
Biotechnological Production of Docosahexaenoic Acid Using Aurantiochytrium limacinum: Carbon Sources Comparison And Growth Characterization

Abstract
:
1. Introduction
1.1. Crude Glycerol Valorisation
1.2. Crude Glycerol to Obtain Long Chain Poly Unsaturated Fatty Acids
2. Results and Discussion

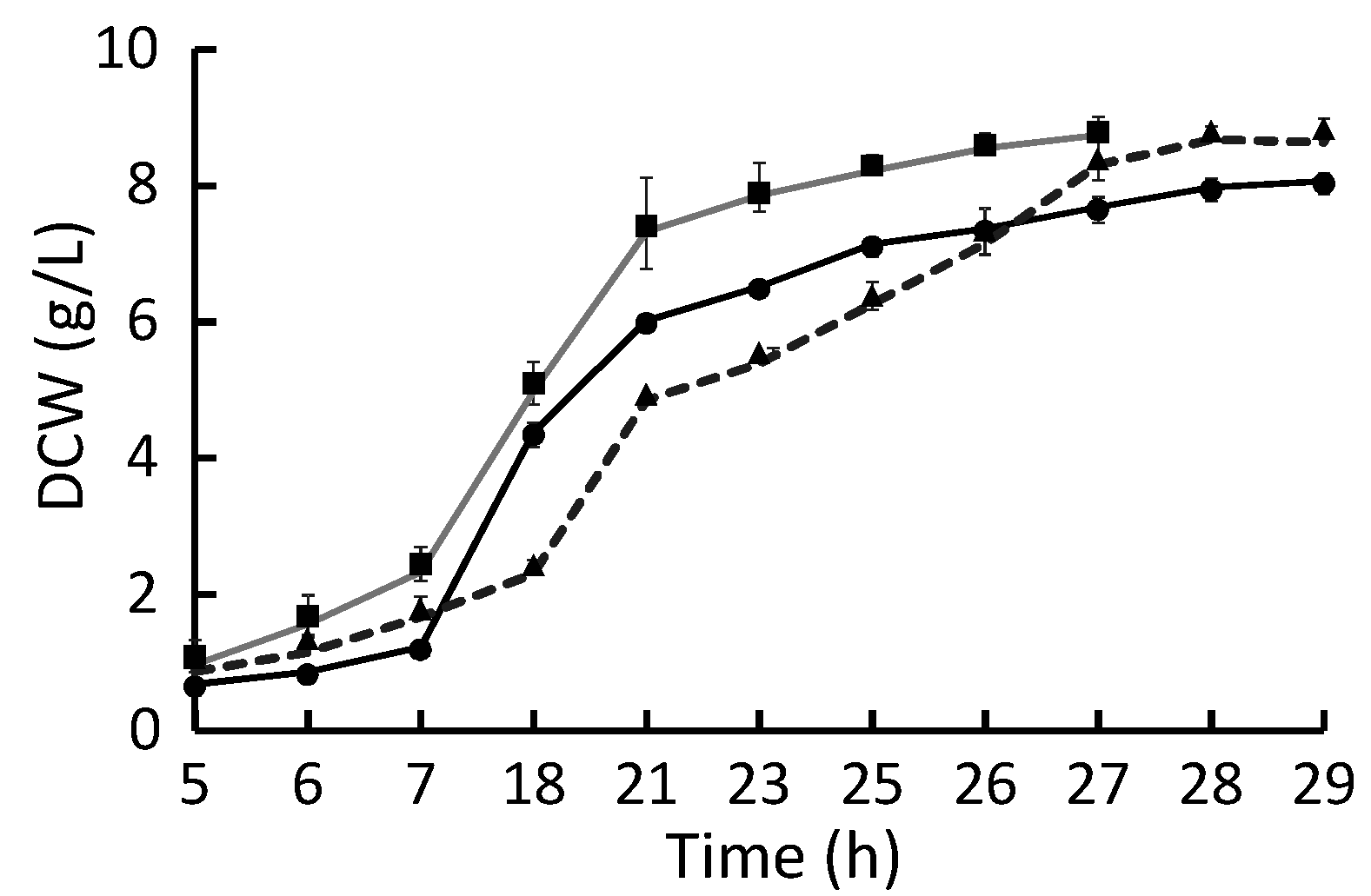
2.1. Growth Rate
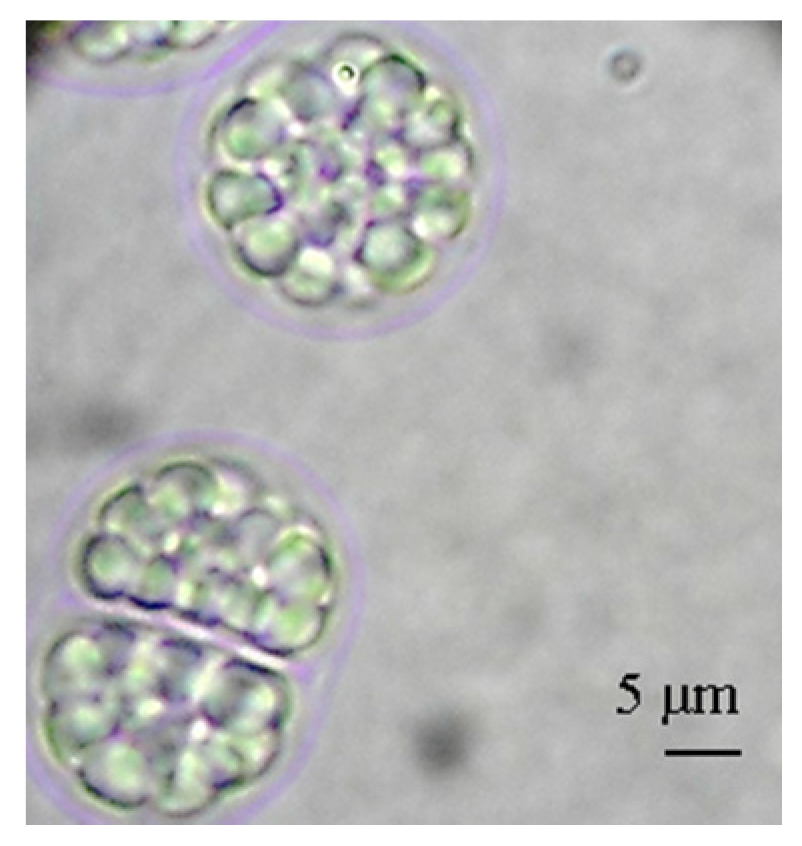
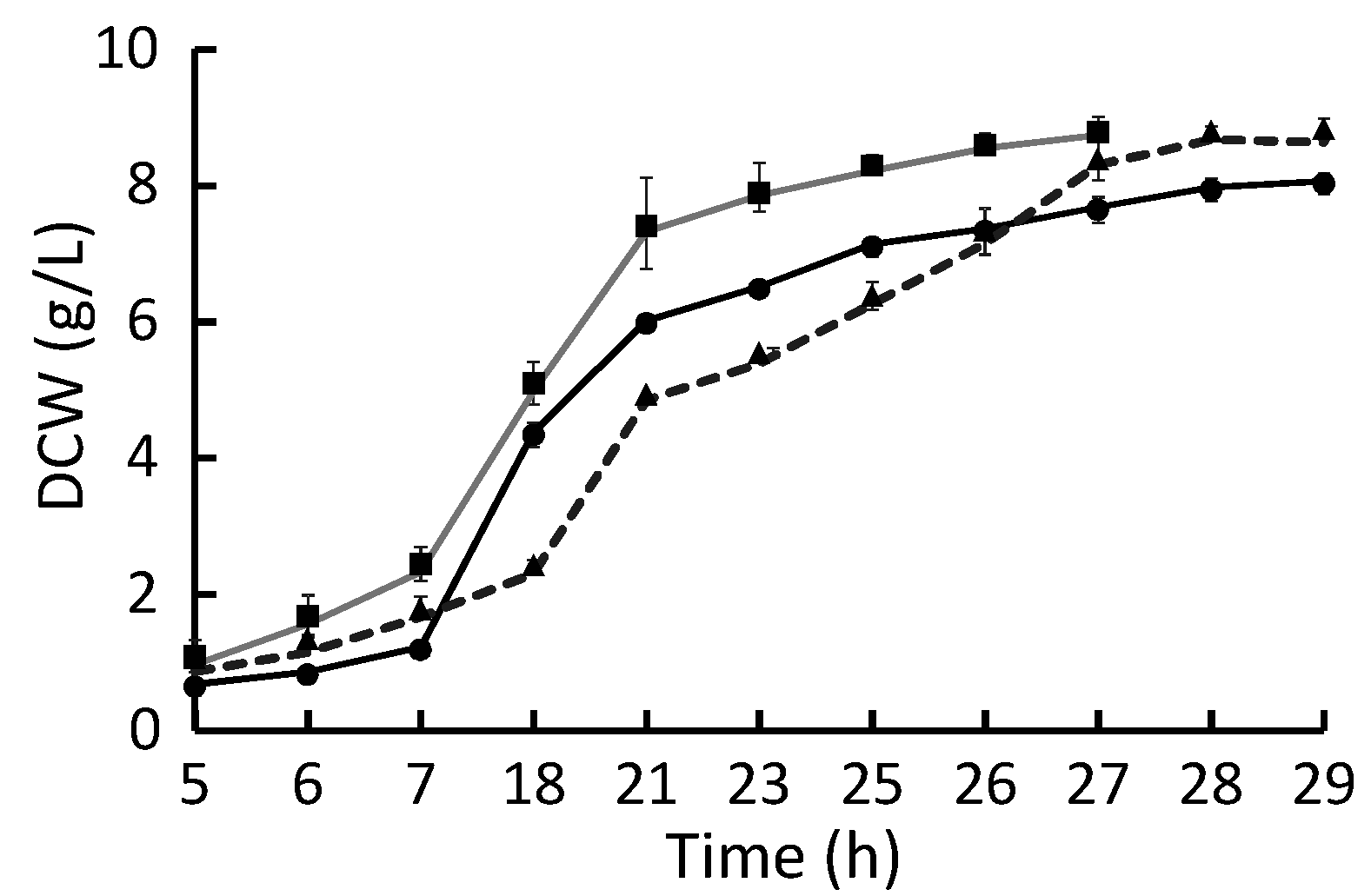
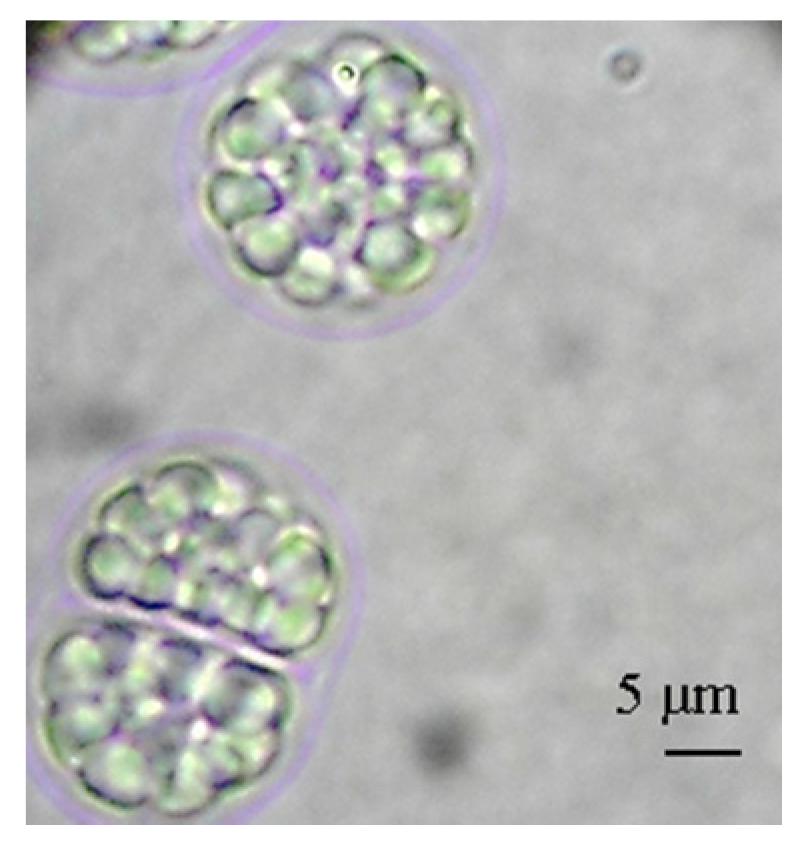
2.2. Growth Yield

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucose | Glycerol | Crude Glycerol | ||||
|---|---|---|---|---|---|---|
| X (g/L) | 8.83 | ±0.15 | 8.21 | ±0.17 | 8.86 | ±0.12 |
| YX/S (g/g) | 0.84 | ±0.05 | 0.74 | ±0.04 | 0.85 | ±0.1 |
| μmax (h−1) | 0.21 | ±0.02 | 0.18 | ±0.01 | 0.19 | ±0.02 |
| μnet (h−1) | 0.10 | ±0.005 | 0.12 | ±0.01 | 0.10 | ±0.01 |
| KS (g/L) | 2.5 | ±0.5 | 5.0 | ±1.5 | 8.0 | ±1.5 |
| YP/X (g/g) | 0.14 | ±0.02 | 0.15 | ±0.04 | 0.15 | ±0.02 |
| Carbon Source | Time (h) | So (g/L) | X (g/L) | Yx/s (g/g) | Yp/x (g/g) | μnet (h−1) | Reference |
|---|---|---|---|---|---|---|---|
| Glucose | 125 | 150 | 59.2 | 0.39 * | 0.26 * | 0.05 * | [39] |
| 50 | 40 | 21 | 0.52 * | 0.22 * | 0.11 * | [35] | |
| 100 | 120 | 50 | 0.41 * | 0.29 * | 0.11 | [40] | |
| 30 | 10 | 8.83 | 0.84 | 0.14 | 0.10 | This study | |
| Glycerol | 198 | 100 | 32.8 | 0.33 * | 0.18 * | 0.023 * | [37] |
| 30 | 10 | 8.21 | 0.74 | 0.15 | 0.12 | This study | |
| Crude glycerol | 30 | 10 | 8.86 | 0,85 | 0.15 | 0.10 | This study |
2.3. Affinity Constant
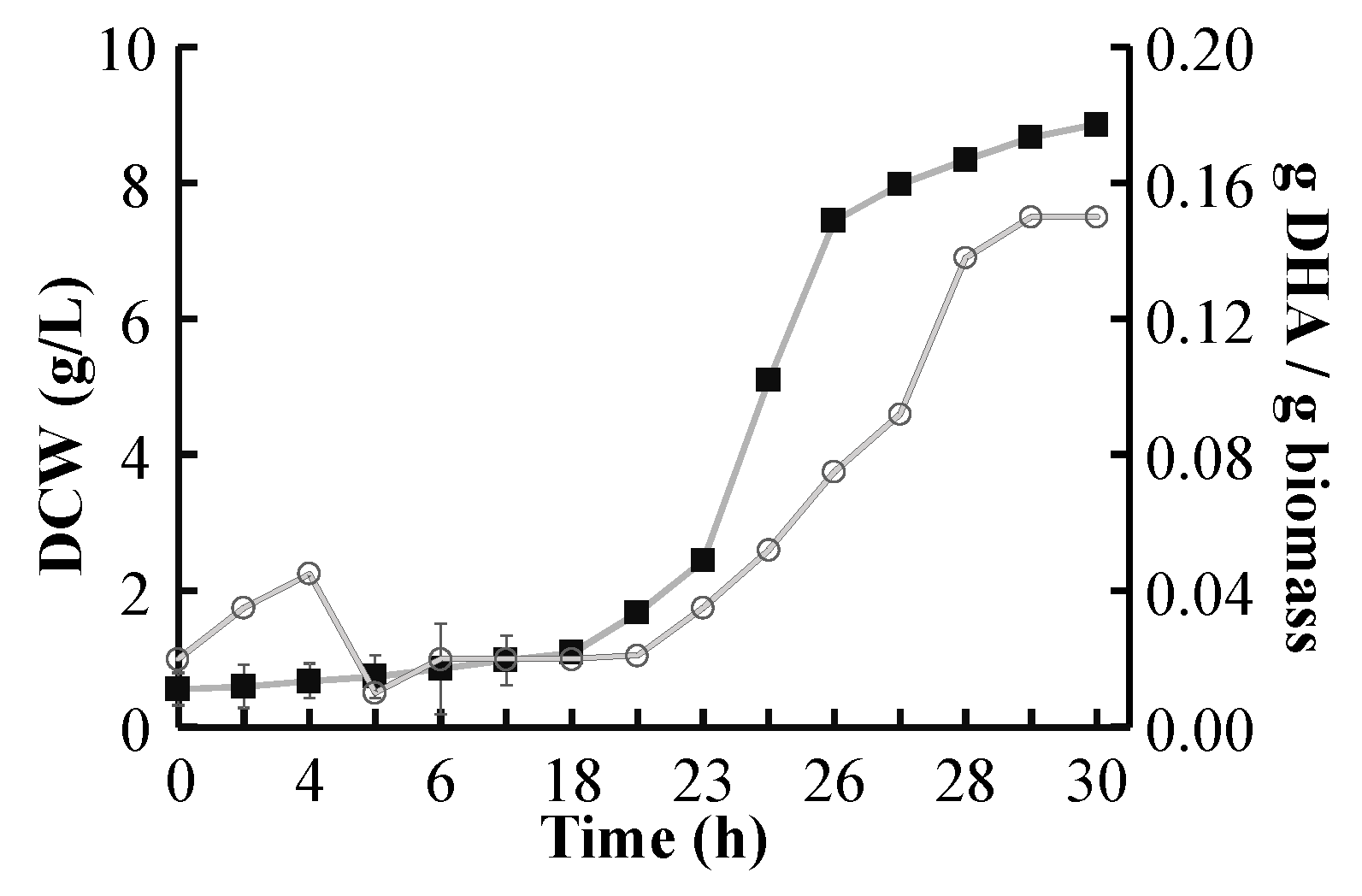
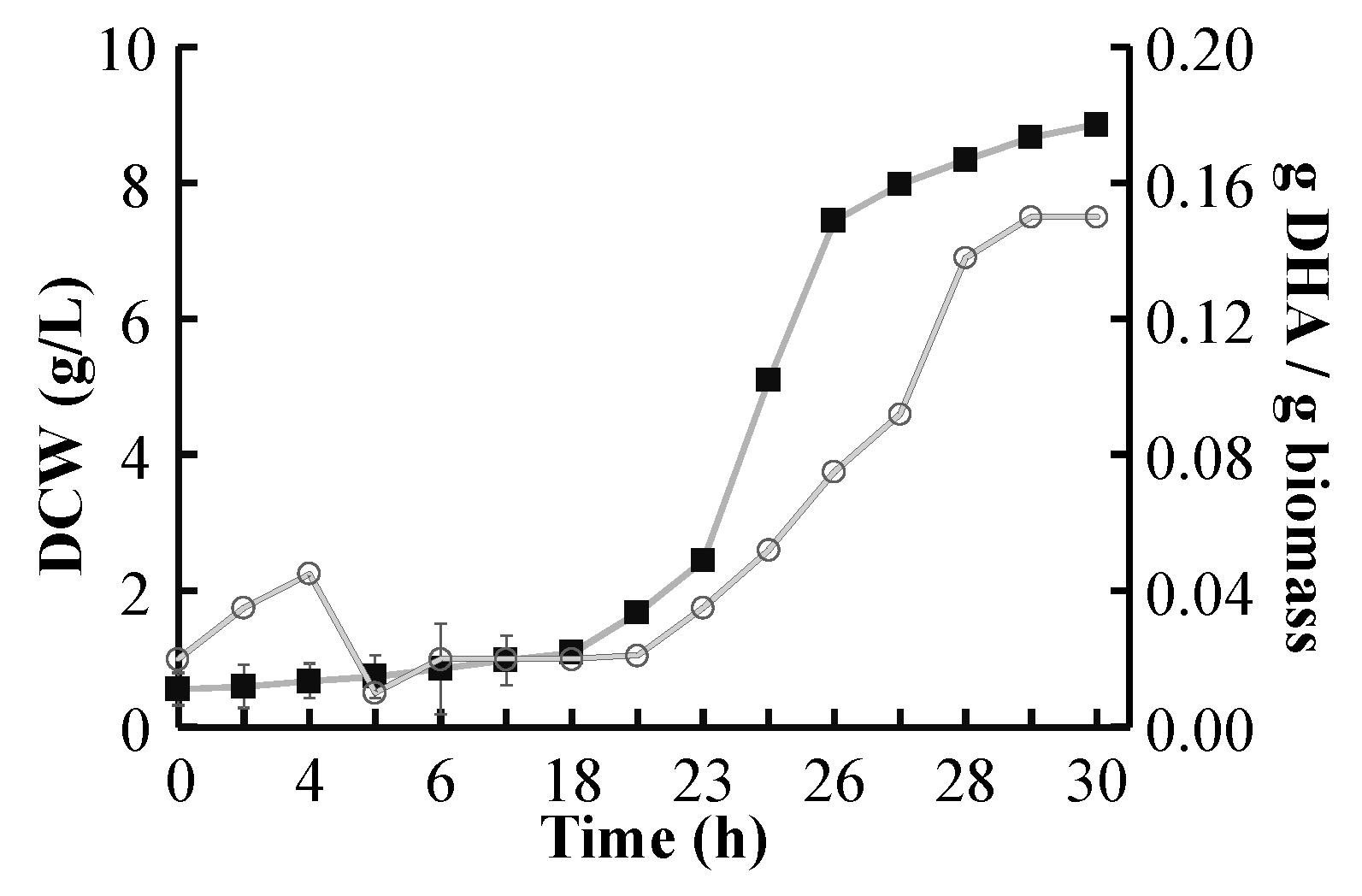
2.4. DHA Yield
3. Experimental Section
3.1. Strain and Culture Media
3.2. Cultivation
3.3. Glucose Cultivation
3.4. Glycerol Cultivation
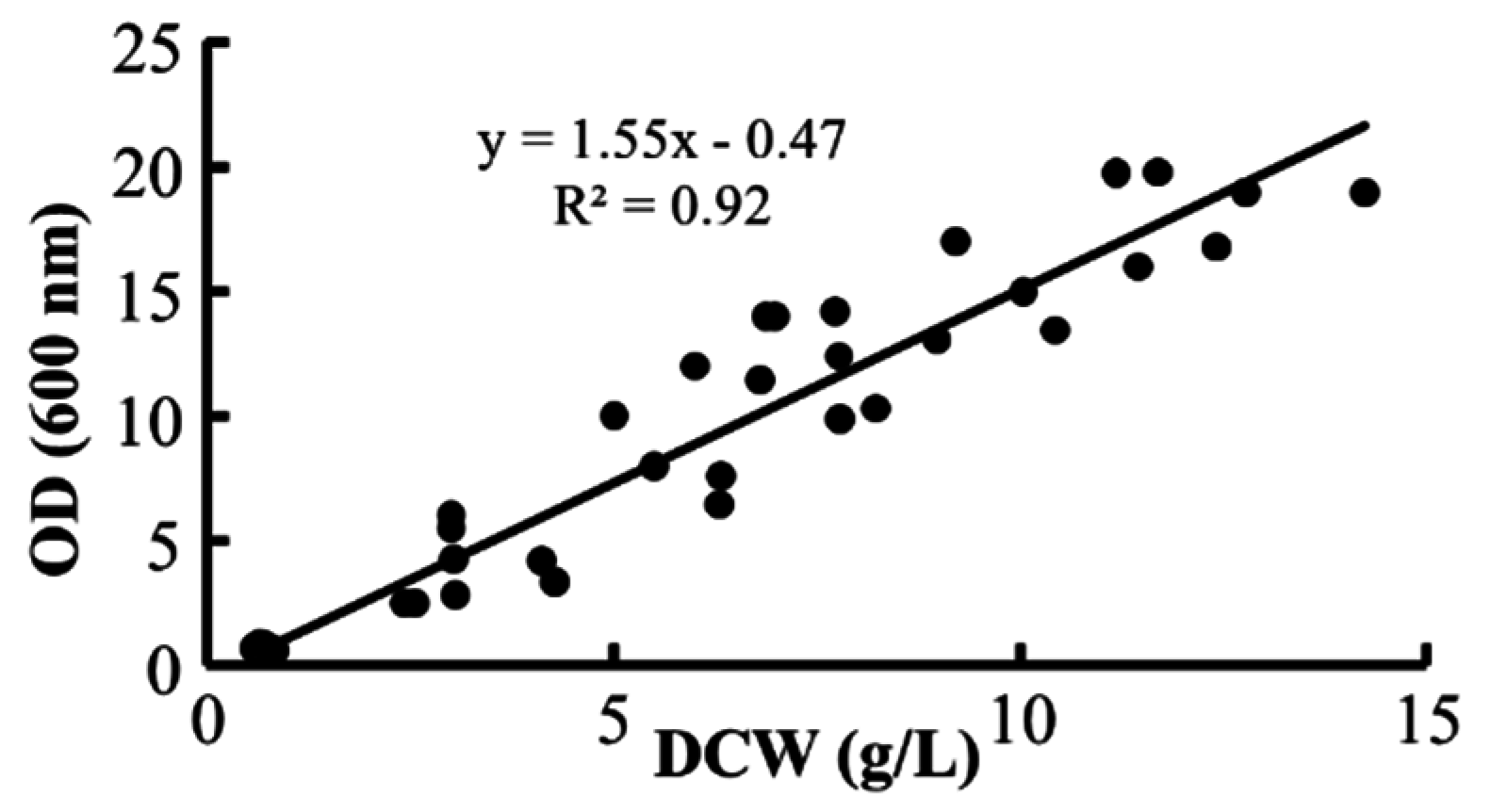
3.5. Analysis
3.6. Kinetic Parameters Calculation
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pyle, D.J. Use of Biodiesel-Derived Crude Glycerol for the Production of Omega-3 Polyunsaturated Fatty Acids by the Microalga Schizochytrium limacinum. Master’s Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 18 April 2008. [Google Scholar]
- Thompson, J.; He, B. Characterization of crude glycerol from biodiesel production from multiple feedstocks. Appl. Eng. Agric. 2006, 22, 2006–2007. [Google Scholar] [CrossRef]
- Johnson, D.T.; Taconi, K.A. The glycerin glut: Options for the value-added conversion of crude glycerol resulting from biodiesel production. Environ. Prog. 2007, 26, 338–348. [Google Scholar] [CrossRef]
- Prielipp, G.G.; Keller, H.H. Purification of crude glycerin by ion exclusion. J. Am. Oil Chem. Soc. 1956, 33, 103–108. [Google Scholar] [CrossRef]
- André, A.; Chatzifragkou, A.; Diamantopoulou, P.; Sarris, D.; Philippoussis, A.; Galiotou-Panayotou, M.; Komaitis, M.; Papanikolaou, S. Biotechnological conversions of bio-diesel-derived crude glycerol by Yarrowia lipolytica strains. Eng. Life Sci. 2009, 9, 468–478. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, Y.; Trushenski, J.; Blackburn, J.W. Converting crude glycerol derived from yellow grease to lipids through yeast fermentation. Bioresour. Technol. 2010, 101, 7581–7586. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Biotechnological valorization of biodiesel derived glycerol waste through production of single cell oil and citric acid by Yarrowia lipolytica. Lipid Technol. 2009, 21, 83–87. [Google Scholar] [CrossRef]
- Cardona, C.; Posada, J.; Montoya, M. Use of Glycerol from biodiesel production: Conversion to added value products. In Proceedings of the European Congress of Chemical Engineering, Copenhagen, Denmark, 16–20 September 2007.
- Casali, S.; Gungormusler, M.; Bertin, L.; Fava, F.; Azbar, N. Development of a biofilm technology for the production of 1,3-propanediol (1,3-PDO) from crude glycerol. J. Biochem. Eng. 2012, 64, 84–90. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Yarrowia lipolytica as a potential producer of citric acid from raw glycerol. J. Appl. Microbiol. 2002, 92, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Himmi, E.H.; Bories, A.; Barbirato, F. Nutrient requirements for glycerol conversion to 1,3-propanediol by Clostridium butyricum. Bioresour. Technol. 1998, 67, 123–128. [Google Scholar] [CrossRef]
- Dietz, D.; Zeng, A.P. Efficient production of 1,3-propanediol from fermentation of crude glycerol with mixed cultures in a simple medium. Bioprocess Biosyst. Eng. 2014, 37, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Pflügl, S.; Marx, H.; Mattanovich, D.; Sauer, M. Heading for an economic industrial upgrading of crude glycerol from biodiesel production to 1,3-propanediol by Lactobacillus diolivorans. Bioresour. Technol. 2014, 152, 499–504. [Google Scholar] [PubMed]
- Cavalheiro, J.M.B.T.; de Almeida, M.C.M.D.; Grandfils, C.; da Fonseca, M.M.R. Poly(3-hydroxybutyrate) production by Cupriavidus necator using waste glycerol. Process Biochem. 2009, 44, 509–515. [Google Scholar] [CrossRef]
- Mothes, G.; Schnorpfeil, C.; Ackermann, J.-U. Production of PHB from Crude Glycerol. Eng. Life Sci. 2007, 7, 475–479. [Google Scholar] [CrossRef]
- Taccari, M.; Canonico, L.; Comitini, F.; Mannazzu, I.; Ciani, M. Screening of yeasts for growth on crude glycerol and optimization of biomass production. Bioresour. Technol. 2012, 110, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Moita, R.; Freches, A.; Lemos, P.C. Crude glycerol as feedstock for polyhydroxyalkanoates production by mixed microbial cultures. Water Res. 2014, 58, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Canadas, R.F.; Cavalheiro, J.M.B.T.; Guerreiro, J.D.T.; de Almeida, M.C.M.D.; Pollet, E.; da Silva, C.L.; da Fonseca, M.M.R.; Ferreira, F.C. Polyhydroxyalkanoates: Waste glycerol upgrade into electrospun fibrous scaffolds for stem cells culture. Int. J. Biol. Macromol. 2014, 71, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Mantzouridou, F.; Naziri, E.; Tsimidou, M.Z. Industrial glycerol as a supplementary carbon source in the production of β-carotene by Blakeslea trispora. J. Agric. Food Chem. 2008, 56, 2668–2675. [Google Scholar] [CrossRef] [PubMed]
- Abad, S.; Turon, X. Valorization of biodiesel derived glycerol as a carbon source to obtain added-value metabolites: Focus on polyunsaturated fatty acids. Biotechnol. Adv. 2012, 30, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Athalye, S.K.; Garcia, R.A.; Wen, Z. Use of biodiesel-derived crude glycerol for producing eicosapentaenoic acid (EPA) by fungus Pythium irregulare. J. Agric. Food Chem. 2009, 57, 2739–2744. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Pyle, D.; Wen, Z.; Frear, C.; Chen, S.; Zhiyou, W.; Shulin, C. A laboratory study of producing docosahexaenoic acid from biodiesel-waste glycerol by microalgal fermentation. Process Biochem. 2007, 42, 1537–1545. [Google Scholar] [CrossRef]
- Ethier, S.; Woisard, K.; Vaughan, D.; Wen, Z. Continuous culture of the microalgae Schizochytrium limacinum on biodiesel-derived crude glycerol for producing docosahexaenoic acid. Bioresour. Technol. 2010, 102, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Pyle, D.J.; Garcia, R.A.; Wen, Z. Producing docosahexaenoic acid (DHA)-rich algae from biodiesel-derived crude glycerol: Effects of impurities on DHA production and algal biomass composition. J. Agric. Food Chem. 2008, 56, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Braune, B.M.; Outridge, P.M.; Fisk, A.T.; Muir, D.C.G.; Helm, P.A.; Hobbs, K.; Hoekstra, P.F.; Kuzyk, Z.A.; Kwan, M.; Letcher, R.J.; et al. Persistent organic pollutants and mercury in marine biota of the Canadian Arctic: An overview of spatial and temporal trends. Sci. Total Environ. 2005, 351–352, 4–56. [Google Scholar] [CrossRef] [PubMed]
- Muir, D.; Wagemann, R.; Hargrave, B.T.; Thomas, D.J.; Peakall, D.B.; Norstrom, R.J. Arctic marine ecosystem contamination. Sci. Total Environ. 1992, 122, 75–134. [Google Scholar] [CrossRef]
- Muir, D.; Braune, B.; Demarch, B.; Norstrom, R.; Wagemann, R.; Lockhart, L.; Hargrave, B.; Bright, D.; Addison, R.; Payne, J.; et al. Spatial and temporal trends and effects of contaminants in the Canadian Arctic marine ecosystem: A review. Sci. Total Environ. 2005, 230, 83–144. [Google Scholar] [CrossRef]
- Jacobs, M.N.; Coraci, M.; Gheorghe, A.; Schepens, A.; Covaci, A.; Schepens, P. Time trend investigation of PCBs, PBDEs, and organochlorine pesticides in selected n-3 polyunsaturated fatty acid rich dietary fish oil and vegetable oil supplements; nutritional relevance for human essential n-3 fatty acid requirements. J. Agric. Food Chem. 2004, 52, 1780–1788. [Google Scholar] [CrossRef] [PubMed]
- Turon, X. Algae oil, a valuable source of long chain polyunsaturated fatty acids. J. Life Med. 2013, 1, 11–14. [Google Scholar] [CrossRef]
- Singh, A.; Wilson, S.; Ward, O.P. Docosahexaenoic Thraustochytrium sp. ATCC 20892, acid (DHA) production. World J. Microbiol. Biotechnol. 1996, 12, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Honda, D.; Yokochi, T.; Nakahara, T.; Erata, M.; Higashihara, T. Schizochytrium limacinum sp. nov., a new thraustochytrid from a mangrove area in the west Pacific Ocean. Mycol. Res. 1998, 102, 439–448. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1997, 49, 72–76. [Google Scholar] [CrossRef]
- Jakobsen, A.N.; Aasen, I.M.; Josefsen, K.D.; Strøm, A.R. Accumulation of docosahexaenoic acid-rich lipid in thraustochytrid Aurantiochytrium sp. strain T66: Effects of N and P starvation and O2 limitation. Appl. Microbiol. Biotechnol. 2008, 80, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Nagano, N.; Taoka, Y.; Honda, D.; Hayashi, M. Optimization of culture conditions for growth and docosahexaenoic acid production by a marine thraustochytrid, Aurantiochytrium limacinum mh0186. J. Oleo Sci. 2009, 58, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Perveen, Z.; Ando, H.; Ueno, A.; Ito, Y.; Yamamoto, Y.; Yamada, Y.; Takagi, T.; Kaneko, T.; Kogame, K.; Okuyama, H. Isolation and characterization of a novel thraustochytrid-like microorganism that efficiently produces docosahexaenoic acid. Biotechnol. Lett. 2006, 28, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Liu, Y.; Frear, C.; Chen, S. Study of a two-stage growth of DHA-producing marine algae Schizochytrium limacinum SR21 with shifting dissolved oxygen level. Appl. Microbiol. Biotechnol. 2009, 81, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Ji, X.-J.; Ren, L.-J.; Nie, Z.-K.; Feng, Y.; Wu, W.-J.; Ouyang, P.-K.; Huang, H. Enhancement of docosahexaenoic acid production by Schizochytrium sp. using a two-stage oxygen supply control strategy based on oxygen transfer coefficient. Lett. Appl. Microbiol. 2010, 52, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, T.; Tanaka, S.; Yokochi, T.; Nakahara, T.; Higashihara, T. Production of high yields of docosahexaenoic acid by Schizochytrium sp. strain SR21. J. Am. Oil Chem. Soc. 1997, 74, 1431–1434. [Google Scholar] [CrossRef]
- Rosa, S.M.; Soria, M.A.; Vélez, C.G.; Galvagno, M.A. Improvement of a two-stage fermentation process for docosahexaenoic acid production by Aurantiochytrium limacinum SR21 applying statistical experimental designs and data analysis. Bioresour. Technol. 2010, 101, 2367–2374. [Google Scholar] [CrossRef] [PubMed]
- Unagul, P.; Assantachai, C.; Phadungruengluij, S.; Suphantharika, M.; Tanticharoen, M.; Verduyn, C. Coconut water as a medium additive for the production of docosahexaenoic acid (C22:6n3) by Schizochytrium mangrovei Sk-02. Bioresour. Technol. 2007, 98, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, R.; Honda, D. Taxonomic rearrangement of the genus Schizochytrium sensu lato based on morphology, chemotaxonomic characteristics, and 18S rRNA gene phylogeny (Thraustochytriaceae, Labyrinthulomycetes): Emendation for Schizochytrium and erection of Aurantiochytrium and Oblongichytrium gen. nov. Mycoscience 2007, 48, 199–211. [Google Scholar]
- Bauer, R.; Katsikis, N.; Varga, S.; Hekmat, D. Study of the inhibitory effect of the product dihydroxyacetone on Gluconobacter oxydans in a semi-continuous two-stage repeated-fed-batch process. Bioprocess Biosyst. Eng. 2005, 28, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Abad, S.; Pérez, X.; Planas, A.; Turon, X. Rapid monitoring of glycerol in fermentation growth media: Facilitating crude glycerol bioprocess development. Talanta 2014, 121, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Miller, G. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Indarti, E.; Majid, M.I.A.; Hashim, R.; Chong, A. Direct FAME synthesis for rapid total lipid analysis from fish oil and cod liver oil. J. Food Compos. Anal. 2005, 18, 161–170. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abad, S.; Turon, X. Biotechnological Production of Docosahexaenoic Acid Using Aurantiochytrium limacinum: Carbon Sources Comparison And Growth Characterization. Mar. Drugs 2015, 13, 7275-7284. https://doi.org/10.3390/md13127064
Abad S, Turon X. Biotechnological Production of Docosahexaenoic Acid Using Aurantiochytrium limacinum: Carbon Sources Comparison And Growth Characterization. Marine Drugs. 2015; 13(12):7275-7284. https://doi.org/10.3390/md13127064
Chicago/Turabian StyleAbad, Sergi, and Xavier Turon. 2015. "Biotechnological Production of Docosahexaenoic Acid Using Aurantiochytrium limacinum: Carbon Sources Comparison And Growth Characterization" Marine Drugs 13, no. 12: 7275-7284. https://doi.org/10.3390/md13127064
APA StyleAbad, S., & Turon, X. (2015). Biotechnological Production of Docosahexaenoic Acid Using Aurantiochytrium limacinum: Carbon Sources Comparison And Growth Characterization. Marine Drugs, 13(12), 7275-7284. https://doi.org/10.3390/md13127064