
Cell Death Inducing Microbial Protein Phosphatase Inhibitors—Mechanisms of Action


Abstract
:
1. Introduction
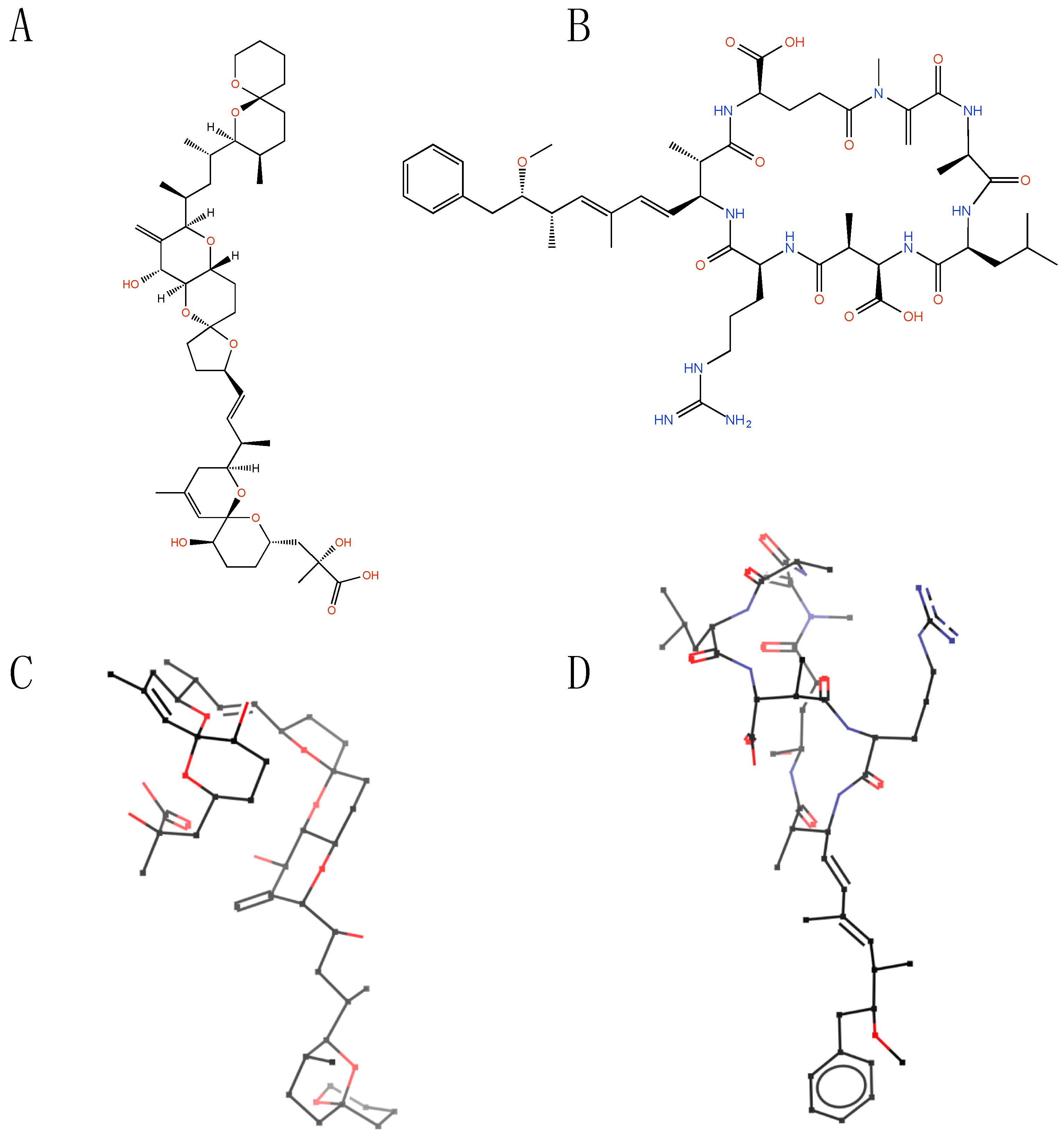
2. Structural Features of Microbial Toxins
3. Comparative Activity of OA and MC-LR against Various Protein Phosphatases
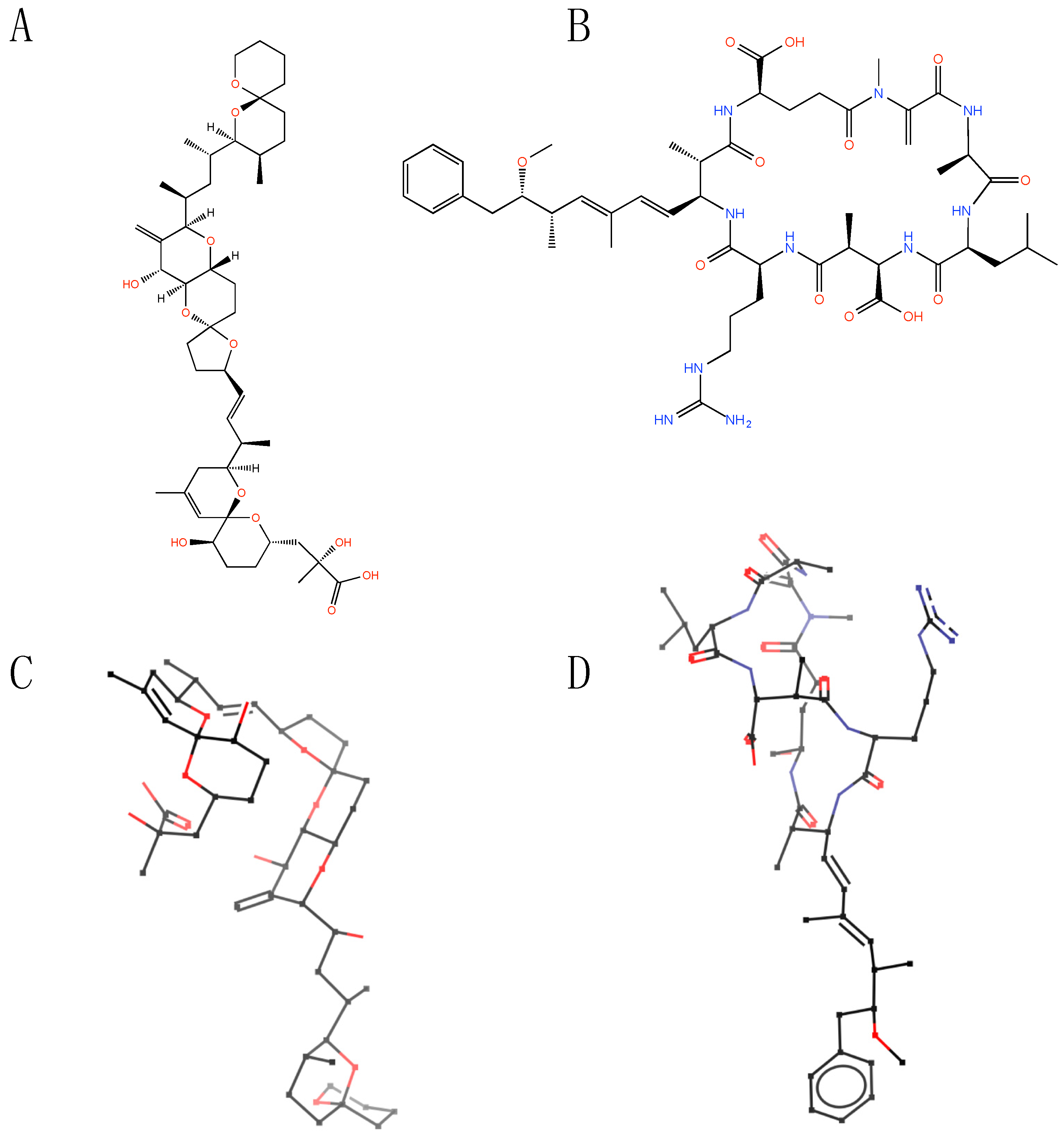

{kind=link}
{kind=link}
{kind=link}
| PP family Member 1 | IC50 (nM) | |||
|---|---|---|---|---|
| Okadaic acid | References | MC-LR | References | |
| PP1 | 15–50 | [17,18] | 0.3–2 | [15,17,18,19,20] |
| PP2A | 0.1–0.3 | [17,18] | 0.05–1 | [17,18,19,20] |
| PP2B | ~4000 | [17] | ~2000 | [17,19] |
| PP3 | 3.7–4 | [18] | 0.2 | [18] |
| PP4 | 0.1 | [17,21] | 0.15 | [17,21] |
| PP5 | 3.5 | [17] | 1.0 | [17] |
| PP7 | >1000 | [17,22] | >1000 | [17,22] |
| Compound | IC50 (nM) |
|---|---|
| Nodularin | 0.015 |
| MC-LR | 0.017 |
| MC-YR | 0.075 |
| OA | 0.100 |
| CalyculinA | 0.250 |
| Tautomycin | 0.560 |
4. Cellular Uptake of Microbial Phosphatase Inhibitors
5. Features of Microbial PP Inhibitor-Induced Cell Death
6. The Pathogenesis of the Acute Hepatotoxicity of MC and Nodularin
7. PP Inhibitors as Tumor Promoters
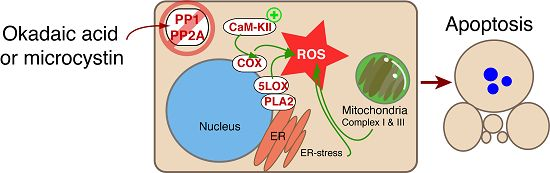
8. The Role of CaM-KII in PP Inhibitor Action—Is CaM-KII an Amplifier of ROS Generation?
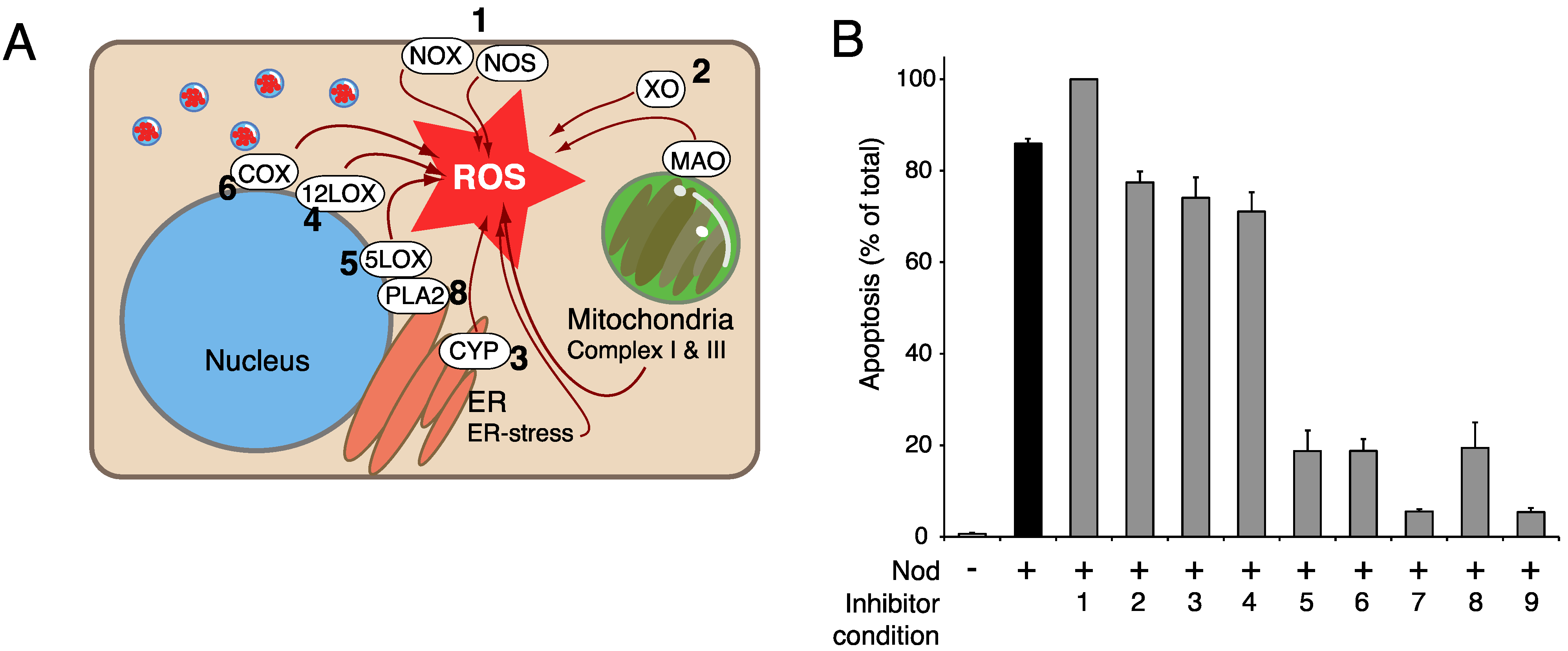
9. The Potential Role of Perturbed Phospholipid Metabolism in PPI-Induced Hepatocyte Apoptosis
10. Gimap5 (Ian5, IROD) is an Inhibitor of OA-Induced Apoptosis
11. Concluding Remarks
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Francis, G. Poisonous Australian lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef]
- Heroes, E.; Lesage, B.; Gornemann, J.; Beullens, M.; van Meervelt, L.; Bollen, M. The PP1 binding code: A molecular-lego strategy that governs specificity. FEBS J. 2013, 280, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Seshacharyulu, P.; Pandey, P.; Datta, K.; Batra, S.K. Phosphatase: PP2A structural importance, regulation and its aberrant expression in cancer. Cancer Lett. 2013, 335, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, K.; Scheuer, P.J.; Tsukitani, Y.; Kikuchi, H.; Vanengen, D.; Clardy, J.; Gopichand, Y.; Schmitz, F.J. Okadaic Acid, a Cyto-Toxic Polyether from 2 Marine Sponges of the Genus Halichondria. J. Am. Chem. Soc. 1981, 103, 2469–2471. [Google Scholar] [CrossRef]
- Bishop, C.T.; Anet, E.F.L.; Gorham, P.R. Isolation and Identification of the Fast-Death Factor in Microcystis aeruginosa NRC-1. Can. J. Biochem. Physiol. 1959, 37, 453–471. [Google Scholar] [PubMed]
- Botes, D.P.; Tuinman, A.A.; Wessels, P.L.; Viljoen, C.C.; Kruger, H.; Williams, D.H.; Santikarn, S.; Smith, R.J.; Hammond, S.J. The Structure of Cyanoginosin-LA, a Cyclic Heptapeptide Toxin from the Cyanobacterium Microcystis aeruginosa. J. Chem. Soc. Perkin Trans. 1 1984. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Eschedor, J.T.; Patterson, G.M.L.; Moore, R.E. Toxicity and Partial Structure of a Hepatotoxic Peptide Produced by the Cyanobacterium Nodularia spumigena Mertens Emend L575 from New Zealand. Appl. Environ. Microbiol. 1988, 54, 2257–2263. [Google Scholar] [PubMed]
- Eriksson, J.E.; Meriluoto, J.A.O.; Kujari, H.P.; Osterlund, K.; Fagerlund, K.; Hallbom, L. Preliminary Characterization of a Toxin Isolated from the Cyanobacterium Nodularia spumigena. Toxicon 1988, 26, 161–166. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Harada, K.; Namikoshi, M.; Chen, C.; Harvis, C.A.; Munro, M.H.G.; Blunt, J.W.; Mulligan, P.E.; Beasley, V.R.; Dahlem, A.M.; et al. Nodularin, Microcystin, and the Configuration of Adda. J. Am. Chem. Soc. 1988, 110, 8557–8558. [Google Scholar] [CrossRef]
- Connor, J.H.; Kleeman, T.; Barik, S.; Honkanen, R.E.; Shenolikar, S. Importance of the beta12-beta13 loop in protein phosphatase-1 catalytic subunit for inhibition by toxins and mammalian protein inhibitors. J. Biol. Chem. 1999, 274, 22366–22372. [Google Scholar] [CrossRef] [PubMed]
- Dawson, J.F.; Holmes, C.F. Molecular mechanisms underlying inhibition of protein phosphatases by marine toxins. Front. Biosci. 1999, 4, 646–658. [Google Scholar] [CrossRef]
- Gauss, C.M.; Sheppeck, J.E.; Nairn, A.C.; Chamberlin, R. A molecular modeling analysis of the binding interactions between the okadaic acid class of natural product inhibitors and the Ser-Thr phosphatases, PP1 and PP2A. Bioorg. Med. Chem. 1997, 5, 1751–1773. [Google Scholar] [CrossRef]
- Goldberg, J.; Huang, H.-b.; Kwon, Y.-g.; Greengard, P.; Nairn, A.C.; Kuriyan, J. Three-dimensional structure of the catalytic subunit of protein serine/threonine phosphatase-1. Nature 1995, 376, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Maynes, J.T.; Bateman, K.S.; Cherney, M.M.; Das, A.K.; Luu, H.A.; Holmes, C.F.; James, M.N. Crystal structure of the tumor-promoter okadaic acid bound to protein phosphatase-1. J. Biol. Chem. 2001, 276, 44078–44082. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, R.W.; Dalby, K.N.; Campbell, D.G.; Cohen, P.T.W.; Cohen, P.; MacKintosh, C. The cyanobacterial toxin microcystin binds covalently to cysteine-273 on protein phosphatase 1. FEBS Lett. 1995, 371, 236–240. [Google Scholar] [CrossRef]
- Honkanen, R.E.; Dukelow, M.; Zwiller, J.; Moore, R.E.; Khatra, B.S.; Boynton, A.L. Cyanobacterial nodularin is a potent inhibitor of type 1 and type 2A protein phosphatases. Mol. Pharmacol. 1991, 40, 577–583. [Google Scholar] [PubMed]
- Swingle, M.; Honkanen, R.E. Small-molecule inhibitors of Ser/Thr protein phosphatases. In Methods in Molecular Biology; Moorhead, G., Ed.; Springer: New York, NY, USA, 2007; Volume 365, pp. 23–38. [Google Scholar]
- Honkanen, R.E.; Codispoti, B.A.; Tse, K.; Boynton, A.L. Characterization of natural toxins with inhibitory activity against serine/threonine protein phosphatases. Toxicon 1994, 32, 339–350. [Google Scholar] [CrossRef]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef]
- Toivola, D.M.; Eriksson, J.E.; Brautigan, D.L. Identification of protein phosphatase 2A as the primary target for microcystin-LR in rat liver homogenates. FEBS Lett. 1994, 344, 175–180. [Google Scholar] [CrossRef]
- Brewis, N.D.; Street, A.J.; Prescott, A.R.; Cohen, P.T. PPX, a novel protein serine/threonine phosphatase localized to centrosomes. EMBO J. 1993, 12, 987–996. [Google Scholar] [PubMed]
- Huang, X.; Honkanen, R.E. Molecular cloning, expression, and characterization of a novel human serine/threonine protein phosphatase, PP7, that is homologous to Drosophila retinal degeneration C gene product (rdgC). J. Biol. Chem. 1998, 273, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Serres, M. H.; Fladmark, K. E.; Døskeland, S. O. An ultrasensitive competitive binding assay for the detection of toxins affecting protein phosphatases. Toxicon 2000, 38, 347–360. [Google Scholar] [CrossRef]
- Nam, K.Y.; Hiro, M.; Kimura, S.; Fujiki, H.; Imanishi, Y. Permeability of a Non-Tpa-Type Tumor Promoter, Okadaic Acid, through Lipid Bilayer-Membrane. Carcinogenesis 1990, 11, 1171–1174. [Google Scholar] [CrossRef] [PubMed]
- Becsi, B.; Kiss, A.; Erdodi, F. Interaction of protein phosphatase inhibitors with membrane lipids assessed by surface plasmon resonance based binding technique. Chem. Phys. Lipids 2014, 183, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Oftedal, L.; Myhren, L.; Jokela, J.; Gausdal, G.; Sivonen, K.; Døskeland, S.O.; Herfindal, L. The lipopeptide toxins anabaenolysin A and B target biological membranes in a cholesterol-dependent manner. Biochim. Biophys. Acta 2012, 1818, 3000–3009. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Choudhuri, S.; Ogura, K.; Csanaky, I.L.; Lei, X.H.; Cheng, X.G.; Song, P.Z.; Klaassen, C.D. Characterization of organic anion transporting polypeptide 1b2-null mice: Essential role in hepatic uptake/toxicity of phalloidin and microcystin-LR. Toxicol. Sci. 2008, 103, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Herfindal, L.; Kasprzykowski, F.; Schwede, F.; Lankiewicz, L.; Fladmark, K.E.; Lukomska, J.; Wahlsten, M.; Sivonen, K.; Grzonka, Z.; Jastorff, B.; et al. Acyloxymethyl esterification of nodularin-R and microcystin-LA produces inactive protoxins that become reactivated and produce apoptosis inside intact cells. J. Med. Chem. 2009, 52, 5758–5762. [Google Scholar] [CrossRef] [PubMed]
- Bøe, R.; Gjertsen, B.T.; Vintermyr, O.K.; Houge, G.; Lanotte, M.; Døskeland, S.O. The protein phosphatase inhibitor okadaic acid induces morphological changes typical of apoptosis in mammalian cells. Exp. Cell Res. 1991, 195, 237–246. [Google Scholar] [CrossRef]
- Herfindal, L. Bioactive Compounds from Culturable Marine Microorganisms. In Proceedings of the 5th International Conference on Bioprospecting, Tromso, Norway, 23–25 February 2011; p. 32.
- Komatsu, M.; Furukawa, T.; Ikeda, R.; Takumi, S.; Nong, Q.; Aoyama, K.; Akiyama, S.; Keppler, D.; Takeuchi, T. Involvement of mitogen-activated protein kinase signaling pathways in microcystin-LR-induced apoptosis after its selective uptake mediated by OATP1B1 and OATP1B3. Toxicol. Sci. 2007, 97, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Monks, N.R.; Liu, S.; Xu, Y.; Yu, H.; Bendelow, A.S.; Moscow, J.A. Potent cytotoxicity of the phosphatase inhibitor microcystin LR and microcystin analogues in OATP1B1- and OATP1B3-expressing HeLa cells. Mol. Cancer Ther. 2007, 6, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Fladmark, K.E.; Serres, M.H.; Larsen, N.L.; Yasumoto, T.; Aune, T.; Døskeland, S.O. Sensitive detection of apoptogenic toxins in suspension cultures of rat and salmon hepatocytes. Toxicon 1998, 36, 1101–1114. [Google Scholar] [CrossRef]
- Liu, L.; Jokela, J.; Herfindal, L.; Wahlsten, M.; Sinkkonen, J.; Permi, P.; Fewer, D.P.; Døskeland, S.O.; Sivonen, K. 4-Methylproline guided natural product discovery: Co-occurrence of 4-hydroxy- and 4-methylprolines in nostoweipeptins and nostopeptolides. ACS Chem. Biol. 2014, 9, 2646–2655. [Google Scholar] [CrossRef] [PubMed]
- Jokela, J.; Herfindal, L.; Wahlsten, M.; Permi, P.; Selheim, F.; Vasconcelos, V.; Døskeland, S.O.; Sivonen, K. A novel cyanobacterial nostocyclopeptide is a potent antitoxin against microcystin. ChemBioChem 2010, 11, 1594–1599. [Google Scholar] [CrossRef] [PubMed]
- Fladmark, K.E.; Brustugun, O.T.; Hovland, R.; Boe, R.; Gjertsen, B.T.; Zhivotovsky, B.; Døskeland, S.O. Ultrarapid caspase-3 dependent apoptosis induction by serine/threonine phosphatase inhibitors. Cell Death Differ. 1999, 6, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Krakstad, C.; Herfindal, L.; Gjertsen, B.T.; Bøe, R.; Vintermyr, O.K.; Fladmark, K.E.; Døskeland, S.O. CaM-kinaseII dependent commitment to microcystin-induced apoptosis is coupled to cell budding, but not to shrinkage or chromatin hypercondensation. Cell Death Differ. 2006, 13, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Houge, G.; Robaye, B.; Eikhom, T.S.; Golstein, J.; Mellgren, G.; Gjertsen, B.T.; Lanotte, M.; Døskeland, S.O. Fine mapping of 28S rRNA sites specifically cleaved in cells undergoing apoptosis. Mol. Cell. Biol. 1995, 15, 2051–2062. [Google Scholar] [CrossRef] [PubMed]
- Samali, A.; Gilje, B.; Doskeland, S.O.; Cotter, T.G.; Houge, G. The ability to cleave 28S ribosomal RNA during apoptosis is a cell-type dependent trait unrelated to DNA fragmentation. Cell Death Differ. 1997, 4, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, T.; Linkermann, A.; Jouan-Lanhouet, S.; Walczak, H.; Vandenabeele, P. Regulated necrosis: The expanding network of non-apoptotic cell death pathways. Nat. Rev. Mol. Cell Biol. 2014, 15, 134–146. [Google Scholar]
- Gjertsen, B.T.; Cressey, L.I.; Ruchaud, S.; Houge, G.; Lanotte, M.; Doskeland, S.O. Multiple apoptotic death types triggered through activation of separate pathways by cAMP and inhibitors of protein phosphatases in one (IPC leukemia) cell line. J. Cell Sci. 1994, 107, 3363–3377. [Google Scholar] [PubMed]
- Pouria, S.; de Andrade, A.; Barbosa, J.; Cavalcanti, R.L.; Barreto, V.T.; Ward, C.J.; Preiser, W.; Poon, G.K.; Neild, G.H.; Codd, G.A. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.S.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.M.; Antunes, M.B.D.; de Melo, D.A.; Lyra, T.M.; Barreto, V.S.T.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. New Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.E.; Paatero, G.I.; Meriluoto, J.A.; Codd, G.A.; Kass, G.E.; Nicotera, P.; Orrenius, S. Rapid microfilament reorganization induced in isolated rat hepatocytes by microcystin-LR, a cyclic peptide toxin. Exp. Cell Res. 1989, 185, 86–100. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Toivola, D.; Meriluoto, J.A.; Karaki, H.; Han, Y.G.; Hartshorne, D. Hepatocyte deformation induced by cyanobacterial toxins reflects inhibition of protein phosphatases. Biochem. Biophys. Res. Commun. 1990, 173, 1347–1353. [Google Scholar] [CrossRef]
- Runnegar, M.T.; Falconer, I.R.; Silver, J. Deformation of isolated rat hepatocytes by a peptide hepatotoxin from the blue-green alga microcystis aeruginosa. Naunyn Schmiedebergs Arch. Pharmacol. 1981, 317, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Toivola, D.M.; Goldman, R.D.; Garrod, D.R.; Eriksson, J.E. Protein phosphatases maintain the organization and structural interactions of hepatic keratin intermediate filaments. J. Cell Sci. 1997, 110, 23–33. [Google Scholar] [PubMed]
- Hirano, K.; Chartier, L.; Taylor, R.G.; Allen, R.E.; Fusetani, N.; Karaki, H.; Hartshorne, D.J. Changes in the cytoskeleton of 3T3 fibroblasts induced by the phosphatase inhibitor, calyculin-A. J. Muscle Res. Cell Motil. 1992, 13, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Huseby, S.; Gausdal, G.; Keen, T.J.; Kjaerland, E.; Krakstad, C.; Myhren, L.; Bronstad, K.; Kunick, C.; Schwede, F.; Genieser, H.G.; et al. Cyclic AMP induces IPC leukemia cell apoptosis via CRE-and CDK-dependent Bim transcription. Cell Death Dis. 2011, 2, e237. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.H.; Kim, D.H.; Choi, J.E.; Chang, E.J.; Yoon, S.Y. Increased phosphorylation of dynamin-related protein 1 and mitochondrial fission in okadaic acid-treated neurons. Brain Res. 2012, 1454, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Suguri, H.; Yoshizawa, S.; Takagi, K.; Uda, N.; Wakamatsu, K.; Yamada, K.; Murata, M.; Yasumoto, T.; et al. Diarrhetic Shellfish Toxin, Dinophysistoxin-1, Is a Potent Tumor Promoter on Mouse Skin. Jpn. J. Cancer Res. 1988, 79, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver tumor promotion by the cyanobacterial cyclic peptide toxin microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Pallas, D.C.; Shahrik, L.K.; Martin, B.L.; Jaspers, S.; Miller, T.B.; Brautigan, D.L.; Roberts, T.M. Polyoma Small and Middle T-Antigens and Sv40 Small T-Antigen Form Stable Complexes with Protein Phosphatase-2A. Cell 1990, 60, 167–176. [Google Scholar] [CrossRef]
- Yang, S.I.; Lickteig, R.L.; Estes, R.; Rundell, K.; Walter, G.; Mumby, M.C. Control of Protein Phosphatase-2A by Simian Virus-40 Small-T Antigen. Mol. Cell. Biol. 1991, 11, 1988–1995. [Google Scholar] [CrossRef] [PubMed]
- Harmala-Brasken, A.S.; Mikhailov, A.; Soderstrom, T.S.; Meinander, A.; Holmstrom, T.H.; Damuni, Z.; Eriksson, J.E. Type-2A protein phosphatase activity is required to maintain death receptor responsiveness. Oncogene 2003, 22, 7677–7686. [Google Scholar] [CrossRef] [PubMed]
- Mellgren, G.; Vintermyr, O.K.; Boe, R.; Døskeland, S.O. Hepatocyte DNA replication is abolished by inhibitors selecting protein phosphatase 2A rather than phosphatase 1. Exp. Cell Res. 1993, 205, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Dean, N.M.; Mordan, L.J.; Tse, K.; Mooberry, S.L.; Boynton, A.L. Okadaic Acid Inhibits PDGF-Induced Proliferation and Decreases PDGF Receptor Number in C3H/10T1/2 Mouse Fibroblasts. Carcinogenesis 1991, 12, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, M.M. Microcystin-LR and okadaic acid-induced cellular effects: A dualistic response. FEBS Lett. 2004, 557, 1–8. [Google Scholar] [CrossRef]
- Nakagama, H.; Kaneko, S.; Shima, H.; Inamori, H.; Fukuda, H.; Kominami, R.; Sugimura, T.; Nagao, M. Induction of minisatellite mutation in NIH 3T3 cells by treatment with the tumor promoter okadaic acid. Proc. Natl. Acad. Sci. USA 1997, 94, 10813–10816. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.S.; Catian, R.; Moukha, S.; Matias, W.G.; Creppy, E.E. Comparative study of Domoic Acid and Okadaic Acid induced-chromosomal abnormalities in the Caco-2 cell line. Int. J. Environ. Res. Public Health 2006, 3, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Fladmark, K.E.; Brustugun, O.T.; Mellgren, G.; Krakstad, C.; Boe, R.; Vintermyr, O.K.; Schulman, H.; Døskeland, S.O. Ca2+/calmodulin-dependent protein kinase II is required for microcystin-induced apoptosis. J. Biol. Chem. 2002, 277, 2804–2811. [Google Scholar] [CrossRef] [PubMed]
- Coultrap, S.J.; Bayer, K.U. CaMKII regulation in information processing and storage. Trends Neurosci. 2012, 35, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.R.; Joiner, M.L.A.; Guan, X.; Kutschke, W.; Yang, J.Y.; Oddis, C.V.; Bartlett, R.K.; Lowe, J.S.; O’Donnell, S.E.; Aykin-Burns, N.; et al. A dynamic pathway for calcium-independent activation of CaMKII by methionine oxidation. Cell 2008, 133, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Denu, J.M.; Tanner, K.G. Specific and reversible inactivation of protein tyrosine phosphatases by hydrogen peroxide: Evidence for a sulfenic acid intermediate and implications for redox regulation. Biochemistry 1998, 37, 5633–5642. [Google Scholar] [CrossRef]
- Low, I.C.; Loh, T.; Huang, Y.; Virshup, D.M.; Pervaiz, S. Ser70 phosphorylation of Bcl-2 by selective tyrosine nitration of PP2A-B56delta stabilizes its antiapoptotic activity. Blood 2014, 124, 2223–2234. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Yin, J.; Luo, Y.; Huang, S. Hydrogen peroxide-induced neuronal apoptosis is associated with inhibition of protein phosphatase 2A and 5, leading to activation of MAPK pathway. Int. J. Biochem. Cell Biol. 2009, 41, 1284–1295. [Google Scholar] [CrossRef] [PubMed]
- Drose, S.; Brandt, U. Molecular mechanisms of superoxide production by the mitochondrial respiratory chain. Adv. Exp. Med. Biol. 2012, 748, 145–169. [Google Scholar] [PubMed]
- Lustgarten, M.S.; Bhattacharya, A.; Muller, F.L.; Jang, Y.C.; Shimizu, T.; Shirasawa, T.; Richardson, A.; van Remmen, H. Complex I generated, mitochondrial matrix-directed superoxide is released from the mitochondria through voltage dependent anion channels. Biochem. Biophys. Res. Commun. 2012, 422, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Fujino, G.; Noguchi, T.; Takeda, K.; Ichijo, H. Thioredoxin and protein kinases in redox signaling. Semin. Cancer Biol. 2006, 16, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, M.; Nishitoh, H.; Fujii, M.; Takeda, K.; Tobiume, K.; Sawada, Y.; Kawabata, M.; Miyazono, K.; Ichijo, H. Mammalian thioredoxin is a direct inhibitor of apoptosis signal-regulating kinase (ASK) 1. EMBO J. 1998, 17, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhao, J.N.; Chang, Z.Y.; Guo, G.D. CaMKII Activates ASK1 to Induce Apoptosis of Spinal Astrocytes under Oxygen-Glucose Deprivation. Cell. Mol. Neurobiol. 2013, 33, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Kashiwase, K.; Higuchi, Y.; Hirotani, S.; Yamaguchi, O.; Hikoso, S.; Takeda, T.; Watanabe, T.; Taniike, M.; Nakai, A.; Tsujimoto, I.; et al. CaMKII activates ASK1 and NF-kappa B to induce cardiomyocyte hypertrophy. Biochem. Biophys. Res. Commun. 2005, 327, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Lorincz, A.M.; Szarvas, G.; Smith, S.M.; Ligeti, E. Role of Rac GTPase activating proteins in regulation of NADPH oxidase in human neutrophils. Free Radic. Biol. Med. 2014, 68, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbeck, M.J.; Hegg, G.G.; Karnovsky, M.J. Arachidonate Activation of the Neutrophil NADPH-Oxidase—Synergistic Effects of Protein Phosphatase Inhibitors Compared with Protein-Kinase Activators. J. Biol. Chem. 1991, 266, 16336–16342. [Google Scholar] [PubMed]
- Farquharson, C.A.; Butler, R.; Hill, A.; Belch, J.J.; Struthers, A.D. Allopurinol improves endothelial dysfunction in chronic heart failure. Circulation 2002, 106, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Kaludercic, N.; Mialet-Perez, J.; Paolocci, N.; Parini, A.; Di Lisa, F. Monoamine oxidases as sources of oxidants in the heart. J. Mol. Cell. Cardiol. 2014, 73, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.D.; Kaufman, S. Products of the Tyrosine-Dependent Oxidation of Tetrahydrobiopterin by Rat-Liver Phenylalanine-Hydroxylase. Arch. Biochem. Biophys. 1993, 304, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Haavik, J.; Almas, B.; Flatmark, T. Generation of reactive oxygen species by tyrosine hydroxylase: A possible contribution to the degeneration of dopaminergic neurons? J. Neurochem. 1997, 68, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.D.; Klaidman, L.K.; Ribeiro, P. Tyrosine hydroxylase: Mechanisms of oxygen radical formation. Redox. Rep. 1997, 3, 273–279. [Google Scholar] [PubMed]
- Doskeland, A.P.; Vintermyr, O.K.; Flatmark, T.; Cotton, R.G.H.; Doskeland, S.O. Phenylalanine Positively Modulates the cAMP-Dependent Phosphorylation and Negatively Modulates the Vasopressin-Induced and Okadaic-Acid-Induced Phosphorylation of Phenylalanine 4-Monooxygenase in Intact Rat Hepatocytes. Eur. J. Biochem. 1992, 206, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Lee, G.H.; Cho, E.Y.; Chae, S.W.; Ahn, T.; Chae, H.J. Bax inhibitor 1 regulates ER-stress-induced ROS accumulation through the regulation of cytochrome P450 2E1. J. Cell Sci. 2009, 122, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Laursen, J.B.; Somers, M.; Kurz, S.; McCann, L.; Warnholtz, A.; Freeman, B.A.; Tarpey, M.; Fukai, T.; Harrison, D.G. Endothelial regulation of vasomotion in apoE-deficient mice: Implications for interactions between peroxynitrite and tetrahydrobiopterin. Circulation 2001, 103, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Guidarelli, A.; Cantoni, O. Pivotal role of superoxides generated in the mitochondrial respiratory chain in peroxynitrite-dependent activation of phospholipase A2. Biochem. J. 2002, 366, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.; Sanchez, A.; Pilar Martinez, M.; Benedito, S.; Lopez-Oliva, M.E.; Garcia-Sacristan, A.; Hernandez, M.; Prieto, D. COX-2 is involved in vascular oxidative stress and endothelial dysfunction of renal interlobar arteries from obese Zucker rats. Free Radic. Biol. Med. 2015, 84, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Fladmark, K.E.; Brustugun, O.T.; Mellgren, G.; Krakstad, C.; Boe, R.; Vintermyr, O.K.; Schulman, H.; Doskeland, S.O. Ca2+/calmodulin-dependent protein kinase II is required for microcystin-induced apoptosis. J. Biol. Chem. 2002, 277, 2804–2811. [Google Scholar] [CrossRef] [PubMed]
- Im, J.Y.; Kim, D.; Paik, S.G.; Han, P.L. Cyclooxygenase-2-dependent neuronal death proceeds via superoxide anion generation. Free Radic. Biol. Med. 2006, 41, 960–972. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.H.; Liou, H.H.; Hour, M.J.; Liou, H.C.; Fu, W.M. Protection of dopaminergic neurons by 5-lipoxygenase inhibitor. Neuropharmacology 2013, 73, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.S.; Dho, S.H.; Youm, T.H.; Kwon, K.S.; Park, S.S. Astrocytic phospholipase A2 contributes to neuronal glutamate toxicity. Brain Res. 2014, 1590, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Christofi, F.L.; Wright, V.P.; Bao, S.; Clanton, T.L. Lipoxygenase-dependent superoxide release in skeletal muscle. J. Appl. Physiol. (1985) 2004, 97, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Kanno, T.; Tsuchiya, A.; Shimizu, T.; Tanaka, A.; Nishizaki, T. Indomethacin Serves as a Potential Inhibitor of Protein Phosphatases. Cell Physiol. Biochem. 2012, 30, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Ozveren, E.; Korkmaz, B.; Buharalioglu, C.K.; Tunctan, B. Involvement of calcium/calmodulin-dependent protein kinase II to endotoxin-induced vascular hyporeactivity in rat superior mesenteric artery. Pharmacol. Res. 2006, 54, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Fatima, S.; Yaghini, F.A.; Ahmed, A.; Khandekar, Z.; Malik, K.U. CaM kinase II alpha mediates norepinephrine-induced translocation of cytosolic phospholipase A(2) to the nuclear envelope. J. Cell Sci. 2003, 116, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Grewal, S.; Ponnambalam, S.; Walker, J.H. Association of cPLA(2)-alpha and COX-1 with the Golgi apparatus of A549 human lung epithelial cells. J. Cell Sci. 2003, 116, 2303–2310. [Google Scholar] [CrossRef] [PubMed]
- Strid, T.; Svartz, J.; Franck, N.; Hallin, E.; Ingelsson, B.; Soderstrom, M.; Hammarstrom, S. Distinct parts of leukotriene C-4 synthase interact with 5-lipoxygenase and 5-lipoxygenase activating protein. Biochem. Biophys. Res. Commun. 2009, 381, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Radmark, O.; Werz, O.; Steinhilber, D.; Samuelsson, B. 5-Lipoxygenase: Regulation of expression and enzyme activity. Trends Biochem. Sci. 2007, 32, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Fredman, G.; Ozcan, L.; Spolitu, S.; Hellmann, J.; Spite, M.; Backs, J.; Tabas, I. Resolvin D1 limits 5-lipoxygenase nuclear localization and leukotriene B-4 synthesis by inhibiting a calcium-activated kinase pathway. Proc. Natl. Acad. Sci. USA 2014, 111, 14530–14535. [Google Scholar] [CrossRef] [PubMed]
- Sandal, T.; Aumo, L.; Hedin, L.; Gjertsen, B.T.; Døskeland, S.O. Irod/Ian5: An inhibitor of gamma-radiation- and okadaic acid-induced apoptosis. Mol. Biol. Cell 2003, 14, 3292–3304. [Google Scholar] [CrossRef] [PubMed]
- Sandal, T.; Ahlgren, R.; Lillehaug, J.; Doskeland, S.O. Establishment of okadaic acid resistant cell clones using a cDNA expression library. Cell Death Differ. 2001, 8, 754–766. [Google Scholar] [CrossRef] [PubMed]
- Schulteis, R.D.; Chu, H.Y.; Dai, X.Z.; Chen, Y.H.; Edwards, B.; Haribhai, D.; Williams, C.B.; Malarkannan, S.; Hessner, M.J.; Glisic-Milosavljevic, S.; et al. Impaired survival of peripheral T cells, disrupted NK/NKT cell development, and liver failure in mice lacking Gimap5. Blood 2008, 112, 4905–4914. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleppe, R.; Herfindal, L.; Døskeland, S.O. Cell Death Inducing Microbial Protein Phosphatase Inhibitors—Mechanisms of Action. Mar. Drugs 2015, 13, 6505-6520. https://doi.org/10.3390/md13106505
Kleppe R, Herfindal L, Døskeland SO. Cell Death Inducing Microbial Protein Phosphatase Inhibitors—Mechanisms of Action. Marine Drugs. 2015; 13(10):6505-6520. https://doi.org/10.3390/md13106505
Chicago/Turabian StyleKleppe, Rune, Lars Herfindal, and Stein Ove Døskeland. 2015. "Cell Death Inducing Microbial Protein Phosphatase Inhibitors—Mechanisms of Action" Marine Drugs 13, no. 10: 6505-6520. https://doi.org/10.3390/md13106505
APA StyleKleppe, R., Herfindal, L., & Døskeland, S. O. (2015). Cell Death Inducing Microbial Protein Phosphatase Inhibitors—Mechanisms of Action. Marine Drugs, 13(10), 6505-6520. https://doi.org/10.3390/md13106505