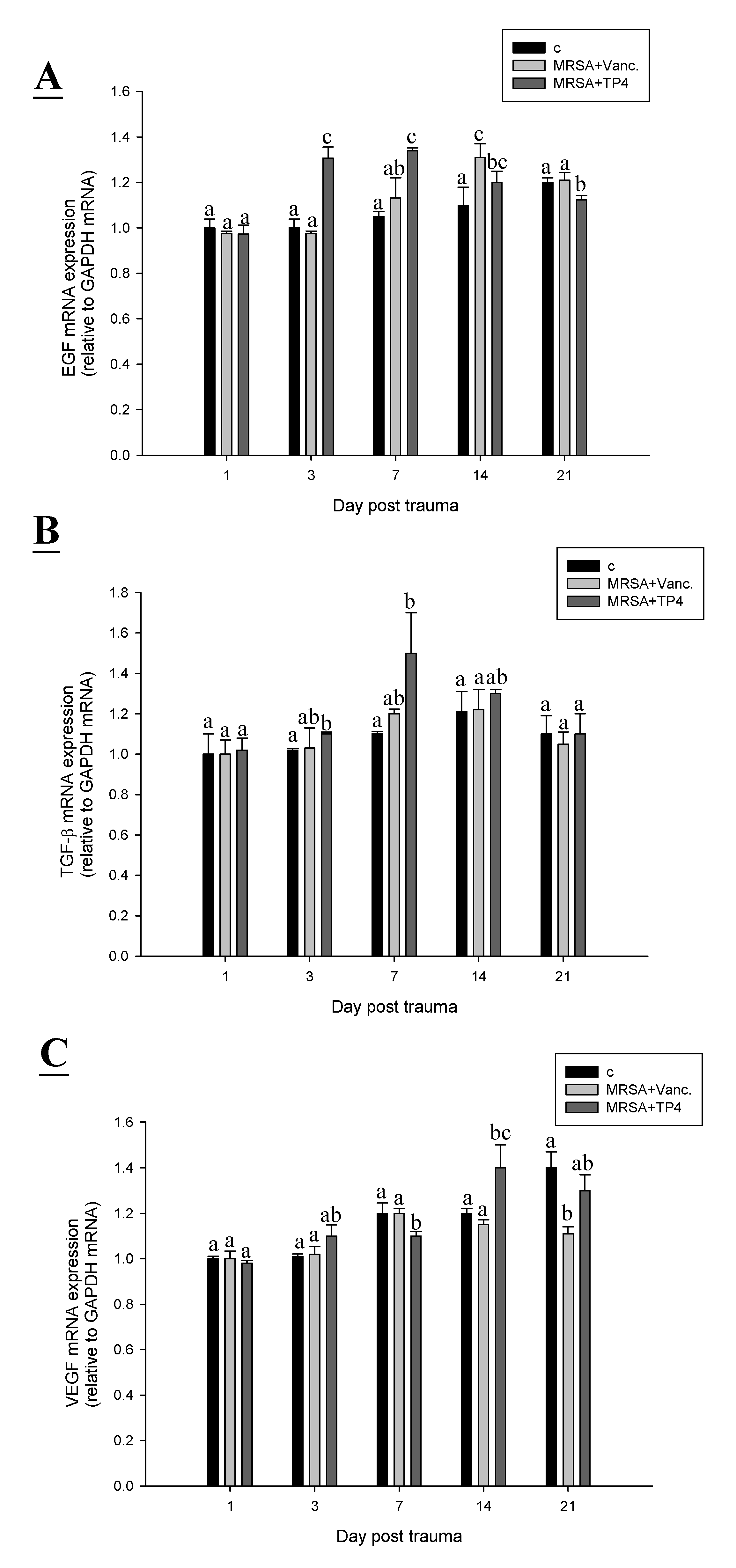

3. Discussion
4. Experimental Section
4.1. Bacteria, Cells, and Mice
4.2. Peptides, Reagents, and Antibodies
4.3. Cell Proliferation
4.4. Measurement of Collagen I, Collagen III, KGF, Keratin 10, and Keratin 17 Expression
| GAPDH | Sense | CGCTCTCTGCTCCTCCTGTTC |
| (Nm_002046) | Antisense | TTGACTCCGACCTTCACCTTCC |
| Collagen I | Sense | ACAGGGCGACAGAGGCATAAAG |
| (NM_000088) | Antisense | CCAGGAGCACCAGCAGAGC |
| Collagen III | Sense | TCCAAAGGGTGACAAGGGTGAAC |
| (NM_000090) | Antisense | AGGAGGACCAATAGGACCAGTAGG |
| KGF | Sense | GCAACTGAACTTACTACGAACTG |
| (S81661) | Antisense | TCATTGACCTCTTCCTATCTGTG |
| Keratin 10 | Sense | CTGCGTAGGGTGCTGGATGAG |
| (AF264085) | Antisense | TTCCTCCTCGTGGTTCTTCTTCAG |
| Keratin 17 | Sense | CTGGCTGCTGATGACTTCC |
| (NM_000422) | Antisense | CCTCCTCGTGGTTCTTCTTC |
4.5. Tilapia Piscidin 4 Peptides and Bacteriostatic Analysis
4.6. In Vivo Toxicity
4.7. Therapeutic Use in a Mouse Model of MRSA Sepsis
4.8. Mouse Models of Wound Healing
4.9. Wound Closure Measurements and Cell Proliferation Gene Expression
| GAPDH | Sense | CTCCAAGGAGTAAGAAACCC |
| (GU214026.1) | Antisense | TGGAAATTGTGAGGGAGATG |
| EGF | Sense | CATATGTGATGGCTACTGCT |
| (AF125256.1) | Antisense | TTAATGTTCCTCAGGGAAGC |
| TGF-β | Sense | CGTGCTCTTCTTCGACAATA |
| (M57639) | Antisense | AACATGAACAAACAGTCCCT |
| VEGF | Sense | ACCTTTGGGAAGAAGATGTC |
| (AB086118.1) | Antisense | CAATAGAACCCTCGAGTGAG |
4.10. IHC and ELISA of Cytokines
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Theuretzbacher, U.; Toney, J.H. Nature’s clarion call of antibacterial resistance: Are we listening? Curr. Opin. Investig. Drugs 2006, 7, 158–166. [Google Scholar] [PubMed]
- Spellberg, B.; Powers, J.H.; Brass, E.P.; Miller, L.G.; Edwards, J.E., Jr. Trends in antimicrobial drug development: Implications for the future. Clin. Infect. Dis. 2004, 38, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.N.; Pan, C.Y.; Rajanbabu, V.; Chan, Y.L.; Wu, C.J.; Chen, J.Y. Modulation of immune responses by the antimicrobial peptide, epinecidinn(Epi)-1, and establishment of an Epi-1-based inactivated vaccine. Biomaterials 2011, 32, 3627–3636. [Google Scholar]
- Peng, K.C.; Lee, S.H.; Hour, A.L.; Pan, C.Y.; Lee, L.H.; Chen, J.Y. Five different piscidins from Nile tilapia, Oreochromis niloticus: Analysis of their expressions and biological functions. PLoS ONE 2012, 7, e50263. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.; Carpio, Y.; Valdés, I.; Velázquez, J.; Zamora, Y.; Morales, R.; Morales, A.; Rodríguez, E.; Estrada, M.P. Co-administration of tilapia alpha-helical antimicrobial peptides with subunit antigens boost immunogenicity in mice and tilapia (Oreochromis niloticus). Vaccine 2014, 32, 223–229. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, S.; Cullum, N.; Majid, M.; Sheldon, T. Systematic reviews of wound care management: (3) antimicrobial agents for chronic wounds; (4) diabetic foot ulceration. Health Technol. Assess. 2000, 4, 1–237. [Google Scholar] [PubMed]
- Pan, C.Y.; Wu, J.L.; Hui, C.F.; Lin, C.H.; Chen, J.Y. Insights into the antibacterial and immunomodulatory functions of the antimicrobial peptide, epinecidin-1, against Vibrio vulnificus infection in zebrafish. Fish Shellf. Immunol. 2011, 31, 1019–1025. [Google Scholar] [CrossRef]
- Huang, T.C.; Chen, J.Y. Proteomic and functional analysis of zebrafish after administration of antimicrobial peptide epinecidin-1. Fish Shellf. Immunol. 2013, 34, 593–598. [Google Scholar] [CrossRef]
- Singer, A.J.; Clark, R.A.F. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Michelson, P.H.; Tigue, M.; Panos, R.J.; Sporn, P.H. Keratinocyte growth factor stimulates bronchial epithelial cell proliferation in vitro and in vivo. Am. J. Physiol. Lung Cell Mol. Physiol. 1999, 277, L737–L742. [Google Scholar]
- Pawankar, R. Epithelial cells as immunoregulators in allergic airway diseases. Curr. Opin. Allergy Clin. Immunol. 2001, 2, 1–5. [Google Scholar] [CrossRef]
- Puddicombe, S.M.; Polosa, R.; Richter, A.; Krishna, M.T.; Howarth, P.H.; Holgate, S.T.; Davies, D.E. Involvement of the epidermal growth factor receptor in epithelial repair in asthma. FASEB J. 2000, 14, 1362–1374. [Google Scholar] [CrossRef] [PubMed]
- Aarbiou, J.; Verhoosel, R.M.; van Wetering, S.; de Boer, W.I.; van Krieken, J.H.; Litvinov, S.V.; Rabe, K.F.; Hiemstra, P.S. Human neutrophil defensins induce lung epithelial cell proliferation in vitro. J. Leukoc. Biol. 2002, 72, 167–174. [Google Scholar] [PubMed]
- Aarbiou, J.; Verhoosel, R.M.; van Wetering, S.; de Boer, W.I.; van Krieken, J.H.; Litvinov, S.V.; Rabe, K.F.; Hiemstra, P.S. Neutrophil defensins enhance lung epithelial wound closure and mucin gene expression in vitro. Am. J. Respir. Cell Mol. Biol. 2004, 30, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Houghton, P.J.; Hylands, P.J.; Mensah, A.Y.; Hensel, A.; Deters, A.M. In vitro tests and ethnopharmacological investigations: Wound healing as an example. J. Ethnopharmacol. 2005, 100, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Martin, A. The use of antioxidants in healing. Dermatol. Surg. 1996, 22, 156–160. [Google Scholar]
- Ruszczak, Z. Effect of collagen matrices on dermal wound healing. Adv. Drug Deliv. Rev. 2003, 55, 1595–1611. [Google Scholar] [CrossRef] [PubMed]
- McGowan, K.M.; Tong, X.; Colucci-Guyon, E.; Langa, F.; Babinet, C.; Coulombe, P.A. Keratin 17 null mice exhibit age and strain-dependent alopecia. Genes Dev. 2002, 16, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Sauder, D.N.; Kilian, P.L.; McLane, J.A.; Quick, T.W.; Jakubovic, H.; Davis, S.C.; Eaglstein, W.H.; Mertz, P.M. Interleukin-1 enhances epidermal wound healing. Lymphokine Res. 1990, 9, 465–473. [Google Scholar] [PubMed]
- Galko, M.J.; Krasnow, M.A. Cellular and genetic analysis of wound healing in Drosophila larvae. PLoS Biol. 2004, 2, E239. [Google Scholar] [CrossRef] [PubMed]
- Beanes, S.R.; Dang, C.; Soo, C.; Ting, K. Skin repair and scar formation: The central role of TGF-beta. Expert Rev. Mol. Med. 2003, 5, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S. Review of epidermal growth factor receptor biology. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Broughton, G., 2nd; Janis, J.E.; Attinger, C.E. Wound healing: An overview. Plast. Reconstr. Surg. 2006, 117 (Suppl. 7), 1e-S–32e-S. [Google Scholar]
- Fox, J.L. Antimicrobial peptides stage a comeback. Nat. Biotechnol. 2013, 31, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Biragyn, A.; Hoover, D.M.; Lubkowski, J.; Oppenheim, J.J. Multiple roles of antimicrobial defensins, cathelicidins, and eosinophil-derived neurotoxin in host defense. Annu. Rev. Immunol. 2004, 22, 181–215. [Google Scholar] [CrossRef] [PubMed]
- Hebda, P.A.; Whaley, D.; Kim, H.G.; Wells, A. Absence of inhibition of cutaneous wound healing in mice by oral doxycycline. Wound Repair Regen. 2003, 11, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Babu, M.; Wells, A. Dermal-epidermal communication in wound healing. Wounds 2001, 13, 183–189. [Google Scholar]
- Hsu, J.C.; Lin, L.C.; Tzen, J.T.; Chen, J.Y. Pardaxin-induced apoptosis enhances antitumor activity in HeLa cells. Peptides 2011, 32, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Kirker, K.R.; Luo, Y.; Nielson, J.H.; Shelby, J.; Prestwich, G.D. Glycosaminoglycan hydrogel films as bio-interactive dressings for wound healing. Biomaterials 2002, 23, 3661–3671. [Google Scholar] [CrossRef] [PubMed]
- Bird, T.A.; Saklatvala, J. IL-1 and TNF transmodulate epidermal growth factor receptors by a protein kinase C-independent mechanism. J. Immunol. 1989, 142, 126–133. [Google Scholar] [PubMed]
- Huang, H.N.; Rajanbabu, V.; Pan, C.Y.; Chan, Y.L.; Wu, C.J.; Chen, J.Y. Use of the antimicrobial peptide Epinecidin-1 to protect against MRSA infection in mice with skin injuries. Biomaterials 2013, 34, 10319–10327. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Dai, C.; Li, Z.; Fan, Z.; Song, Y.; Wu, Y.; Cao, Z.; Li, W. Antibacterial activity and mechanism of a scorpion venom peptide derivative in vitro and in vivo. PLoS ONE 2012, 7, e40135. [Google Scholar] [CrossRef] [PubMed]
- Yates, C.C.; Whaley, D.; Babu, R.; Zhang, J.; Krishna, P.; Beckman, E.; Pasculle, A.W.; Wells, A. The effect of multifunctional polymer-based gels on wound healing in full thickness bacteria-contaminated mouse skin wound models. Biomaterials 2007, 28, 3977–3986. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.-N.; Chan, Y.-L.; Wu, C.-J.; Chen, J.-Y. Tilapia Piscidin 4 (TP4) Stimulates Cell Proliferation and Wound Closure in MRSA-Infected Wounds in Mice. Mar. Drugs 2015, 13, 2813-2833. https://doi.org/10.3390/md13052813
Huang H-N, Chan Y-L, Wu C-J, Chen J-Y. Tilapia Piscidin 4 (TP4) Stimulates Cell Proliferation and Wound Closure in MRSA-Infected Wounds in Mice. Marine Drugs. 2015; 13(5):2813-2833. https://doi.org/10.3390/md13052813
Chicago/Turabian StyleHuang, Hang-Ning, Yi-Lin Chan, Chang-Jer Wu, and Jyh-Yih Chen. 2015. "Tilapia Piscidin 4 (TP4) Stimulates Cell Proliferation and Wound Closure in MRSA-Infected Wounds in Mice" Marine Drugs 13, no. 5: 2813-2833. https://doi.org/10.3390/md13052813
APA StyleHuang, H.-N., Chan, Y.-L., Wu, C.-J., & Chen, J.-Y. (2015). Tilapia Piscidin 4 (TP4) Stimulates Cell Proliferation and Wound Closure in MRSA-Infected Wounds in Mice. Marine Drugs, 13(5), 2813-2833. https://doi.org/10.3390/md13052813