
Selective Blocking Effects of 4,9-Anhydrotetrodotoxin, Purified from a Crude Mixture of Tetrodotoxin Analogues, on NaV1.6 Channels and Its Chemical Aspects

Abstract
:1. Introduction
2. NaV Channels
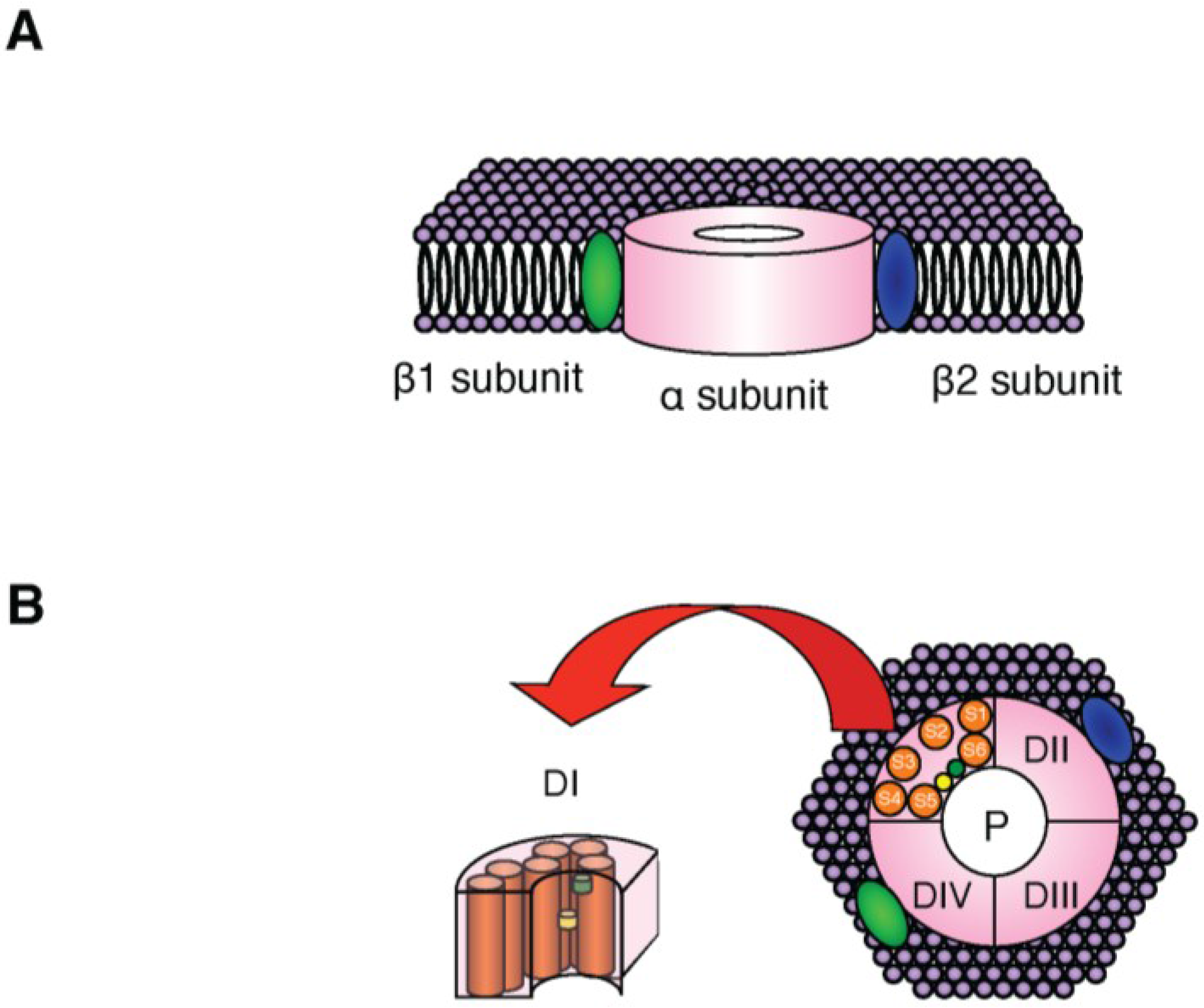

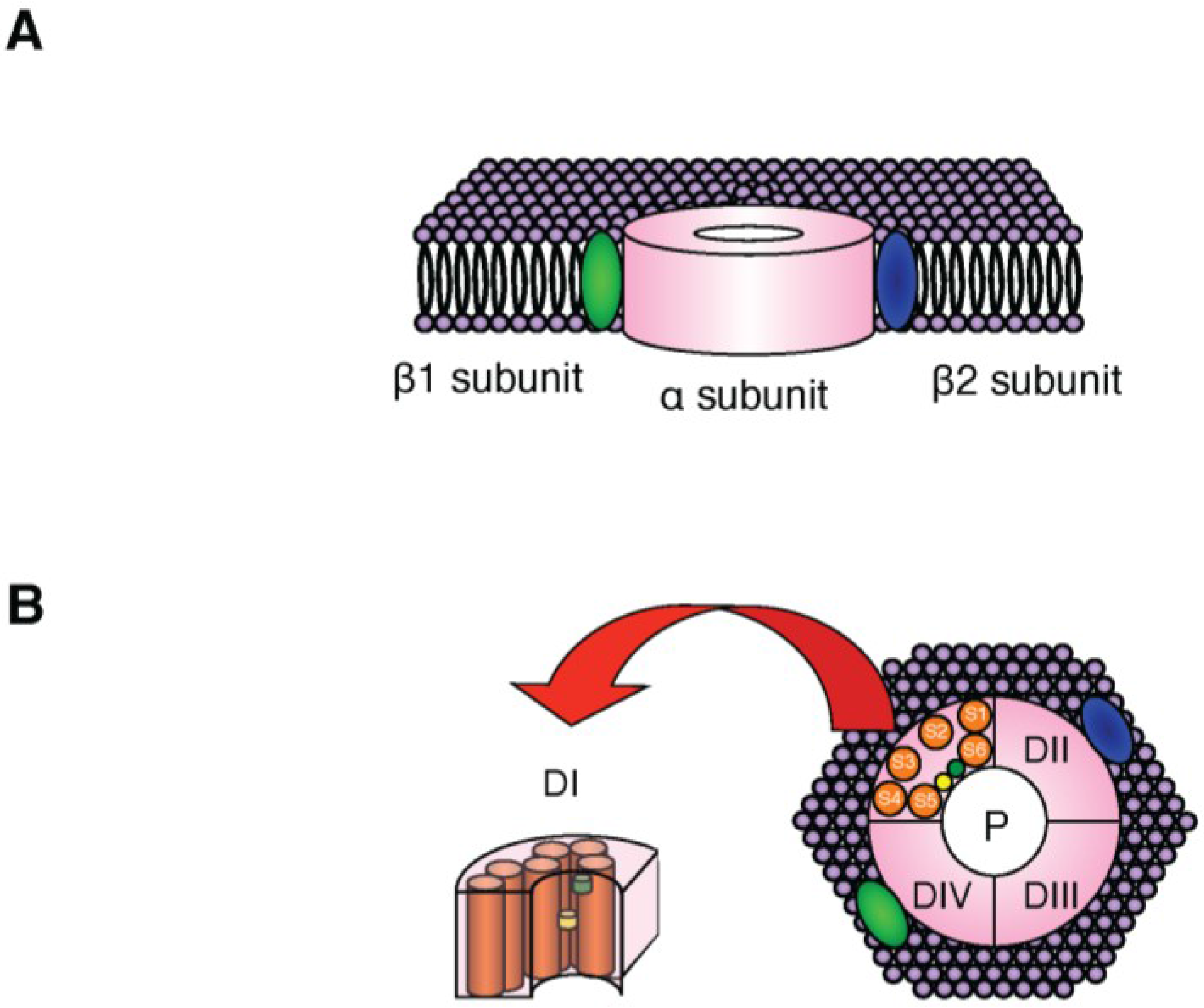{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α Subunit (Channel pore) | Coding gene | TTX-Sensitivity | Predominant location |
|---|---|---|---|
| NaV1.1 | Scn1A | S | CNS, PNS |
| NaV1.2 | Scn2A | S | CNS |
| NaV1.3 | Scn3A | S | CNS (embryonic) |
| NaV1.4 | Scn4A | S | Skeletal muscle |
| NaV1.5 | Scn5A | R | Heart muscle |
| NaV1.6 | Scn8A | S | CNS, PNS, Glia, Nodes of Ranvier, SMCs |
| NaV1.7 | Scn9A | S | PNS, Schwann cells |
| NaV1.8 | Scn10A | R | PNS (sensory neurons) |
| NaV1.9 | Scn11A | R | PNS |
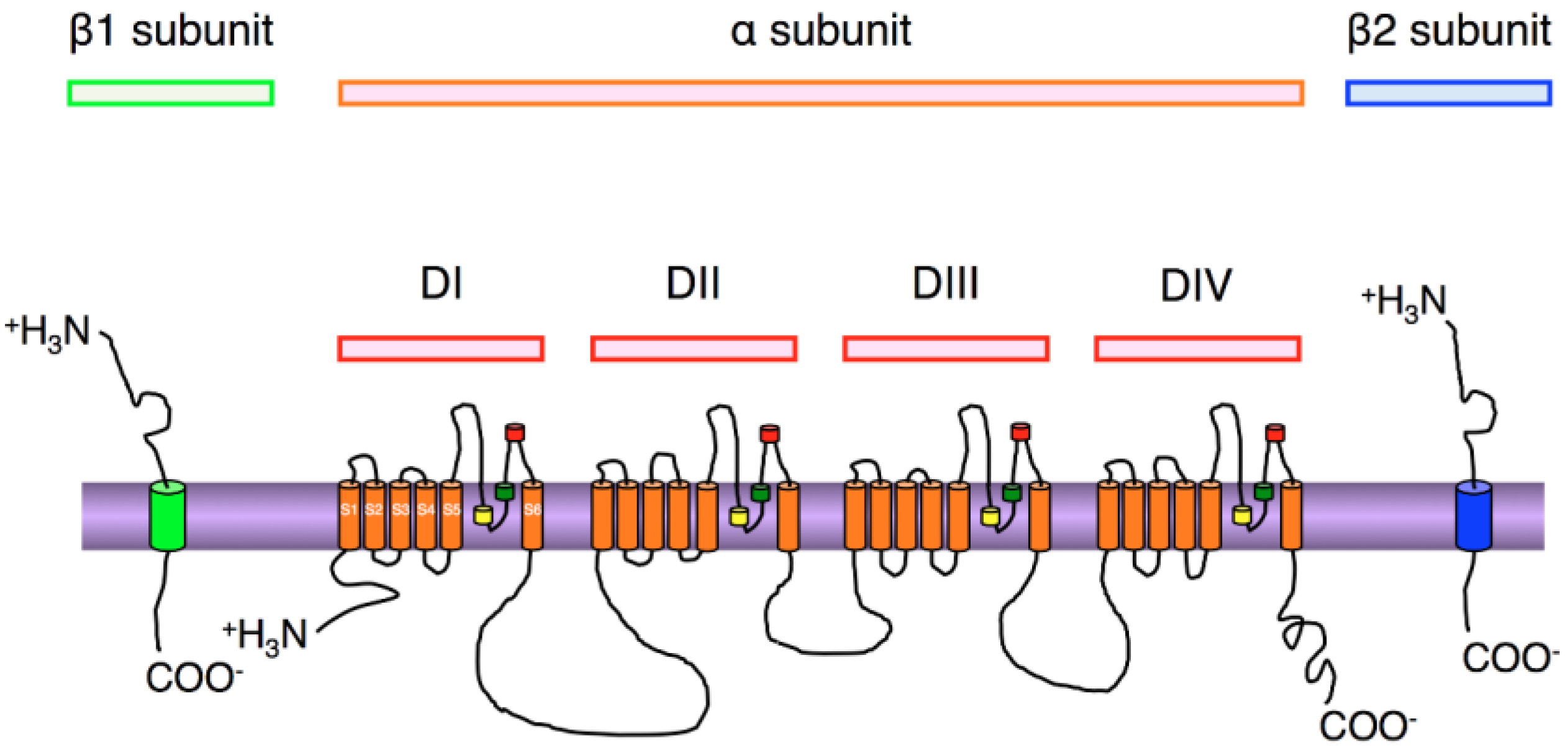
3. Localization of NaV1.6 Channels
4. NaV1.6 Channel-Null Mice (med Mice) Lacking Expression of Scn8a
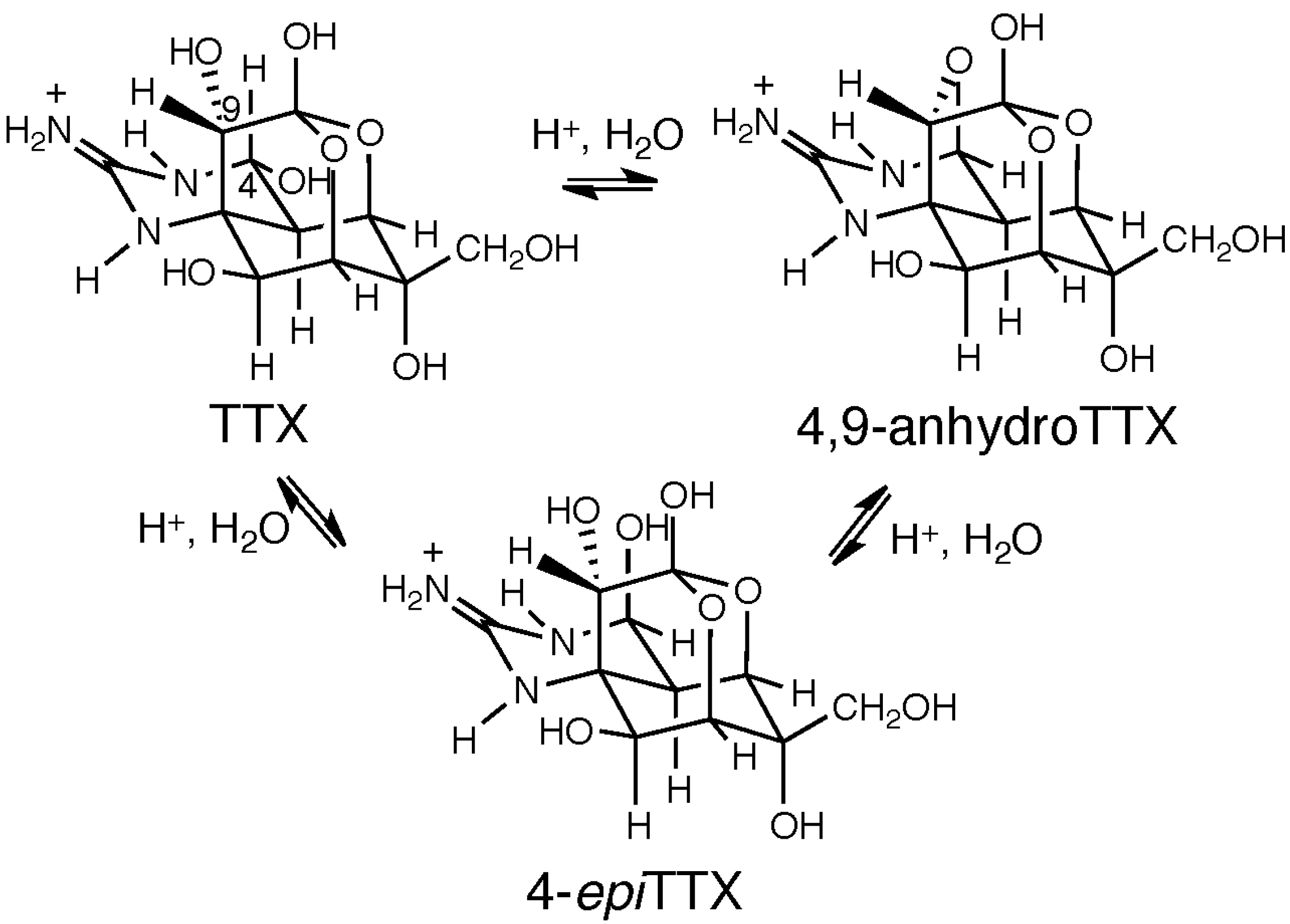
5. Chemical and Toxicological Aspects of 4,9-AnhydroTTX
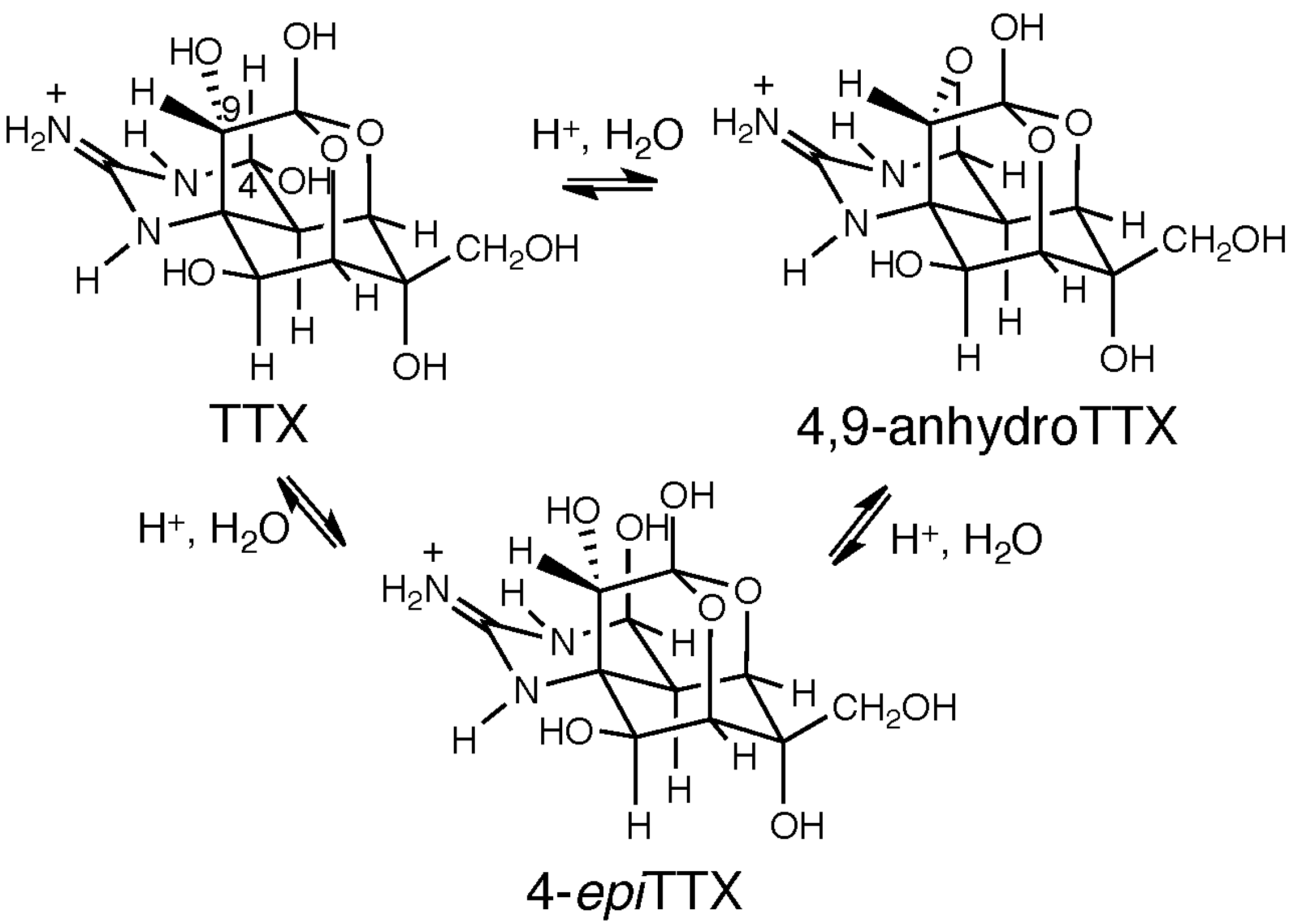
6. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tahara, Y. Studies on globefish poison. J. Pharm. Soc. Jpn. 1909, 29, 587–625. [Google Scholar]
- Narahashi, T. Tetrodotoxin: A brief history. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2008, 84, 147–154. [Google Scholar] [PubMed]
- Hille, B. The superfamily of voltage-gated channels. In Ionic Channels of Excitable Membranes, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001; pp. 61–94. [Google Scholar]
- Nakamura, M.; Yasumoto, T. Tetrodotoxin derivatives in puffer fish. Toxicon 1985, 23, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Yasumoto, T. Actions of 4-epitetrodotoxin and anhydrotetrodotoxin on the squid axon. Toxicon 1985, 23, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Sugimoto, A.; Takai, A.; Yasumoto, T. Effects of specific modifications of several hydroxyls of tetrodotoxin on its affinity to rat brain membrane. J. Pharmacol. Exp. Ther. 1999, 289, 1688–1696. [Google Scholar] [PubMed]
- Yasumoto, T.; Michishita, T. Fluorometric determination of tetrodotoxin by high performance liquid chromatography. Agric. Biol. Chem. 1985, 49, 3077–3080. [Google Scholar] [CrossRef]
- Shoji, Y.; Yotsu-Yamashita, M.; Miyazawa, T.; Yasumoto, T. Electrospray ionization mass spectrometry of tetrodotoxin and its analogs: Liquid mass spectrometry, and liquid chromatography/tandem mass spectrometry. Anal. Biochem. 2001, 290, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Yotsu-Yamashita, M. Distribution of tetrodotoxin, saxitoxin, and their analogs among tissues of the puffer fish Fugu pardalis. Toxicon 2006, 48, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Jang, J.; Yotsu-Yamashita, M. Hydrophilic interaction liquid chromatography-electrospray ionization mass spectrometry of tetrodotoxin and its analogs. Anal. Biochem. 2006, 352, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Jang, J.H.; Cho, Y.; Konoki, K. Optimization of simultaneous analysis of tetrodotoxin, 4-epitetrodotoxin, 4,9-anhydrotetrodotoxin, and 5,6,11-trideoxytetrodotoxin by hydrophilic interaction liquid chromatography-tandem mass spectrometry. Forensic Toxicol. 2011, 29, 61–64. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.J.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T.; et al. First identification of 5,11-dideoxytetrodotoxin in marine animals, and characterization of major fragment ions of tetrodotoxin and its analogs by high resolution ESI-MS/MS. Mar. Drugs 2013, 11, 2799–2813. [Google Scholar] [CrossRef] [PubMed]
- Rosker, C.; Lohberger, B.; Hofer, D.; Steinecker, B.; Quasthoff, S.; Schreibmayer, W. The TTX metabolite 4,9-anhydro-TTX is a highly specific blocker of the NaV1.6 voltage-dependent sodium channel. Am. J. Physiol. Cell Physiol. 2007, 293, C783–C789. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, N.; Zhu, H.L.; Yotsu-Yamashita, M.; Inai, T.; Cunnane, T.C. Resurgent-like currents in mouse vas deferens myocytes are mediated by NaV1.6 voltage-gated sodium channels. Pflügers Arch. Eur. J. Physiol. 2012, 464, 493–502. [Google Scholar] [CrossRef]
- Raman, I.M.; Bean, B.P. Resurgent sodium current and action potential formation in dissociated cerebellar Purkinje neurons. J. Neurosci. 1997, 17, 4517–4526. [Google Scholar] [PubMed]
- Dib-Hajj, S.D.; Cummins, T.R.; Black, J.A.; Waxman, S.G. Sodium channels in normal and pathological pain. Annu. Rev. Neurosci. 2010, 33, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Brackenbury, W.J. Voltage-gated sodium channels and metastatic disease. Channels 2012, 6, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. From ionic currents to molecular mechanisms: The structure and function of voltage-gated sodium channels. Neuron 2000, 26, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Casadei, J.M.; Gordon, R.D.; Barchi, R.L. Immunoaffinity isolation of Na+ channels from rat skeletal muscle. Analysis of subunits. J. Biol. Chem. 1986, 261, 4318–4323. [Google Scholar] [PubMed]
- Scheinman, R.I.; Auld, V.J.; Goldin, A.L.; Davidson, N.; Dunn, R.J.; Catterall, W.A. Developmental regulation of sodium channel expression in the rat forebrain. J. Biol. Chem. 1989, 264, 10660–10666. [Google Scholar] [PubMed]
- Isom, L.L. Sodium channel beta subunits: Anything but auxiliary. Neuroscientist 2001, 7, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Goldin, A.L. Resurgence of sodium channel research. Annu. Rev. Physiol. 2001, 63, 871–894. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, E.; Fujikawa, A.; Matsunaga, H.; Yasoshima, Y.; Sako, N.; Yamamoto, T.; Saegusa, C.; Noda, M. Nav2/NaG channel is involved in control of salt-intake behavior in the CNS. J. Neurosci. 2000, 20, 7743–7751. [Google Scholar] [PubMed]
- Hiyama, T.Y.; Yoshida, M.; Matsumoto, M.; Suzuki, R.; Matsuda, T.; Watanabe, E.; Noda, M. Endothelin-3 expression in the subfornical organ enhances the sensitivity of NaX, the brain sodium-level sensor, to suppress salt intake. Cell Metab. 2013, 17, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Ulbricht, W. Sodium channel inactivation: Molecular determinants and modulation. Physiol. Rev. 2005, 85, 1271–1301. [Google Scholar] [CrossRef] [PubMed]
- Fozzard, H.A.; Lipkind, G.M. The tetrodotoxin binding site is with the outer vestibule of the sodium channel. Mar. Drugs 2010, 8, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Chahine, M.; Ziane, R.; Vijayaragayan, K.; Okamura, Y. Regulation of NaV channels in sensory neurons. Trends Pharmacol. Sci. 2005, 26, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Burgess, D.L.; Kohrman, D.C.; Galt, J.; Plummer, N.W.; Jones, J.M.; Spear, B.; Meisler, M.H. Mutation of a new sodium channel gene, Scn8a, in the mouse mutant ‘motor endplate disease’. Nat. Genet. 1995, 10, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Schaller, K.L.; Krzemien, D.M.; Yarowsky, P.J.; Krueger, B.K.; Caldwell, J.H. A novel, abundant sodium channel expressed in neurons and glia. J. Neurosci. 1995, 15, 3231–3242. [Google Scholar] [PubMed]
- Caldwell, J.H.; Schaller, K.L.; Lasher, R.S.; Peles, E.; Levinson, S.R. Sodium channel NaV1.6 is localized at nodes of ranvier, dendrites, and synapses. Proc. Natl. Acad. Sci. USA 2000, 97, 5616–5620. [Google Scholar] [CrossRef] [PubMed]
- Tzoumaka, E.; Tischler, A.C.; Sangameswaran, L.; Eglen, R.M.; Hunter, J.C.; Novakovic, S.D. Differential distribution of the tetrodotoxin-sensitive rPN4/NaCh6/Scn8a sodium channel in the nervous system. J. Neurosci. Res. 2000, 60, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Black, J.A.; Renganathan, M.; Waxman, S.G. Sodium channel NaV1.6 is expressed along nonmyelinated axons and it contributes to conduction. Brain Res. Mol. Brain Res. 2002, 105, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Royeck, M.; Horstmann, M.T.; Remy, S.; Reitze, M.; Yaari, Y.; Beck, H. Role of axonal NaV1.6 sodium channels in action potential initiation of CA1 pyramidal neurons. J. Neurophysiol. 2008, 100, 2361–2380. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, T.; Kobayashi, K.; Noguchi, K. Laminae-specific distribution of alpha-subunits of voltage-gated sodium channels in the adult rat spinal cord. Neuroscience 2010, 169, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, T.; Kobayashi, K.; Yamanaka, H.; Obata, K.; Dai, Y.; Noguchi, K. Comparative study of the distribution of the alpha-subunits of voltage-gated sodium channels in normal and axotomized rat dorsal root ganglion neurons. J. Comp. Neurol. 2008, 510, 188–206. [Google Scholar] [CrossRef] [PubMed]
- Persson, A.K.; Black, J.A.; Gasser, A.; Cheng, X.; Fischer, T.Z.; Waxman, S.G. Sodium-calcium exchanger and multiple sodium channel isoforms in intra-epidermal nerve terminals. Mol. Pain 2010, 6, 84–97. [Google Scholar] [PubMed]
- Zhao, P.; Barr, T.P.; Hou, Q.; Dib-Hajj, S.D.; Black, J.A.; Albrecht, P.J.; Petersen, K.; Eisenberg, E.; Wymer, J.P.; Rice, F.L.; et al. Voltage-gated sodium channel expression in rat and human epidermal keratinocytes: Evidence for a role in pain. Pain 2008, 139, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Costigan, M.; Scholz, J.; Woolf, C.J. Neuropathic pain: A maladaptive response of the nervous system to damage. Annu. Rev. Neurosci. 2009, 32, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Kohrman, D.C.; Harris, J.B.; Meisler, M.H. Mutation detection in the med and medJ alleles of the sodium channel Scn8a. Unusual splicing due to a minor class AT-AC intron. J. Biol. Chem. 1996, 271, 17576–17581. [Google Scholar] [CrossRef] [PubMed]
- Sidman, R.L.; Cowen, J.S.; Eicher, E.M. Inherited muscle and nerve diseases in mice: A tabulation with commentary. Ann. N. Y. Acad. Sci. 1979, 317, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Duchen, L.W.; Stefani, E. Electrophysiological studies of neuromuscular transmission in hereditary “motor end-plate disease” of the mouse. J. Physiol. 1971, 212, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Angaut-Petit, D.; McArdle, J.J.; Mallart, A.; Bournaud, R.; Pincon-Raymond, M.; Riegers, F. Electrophysiological and morphological studies of a motor nerve in ‘motor endplate disease’ of the mouse. Proc. R. Soc. Lond. B Biol. Sci. 1982, 215, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Dick, D.J.; Boakes, R.J.; Harris, J.B. A cerebellar abnormality in the mouse with motor end-plate disease. Neuropathol. Appl. Neurobiol. 1985, 11, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.L.; Aishima, M.; Wassall, R.D.; Shibata, A.; Iwasa, K.; Nomura, M.; Nagao, M.; Sueishi, K.; Cunnane, T.C.; Teramoto, N. Molecular and biophysical properties of voltage-gated Na+ currents in murine vas deferens. Biophys. J. 2008, 94, 3340–3351. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhu, H.L.; Shibata, A.; Inai, T.; Nomura, M.; Shibata, Y.; Brock, J.A.; Teramoto, N. Characterization of NaV1.6-mediated Na+ currents in smooth muscle cells isolated from mouse vas deferens. J. Cell. Physiol. 2010, 233, 234–243. [Google Scholar]
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O. Tetrodotoxin. VII. On the structure of tetrodotoxin and its derivatives. Chem. Pharm. Bull. 1964, 12, 1357–1374. [Google Scholar] [CrossRef] [PubMed]
- Woodward, R.B. The structure of tetrodotoxin. Pure Appl. Chem. 1964, 9, 49–74. [Google Scholar] [CrossRef]
- Goto, T.; Kishi, Y.; Takahashi, S.; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yotsu, M.; Murata, M.; Naoki, H. New tetrodotoxin analogue from the newt Cynops ensicauda. J. Am. Chem. Soc. 1988, 110, 2344–2345. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M. Chemistry of puffer fish toxin. J. Toxicol. Toxin Rev. 2001, 20, 51–66. [Google Scholar] [CrossRef]
- Kudo, Y.; Yamashita, Y.; Mebs, D.; Cho, Y.; Konoki, K.; Yasumoto, T.; Yotsu-Yamashita, M. C5–C10 Directly bonded tetrodotoxin analogues: Possible biosynthetic precursors of tetrodotoxin from newts. Angew. Chem. Int. Ed. 2014, 53, 14546–14549. [Google Scholar] [CrossRef]
- Kono, M.; Matsui, T.; Furukawa, K.; Takase, T.; Yamamori, K.; Kaneda, H.; Aoki, D.; Jang, J.H.; Yotsu-Yamashita, M. Examination of transformation among tetrodotoxin and its analogs in the living cultured juvenile puffer fish, kusafugu, Fugu niphobles by intramuscular administration. Toxicon 2008, 52, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Goto, A.; Nakagawa, T. Isolation of 4-S-cysteinyltetrodotoxin from the liver of the puffer fish Fugu pardalis, and formation of the adducts of 4,9-anhydrotetrodotoxin with thiols. Chem. Res. Toxicol. 2005, 18, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Isobe, M. Synthesis of tetrodotoxin, a classic but still fascinating natural product. Chem. Rec. 2013, 13, 286–302. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teramoto, N.; Yotsu-Yamashita, M. Selective Blocking Effects of 4,9-Anhydrotetrodotoxin, Purified from a Crude Mixture of Tetrodotoxin Analogues, on NaV1.6 Channels and Its Chemical Aspects. Mar. Drugs 2015, 13, 984-995. https://doi.org/10.3390/md13020984
Teramoto N, Yotsu-Yamashita M. Selective Blocking Effects of 4,9-Anhydrotetrodotoxin, Purified from a Crude Mixture of Tetrodotoxin Analogues, on NaV1.6 Channels and Its Chemical Aspects. Marine Drugs. 2015; 13(2):984-995. https://doi.org/10.3390/md13020984
Chicago/Turabian StyleTeramoto, Noriyoshi, and Mari Yotsu-Yamashita. 2015. "Selective Blocking Effects of 4,9-Anhydrotetrodotoxin, Purified from a Crude Mixture of Tetrodotoxin Analogues, on NaV1.6 Channels and Its Chemical Aspects" Marine Drugs 13, no. 2: 984-995. https://doi.org/10.3390/md13020984
APA StyleTeramoto, N., & Yotsu-Yamashita, M. (2015). Selective Blocking Effects of 4,9-Anhydrotetrodotoxin, Purified from a Crude Mixture of Tetrodotoxin Analogues, on NaV1.6 Channels and Its Chemical Aspects. Marine Drugs, 13(2), 984-995. https://doi.org/10.3390/md13020984