Impact of DHA on Metabolic Diseases from Womb to Tomb


Abstract
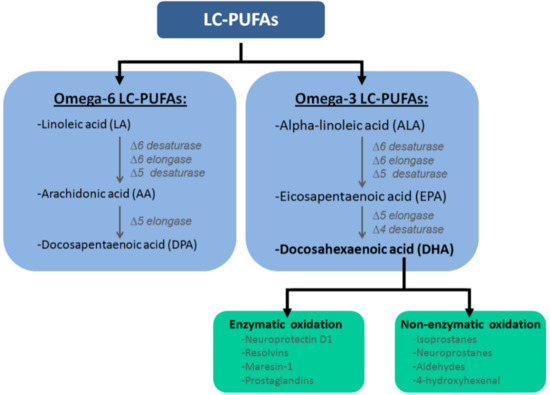
:
1. Introduction
Docosahexaenoic Acid (DHA)
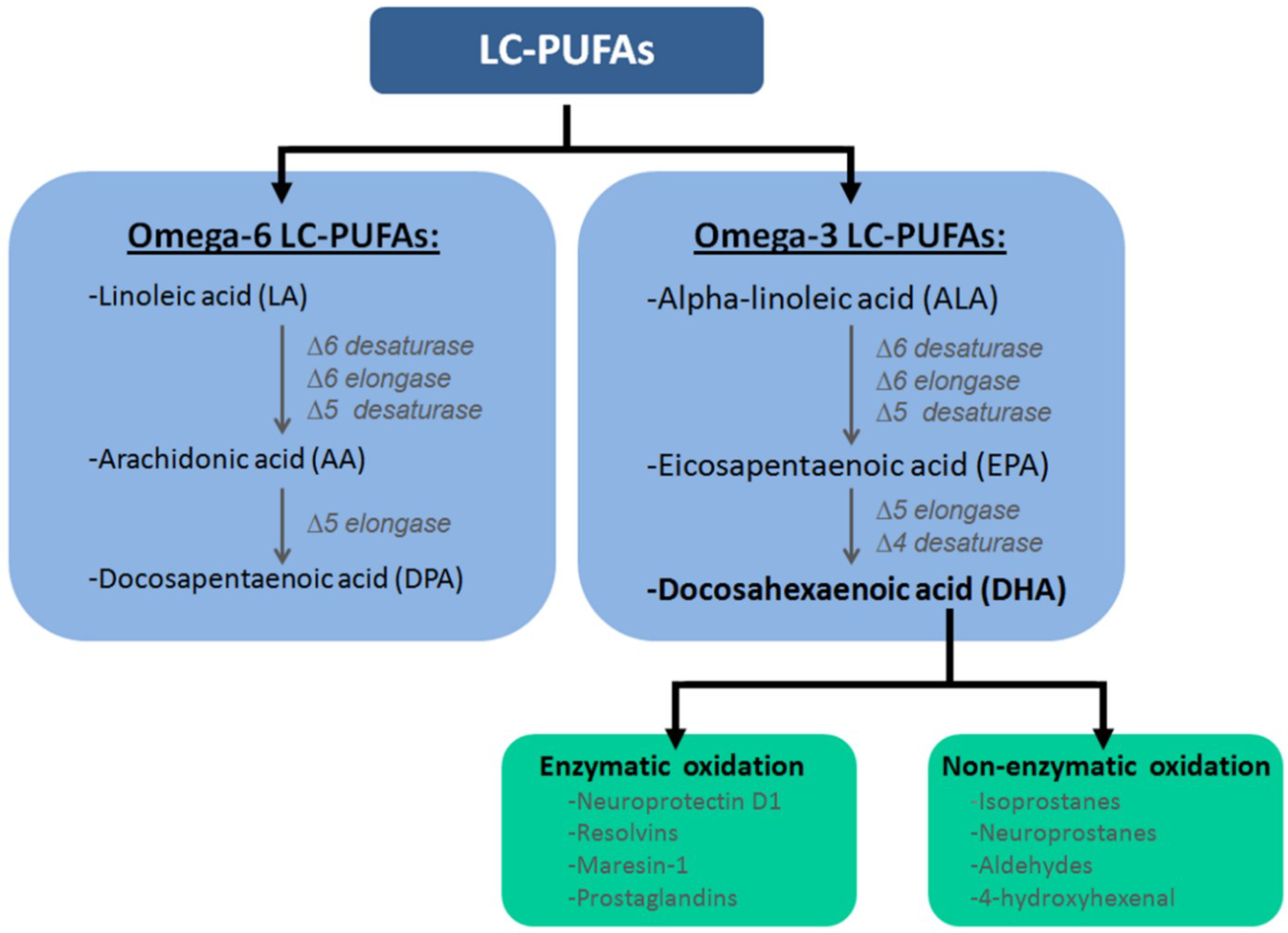
2. Metabolic Diseases
2.1. Processes Mediated by DHA
2.1.1. Inflammation
2.1.2. Insulin Metabolism
2.1.3. Adipocytes
2.1.4. Cardiovascular Health
2.2. DHA & Metabolic Diseases in Childhood
2.3. DHA & Metabolic Diseases in Adolescence
2.4. DHA and Metabolic Diseases in Adults

{kind=link}
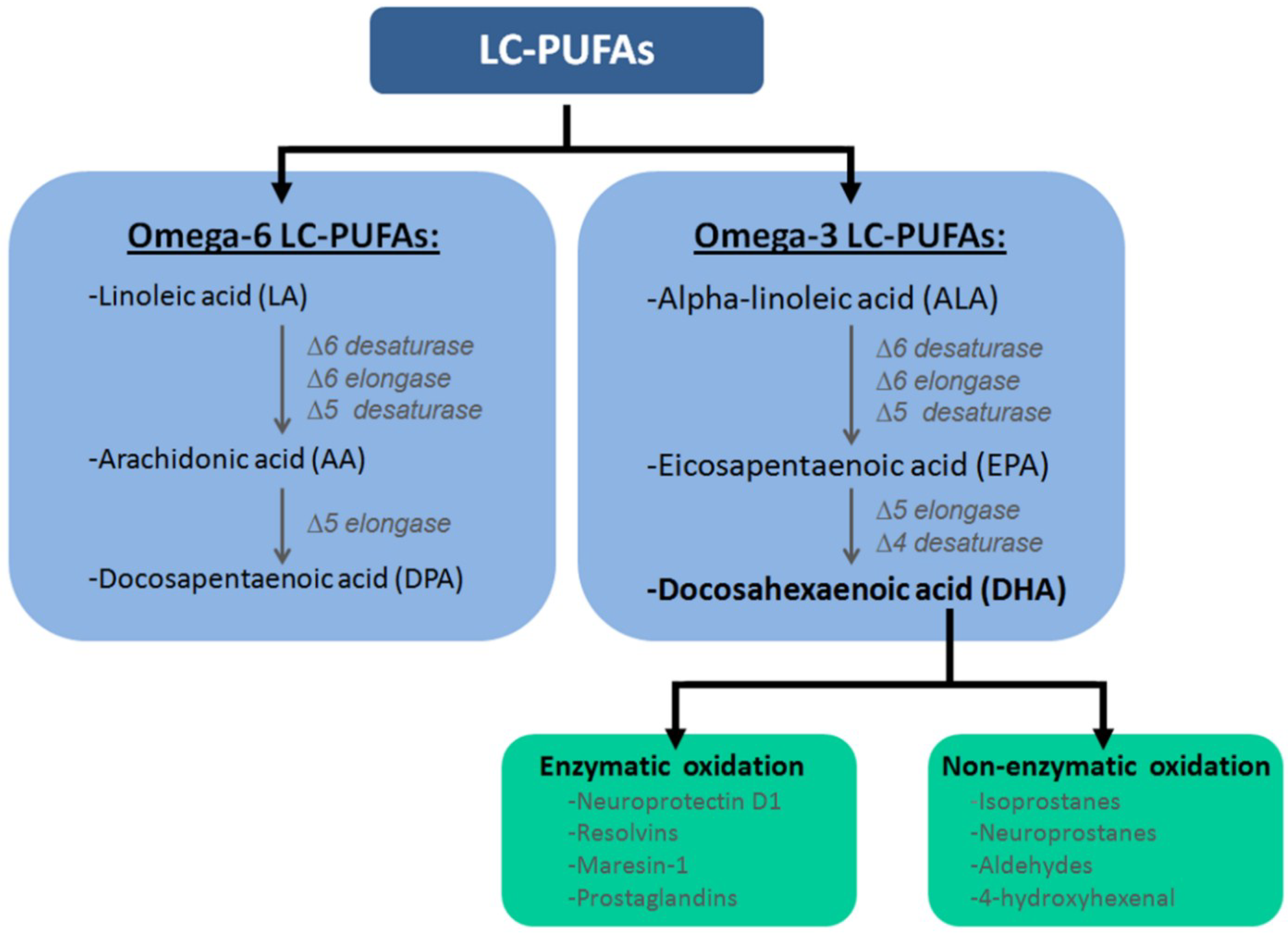{kind=link}
| Lifetime | Author | Year | N | Age (Years) | DHA | Outcome |
|---|---|---|---|---|---|---|
| Childhood | ||||||
| Desci [3] | 2002 | 80 ♂ & ♀ | 12 | Plasma phospholipids ARA & DHA | Values of arachidonic acid and docosahexaenoic acid were significantly lower in diabetic children than in controls. | |
| Burrows [2] | 2011 | 48 ♂ & ♀ | Non-obese: 9.0 ± 0.9 Obese: 8.9 ± 1.2 | Erythrocyte fatty acid; the Omega-3 index (O3I) composition | Obese children had altered erythrocyte fatty acid composition unrelated to reported dietary intake. A greater proportion of obese children had an omega-3 index of <4.0 (high risk) compared with non-obese children. | |
| Vasickova [87] | 2011 | 120 ♂ & ♀ (obese) | 10.0 ± 1.9 | 300 mg/day DHA + 42 mg/d EPA for 3 weeks | Daily consumption of 300mg DHA and 42 mg EPA for three weeks leads to an improvement of the anthropometric and lipid parameters in obese children [87]. | |
| Juarez Lopez [89] | 2013 | 201 ♂ & ♀ (obese and insulin resistant) | 11.6 ± 0.7 | 12 weeks LC-PUFA supplementation, 360 mg EPA & 240 mg DHA daily | LC-PUFA supplementation for 12 weeks decreased the concentrations of glucose, insulin, triglyceride-levels and BMI. | |
| Damsgaardt [88] | 2013 | 73 ♂ & ♀ | 10.29 ± 0.58 | Plasma DHA &EPA concentrations | DHA was positively associated with mean arterial pressure in boys. | |
| Adolescence | ||||||
| Dangardt [92] | 2012 | 25 ♂ & ♀ | 15.6 ± 0.9 ♀ 15.7 ± 1.0 ♂ | 1,2 g/d LC-PUFAs (DHA & EPA) for 3 months | Three months of supplementation of omega-3 LCPUFA improved glucose and insulin homeostasis in obese girls without influencing body weight. | |
| Adulthood | ||||||
| Rivellese [107] | 1997 | 16 ♂ & ♀ (NIDDM patients with hypertriglyeridemia) | 56.0 ± 3.0 | First two months: 0.96gr EPA and 1.59 g DHA per day Last four months: 0.64 gr EPA and 1.06 gr DHA per day | DHA and EPA significantly reduced plasma triglycerides and VLDL- triglycerides without significant changes in blood glucose control. | |
| Mori [105] | 1999 | 56 ♂(overweight & hyperlipidemic) | 49.1 ± 1.2 | 4 g/day DHA, EPA or olive oil (placebo) for 6 weeks | Purified DHA but not EPA reduced ambulatory BP and HR in mildly hyperlipidemic men. | |
| Mori [104] | 2000 | 59 ♂ (overweight & hyperlipidemic) | 50.6 ± 1.4 | 4 g/day DHA, EPA or olive oil (placebo) for 6 weeks | DHA enhances vasodilator mechanisms and attenuates constrictor responses in the forearm microcirculation. | |
| Woodman [102] | 2003 | ♂ & ♀ (Hypertensive and diabetic) | 40–75 | 4 g/day DHA, EPA or olive oil (placebo) for 6 weeks | DHA increased low density lipoprotein particle size | |
| Kelley [106] | 2007 | 34 ♂ | 55.0 ± 2.0 | 7.5 g DHA-oil for 90 days | DHA supplementation for 45 d significantly decreased concentrations of fasting triacylglycerol, large VLDL, and intermediate-density lipoproteins and the mean diameter of VLDL particles. | |
| Sneddon [108] | 2008 | 69 ♂ | 32.4 ± 2.3 | 3 g/day CLA + 3 g/day omega-3 LC-PUFAs | Supplementation with conjugated linoleic acids (CLAs) plus omega-3 LC-PUFAs prevents increased abdominal fat mass and raises fat-free mass and adiponectin levels in obese adults | |
| Micallef [6] | 2009 | 124 ♂ & ♀ | 43.79 ± 2.22 | Plasma levels of DHA & EPA | BMI, waist circumference and hip circumference were inversely correlated with n-3 PUFA, EPA and DHA (p < 0.05 for all) in the obese group. Obese individuals had significantly lower plasma concentrations of total n-3 PUFA, compared with healthy-weight individuals. | |
| Stirban [98] | 2010 | 34 ♂ & ♀ (T2DM) | 56.8 ± 8.3 | 2 g/d EPA & DHA for 6 weeks | Six weeks of supplementation with LC-PUFAs reduced the postprandial decrease in macrovascular function relative to placebo. LC-PUFAs supplementation improved postprandial microvascular function. | |
| Itariu [109] | 2012 | 55 ♂ & ♀ (obese) | 39.0 ± 2.0 | 3,36 g/d EPA & DHA for 8 weeks | n-3 PUFAs, which was well tolerated, decreased the gene expression of most analyzed inflammatory genes in subcutaneous adipose tissue (p <0.05) and increased production of anti-inflammatory eicosanoids in visceral adipose tissue and subcutaneous adipose tissue (p <0.05). | |
| Labonte [96] | 2013 | 12 ♂ (obese +T2DM) | 54.1 ± 7.2 | 3 g/d EPA & DHA for 8 weeks | In obese patients with T2DM, EPA&DHA supplementation did not affect the gene expression of pro-inflammatory cytokines in duodenal cells. | |
| Singhal [7] | 2013 | 328 ♂ & ♀ | 28.1 ± 4.8 | 1.6 g/day DHA | DHA supplementation did not improve endothelial function in healthy adolescents. Only triglyceride and very low-density lipoprotein concentrations were significant lower in DHA-supplemented individuals compared with controls. | |
| McDonald [101] | 2013 | 22 ♂ & ♀ Hypertensive and T2DM | 58.6 ± 8.8 | Daily supplementation of 1.8 g EPA and 1.5 g DHA for 8 weeks | LC-PUFAs diminish platelet superoxide production in T2DM hypertensive patients in vivo. | |
| Virtanen [99] | 2013 | 2122 ♂ | 53.1 ± 5.1 | Serum levels DHA, EPA, DPA | Men with higher serum level of EPA+DHA+DPA had a 33% lower multivariate-adjusted risk for T2DM. (Trend: p = 0.01) | |
| Mature and late adulthood | ||||||
| Woodman [100] | 2003 | 51 (39-♂ & 12-♀) (Hypertensive and diabetic) | 61.2 ± 1.2 | 4 g/day DHA, EPA or olive oil (placebo) for 6 weeks | DHA supplementation significantly reduced collagen aggregation and collagen-stimulated thromboxane release. | |
| Lemaitre [122] | 2003 | 54: Ischemic heart disease 125:non-fatal myocardial infarction 179: matched controls ♂ & ♀ | 79.1 ± 7.5 | DHA & EPA plasma phospholipids | Higher combined dietary intake of DHA and EPA, and possibly α-linolenic acid, may lower the risk of fatal ischemic heart disease in older adults. | |
| Tsitouras [123] | 2008 | 12 ♂ & ♀ | 66.1 ± 4.5 | Supplemented with 4 g/day EPA and DHA | Insulin sensitivity increased significantly after 8 weeks on the EPA- and DHA-diet, and serum C-reactive protein was significantly reduced. | |
| Heine-Böring [5] | 2010 | 1570 (686-♂ & 884-♀) | 64.0 ± 5.42 - ♂ 64.0 ± 5.6 -♀ | Food intake questionnaire; Dutch food composition table (DHA & EPA levels) | Subjects with a fish intake >19 g/d had a significantly lower prevalence of mild/moderate calcification. EPA plus DHA intake showed no significant associations. | |
| Djousse [48] | 2011 | 3088 ♂ & ♀ | 75.0 | Plasma phospholipids DHA and EPA | DHA is not associated with a higher incidence of T2DM, and individuals with higher EPA and DHA plasma concentrations had lower risk on T2DM. | |
2.5. DHA and Metabolic Diseases in Mature and Late Adulthood
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Balakumar, P.; Taneja, G. Fish oil and vascular endothelial protection: Bench to bedside. Free Radic. Biol. Med. 2012, 53, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Burrows, T.; Collins, C.E.; Garg, M.L. Omega-3 index, obesity and insulin resistance in children. Int. J. Pediatr. Obes. 2011, 6, e532–e539. [Google Scholar] [CrossRef] [PubMed]
- Decsi, T.; Minda, H.; Hermann, R.; Kozari, A.; Erhardt, E.; Burus, I.; Molnar, S.; Soltesz, G. Polyunsaturated fatty acids in plasma and erythrocyte membrane lipids of diabetic children. Prostaglandins Leukot. Essent. Fatty Acids 2002, 67, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Stehle, P. Impact of n-3 fatty acids on endothelial function: Results from human interventions studies. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Heine-Broring, R.C.; Brouwer, I.A.; Proenca, R.V.; van Rooij, F.J.; Hofman, A.; Oudkerk, M.; Witteman, J.C.; Geleijnse, J.M. Intake of fish and marine n-3 fatty acids in relation to coronary calcification: The rotterdam study. Am. J. Clin. Nutr. 2010, 91, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Micallef, M.; Munro, I.; Phang, M.; Garg, M. Plasma n-3 polyunsaturated fatty acids are negatively associated with obesity. Br. J. Nutr. 2009, 102, 1370–1374. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Lanigan, J.; Storry, C.; Low, S.; Birbara, T.; Lucas, A.; Deanfield, J. Docosahexaenoic acid supplementation, vascular function and risk factors for cardiovascular disease: A randomized controlled trial in young adults. J. Am. Heart Assoc. 2013, 2, e000283. [Google Scholar] [CrossRef] [PubMed]
- Singh, M. Essential fatty acids, dha and human brain. Indian J. Pediatr. 2005, 72, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Clandinin, M.T.; Jumpsen, J.; Suh, M. Relationship between fatty acid accretion, membrane composition, and biologic functions. J. Pediatr. 1994, 125, S25–S32. [Google Scholar] [CrossRef] [PubMed]
- Petrie, J.R.; Shrestha, P.; Liu, Q.; Mansour, M.P.; Wood, C.C.; Zhou, X.R.; Nichols, P.D.; Green, A.G.; Singh, S.P. Rapid expression of transgenes driven by seed-specific constructs in leaf tissue: Dha production. Plant Methods 2010, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.A. Polyunsaturated fatty acid synthesis and release by brain-derived cells in vitro. J. Mol. Neurosci. 2001, 16, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Demar, J.C., Jr.; Ma, K.; Chang, L.; Bell, J.M.; Rapoport, S.I. Alpha-linolenic acid does not contribute appreciably to docosahexaenoic acid within brain phospholipids of adult rats fed a diet enriched in docosahexaenoic acid. J. Neurochem. 2005, 94, 1063–1076. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Tanabe, Y.; Fujii, Y.; Kikuta, T.; Shibata, H.; Shido, O. Chronic administration of docosahexaenoic acid ameliorates the impairment of spatial cognition learning ability in amyloid beta-infused rats. J. Nutr. 2005, 135, 549–555. [Google Scholar] [PubMed]
- Suzuki, H.; Park, S.J.; Tamura, M.; Ando, S. Effect of the long-term feeding of dietary lipids on the learning ability, fatty acid composition of brain stem phospholipids and synaptic membrane fluidity in adult mice: A comparison of sardine oil diet with palm oil diet. Mech. Ageing Dev. 1998, 101, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.; Mirakaj, V.; Serhan, C.N. Functional metabolomics reveals novel active products in the dha metabolome. Front. Immunol. 2012, 3, 81. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.L. Resolvins: A family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Yang, R.; Martinod, K.; Kasuga, K.; Pillai, P.S.; Porter, T.F.; Oh, S.F.; Spite, M. Maresins: Novel macrophage mediators with potent antiinflammatory and proresolving actions. J. Exp. Med. 2009, 206, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Systems approach with inflammatory exudates uncovers novel anti-inflammatory and pro-resolving mediators. Prostaglandins Leukot. Essent. Fatty Acids 2008, 79, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Petasis, N.A. Resolvins and protectins in inflammation resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Torpy, J.M.; Lynm, C.; Glass, R.M. Jama patient page. Eating fish: Health benefits and risks. JAMA 2006, 296, 1926. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, J. Docosahexaenoic acid (DHA): An ancient nutrient for the modern human brain. Nutrients 2011, 3, 529–554. [Google Scholar] [CrossRef] [PubMed]
- Cossins, A.R.; Macdonald, A.G. The adaptation of biological membranes to temperature and pressure: Fish from the deep and cold. J. Bioenerg. Biomembr. 1989, 21, 115–135. [Google Scholar] [CrossRef] [PubMed]
- Luchtman, D.W.; Song, C. Cognitive enhancement by omega-3 fatty acids from child-hood to old age: Findings from animal and clinical studies. Neuropharmacology 2013, 64, 550–565. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, M.; Saint-Pierre, M.; Julien, C.; Salem, N., Jr.; Cicchetti, F.; Calon, F. Beneficial effects of dietary omega-3 polyunsaturated fatty acid on toxin-induced neuronal degeneration in an animal model of parkinson’s disease. FASEB J. 2008, 22, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Chytrova, G.; Ying, Z.; Gomez-Pinilla, F. Exercise contributes to the effects of DHA dietary supplementation by acting on membrane-related synaptic systems. Brain Res. 2010, 1341, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hossain, S.; Shimada, T.; Shido, O. Docosahexaenoic acid-induced protective effect against impaired learning in amyloid beta-infused rats is associated with increased synaptosomal membrane fluidity. Clin. Exp. Pharmacol. Physiol. 2006, 33, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Teague, W.E.; Fuller, N.L.; Rand, R.P.; Gawrisch, K. Polyunsaturated lipids in membrane fusion events. Cell. Mol. Biol. Lett. 2002, 7, 262–264. [Google Scholar] [PubMed]
- Ouellet, M.; Emond, V.; Chen, C.T.; Julien, C.; Bourasset, F.; Oddo, S.; LaFerla, F.; Bazinet, R.P.; Calon, F. Diffusion of docosahexaenoic and eicosapentaenoic acids through the blood-brain barrier: An in situ cerebral perfusion study. Neurochem. Int. 2009, 55, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. Fast flip-flop of cholesterol and fatty acids in membranes: Implications for membrane transport proteins. Curr. Opin. Lipidol. 2003, 14, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Doege, H.; Stahl, A. Protein-mediated fatty acid uptake: Novel insights from in vivo models. Physiology 2006, 21, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Nikolakopoulou, Z.; Shaikh, M.H.; Dehlawi, H.; Michael-Titus, A.T.; Parkinson, E.K. The induction of apoptosis in pre-malignant keratinocytes by omega-3 polyunsaturated fatty acids docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) is inhibited by albumin. Toxicol. Lett. 2013, 218, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Coburn, C.T.; Hajri, T.; Ibrahimi, A.; Abumrad, N.A. Role of CD36 in membrane transport and utilization of long-chain fatty acids by different tissues. J. Mol. Neurosci. 2001, 16, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Gargiulo, C.E.; Stuhlsatz-Krouper, S.M.; Schaffer, J.E. Localization of adipocyte long-chain fatty acyl-coa synthetase at the plasma membrane. J. Lipid Res. 1999, 40, 881–892. [Google Scholar] [PubMed]
- Kazantzis, M.; Stahl, A. Fatty acid transport proteins, implications in physiology and disease. Biochim. Biophys. Acta 2012, 1821, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.; Neergaard, T.B.; Gaigg, B.; Jensen, M.V.; Hansen, J.K. Role of acyl-CoA binding protein in acyl-CoA metabolism and acyl-CoA-mediated cell signaling. J. Nutr. 2000, 130, 294S–298S. [Google Scholar] [PubMed]
- Storch, J.; McDermott, L. Structural and functional analysis of fatty acid-binding proteins. J. Lipid Res. 2009, 50 (Suppl.), S126–S131. [Google Scholar] [PubMed]
- Storch, J.; Thumser, A.E. Tissue-specific functions in the fatty acid-binding protein family. J. Biol. Chem. 2010, 285, 32679–32683. [Google Scholar] [CrossRef] [PubMed]
- Glatz, J.F.; Luiken, J.J.; van Bilsen, M.; van der Vusse, G.J. Cellular lipid binding proteins as facilitators and regulators of lipid metabolism. Mol. Cell. Biochem. 2002, 239, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Khnykin, D.; Miner, J.H.; Jahnsen, F. Role of fatty acid transporters in epidermis: Implications for health and disease. Dermato-Endocrinology 2011, 3, 53–61. [Google Scholar] [CrossRef] [PubMed]
- WHO. Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/index.html (accessed on 9 August 2012).
- Van Greevenbroek, M.M.; Schalkwijk, C.G.; Stehouwer, C.D. Obesity-associated low-grade inflammation in type 2 diabetes mellitus: Causes and consequences. Neth. J. Med. 2013, 71, 174–187. [Google Scholar]
- Bender, S.B.; McGraw, A.P.; Jaffe, I.Z.; Sowers, J.R. Mineralocorticoid receptor-mediated vascular insulin resistance: An early contributor to diabetes-related vascular disease? Diabetes 2013, 62, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Lean, M.E. Obesity: Burdens of illness and strategies for prevention or management. Drugs Today (Barc.) 2000, 36, 773–784. [Google Scholar] [CrossRef]
- Andersson, C.X.; Gustafson, B.; Hammarstedt, A.; Hedjazifar, S.; Smith, U. Inflamed adipose tissue, insulin resistance and vascular injury. Diabetes Metab. Res. Rev. 2008, 24, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Morrish, N.J.; Wang, S.L.; Stevens, L.K.; Fuller, J.H.; Keen, H. Mortality and causes of death in the who multinational study of vascular disease in diabetes. Diabetologia 2001, 44, S14–S21. [Google Scholar] [CrossRef] [PubMed]
- Dangardt, F.; Osika, W.; Chen, Y.; Nilsson, U.; Gan, L.M.; Gronowitz, E.; Strandvik, B.; Friberg, P. Omega-3 fatty acid supplementation improves vascular function and reduces inflammation in obese adolescents. Atherosclerosis 2010, 212, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Djousse, L.; Biggs, M.L.; Lemaitre, R.N.; King, I.B.; Song, X.; Ix, J.H.; Mukamal, K.J.; Siscovick, D.S.; Mozaffarian, D. Plasma omega-3 fatty acids and incident diabetes in older adults. Am. J. Clin. Nutr. 2011, 94, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Huang, H.; Sacks, F.M.; Rimm, E.B.; Wang, M.; Siscovick, D.S. Plasma phospholipid long-chain omega-3 fatty acids and total and cause-specific mortality in older adults: A cohort study. Ann. Intern. Med. 2013, 158, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Spiegelman, D.; Sacks, F.M.; Rimm, E.B.; Siscovick, D.S. Circulating long-chain omega-3 fatty acids and incidence of congestive heart failure in older adults: The cardiovascular health study: A cohort study. Ann. Intern. Med. 2011, 155, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Antuna-Puente, B.; Feve, B.; Fellahi, S.; Bastard, J.P. Adipokines: The missing link between insulin resistance and obesity. Diabetes Metab. 2008, 34, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.; Feve, B.; Ferre, P.; Halimi, S.; Baizri, H.; Bordier, L.; Guiu, G.; Dupuy, O.; Bauduceau, B.; Mayaudon, H. Diabetes and inflammation: Fundamental aspects and clinical implications. Diabetes Metab. 2010, 36, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Periz, A.; Horrillo, R.; Ferre, N.; Gronert, K.; Dong, B.; Moran-Salvador, E.; Titos, E.; Martinez-Clemente, M.; Lopez-Parra, M.; Arroyo, V.; et al. Obesity-induced insulin resistance and hepatic steatosis are alleviated by omega-3 fatty acids: A role for resolvins and protectins. FASEB J. 2009, 23, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, T.; Wissuwa, B.; Tian, Y.; Tajima, N.; Xu, R.; Bauer, M.; Heinemann, S.H.; Hou, S. Omega-3 fatty acids lower blood pressure by directly activating large-conductance Ca2+-dependent K+ channels. Proc. Natl. Acad. Sci. USA 2013, 110, 4816–4821. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, O.Y.; Cho, Y.; Chung, J.H.; Jung, Y.S.; Hwang, G.S.; Shin, M.J. Plasma phospholipid fatty acid composition in ischemic stroke: Importance of docosahexaenoic acid in the risk for intracranial atherosclerotic stenosis. Atherosclerosis 2012, 225, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Pickup, J.C.; Mattock, M.B.; Chusney, G.D.; Burt, D. Niddm as a disease of the innate immune system: Association of acute-phase reactants and interleukin-6 with metabolic syndrome X. Diabetologia 1997, 40, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Feve, B.; Bastard, J.P. The role of interleukins in insulin resistance and type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2009, 5, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, K.; Vallance, P. Inflammatory cytokines impair endothelium-dependent dilatation in human veins in vivo. Circulation 1997, 96, 3042–3047. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Ba, Z.F.; Chaudry, I.H. Administration of tumor necrosis factor-alpha in vivo depresses endothelium-dependent relaxation. Am. J. Physiol. 1994, 266, H2535–H2541. [Google Scholar] [PubMed]
- Das, U.N. Is obesity an inflammatory condition? Nutrition 2001, 17, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Murumalla, R.K.; Gunasekaran, M.K.; Padhan, J.K.; Bencharif, K.; Gence, L.; Festy, F.; Cesari, M.; Roche, R.; Hoareau, L. Fatty acids do not pay the toll: Effect of sfa and pufa on human adipose tissue and mature adipocytes inflammation. Lipids Health Dis. 2012, 11, 175. [Google Scholar] [CrossRef] [PubMed]
- Titos, E.; Rius, B.; Gonzalez-Periz, A.; Lopez-Vicario, C.; Moran-Salvador, E.; Martinez-Clemente, M.; Arroyo, V.; Claria, J. Resolvin d1 and its precursor docosahexaenoic acid promote resolution of adipose tissue inflammation by eliciting macrophage polarization toward an m2-like phenotype. J. Immunol. 2011, 187, 5408–5418. [Google Scholar] [CrossRef] [PubMed]
- Bonadonna, R.C.; Groop, L.; Kraemer, N.; Ferrannini, E.; del Prato, S.; deFronzo, R.A. Obesity and insulin resistance in humans: A dose-response study. Metabolism 1990, 39, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Steinberger, J.; Daniels, S.R.; American Heart Association Atherosclerosis, Hypertension; Obesity in the Young Committee; American Heart Association Diabetes Committee. Obesity, insulin resistance, diabetes, and cardiovascular risk in children: An american heart association scientific statement from the atherosclerosis, hypertension, and obesity in the young committee (council on cardiovascular disease in the young) and the diabetes committee (council on nutrition, physical activity, and metabolism). Circulation 2003, 107, 1448–1453. [Google Scholar] [CrossRef] [PubMed]
- Makni, E.; Moalla, W.; Lac, G.; Aouichaoui, C.; Cannon, D.; Elloumi, M.; Tabka, Z. The homeostasis model assessment-adiponectin (HOMA-AD) is the most sensitive predictor of insulin resistance in obese children. Ann. Endocrinol. (Paris) 2012, 73, 26–33. [Google Scholar] [CrossRef]
- Stienstra, R.; Tack, C.J.; Kanneganti, T.D.; Joosten, L.A.; Netea, M.G. The inflammasome puts obesity in the danger zone. Cell Metab. 2012, 15, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Bastard, J.P.; Maachi, M.; Van Nhieu, J.T.; Jardel, C.; Bruckert, E.; Grimaldi, A.; Robert, J.J.; Capeau, J.; Hainque, B. Adipose tissue IL-6 content correlates with resistance to insulin activation of glucose uptake both in vivo and in vitro. J. Clin. Endocrinol. Metab. 2002, 87, 2084–2089. [Google Scholar] [CrossRef] [PubMed]
- Botton, J.; Heude, B.; Kettaneh, A.; Borys, J.M.; Lommez, A.; Bresson, J.L.; Ducimetiere, P.; Charles, M.A. Cardiovascular risk factor levels and their relationships with overweight and fat distribution in children: The Fleurbaix Laventie Ville Sante II study. Metabolism 2007, 56, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Bandyopadhyay, A. Inflammation: The link between insulin resistance, obesity and diabetes. Trends Immunol. 2004, 25, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.; Wilhelmsen, L.; Welin, C.; Eriksson, H.; Welin, L.; Rosengren, A. Obesity, smoking and secular trends in cardiovascular risk factors in middle-aged women: Data from population studies in goteborg from 1980 to 2003. J. Intern. Med. 2010, 268, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [PubMed]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Sorisky, A. Molecular links between obesity and cardiovascular disease. Am. J. Ther. 2002, 9, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Della-Fera, M.; Lin, J.; Baile, C.A. Docosahexaenoic acid inhibits adipocyte differentiation and induces apoptosis in 3T3-L1 preadipocytes. J. Nutr. 2006, 136, 2965–2969. [Google Scholar] [PubMed]
- Bang, H.O.; Dyerberg, J.; Hjoorne, N. The composition of food consumed by greenland eskimos. Acta Med. Scand. 1976, 200, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Calo, L.; Bianconi, L.; Colivicchi, F.; Lamberti, F.; Loricchio, M.L.; de Ruvo, E.; Meo, A.; Pandozi, C.; Staibano, M.; Santini, M. N-3 fatty acids for the prevention of atrial fibrillation after coronary artery bypass surgery: A randomized, controlled trial. J. Am. Coll. Cardiol. 2005, 45, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Dallongeville, J.; Yarnell, J.; Ducimetiere, P.; Arveiler, D.; Ferrieres, J.; Montaye, M.; Luc, G.; Evans, A.; Bingham, A.; Hass, B.; et al. Fish consumption is associated with lower heart rates. Circulation 2003, 108, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.B.; Niaz, M.A.; Sharma, J.P.; Kumar, R.; Rastogi, V.; Moshiri, M. Randomized, double-blind, placebo-controlled trial of fish oil and mustard oil in patients with suspected acute myocardial infarction: The indian experiment of infarct survival—4. Cardiovasc. Drugs Ther. 1997, 11, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: Results of the gissi-prevenzione trial. Gruppo italiano per lo studio della sopravvivenza nell'infarto miocardico. Lancet 1999, 354, 447–455.
- Lorente-Cebrian, S.; Costa, A.G.; Navas-Carretero, S.; Zabala, M.; Martinez, J.A.; Moreno-Aliaga, M.J. Role of omega-3 fatty acids in obesity, metabolic syndrome, and cardiovascular diseases: A review of the evidence. J. Physiol. Biochem. 2013, 69, 633–651. [Google Scholar] [CrossRef] [PubMed]
- Thies, F.; Garry, J.M.; Yaqoob, P.; Rerkasem, K.; Williams, J.; Shearman, C.P.; Gallagher, P.J.; Calder, P.C.; Grimble, R.F. Association of n-3 polyunsaturated fatty acids with stability of atherosclerotic plaques: A randomised controlled trial. Lancet 2003, 361, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Baumann, K.H.; Hessel, F.; Larass, I.; Muller, T.; Angerer, P.; Kiefl, R.; von Schacky, C. Dietary omega-3, omega-6, and omega-9 unsaturated fatty acids and growth factor and cytokine gene expression in unstimulated and stimulated monocytes. A randomized volunteer study. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Goodfellow, J.; Bellamy, M.F.; Ramsey, M.W.; Jones, C.J.; Lewis, M.J. Dietary supplementation with marine omega-3 fatty acids improve systemic large artery endothelial function in subjects with hypercholesterolemia. J. Am. Coll. Cardiol. 2000, 35, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Kasbi Chadli, F.; Andre, A.; Prieur, X.; Loirand, G.; Meynier, A.; Krempf, M.; Nguyen, P.; Ouguerram, K. N-3 PUFA prevent metabolic disturbances associated with obesity and improve endothelial function in golden syrian hamsters fed with a high-fat diet. Br. J. Nutr. 2011, 107, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Van Emmerik, N.M.; Renders, C.M.; van de Veer, M.; van Buuren, S.; van der Baan-Slootweg, O.H.; Kist-van Holthe, J.E.; Hirasing, R.A. High cardiovascular risk in severely obese young children and adolescents. Arch. Dis. Child. 2012, 97, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, D.; Satnick, A.; Abell, R.; Messina, C.R.; Chawla, A. The prevalence of abnormal metabolic parameters in obese and overweight children. JPEN J. Parenter Enteral. Nutr. 2014, 38, 852–855. [Google Scholar] [CrossRef] [PubMed]
- Vasickova, L.; Stavek, P.; Suchanek, P. Possible effect of dha intake on body weight reduction and lipid metabolism in obese children. Neuro Endocrinol. Lett. 2011, 32 (Suppl. 2), 64–67. [Google Scholar] [PubMed]
- Damsgaard, C.T.; Stark, K.D.; Hjorth, M.F.; Biltoft-Jensen, A.; Astrup, A.; Michaelsen, K.F.; Lauritzen, L. N-3 PUFA status in school children is associated with beneficial lipid profile, reduced physical activity and increased blood pressure in boys. Br. J. Nutr. 2013, 110, 1304–1312. [Google Scholar] [CrossRef] [PubMed]
- Juarez-Lopez, C.; Klunder-Klunder, M.; Madrigal-Azcarate, A.; Flores-Huerta, S. Omega-3 polyunsaturated fatty acids reduce insulin resistance and triglycerides in obese children and adolescents. Pediatr. Diabetes 2013, 14, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Raiko, J.R.; Oikonen, M.; Wendelin-Saarenhovi, M.; Siitonen, N.; Kahonen, M.; Lehtimaki, T.; Viikari, J.; Jula, A.; Loo, B.M.; Huupponen, R.; et al. Plasminogen activator inhitor-1 associates with cardiovascular risk factors in healthy young adults in the cardiovascular risk in young finns study. Atherosclerosis 2012, 224, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Yau, P.L.; Javier, D.C.; Ryan, C.M.; Tsui, W.H.; Ardekani, B.A.; Ten, S.; Convit, A. Preliminary evidence for brain complications in obese adolescents with type 2 diabetes mellitus. Diabetologia 2010, 53, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Dangardt, F.; Chen, Y.; Gronowitz, E.; Dahlgren, J.; Friberg, P.; Strandvik, B. High physiological omega-3 fatty acid supplementation affects muscle fatty acid composition and glucose and insulin homeostasis in obese adolescents. J. Nutr. Metab. 2012, 2012, 395757. [Google Scholar] [CrossRef] [PubMed]
- Rise, P.; Tragni, E.; Ghezzi, S.; Agostoni, C.; Marangoni, F.; Poli, A.; Catapano, A.L.; Siani, A.; Iacoviello, L.; Galli, C.; et al. Different patterns characterize omega 6 and omega 3 long chain polyunsaturated fatty acid levels in blood from Italian infants, children, adults and elderly. Prostaglandins Leukot. Essent. Fatty Acids 2013, 89, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Federation, I.D. IDF Diabetes Atlas. Available online: http://www.idf.org/diabetesatlas (accessed on 18 June 2014).
- Ergotron, I. Obesity, Cardiovascular Disease and Diabetes Regional Statistics by Country/State/Province. Available online: http://www.juststand.org/Portals/3/literature/regionalStats.pdf (accessed on 2 April 2014).
- Labonte, M.E.; Couture, P.; Tremblay, A.J.; Hogue, J.C.; Lemelin, V.; Lamarche, B. Eicosapentaenoic and docosahexaenoic acid supplementation and inflammatory gene expression in the duodenum of obese patients with type 2 diabetes. Nutr. J. 2013, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Brostow, D.P.; Odegaard, A.O.; Koh, W.P.; Duval, S.; Gross, M.D.; Yuan, J.M.; Pereira, M.A. Omega-3 fatty acids and incident type 2 diabetes: The singapore chinese health study. Am. J. Clin. Nutr. 2011, 94, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Stirban, A.; Nandrean, S.; Gotting, C.; Tamler, R.; Pop, A.; Negrean, M.; Gawlowski, T.; Stratmann, B.; Tschoepe, D. Effects of n-3 fatty acids on macro- and microvascular function in subjects with type 2 diabetes mellitus. Am. J. Clin. Nutr. 2010, 91, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, J.K.; Mursu, J.; Voutilainen, S.; Uusitupa, M.; Tuomainen, T.P. Serum omega-3 polyunsaturated fatty acids and risk of incident type 2 diabetes in men: The kuopio ischaemic heart disease risk factor study. Diabetes Care 2014, 37, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Woodman, R.J.; Mori, T.A.; Burke, V.; Puddey, I.B.; Barden, A.; Watts, G.F.; Beilin, L.J. Effects of purified eicosapentaenoic acid and docosahexaenoic acid on platelet, fibrinolytic and vascular function in hypertensive type 2 diabetic patients. Atherosclerosis 2003, 166, 85–93. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.M.; O’Kane, F.; McConville, M.; Devine, A.B.; McVeigh, G.E. Platelet redox balance in diabetic patients with hypertension improved by n-3 fatty acids. Diabetes Care 2013, 36, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Woodman, R.J.; Mori, T.A.; Burke, V.; Puddey, I.B.; Watts, G.F.; Best, J.D.; Beilin, L.J. Docosahexaenoic acid but not eicosapentaenoic acid increases ldl particle size in treated hypertensive type 2 diabetic patients. Diabetes Care 2003, 26, 253. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Burke, V.; Puddey, I.B.; Watts, G.F.; O’Neal, D.N.; Best, J.D.; Beilin, L.J. Purified eicosapentaenoic and docosahexaenoic acids have differential effects on serum lipids and lipoproteins, ldl particle size, glucose, and insulin in mildly hyperlipidemic men. Am. J. Clin. Nutr. 2000, 71, 1085–1094. [Google Scholar] [PubMed]
- Mori, T.A.; Watts, G.F.; Burke, V.; Hilme, E.; Puddey, I.B.; Beilin, L.J. Differential effects of eicosapentaenoic acid and docosahexaenoic acid on vascular reactivity of the forearm microcirculation in hyperlipidemic, overweight men. Circulation 2000, 102, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Bao, D.Q.; Burke, V.; Puddey, I.B.; Beilin, L.J. Docosahexaenoic acid but not eicosapentaenoic acid lowers ambulatory blood pressure and heart rate in humans. Hypertension 1999, 34, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Siegel, D.; Vemuri, M.; Mackey, B.E. Docosahexaenoic acid supplementation improves fasting and postprandial lipid profiles in hypertriglyceridemic men. Am. J. Clin. Nutr. 2007, 86, 324–333. [Google Scholar] [PubMed]
- Rivellese, A.A.; Maffettone, A.; Iovine, C.; Di Marino, L.; Annuzzi, G.; Mancini, M.; Riccardi, G. Long-term effects of fish oil on insulin resistance and plasma lipoproteins in niddm patients with hypertriglyceridemia. Diabetes Care 1996, 19, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, A.A.; Tsofliou, F.; Fyfe, C.L.; Matheson, I.; Jackson, D.M.; Horgan, G.; Winzell, M.S.; Wahle, K.W.; Ahren, B.; Williams, L.M. Effect of a conjugated linoleic acid and omega-3 fatty acid mixture on body composition and adiponectin. Obesity (Silver Spring) 2008, 16, 1019–1024. [Google Scholar] [CrossRef]
- Itariu, B.K.; Zeyda, M.; Hochbrugger, E.E.; Neuhofer, A.; Prager, G.; Schindler, K.; Bohdjalian, A.; Mascher, D.; Vangala, S.; Schranz, M.; et al. Long-chain n-3 pufas reduce adipose tissue and systemic inflammation in severely obese nondiabetic patients: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 96, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; AHA Nutrition Committee. American Heart Association. Omega-3 fatty acids and cardiovascular disease: New recommendations from the american heart association. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Geleijnse, J.M.; Giltay, E.J.; Grobbee, D.E.; Donders, A.R.; Kok, F.J. Blood pressure response to fish oil supplementation: Metaregression analysis of randomized trials. J. Hypertens. 2002, 20, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Von Schacky, C.; Harris, W.S. Cardiovascular risk and the omega-3 index. J. Cardiovasc. Med. 2007, 8 (Suppl. 1), S46–S49. [Google Scholar] [CrossRef]
- Connor, W.E. Importance of n-3 fatty acids in health and disease. Am. J. Clin. Nutr. 2000, 71, 171S–175S. [Google Scholar] [PubMed]
- Okuda, N.; Ueshima, H.; Okayama, A.; Saitoh, S.; Nakagawa, H.; Rodriguez, B.L.; Sakata, K.; Choudhury, S.R.; Curb, J.D.; Stamler, J.; et al. Relation of long chain n-3 polyunsaturated fatty acid intake to serum high density lipoprotein cholesterol among japanese men in japan and japanese-american men in hawaii: The interlipid study. Atherosclerosis 2005, 178, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Leaf, A.; Kang, J.X.; Xiao, Y.F.; Billman, G.E. Clinical prevention of sudden cardiac death by n-3 polyunsaturated fatty acids and mechanism of prevention of arrhythmias by n-3 fish oils. Circulation 2003, 107, 2646–2652. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Lu, G.; Rambjor, G.S.; Walen, A.I.; Ontko, J.A.; Cheng, Q.; Windsor, S.L. Influence of n-3 fatty acid supplementation on the endogenous activities of plasma lipases. Am. J. Clin. Nutr. 1997, 66, 254–260. [Google Scholar] [PubMed]
- Calder, P.C.; Yaqoob, P. Omega-3 (n-3) fatty acids, cardiovascular disease and stability of atherosclerotic plaques. Cell. Mol. Biol. 2010, 56, 28–37. [Google Scholar] [PubMed]
- Dyerberg, J.; Bang, H.O. Lipid metabolism, atherogenesis, and haemostasis in eskimos: The role of the prostaglandin-3 family. Haemostasis 1979, 8, 227–233. [Google Scholar] [PubMed]
- He, K.; Song, Y.; Daviglus, M.L.; Liu, K.; Van Horn, L.; Dyer, A.R.; Greenland, P. Accumulated evidence on fish consumption and coronary heart disease mortality: A meta-analysis of cohort studies. Circulation 2004, 109, 2705–2711. [Google Scholar] [CrossRef] [PubMed]
- Janssen, C.I.; Kiliaan, A.J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: The influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid Res. 2014, 53, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cottin, S.C.; Sanders, T.A.; Hall, W.L. The differential effects of epa and dha on cardiovascular risk factors. Proc. Nutr. Soc. 2011, 70, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, R.N.; King, I.B.; Mozaffarian, D.; Kuller, L.H.; Tracy, R.P.; Siscovick, D.S. N-3 polyunsaturated fatty acids, fatal ischemic heart disease, and nonfatal myocardial infarction in older adults: The cardiovascular health study. Am. J. Clin. Nutr. 2003, 77, 319–325. [Google Scholar] [PubMed]
- Tsitouras, P.D.; Gucciardo, F.; Salbe, A.D.; Heward, C.; Harman, S.M. High omega-3 fat intake improves insulin sensitivity and reduces CRP and IL6, but does not affect other endocrine axes in healthy older adults. Horm. Metab. Res. 2008, 40, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Manzato, E.; Romanato, G.; Zambon, S.; Corti, M.C.; Baggio, G.; Sartori, L.; Musacchio, E.; Zanoni, S.; Marin, R.; Crepaldi, G. Metabolic syndrome and cardiovascular disease in the elderly: The Progetto Veneto Anziani (Pro.V.A.) study. Aging Clin. Exp. Res. 2008, 20, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Czernichow, S.; Greenfield, J.R.; Galan, P.; Bastard, J.P.; Charnaux, N.; Samaras, K.; Safar, M.E.; Blacher, J.; Hercberg, S.; Levy, B.I. Microvascular dysfunction in healthy insulin-sensitive overweight individuals. J. Hypertens. 2010, 28, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, J.K.; Nyantika, A.N.; Kauhanen, J.; Voutilainen, S.; Tuomainen, T.P. Serum long-chain n-3 polyunsaturated fatty acids, methylmercury and blood pressure in an older population. Hypertens. Res. 2012, 35, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Engler, M.M.; Engler, M.B.; Malloy, M.; Chiu, E.; Besio, D.; Paul, S.; Stuehlinger, M.; Morrow, J.; Ridker, P.; Rifai, N.; et al. Docosahexaenoic acid restores endothelial function in children with hyperlipidemia: Results from the early study. Int. J. Clin. Pharmacol. Ther. 2004, 42, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Saito, E.; Okada, T.; Abe, Y.; Kuromori, Y.; Miyashita, M.; Iwata, F.; Hara, M.; Ayusawa, M.; Mugishima, H.; Kitamura, Y. Docosahexaenoic acid content in plasma phospholipids and desaturase indices in obese children. J. Atheroscler. Thromb. 2011, 18, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Kalofoutis, C.; Piperi, C.; Kalofoutis, A.; Harris, F.; Phoenix, D.; Singh, J. Type II diabetes mellitus and cardiovascular risk factors: Current therapeutic approaches. Exp. Clin. Cardiol. 2007, 12, 17–28. [Google Scholar] [PubMed]
- Stanley, J.C.; Elsom, R.L.; Calder, P.C.; Griffin, B.A.; Harris, W.S.; Jebb, S.A.; Lovegrove, J.A.; Moore, C.S.; Riemersma, R.A.; Sanders, T.A. UK food standards agency workshop report: The effects of the dietary n-6:N-3 fatty acid ratio on cardiovascular health. Br. J. Nutr. 2007, 98, 1305–1310. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Harris, M.F.; Powell-Davies, G.; Jayasinghe, U.W.; Flack, J.; Georgiou, A.; Burns, J.R.; Penn, D.L. Cardiovascular risk levels in general practice patients with type 2 diabetes in rural and urban areas. Aust. J. Rural Health 2007, 15, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.N.; Harris, W.S. New evidence for the cardiovascular benefits of long chain omega-3 fatty acids. Curr. Atheroscler. Rep. 2007, 9, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Harris, M.F.; Davies, G.P.; Jayasinghe, U.W.; Flack, J.; Georgiou, A.; Burns, J.R.; Penn, D.L. Cardiovascular risk management and its impact in Australian general practice patients with type 2 diabetes in urban and rural areas. Int. J. Clin. Pract. 2008, 62, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Daniels, S.R.; Long, B.; Crow, S.; Styne, D.; Sothern, M.; Vargas-Rodriguez, I.; Harris, L.; Walch, J.; Jasinsky, O.; Cwik, K.; et al. Cardiovascular effects of sibutramine in the treatment of obese adolescents: Results of a randomized, double-blind, placebo-controlled study. Pediatrics 2007, 120, e147–e157. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D. Adiposity indices and dementia. Lancet Neurol. 2006, 5, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D.R. A life course of adiposity and dementia. Eur. J. Pharmacol. 2008, 585, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D.R. Adiposity hormones and dementia. J. Neurol. Sci. 2010, 299, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D.R. Adiposity and cognitive decline: Underlying mechanisms. J. Alzheimers. Dis. 2012, 30, S97–S112. [Google Scholar] [PubMed]
- Gustafson, D.R.; Steen, B.; Skoog, I. Body mass index and white matter lesions in elderly women. An 18-year longitudinal study. Int. Psychogeriatr. 2004, 16, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Profenno, L.A.; Porsteinsson, A.P.; Faraone, S.V. Meta-analysis of alzheimer’s disease risk with obesity, diabetes, and related disorders. Biol. Psychiatry 2010, 67, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D.; Rothenberg, E.; Blennow, K.; Steen, B.; Skoog, I. An 18-year follow-up of overweight and risk of alzheimer disease. Arch. Intern. Med. 2003, 163, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D.R.; Backman, K.; Waern, M.; Ostling, S.; Guo, X.; Zandi, P.; Mielke, M.M.; Bengtsson, C.; Skoog, I. Adiposity indicators and dementia over 32 years in sweden. Neurology 2009, 73, 1559–1566. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arnoldussen, I.A.C.; Kiliaan, A.J. Impact of DHA on Metabolic Diseases from Womb to Tomb. Mar. Drugs 2014, 12, 6190-6212. https://doi.org/10.3390/md12126190
Arnoldussen IAC, Kiliaan AJ. Impact of DHA on Metabolic Diseases from Womb to Tomb. Marine Drugs. 2014; 12(12):6190-6212. https://doi.org/10.3390/md12126190
Chicago/Turabian StyleArnoldussen, Ilse A. C., and Amanda J. Kiliaan. 2014. "Impact of DHA on Metabolic Diseases from Womb to Tomb" Marine Drugs 12, no. 12: 6190-6212. https://doi.org/10.3390/md12126190
APA StyleArnoldussen, I. A. C., & Kiliaan, A. J. (2014). Impact of DHA on Metabolic Diseases from Womb to Tomb. Marine Drugs, 12(12), 6190-6212. https://doi.org/10.3390/md12126190