Deferoxamine Compensates for Decreases in B Cell Counts and Reduces Mortality in Enterovirus 71-Infected Mice

Abstract
:
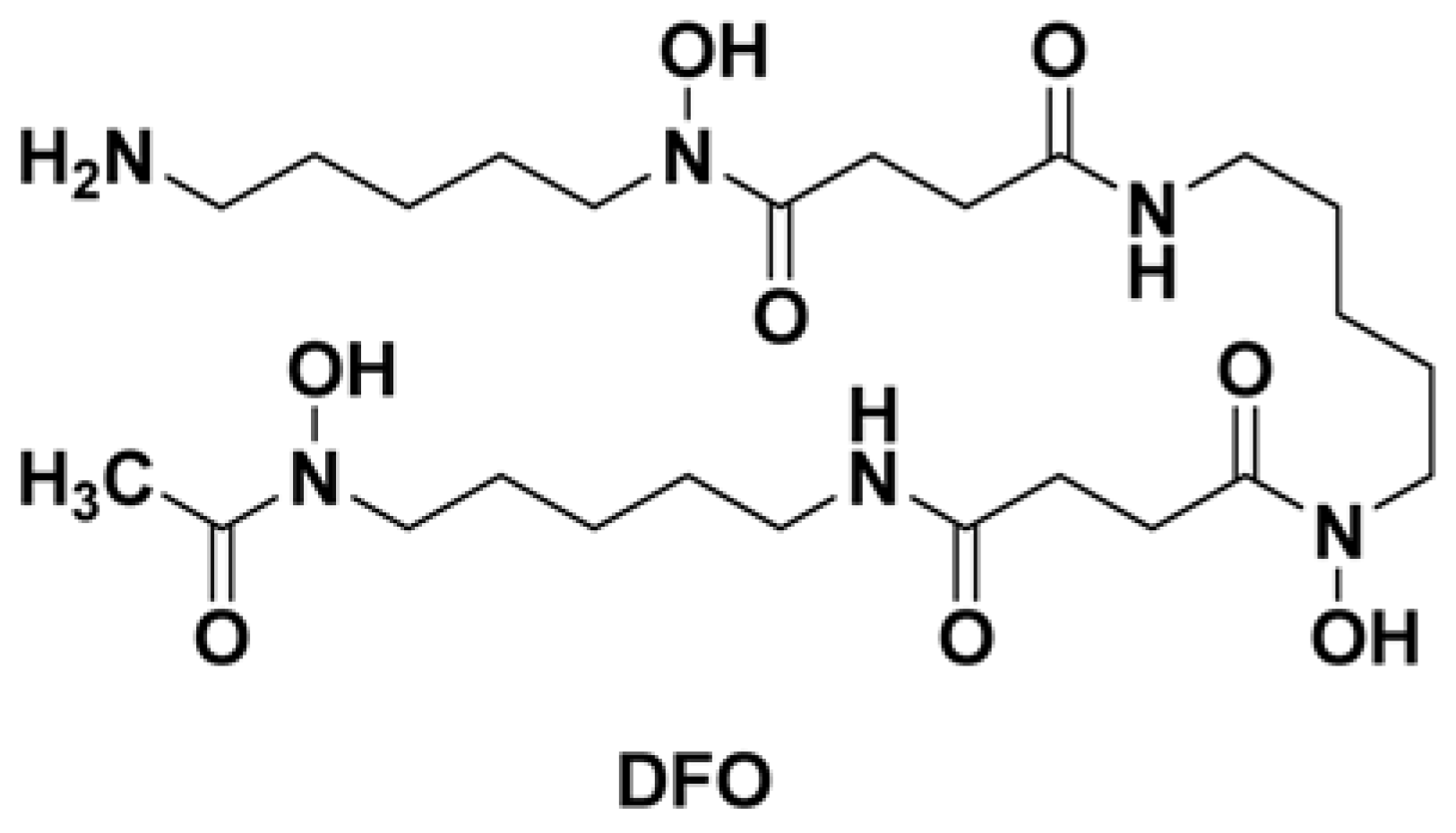
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
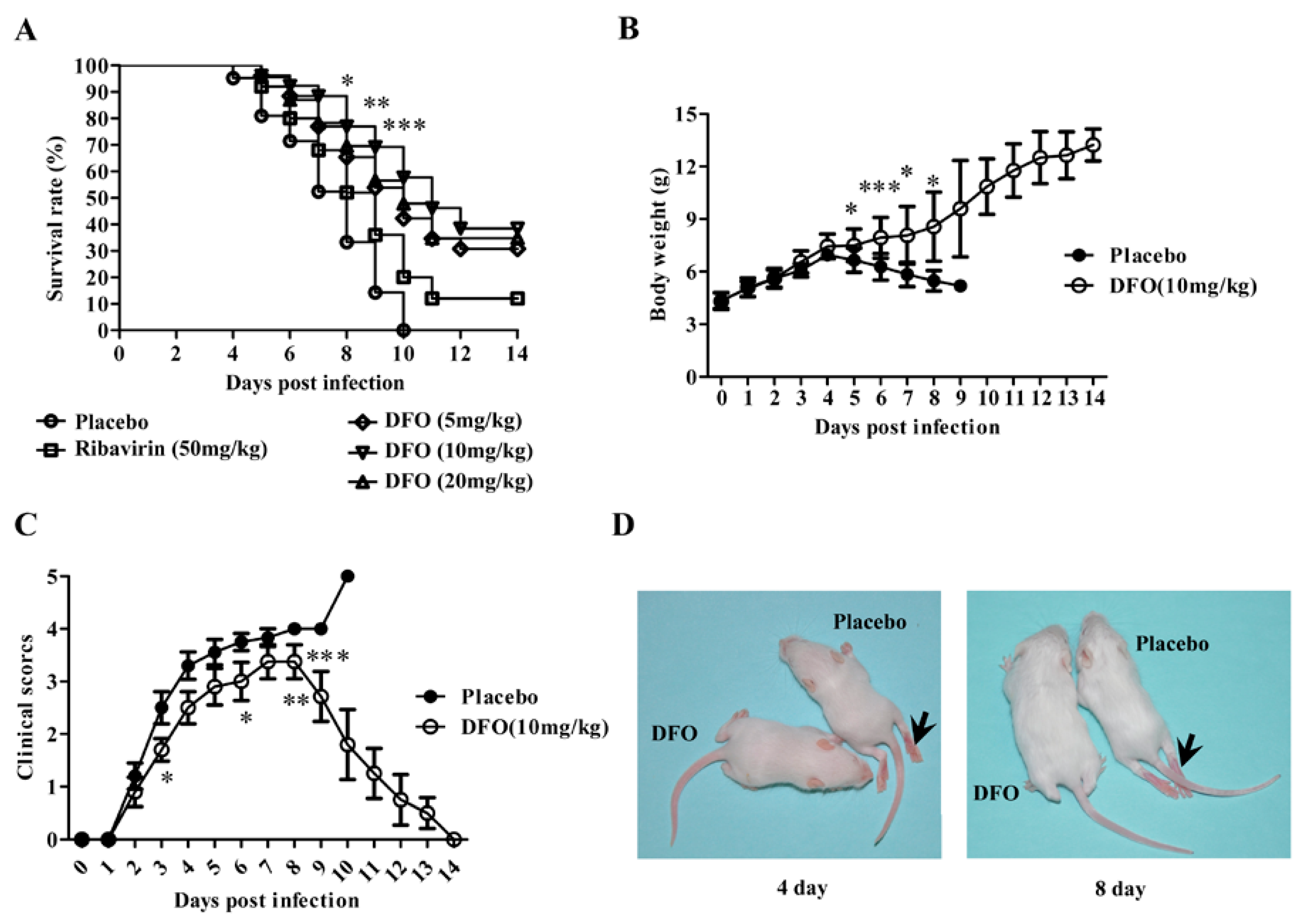
2.1. DFO Treatment Reduced Mouse Mortality upon Lethal EV71 Challenge
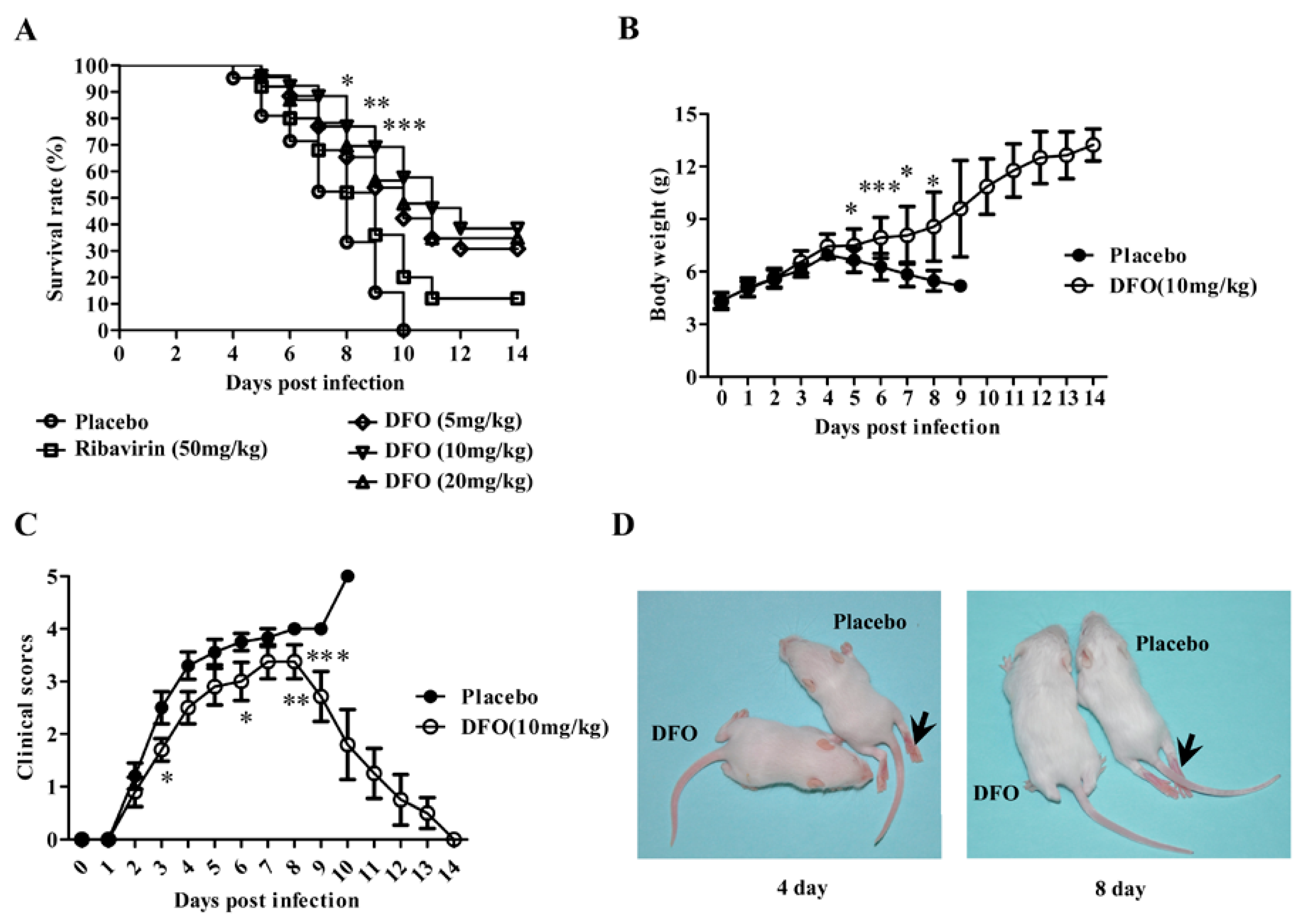
2.2. DFO Treatment Relieved the Symptoms of the Infected Mice
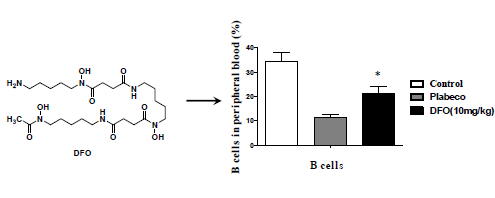
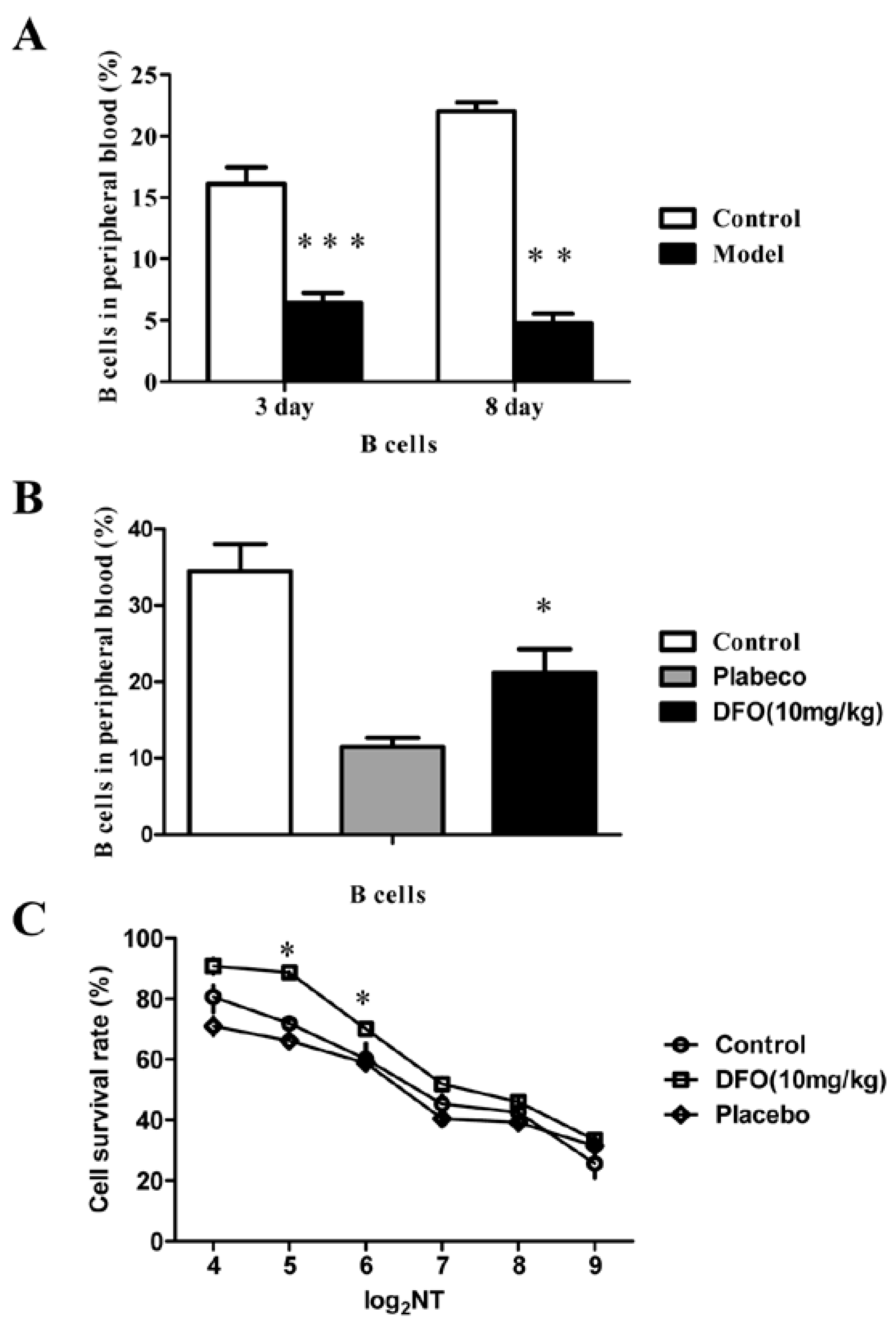
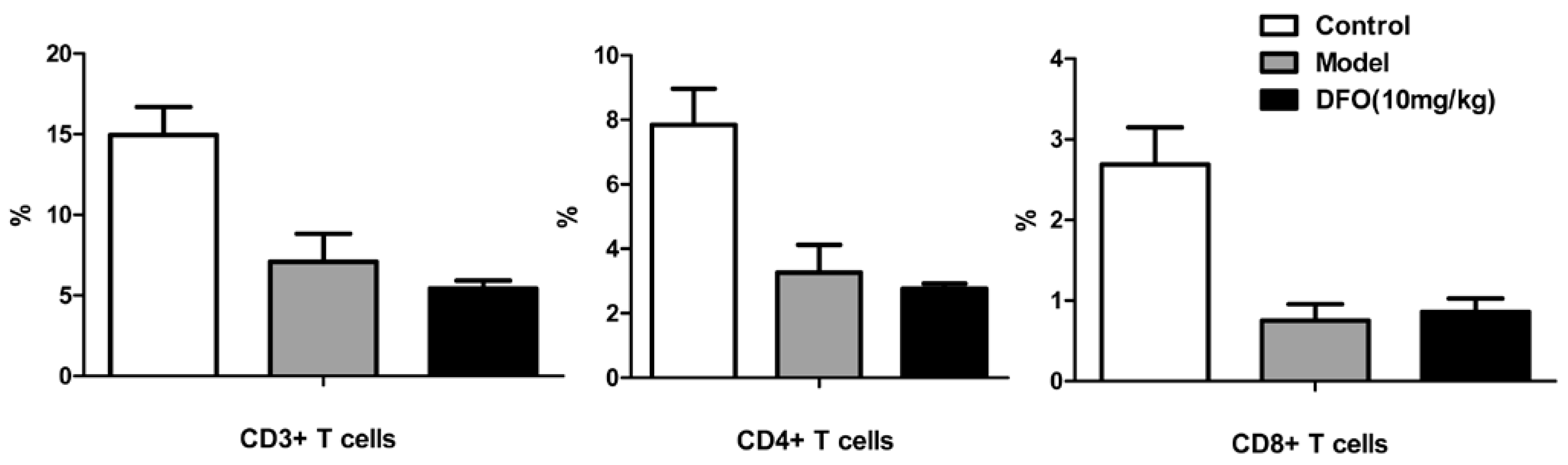
2.3. DFO Treatment Regulates the B Cell Levels of the Infected Mice
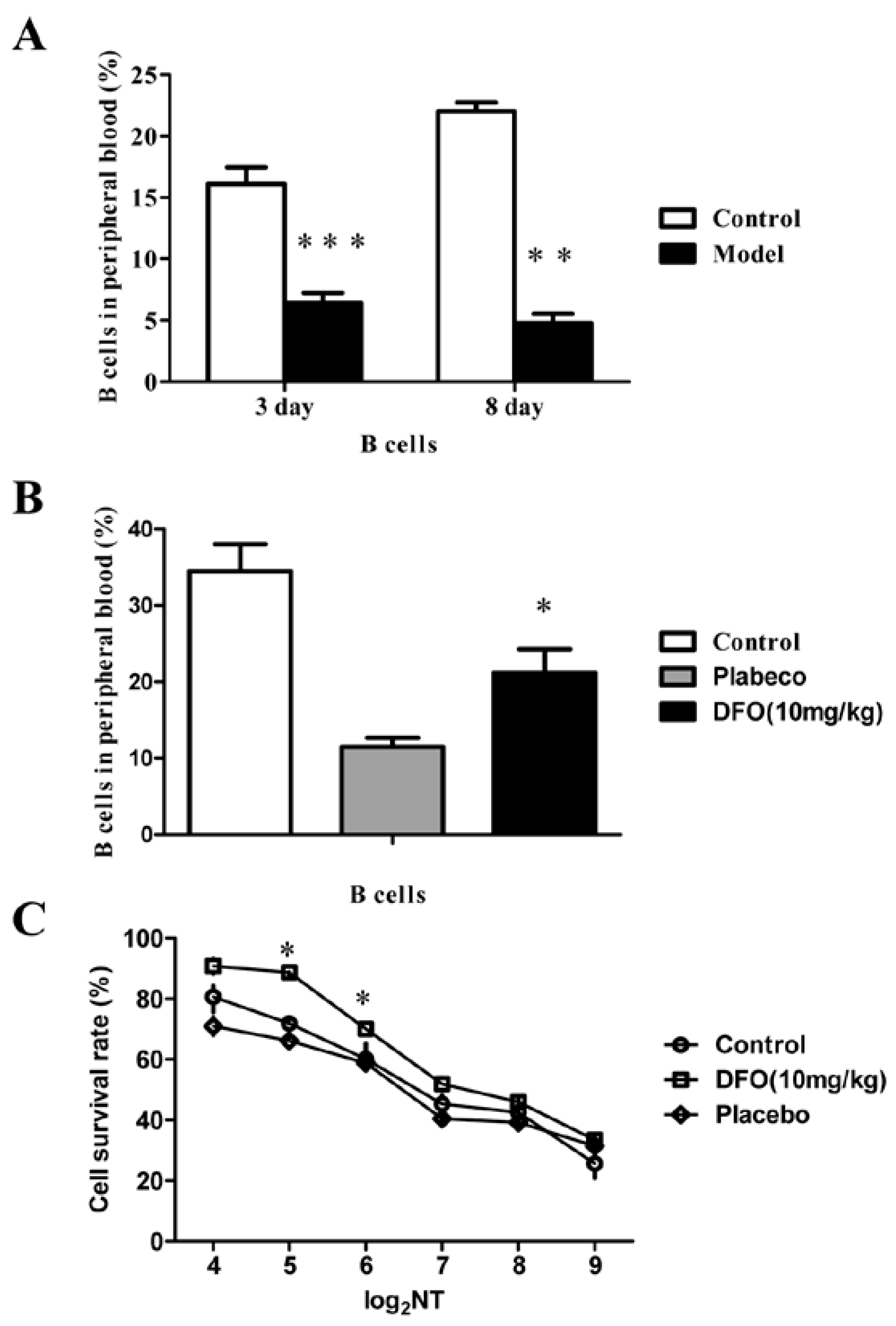
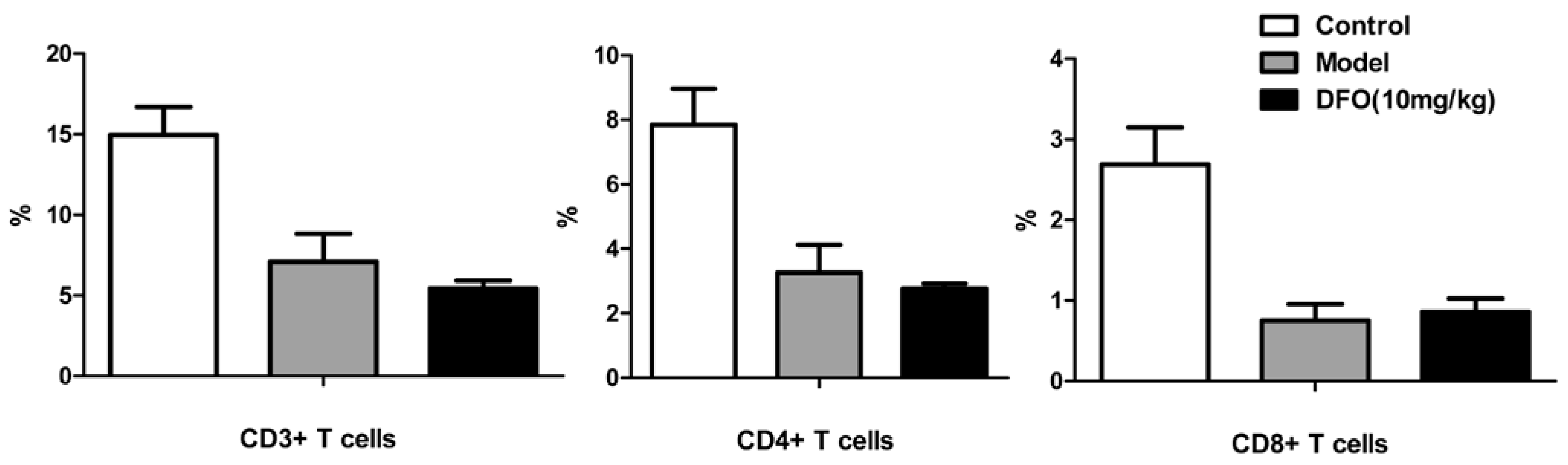
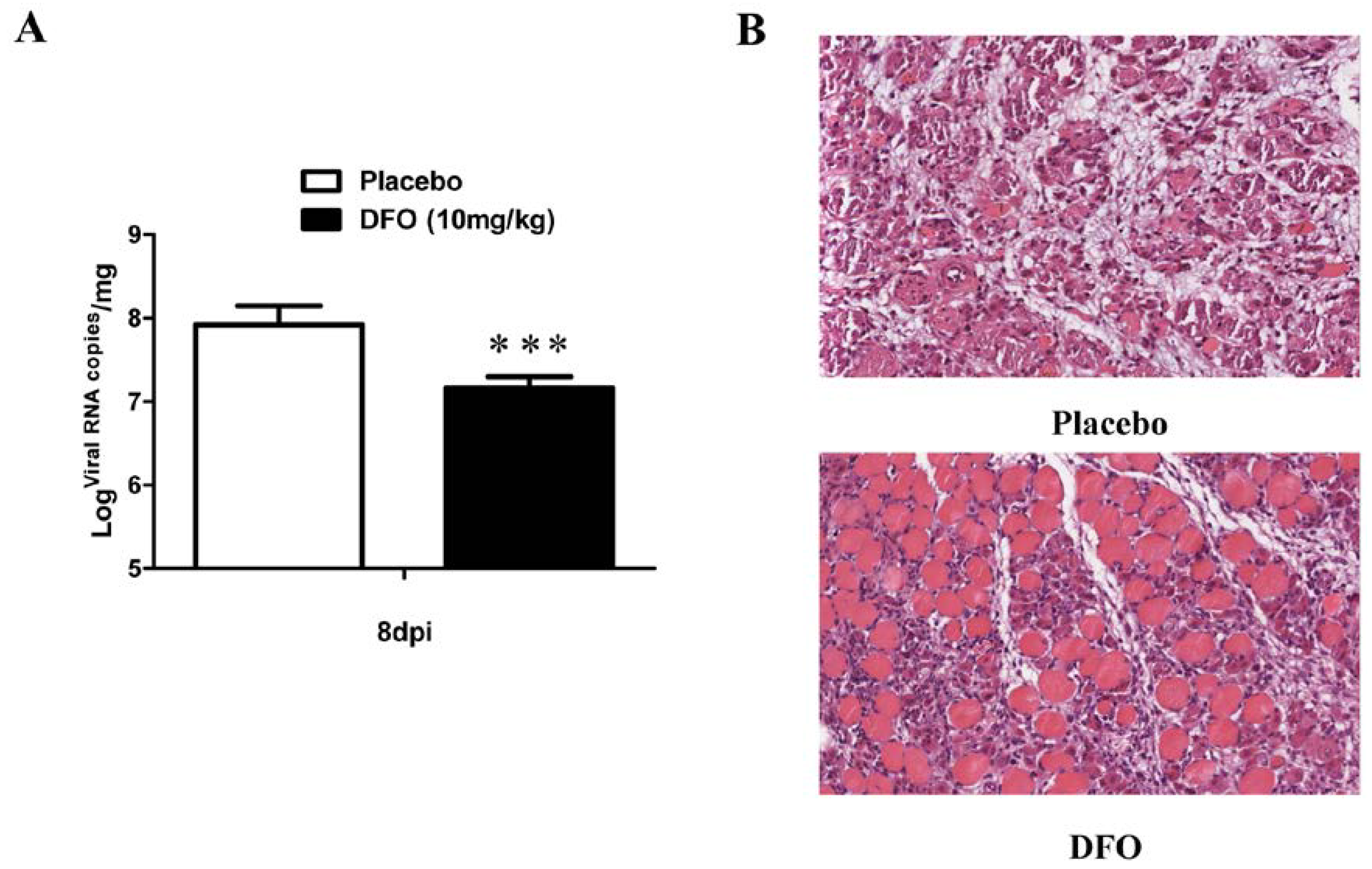
2.4. DFO Treatment Slightly Reduced the Viral Load of the Infected Mice
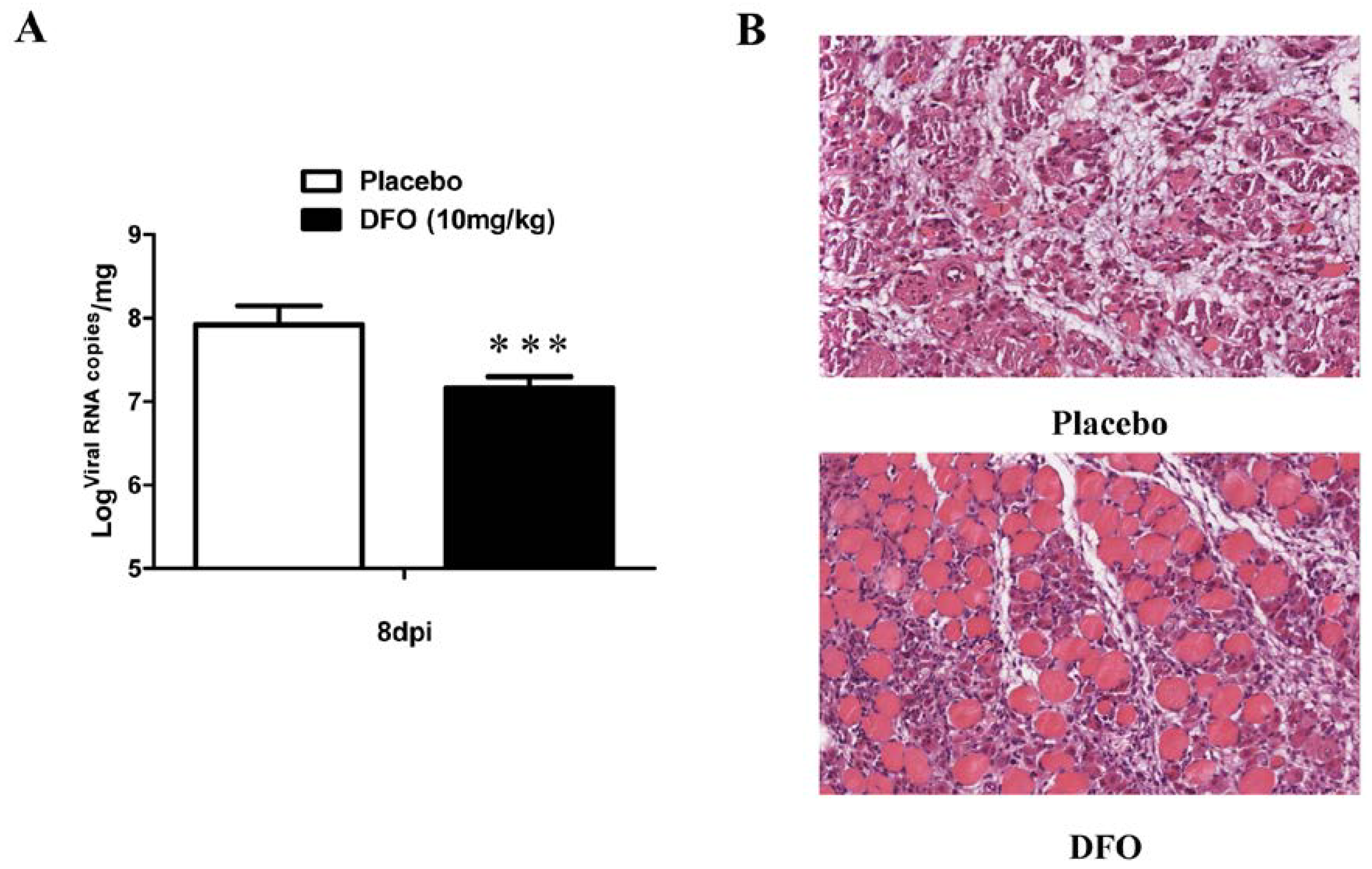
2.5. DFO Treatment Reduced the Muscle Damage of the Infected Mice
3. Experimental Section
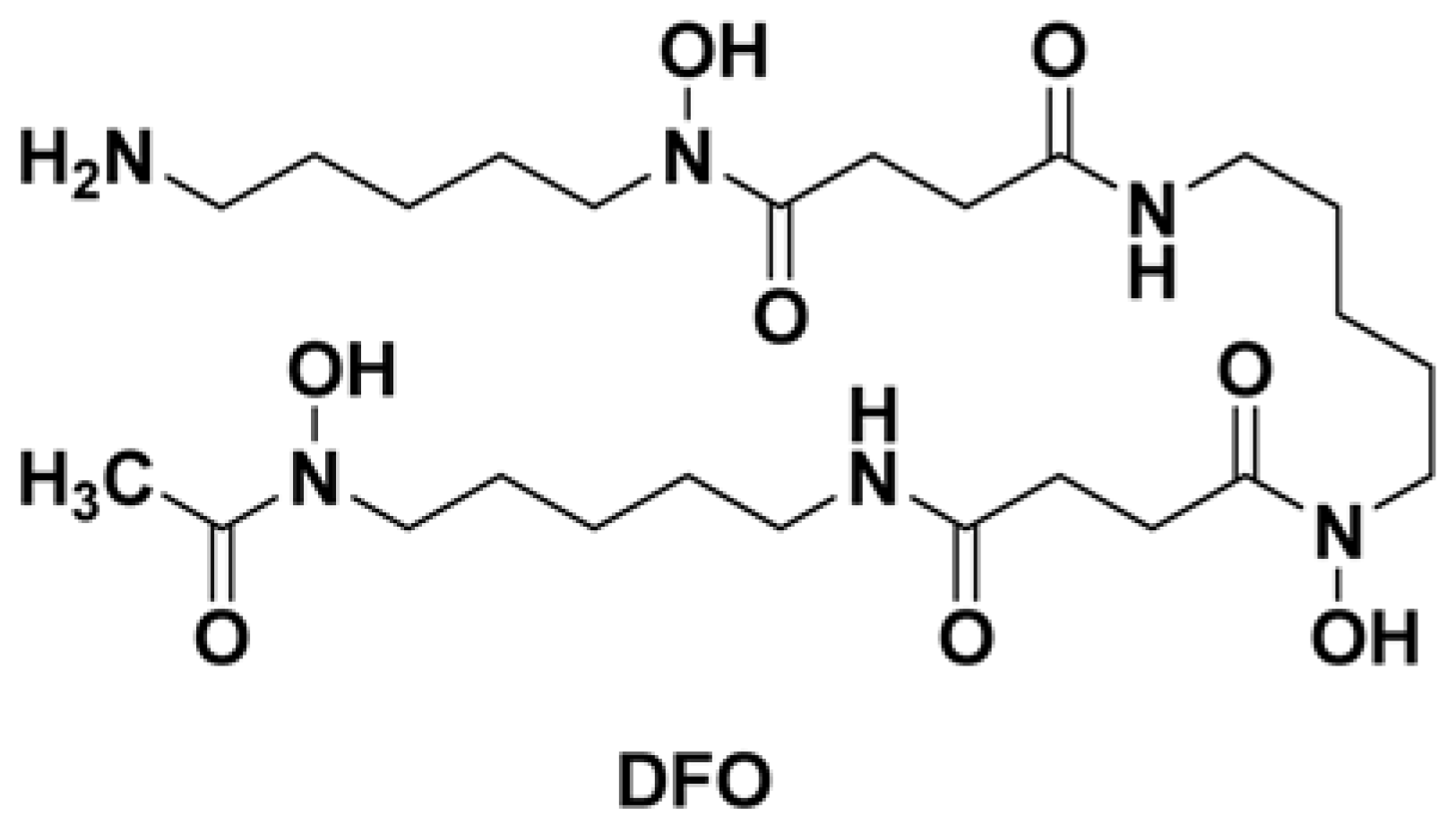
3.1. Cells, Viruses and Reagents
3.3. Mouse Protection Assay
3.4. Flow Cytometry
3.5. Neutralization Assay
3.6. Determination of the Viral Load
3.7. Pathology
3.8. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wu, K.X.; Ng, M.M.; Chu, J.J. Developments towards antiviral therapies against enterovirus 71. Drug Discov. Today 2010, 15, 1041–1051. [Google Scholar]
- Chang, C.W.; Leu, Y.L.; Horng, J.T. Daphne Genkwa sieb. et Zucc. Water-soluble extracts act on enterovirus 71 by inhibiting viral entry. Viruses 2012, 4, 539–556. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, T.; Du, J.; Cui, S.; Yang, F.; Jin, Q. Anti-enterovirus 71 effects of chrysin and its phosphate ester. PLoS One 2014, 9, e89668. [Google Scholar]
- Yang, F.; Zhang, T.; Hu, Y.; Wang, X.; Du, J.; Li, Y.; Sun, S.; Sun, X.; Li, Z.; Jin, Q. Survey of enterovirus infections from hand, foot and mouth disease outbreak in China, 2009. Virol. J. 2011, 8, 508. [Google Scholar] [CrossRef]
- Lin, Y.J.; Lai, C.C.; Lai, C.H.; Sue, S.C.; Lin, C.W.; Hung, C.H.; Lin, T.H.; Hsu, W.Y.; Huang, S.M.; Hung, Y.L.; et al. Inhibition of enterovirus 71 infections and viral IRES activity by Fructus gardeniae and geniposide. Eur. J. Med. Chem. 2013, 62, 206–213. [Google Scholar] [CrossRef]
- Wang, L.C.; Kao, C.M.; Ling, P.; Su, I.J.; Chang, T.M.; Chen, S.H. CD4 T-cell-independent antibody response reduces enterovirus 71 lethality in mice by decreasing tissue viral loads. Clin. Dev. Immunol. 2012, 2012. [Google Scholar] [CrossRef]
- Lin, Y.W.; Chang, K.C.; Kao, C.M.; Chang, S.P.; Tung, Y.Y.; Chen, S.H. Lymphocyte and antibody responses reduce enterovirus 71 lethality in mice by decreasing tissue viral loads. J. Virol. 2009, 83, 6477–6483. [Google Scholar] [CrossRef]
- Abzug, M.J.; Keyserling, H.L.; Lee, M.L.; Levin, M.J.; Rotbart, H.A. Neonatal enterovirus infection: Virology, serology, and effects of intravenous immune globulin. Clin. Infect. Dis. 1995, 20, 1201–1206. [Google Scholar] [CrossRef]
- Cao, R.Y.; Dong, D.Y.; Liu, R.J.; Han, J.F.; Wang, G.C.; Zhao, H.; Li, X.F.; Deng, Y.Q.; Zhu, S.Y.; Wang, X.Y.; et al. Human IgG subclasses against enterovirus Type 71: Neutralization versus antibody dependent enhancement of infection. PLoS One 2013, 8, e64024. [Google Scholar]
- Xiong, Z.Q.; Wang, J.F.; Hao, Y.Y.; Wang, Y. Recent advances in the discovery and development of marine microbial natural products. Mar. Drugs 2013, 11, 700–717. [Google Scholar] [CrossRef]
- Meyer, D. Iron chelation as therapy for HIV and Mycobacterium tuberculosis co-infection under conditions of iron overload. Curr. Pharm. Des. 2006, 12, 1943–1947. [Google Scholar] [CrossRef]
- Williams, A.; Meyer, D. Desferrioxamine as immunomodulatory agent during microorganism infection. Curr. Pharm. Des. 2009, 15, 1261–1268. [Google Scholar] [CrossRef]
- Yang, Y.J.; Zhang, L.F.; Fan, X.X.; Qin, C.; Liu, J.N. Antiviral effect of geraniin on human enterovirus 71 in vitro and in vivo. Bioorg. Med. Chem. Lett. 2012, 22, 2209–2211. [Google Scholar] [CrossRef]
- Yang, Y.J.; Xiu, J.H.; Zhang, X.; Zhang, L.; Yan, K.; Qin, C.; Liu, J.N. Antiviral effect of matrine against human enterovirus 71. Molecules 2012, 17, 10370–10376. [Google Scholar] [CrossRef]
- Liu, J.N.; Wang, W.; Duo, J.Y.; Hao, Y.; Ma, C.M.; Li, W.B.; Lin, S.Z.; Gao, X.Z.; Liu, X.L.; Xu, Y.F.; et al. Combined peptides of human enterovirus 71 protect against virus infection in mice. Vaccine 2010, 8, 7444–7451. [Google Scholar]
- Yang, Y.J.; Xiu, J.H.; Liu, J.N.; Zhang, L.; Li, X.Y.; Xu, Y.F.; Qin, C.; Zhang, L.F. Chebulagic Acid, a Hydrolyzable Tannin, Exhibited Antiviral Activity in Vitro and in Vivo against Human Enterovirus 71. Int. J. Mol. Sci. 2013, 14, 9618–9627. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, Y.; Ma, J.; Xiu, J.; Bai, L.; Guan, F.; Zhang, L.; Liu, J.; Zhang, L. Deferoxamine Compensates for Decreases in B Cell Counts and Reduces Mortality in Enterovirus 71-Infected Mice. Mar. Drugs 2014, 12, 4086-4095. https://doi.org/10.3390/md12074086
Yang Y, Ma J, Xiu J, Bai L, Guan F, Zhang L, Liu J, Zhang L. Deferoxamine Compensates for Decreases in B Cell Counts and Reduces Mortality in Enterovirus 71-Infected Mice. Marine Drugs. 2014; 12(7):4086-4095. https://doi.org/10.3390/md12074086
Chicago/Turabian StyleYang, Yajun, Jing Ma, Jinghui Xiu, Lin Bai, Feifei Guan, Li Zhang, Jiangning Liu, and Lianfeng Zhang. 2014. "Deferoxamine Compensates for Decreases in B Cell Counts and Reduces Mortality in Enterovirus 71-Infected Mice" Marine Drugs 12, no. 7: 4086-4095. https://doi.org/10.3390/md12074086
APA StyleYang, Y., Ma, J., Xiu, J., Bai, L., Guan, F., Zhang, L., Liu, J., & Zhang, L. (2014). Deferoxamine Compensates for Decreases in B Cell Counts and Reduces Mortality in Enterovirus 71-Infected Mice. Marine Drugs, 12(7), 4086-4095. https://doi.org/10.3390/md12074086