New Invertebrate Vectors for PST, Spirolides and Okadaic Acid in the North Atlantic


 and
and 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Trophic level | Edibility | Monitored | References |
|---|---|---|---|---|
| Gibbula sp. | Grazer | Yes | No | [20,50] |
| Monodonta sp. | Grazer | Yes | No | [20,50] |
| Littorina sp. | Grazer | Yes | No | [20,50] |
| Patella intermedia | Grazer | Yes | No | [20,51] |
| Paracentrotus lividus | Grazer | Yes | No | [20,52] |
| Echinus esculentus | Grazer | Yes | No | [20,53] |
| Aplysia depilans | Grazer | No | No | [20,54] |
| Mytilus galloprovincialis | Filter feeder | Yes | Yes | [20,51] |
| Nucella lapillus | 1st level Predator | Yes | No | [20,51] |
| Marthasterias glacialis | 2nd level Predator | No | No | [20,51] |
| Charonia lampas | 3rd level Predator | Yes | No | [20,55] |
2. Results and Discussion
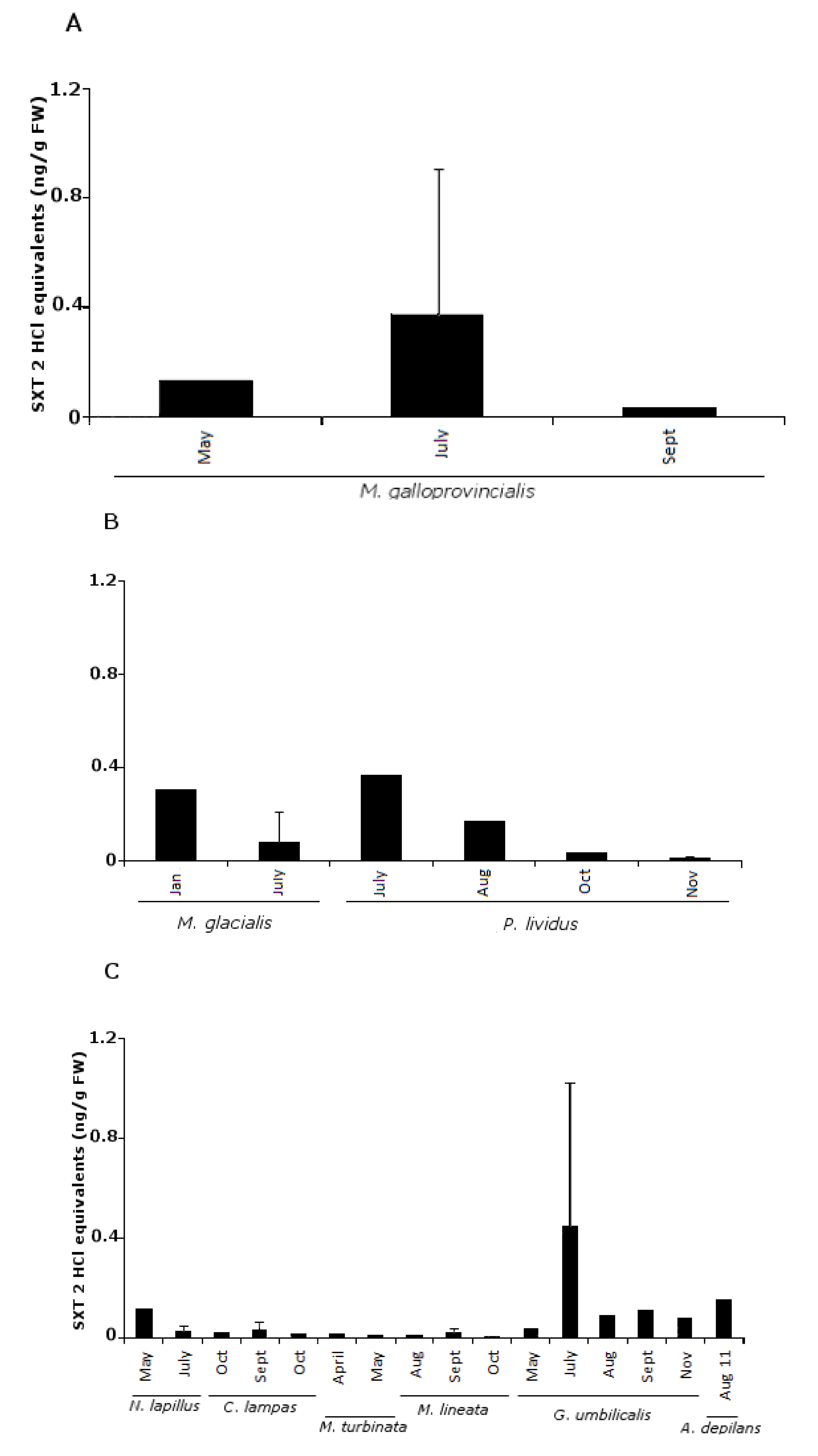
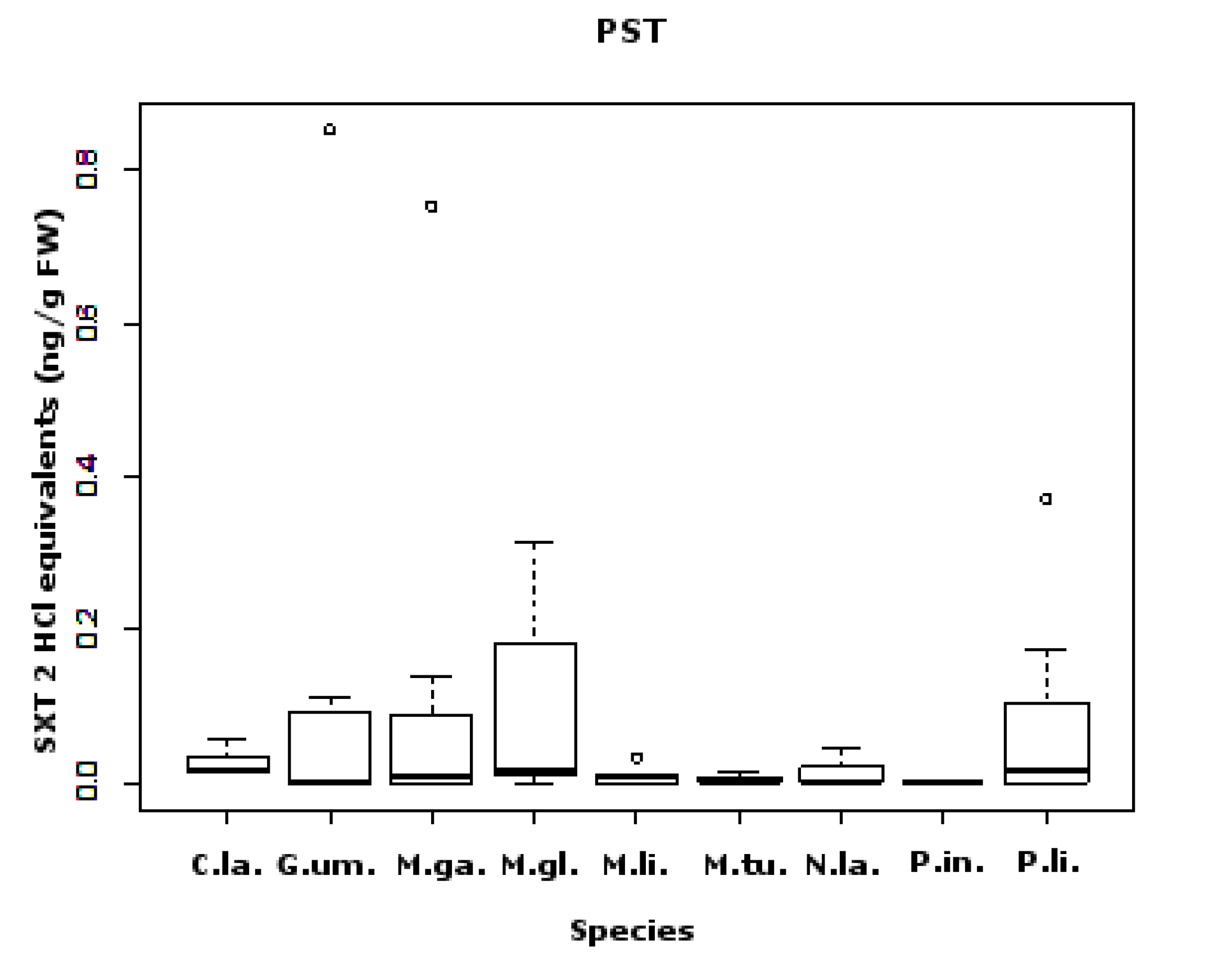
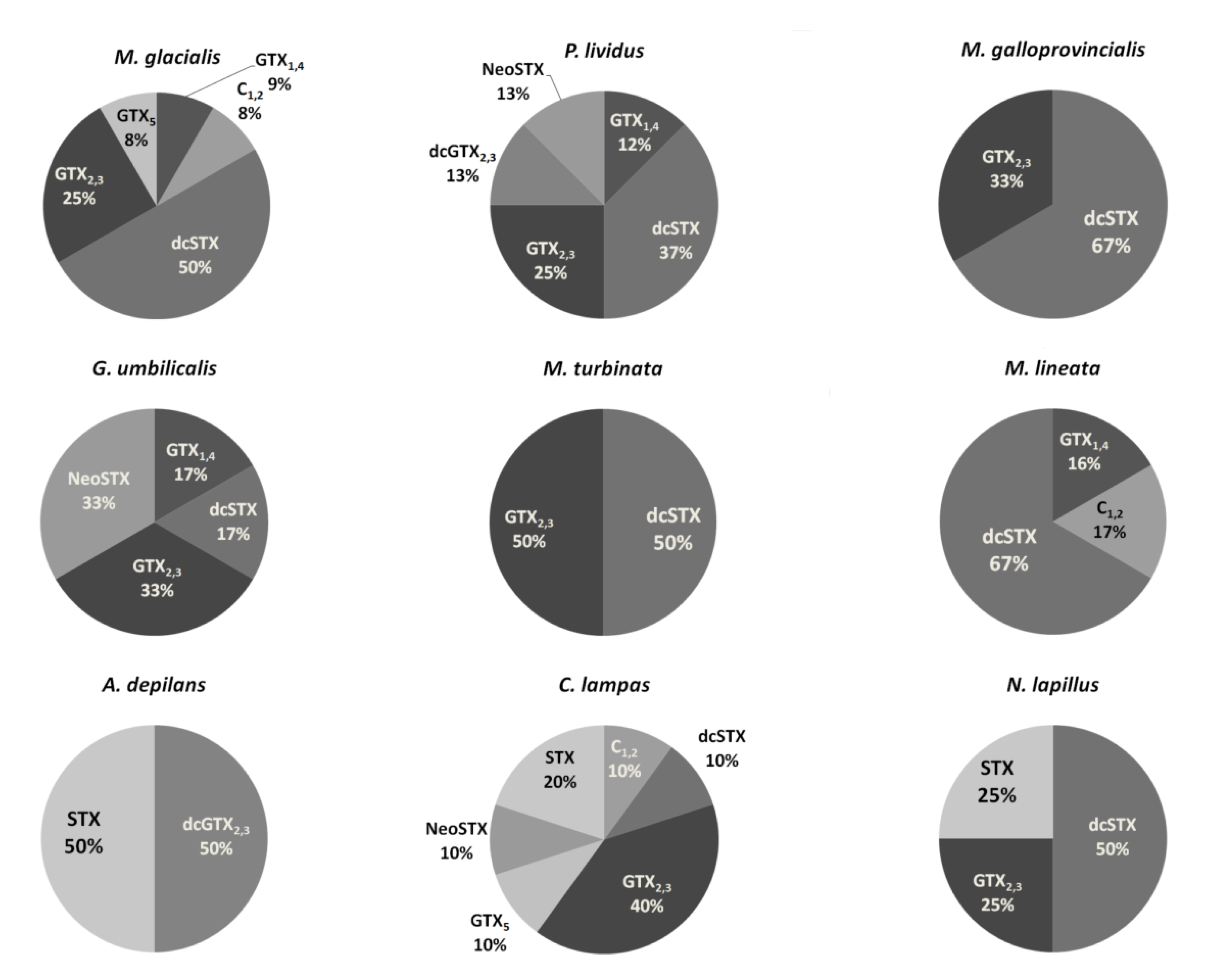
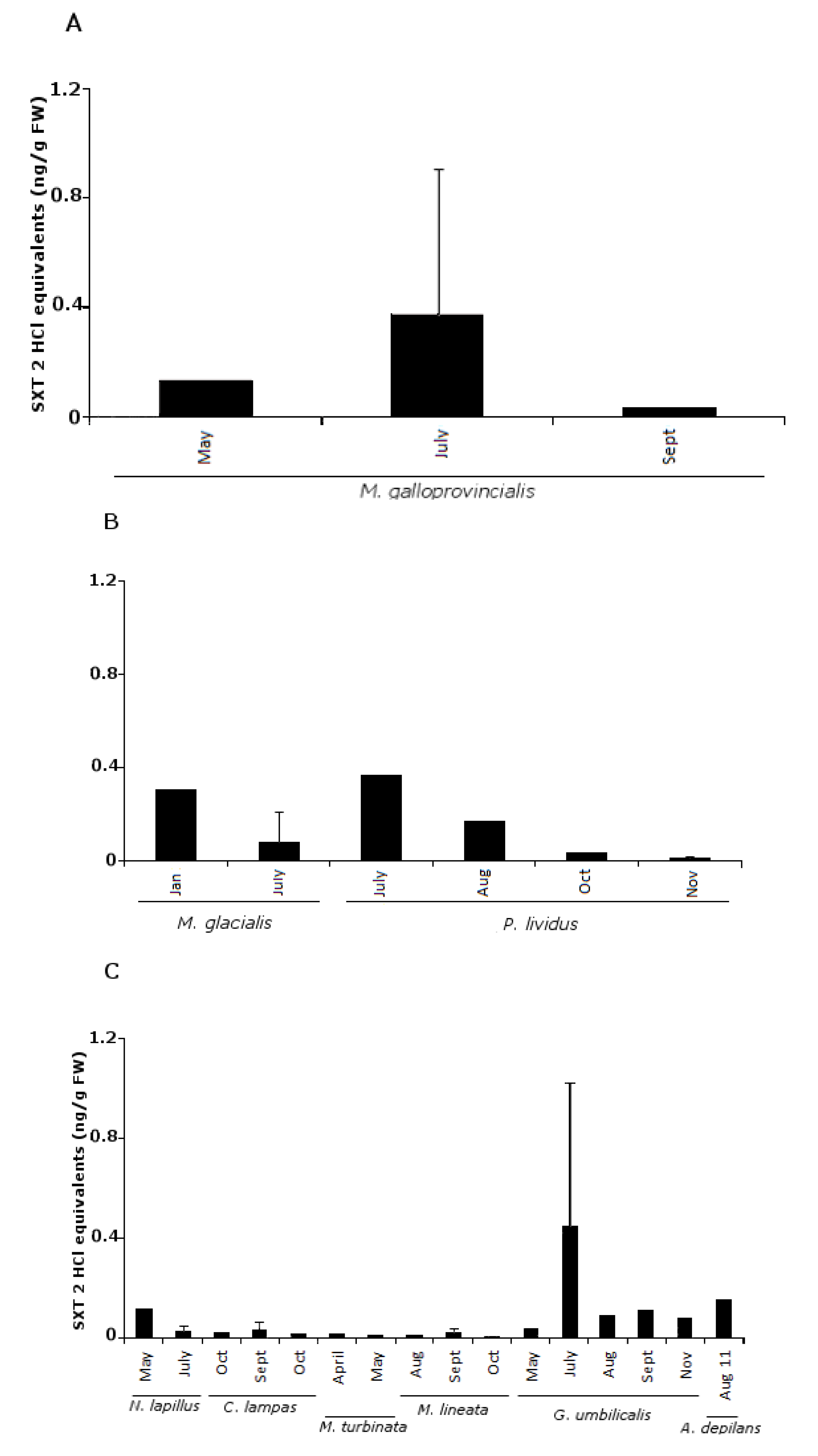
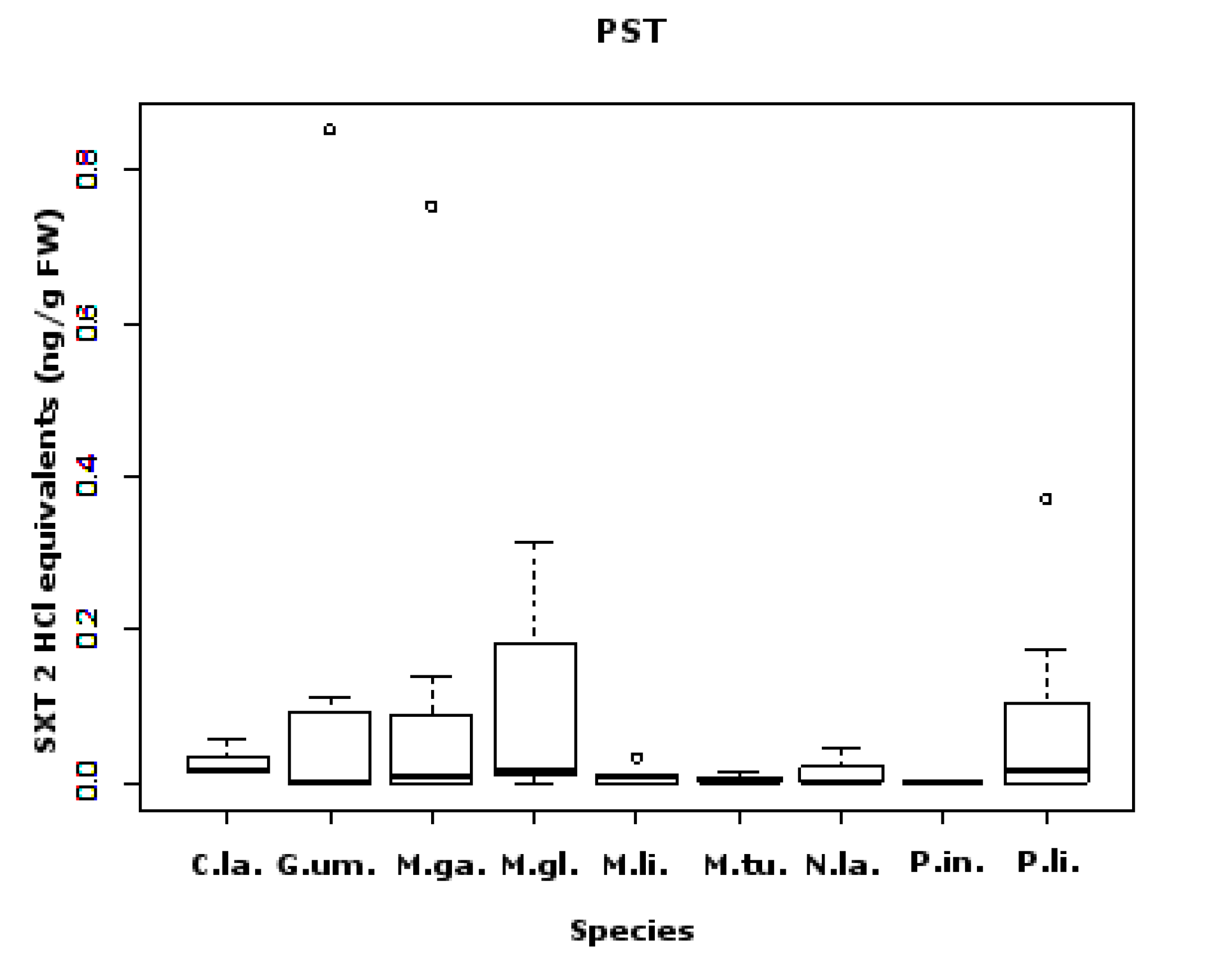
2.1. PSTs

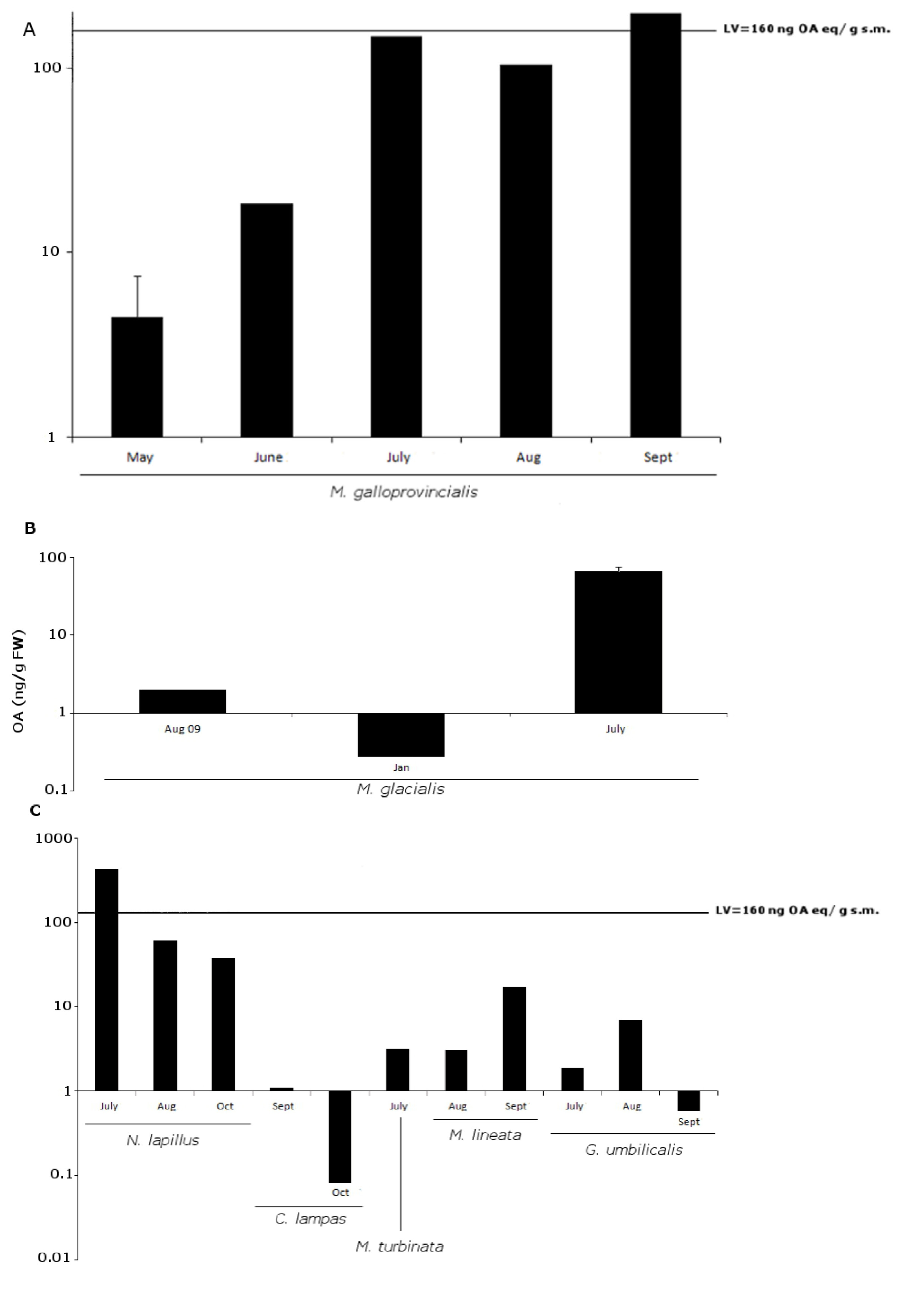
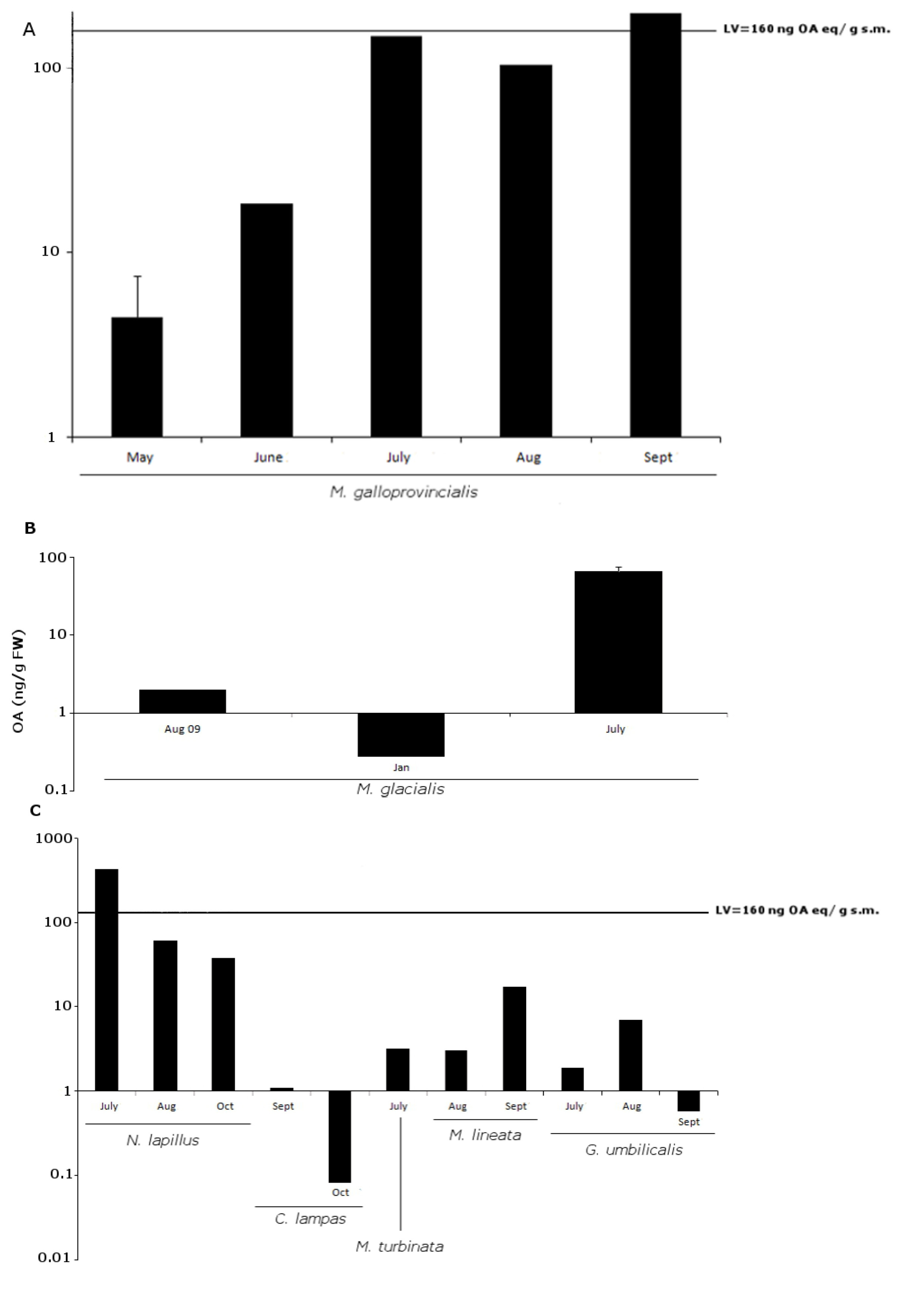
2.2. OA

| Species pairwise comparisons | z | p |
|---|---|---|
| N. lapillus–C. lampas | 3.3 | 0.03 |
| M. glacialis–G. umbilicalis | 3.8 | <0.01 |
| M. galloprovincialis–G. umbilicalis | 5.2 | <0.001 |
| N. lapillus–G. umbilicalis | 4.7 | <0.001 |
| M. lineata–M. glacialis | −3.6 | <0.01 |
| M. turbinata–M. glacialis | −3.4 | 0.02 |
| P. lividus–M. glacialis | −5.6 | <0.001 |
| P. intermedia–M. glacialis | −5.5 | <0.001 |
| M. galloprovincialis–M. lineata | 4.8 | <0.001 |
| N. lapillus–M. lineata | 4.6 | <0.001 |
| M. galloprovincialis–M. turbinata | 4.1 | <0.01 |
| N. lapillus–M. turbinata | 4.0 | <0.01 |
| P. lividus–M. galloprovincialis | −7.1 | <0.001 |
| P. intermedia–M. galloprovincialis | −6.4 | <0.001 |
| P. lividus–N. lapillus | −6.4 | <0.001 |
| P. intermedia–N. lapillus | −5.9 | <0.001 |
| P. intermedia–P. lividus | −0.6 | <0.001 |

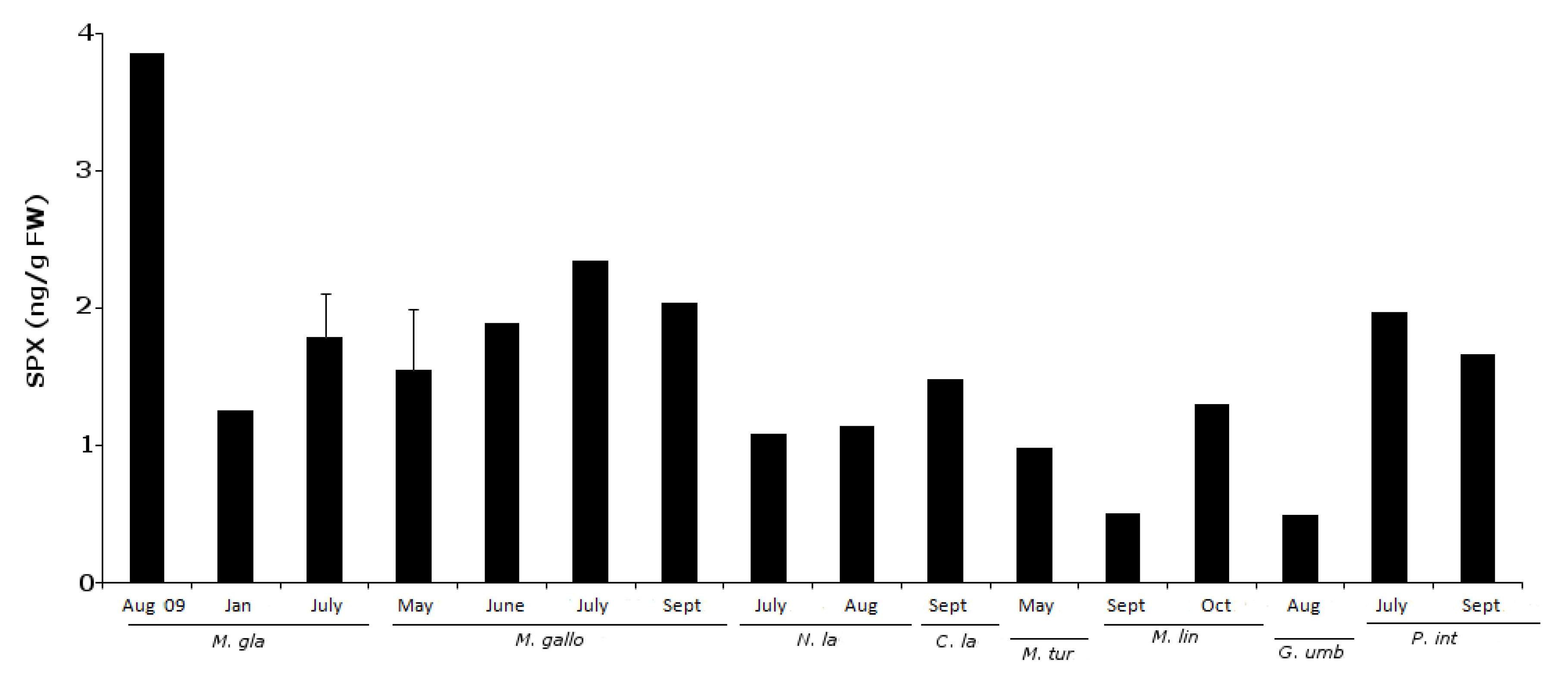
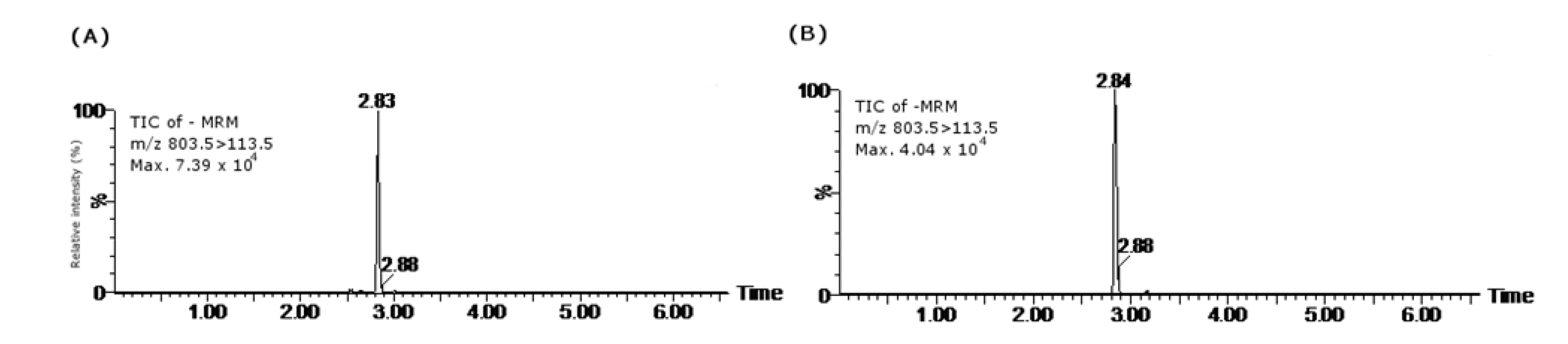
2.3. 13-Desmethyl Spirolide C
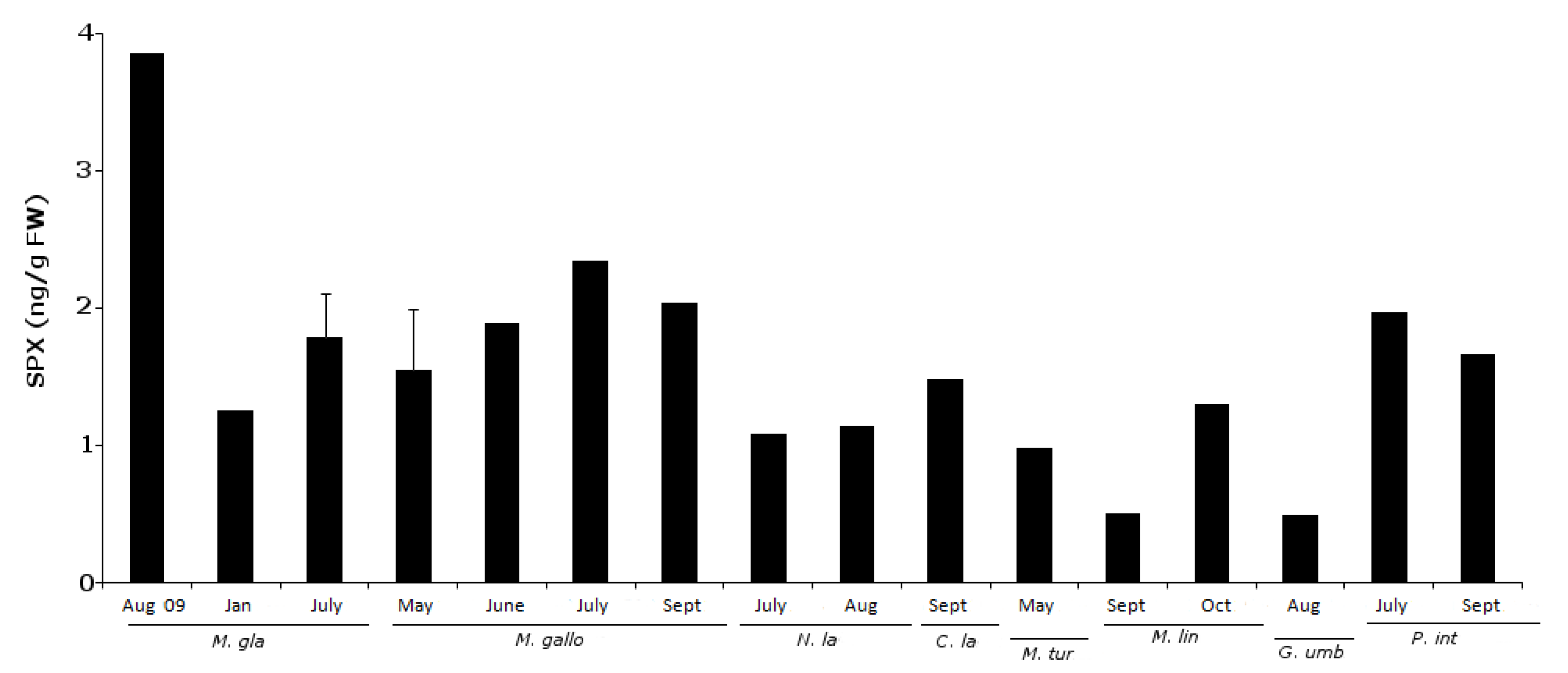
| Species pairwise comparisons | z | p |
|---|---|---|
| M. glacialis–G. umbilicalis | 4.3 | <0.001 |
| M. galloprovincialis–G. umbilicalis | 3.2 | 0.02 |
| M. lineata–M. glacialis | −3.9 | <0.01 |
| M. turbinata–M. glacialis | −3.5 | 0.01 |
| P. lividus–M. glacialis | −4.2 | <0.001 |
| P. lividus–M. galloprovincialis | −3.1 | 0.04 |
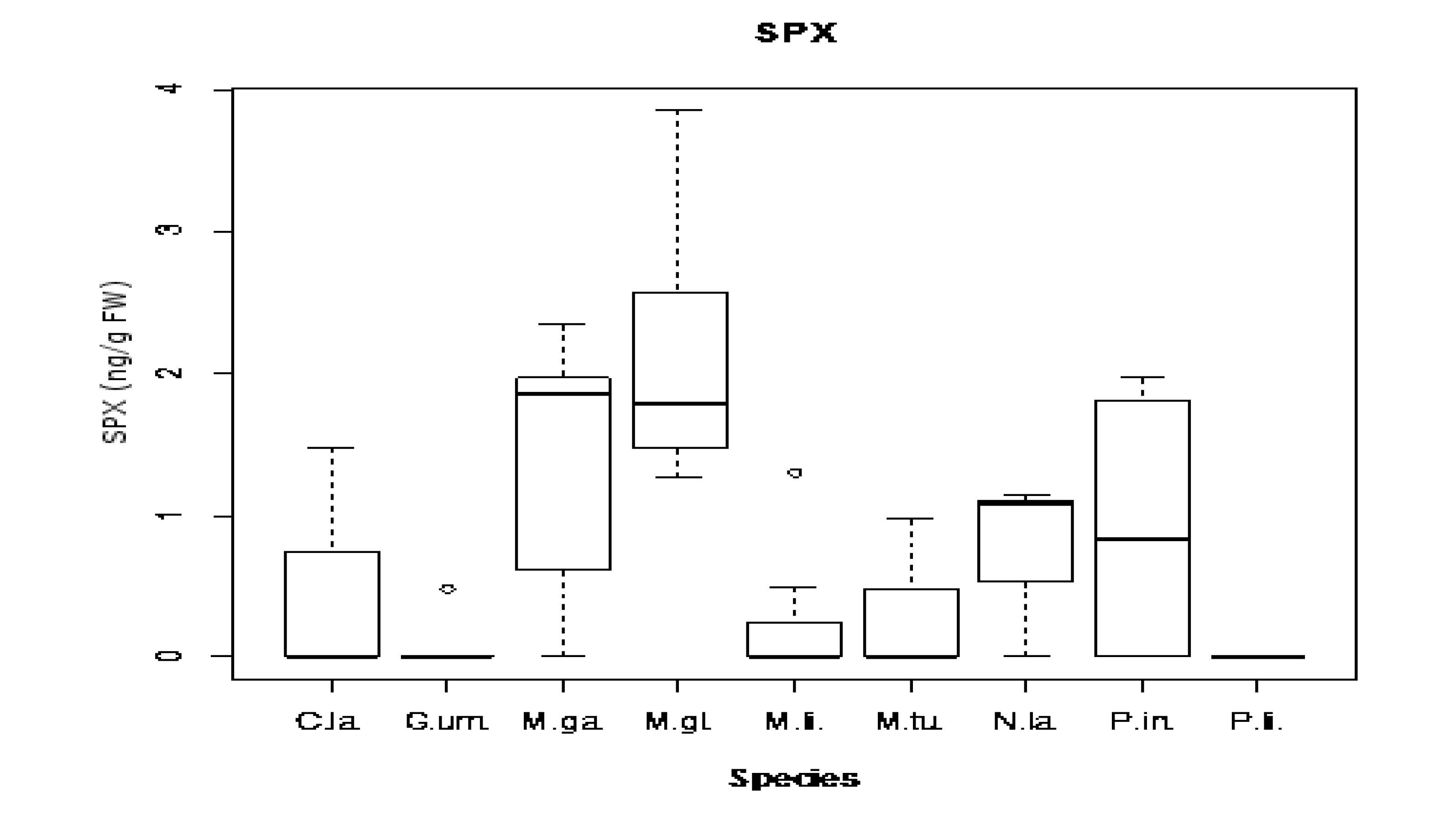
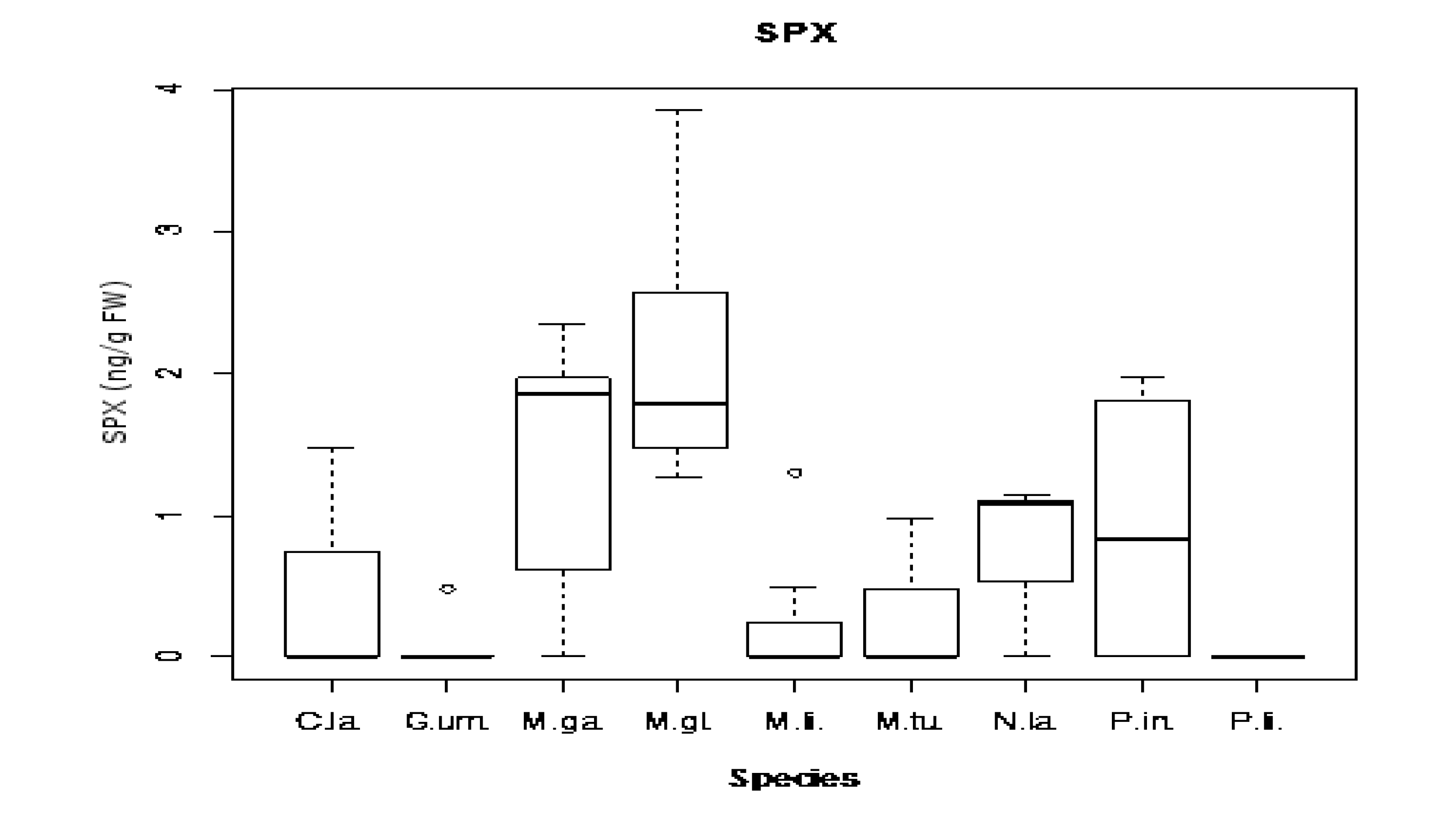
2.4. Toxins Concentrations Found: Comparison between Species and Sampling Sites
| Pairwise Comparison | p |
|---|---|
| C. lampas–P. intermedia | 0.01 |
| G. umbilicalis–M. glacialis | <0.001 |
| G. umbilicalis–M. galloprovincialis | 0.03 |
| G. umbilicalis–N. lapillus | <0.01 |
| P. intermedia–M. glacialis | <0.001 |
| P. intermedia–M. galloprovincialis | 0.02 |
| P. intermedia–N. lapillus | <0.001 |
| M. lineata–M. glacialis | 0.01 |
| M. turbinata–M. glacialis | <0.01 |
| M. galloprovincialis–P. lividus | <0.001 |
| P. lividus–M. glacialis | 0 |
| P. lividus–N. lapillus | |
| Angeiras–Memória | 0.03 |
| Memória–Porto Côvo | 0.01 |
| Memória–Viana do Castelo | 0.03 |
| Memória–Almograve | 0.01 |
3. Experimental Section
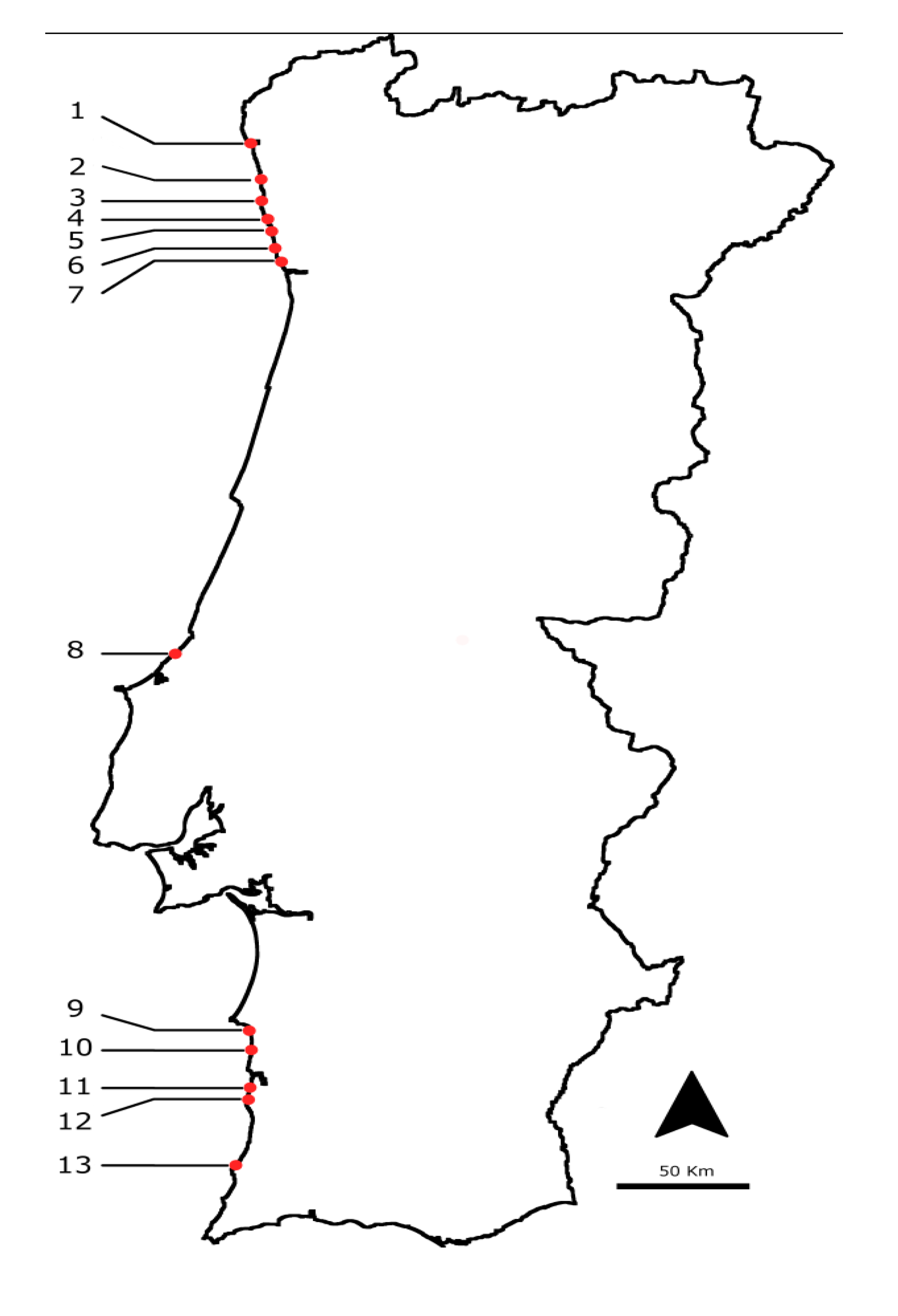
3.1. Selected Species and Sampling Points
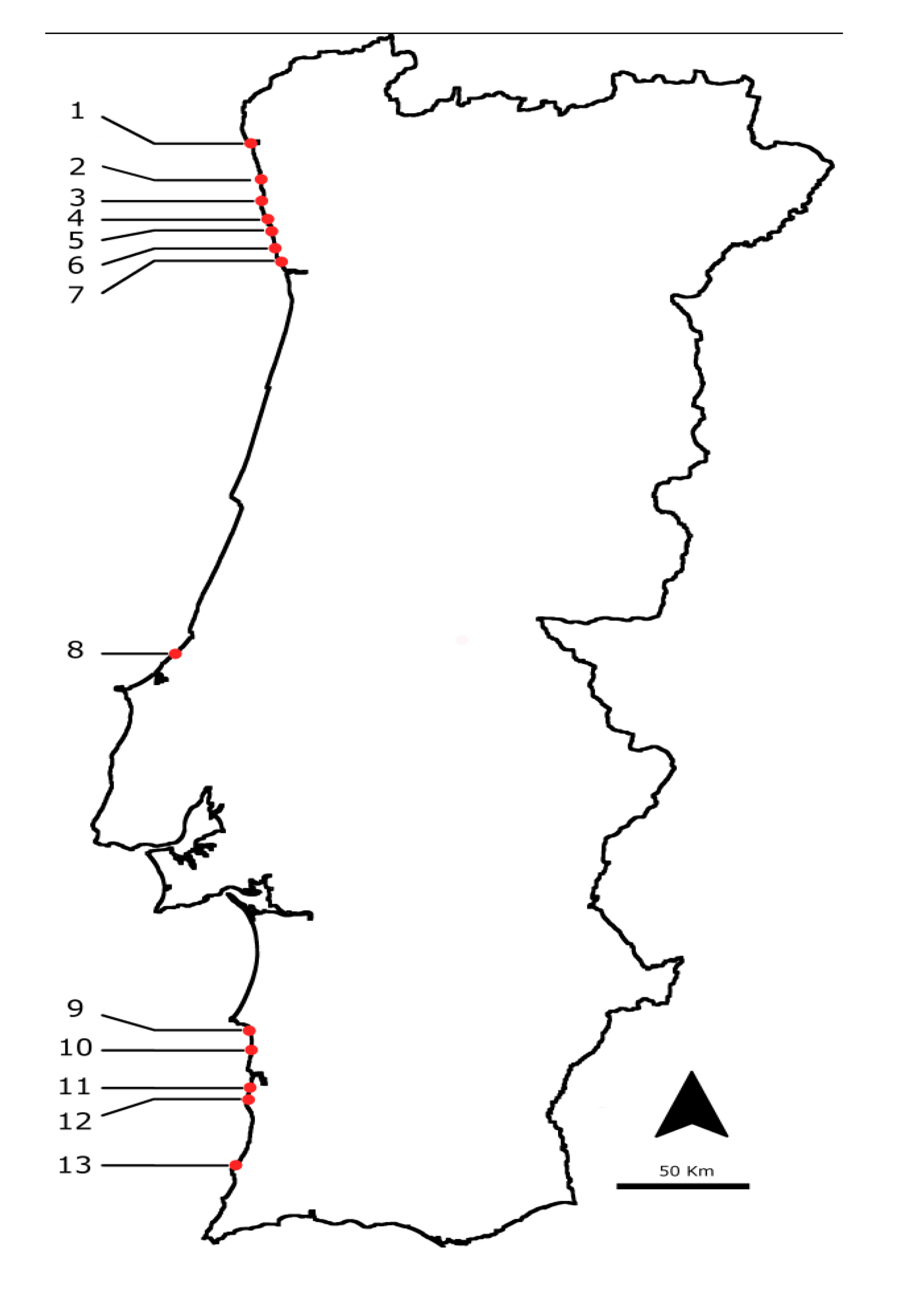
| Species | Number of pooled samples collected from July 2009 till end 2010 | Average number of animals collected to set a pooled sample |
|---|---|---|
| Gibbula umbilicalis | 34 | 100 |
| Gibbula magus | 1 | 90 |
| Monodonta lineata | 20 | 86 |
| Monodonta turbinata | 21 | 86 |
| Nucella lapillus | 13 | 15 |
| Littorina littorea | 2 | 10 |
| Littorina saxatilis | 4 | 15 |
| Patella intermedia | 4 | 15 |
| Charonia lampas | 5 | 1 |
| Mytilus galloprovincialis | 7 | 30 |
| Paracentrotus lividus | 10 | 10 |
| Marthasterias glacialis | 8 | 1 |
| Echinus esculentus | 2 | 1 |
| Aplysia depilans | 1 | 1 |
3.2. Sample Preparation
3.2.1. Paralytic Shellfish Toxin (PST) Extraction
3.2.2. Lipophilic Toxins Extraction
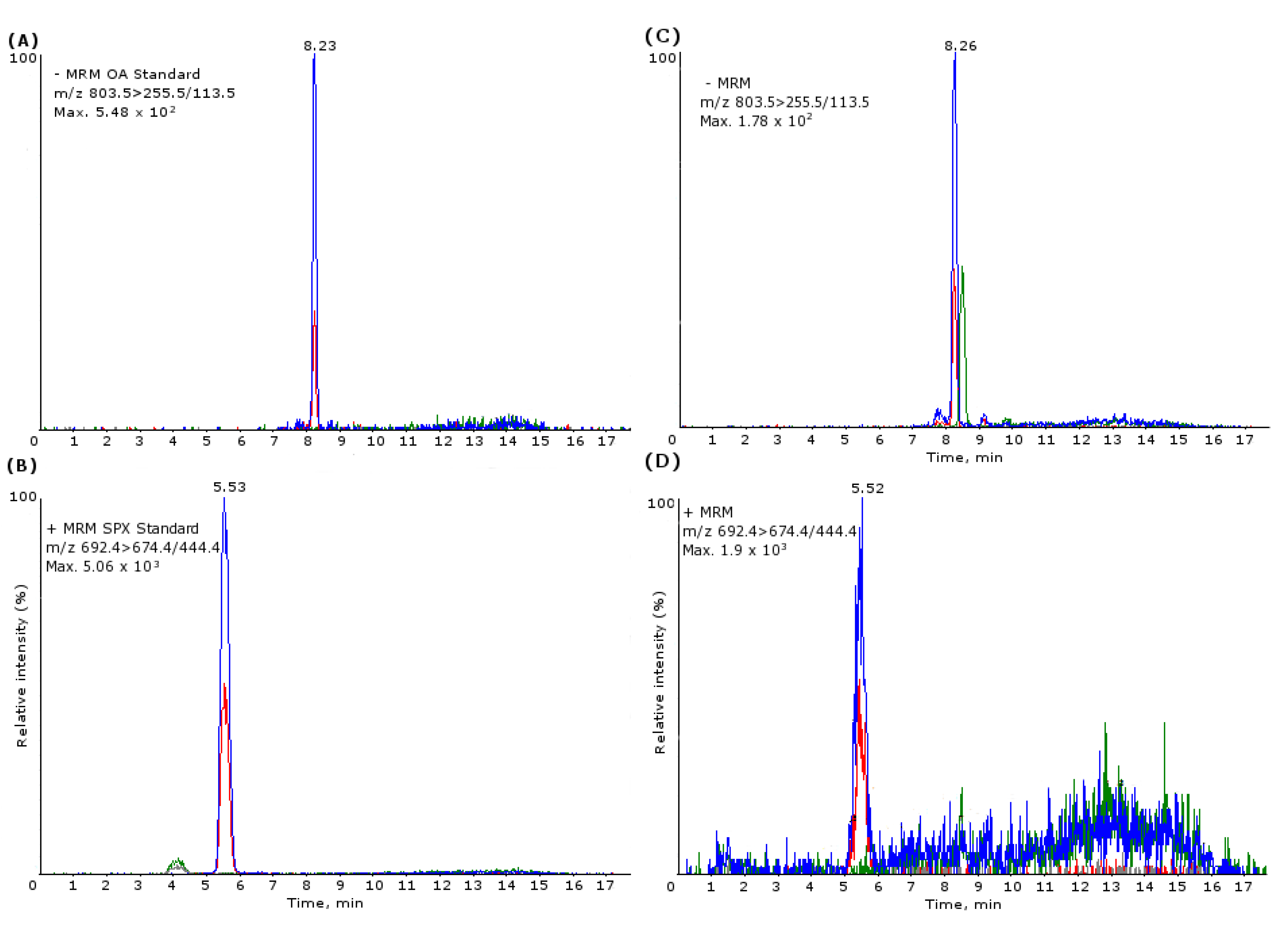
3.3. Sample Analysis
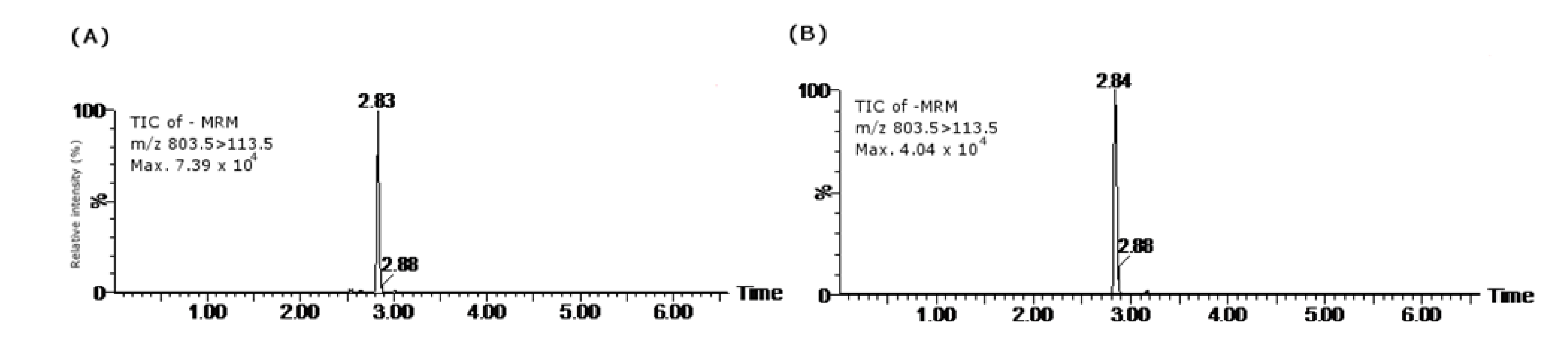
3.3.1. PSP HPLC-FLD Conditions
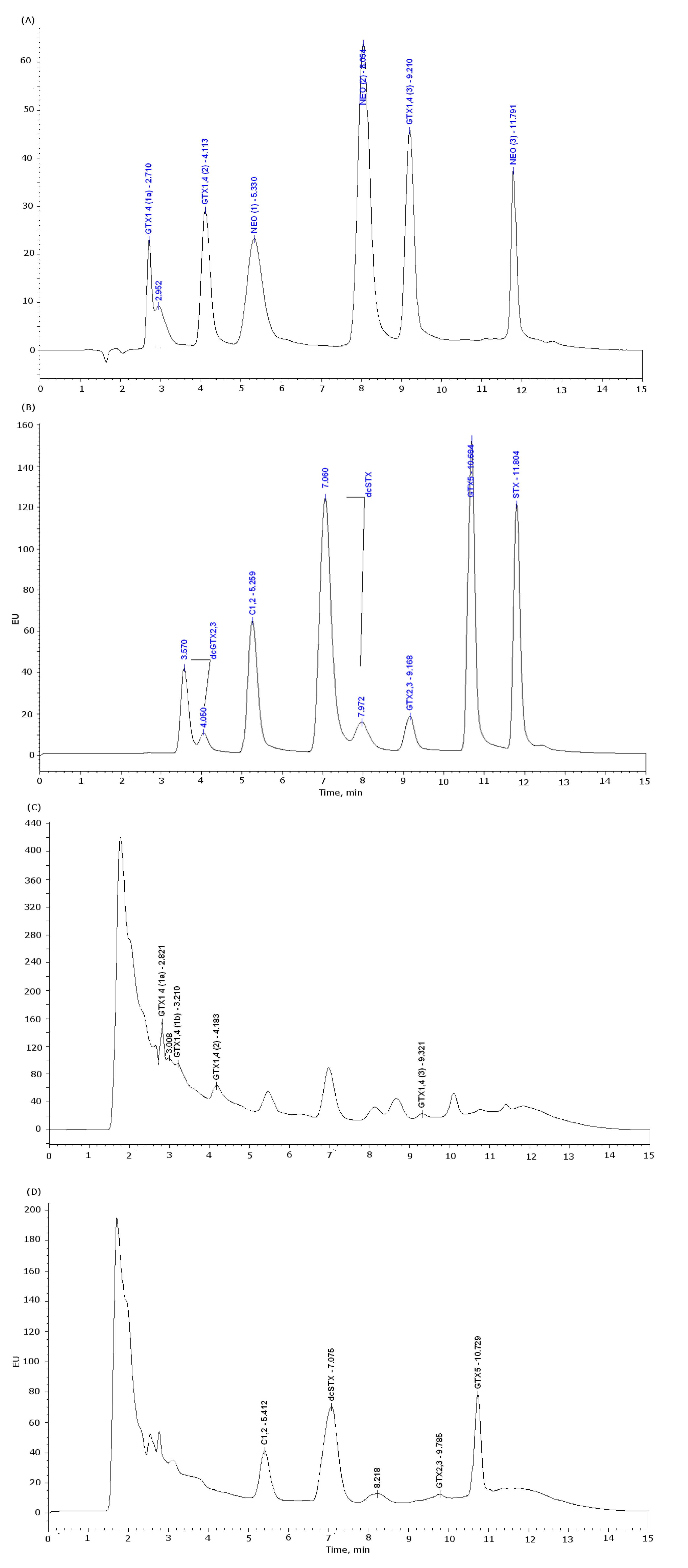
| Toxin | LOD | LOQ |
|---|---|---|
| dcGTX2,3 | 2.7 | 8 |
| C1,2 | 3 | 11 |
| dcSTX | 0.2 | 6 |
| GTX2,3 | 1.5 | 9 |
| GTX5 | 0.4 | 8 |
| SXT | 0.4 | 7 |
| GTX1,4 | 3 | 9 |
| NEO | 1.2 | 8 |
3.3.2. Lipophilic Toxins LC-MS/MS Conditions
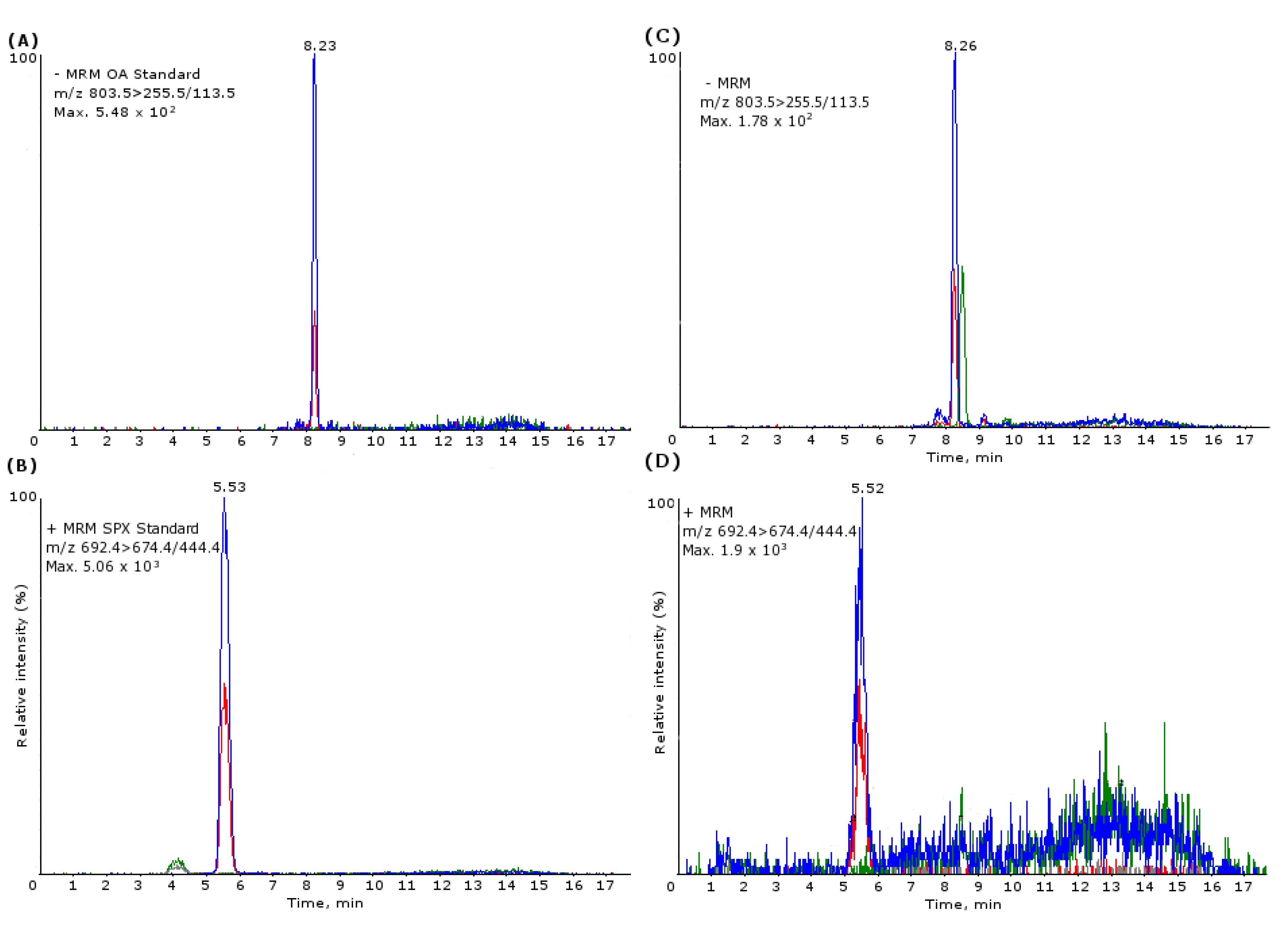
3.3.3. Lipophilic Toxin UPLC-MS/MS Conditions
3.4. Statistical Analyses
4. Conclusions
Acknowledgments
References
- Hallegraeff, G.M. Algal blooms are not a simple toxic broth. Search 1993, 24, 179. [Google Scholar]
- Glibert, P.M.; Anderson, D.; Gentien, P.; Granéli, E.; Sellner, K. The global, complex phenomena of harmful algal blooms. Oceanography 2005, 18, 136–147. [Google Scholar]
- Kat, M. The Occurence of Prorocentrum Species and Coincidental Gastrointestinal Illness of Mussel Consumers. In Toxic Dinoflagellate Blooms; Taylor, D.L., Seliger, H.H., Eds.; Elsevier: New York, NY, USA, 1979; pp. 215–220. [Google Scholar]
- Rodrigues, S.M.; de Carvalho, M.; Mestre, T.; Ferreira, J.J.; Coelho, M.; Peralta, R.; Vale, P. Paralytic shellfish poisoning due to ingestion of Gymnodinium catenatum contaminated cockles—Application of the AOAC HPLC official method. Toxicon 2012, 59, 558–566. [Google Scholar] [CrossRef]
- Meyer, K.F.; Sommer, H.; Schoenholz, P. Mussel poisoning. Am. J. Prev. Med. 1928, 2, 365–394. [Google Scholar]
- Steward, I.; Seawright, A.A.; Shaw, G.R. Cyanobacterial Poisoning in Livestock, Wild Mammals and Birds—An Overview. In Cyanobacterial Harmful Algal Blooms: State of Science and Research Needs; Hudnell, H.K., Ed.; Springer: New York, NY, USA, 2008; pp. 613–637. [Google Scholar]
- Vale, P.; Antonia, M.; Sampayo, M. Esters of okadaic acid and dinophysistoxin-2 in Portuguese bivalves related to human poisonings. Toxicon 1999, 37, 1109–1121. [Google Scholar] [CrossRef]
- Gerssen, A.; Pol-Hofstad, I.E.; Poelman, M.; Mulder, P.P.; van den Top, H.J.; de Boer, J. Marine toxins: Chemistry, toxicity, occurrence and detection, with special reference to the Dutch situation. Toxins 2010, 2, 878–904. [Google Scholar]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef]
- Gainey, L.S.J.; Shumway, S. A compendium of the responses of bivalve molluscs to toxic dinoflagellates. J. Shellfish Res. 1988, 7, 623–628. [Google Scholar]
- Shumway, S.E. Phycotoxin-related shellfish poisoning: Bivalve molluscs are not the only vectors. Rev. Fish. Sci. 1995, 3, 1–31. [Google Scholar] [CrossRef]
- Correia, F.S. Um caso raro de intoxicação alimentar colectiva. Bol. Inst. Super. Hig. Doutor Ricardo Jorge 1946, 3, 216–221. [Google Scholar]
- Vale, P.; Botelho, M.J.; Rodrigues, S.M.; Gomes, S.S.; Sampayo, M.A.D. Two decades of marine biotoxin monitoring in bivalves from Portugal (1986–2006): A review of exposure assessment. Harmful Algae 2008, 7, 11–25. [Google Scholar] [CrossRef]
- Alexander, J.; Benford, D.; Cockburn, A.; Cravedi, J.P.; Dogliotti, E.; Di Domenico, A.; Fernández-Cruz, M.L.; Fink-Gremmels, J.; Fürst, P.; Galli, C.; et al. Scientific opinion of the panel on contaminants in the food chain on a request from the European commission on marine biotoxins in shellfish—Saxitoxin Group. EFSA J. 2009, 1019, 1–76. [Google Scholar]
- Codd, G.A. Cyanobacterial toxins: Occurrence, properties and biological significance. Water Sci. Technol. 1995, 32, 149–156. [Google Scholar]
- Clemente, Z.; Busato, R.H.; Oliveira Ribeiro, C.A.; Cestari, M.M.; Ramsdorf, W.A.; Magalhaes, V.F.; Wosiack, A.C.; Silva de Assis, H.C. Analyses of paralytic shellfish toxins and biomarkers in a southern Brazilian reservoir. Toxicon 2010, 55, 396–406. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Evans, W.R.; Yin, Q.Q.; Bell, P.; Moczydlowski, E. Evidence for paralytic shellfish poisons in the freshwater cyanobacterium Lyngbya wollei (Farlow ex Gomont) comb. nov. Appl. Environ. Microbiol. 1997, 63, 3104–3110. [Google Scholar]
- Ballot, A.; Fastner, J.; Wiedner, C. Paralytic shellfish poisoning toxin-producing cyanobacterium Aphanizomenon gracile in northeast Germany. Appl. Environ. Microbiol. 2010, 76, 1173–1180. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxin Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; WHO E & FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- European Commission. Regulation (EC) No. 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Available online: http://eur-lex.europa.eu (27 December 2010).
- Mackinnon, S.L.; Cembella, A.D.; Burton, I.W.; Lewis, N.; LeBlanc, P.; Walter, J.A. Biosyntheis of 13-desmethyl spirolid C by the dinoflagellate Alexandrium ostenfeldii. J. Org. Chem. 2006, 71, 8724–8731. [Google Scholar] [CrossRef]
- MacKinnon, S.L.; Walter, J.A.; Quilliam, M.A.; Cembella, A.D.; LeBlanc, P.; Burton, I.W.; Hardstaff, W.R.; Lewis, N.I. Spirolides isolated of the danish strains of the toxigenic dinoflagellate Alexandrium ostenfeldii. J. Nat. Prod. 2006, 69, 983–987. [Google Scholar] [CrossRef]
- Munday, R. Toxicology of Cyclic Imines: Gymnodimine, Spirolides, Pinnatoxins, Pteriatoxins, Prorocentrolide, Spiro-Prorocentrimine, and Symbioimines. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press (Taylor and Francys Group): Boca Raton, FL, USA, 2008; pp. 581–594. [Google Scholar]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; van Ginkel, R.; Holland, P.; Munday, R. Production of pinnatoxins by a peridinoid dinoflagellate isolated from Northland, New Zealand. Harmful Algae 2010, 9, 384–389. [Google Scholar] [CrossRef]
- Hu, T.; Curtis, J.M.; Walter, J.A.; Wright, J.L.C. Characterization of biologically inactive spirolides E and F: Identification of the spirolide pharmacophore. Tetrahedron Lett. 1996, 37, 7671–7674. [Google Scholar]
- Hu, T.; Curtis, J.M.; Oshima, Y.; Quilliam, M.A.; Walter, J.A.; Watson-Wright, W.M.; Wright, J.L.C. Spirolides B and D, two novel macrocycles isolated from the digestive glands of shellfish. J. Chem. Soc. Chem. Commun. 1995, 20, 2159–2161. [Google Scholar]
- Hu, T.; Burton, I.W.; Cembella, A.D.; Curtis, J.M.; Quilliam, M.A.; Walter, J.A.; Wright, J.L. Characterization of spirolides a, c, and 13-desmethyl c, new marine toxins isolated from toxic plankton and contaminated shellfish. J. Nat. Prod. 2001, 64, 308–312. [Google Scholar] [CrossRef]
- John, U.; Cembella, A.D.; Hummert, C.; Elbrächter, M.; Groben, R.; Medlin, L.K. Discrimination of the toxigenic dinoflagellate species Alexandrium tamarense and Alexandrium ostenfeldii in co-occuring natural populations from Scottish coastal waters. Eur. J. Phycol. 2003, 38, 25–40. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Keafer, B.A.; Gribble, K.E.; Marin, R.; Scholin, C.A. Identification and enumeration of Alexandrium spp. from the Gulf of Maine using molecular probes. Deep Sea Res. II 2005, 54, 2467–2490. [Google Scholar]
- Aasen, J.A.B.; MacKinnon, S.L.; LeBlanc, P.; Walter, J.A.; Hovgaard, P.; Aune, T.; Quilliam, M.A. Detection and identification of spirolides in Norwegian shellfish and plankton. Chem. Res. Toxicol. 2005, 18, 509–515. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Magno, S.; Tartaglione, L.; Cangini, M.; Pompei, M.; Guerrini, F.; Boni, L.; Pistocchi, R. Toxin profile of Alexandrium ostenfeldii (Dinophyceae) from the Northern Adriatic Sea revealed by liquid chromatography-mass spectometry. Toxicon 2006, 47, 597–604. [Google Scholar] [CrossRef]
- Amzil, Z.; Sibat, M.; Royer, F.; Masson, N.; Abadie, E. Report on the first detection of pectenotoxin-2, spirolide a and their derivatives in French shellfish. Mar. Drugs 2007, 5, 168–179. [Google Scholar] [CrossRef]
- Villar Gonzalez, A.; Rodriguez-Velasco, M.L.; Ben-Gigirey, B.; Botana, L.M. First evidence of spirolides in Spanish shellfish. Toxicon 2010, 48, 1068–1074. [Google Scholar]
- Álvarez, G.; Uribe, E.; Ávalos, P.; Mariño, C.; Blanco, J. First identification of azaspiracid and spirolides in Mesodesma donacium and Mulinia edulis from Northern Chile. Toxicon 2010, 55, 638–641. [Google Scholar] [CrossRef]
- Seki, T.; Satake, M.; Mackenzie, A.L.; Kaspar, H.F.; Yasumoto, T. Gimnodimine, a new marine toxin of unprecedented structure isolated from New Zealand oysters and the dinoflagellate, Gymnodinium sp. Tetrahedron Lett. 1995, 36, 7093–7096. [Google Scholar]
- Touzet, N.; Franco, J.M.; Raine, R. Morphogenetic diversity and biotoxin composition of Alexandrium (Dinophyceae) in Irish coastal waters. Harmful Algae 2008, 7, 782–797. [Google Scholar] [CrossRef]
- Otero, P.; Alfonso, A.; Alfonso, C.; Araoz, R.; Molgo, J.; Vieytes, M.R.; Botana, L.M. First direct fluorescence polarization assay for the detection and quantification of spirolides in mussel samples. Anal. Chim. Acta 2011, 701, 200–208. [Google Scholar] [CrossRef]
- Alexander, J.; Benford, D.; Boobis, A.; Ceccatelli, S.; Cravedi, J.P.; di Domenico, A.; Doerge, D.; Dogliotti, E.; Edler, L.; Farmer, P.; et al. Scientific Opinion on marine biotoxins in shellfish—Cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8, 1628–1887. [Google Scholar]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine sponge toxin, okadaic acid, on protein phosphatases. Biochem. J. 1988, 256, 283–290. [Google Scholar]
- Maneiro, E.; Rodas, V.L.; Costas, E.; Hernandez, J.M. Shellfish consumption: A major risk factor for colorectal cancer. Med. Hypotheses 2008, 70, 409–412. [Google Scholar] [CrossRef]
- Elgarch, A.; Vale, P.; Rifai, S.; Fassouane, A. Detection of diarrheic shellfish poisoning and azaspiracid toxins in moroccan mussels: Comparison of the LC-MS method with the commercial immunoassay kit. Mar. Drugs 2008, 6, 587–594. [Google Scholar] [CrossRef]
- Garcia, C.; Truan, D.; Lagos, M.; Santelices, J.P.; Diaz, J.C.; Lagos, N. Metabolic transformation of dinophysistoxin-3 into dinophysistoxin-1 causes human intoxication by consumption of O-acyl-derivatives dinophysistoxins contaminated shellfish. J. Toxicol. Sci. 2005, 30, 287–296. [Google Scholar] [CrossRef]
- Scoging, A.; Bahl, M. Diarrhetic shellfish poisoning in the UK. Lancet 1998, 352, 117. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in tohoku district. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- MacKenzie, L.; Beuzenberg, V.; Holland, P.; McNabb, P.; Suzuki, T.; Selwood, A. Pectenotoxin and okadaic acid-based toxin profiles in Dinophysis acuta and Dinophysis acuminata from New Zealand. Harmful Algae 2005, 4, 75–85. [Google Scholar] [CrossRef]
- Hu, T.M.; Curtis, J.M.; Walter, J.A.; Wright, J.L.C. Identification of DTX-4, a new water-soluble phosphatase inhibitor from the toxic dinoflagellate Prorocentrum lima. J. Chem. Soc. Chem. Comm. 1995, 5, 597–599. [Google Scholar]
- Caroppo, C.; Congestri, R.; Bruno, M. On the presence of Phalacroma rotundatum in the southern Adriatic Sea (Italy). Aquat. Microb. Ecol. 1999, 17, 301–310. [Google Scholar] [CrossRef]
- Jorgensen, K.; Andersen, P. Relation between the concentration of Dinophysis acuminata and diarrheic shellfish poisoning toxins in blue mussels (Mytilus edulis) during a toxic episode in the Limfjord (Denmark), 2006. J. Shellfish Res. 2007, 26, 1081–1087. [Google Scholar] [CrossRef]
- Spatharis, S.; Dolapsakis, N.P.; Economou-Amilli, A.; Tsirtsis, G.; Danielidis, D.B. Dynamics of potentially harmful microalgae in a confined Mediterranean Gulf—Assessing the risk of bloom formation. Harmful Algae 2009, 8, 736–743. [Google Scholar] [CrossRef]
- Crothers, J.H. Common topshells: An introduction to the biology of Osilinus lineatus with notes on other species in the genus. Field Stud. 2001, 10, 115–160. [Google Scholar]
- Knox, G.A. Hard Shores. In The Ecology of Seashores; Kennish, M.J., Ed.; CRC Press: Boca Raton, FL, USA, 2001; pp. 20–86. [Google Scholar]
- Lemée, R.; Boudouresque, C.F.; Gobert, J.; Malestroit, P.; Mari, X.; Meinesz, A.; Menager, V.; Ruitton, S. Feeding behaviour of Paracentrotus lividus in the presence of Caulerpa taxifolia introduced in the Mediterranean Sea. Oceanol. Acta 1995, 19, 245–253. [Google Scholar]
- Forster, G.R. The ecology of Echinus esculentus L. Quantitative distribution and rate of feeding. J. Mar. Biol. Assoc. UK 1959, 38, 361–367. [Google Scholar] [CrossRef]
- Carefoot, T.H. Aplysia: Its biology and ecology. Oceanogr. Mar. Biol. Annu. Rev. 1987, 25, 167–284. [Google Scholar]
- Lin, S.J.; Hwang, D.F. Possible source of tetrodotoxin in the starfish Astropecten scoparius. Toxicon 2001, 39, 573–579. [Google Scholar]
- Vale, P. Hydrolysis of hydroxybenzoate saxitoxin analogues originating from Gymnodium catenatum. Food Chem. 2011, 125, 1160–1165. [Google Scholar] [CrossRef]
- Ogata, T.; Kodama, M.; Fukuyo, Y.; Inoue, T.; Kamiya, H.; Matsuura, F.; Sekiguchi, K.; Watanabe, S. Studies on paralytic shellfish poisoning in ofunato bay. 1. The occurrence of protogonyaulax spp in Ofunato Bay, in association with the toxification of the scallop Patinopecten yessoensis. Bull. Jpn. Soc. Sci. Fish. 1982, 48, 563–566. [Google Scholar]
- Montojo, U.M.; Sakamoto, S.; Cayme, M.F.; Gatdula, N.C.; Furio, E.F.; Relox, J.R.; Sato, S.; Fukuyo, Y.; Kodama, M. Remarkable difference in accumulation of paralytic shellfish poisoning toxins among bivalve species exposed to Pyrodinium bahamense var. compressum bloom in Masinloc bay, Philippines. Toxicon 2006, 48, 85–92. [Google Scholar]
- Sekiguchi, K.; Sato, S.; Kaga, S.; Ogata, T.; Kodama, M. Accumulation of paralytic shellfish poisoning toxins in bivalves and an ascidian fed on Alexandrium tamarense cells. Fish. Sci. 2001, 67, 301–305. [Google Scholar] [CrossRef]
- Asakawa, M.; Takagi, M.; Iida, A.; Oishi, K. Studies on the conversion of paralytic shellfish poison (Psp) components by biochemical reducing agents. Eisei Kagaku 1987, 33, 50–55. [Google Scholar] [CrossRef]
- Oshima, Y. Chemical and Enzymatic Transformation of Paralytic Shellfish Toxins in Marine Organisms. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier Intercept Ltd.: Paris, France, 1995; p. 475. [Google Scholar]
- Noguchi, T. Marine toxins. Nippon Suisan Gakkaishi 2003, 69, 895–909. [Google Scholar]
- Nagashima, Y.; Arakawa, O.; Shiomi, K.; Noguchi, T. Paralytic Shellfish Toxins in a Trumpet Shell. Charonia lampas from Spain. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; Intergovernmental Oceanic Comission of UNESCO: Sendai, Japan, 1996; pp. 425–427. [Google Scholar]
- Asakawa, M.; Nishimura, F.; Miyazawa, K.; Noguchi, T. Occurrence of paralytic shellfish poison in the starfish Asterias amurensis in Kure Bay, Hiroshima Prefecture, Japan. Toxicon 1997, 35, 1081–1087. [Google Scholar] [CrossRef]
- Lin, S.J.; Tsai, Y.H.; Lin, H.P.; Hwang, D.F. Paralytic toxins in Taiwanese starfish Astropecten scoparius. Toxicon 1998, 36, 799–803. [Google Scholar] [CrossRef]
- Brusca, R.C.; Brusca, G.J. Invertebrates, 2nd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2002. [Google Scholar]
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: Collaborative study. J. AOAC Int. 2005, 88, 1714–1732. [Google Scholar]
- Otero, P.; Alfonso, A.; Alfonso, C.; Vieytes, M.R.; Louzao, M.C.; Botana, A.M.; Botana, L.M. New protocol to obtain spirolides from Alexandrium ostenfeldii cultures with high recovery and purity. Biomed. Chromatogr. 2010, 24, 878–886. [Google Scholar]
- Rodriguez, P.; Alfonso, A.; Botana, A.M.; Vieytes, M.R.; Botana, L.M. Comparative analysis of pre- and post-column oxidation methods for detection of paralytic shellfish toxins. Toxicon 2010, 56, 448–457. [Google Scholar] [CrossRef]
- Otero, P.; Alfonso, A.; Alfonso, C.; Rodriguez, P.; Vieytes, M.R.; Botana, L.M. Effect of uncontrolled factors in a validated liquid chromatography-tandem mass spectrometry method question its use as a reference method for marine toxins: Major causes for concern. Anal. Chem. 2011, 83, 5903–5911. [Google Scholar] [CrossRef]
- Guisande, C.G.; Vaamonde, A.L.; Barreiro, A. Tratamiento de datos con R, Statistica y SPSS; Díaz de Santos: Madrid, Spain, 2011; p. 978. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Silva, M.; Barreiro, A.; Rodriguez, P.; Otero, P.; Azevedo, J.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New Invertebrate Vectors for PST, Spirolides and Okadaic Acid in the North Atlantic. Mar. Drugs 2013, 11, 1936-1960. https://doi.org/10.3390/md11061936
Silva M, Barreiro A, Rodriguez P, Otero P, Azevedo J, Alfonso A, Botana LM, Vasconcelos V. New Invertebrate Vectors for PST, Spirolides and Okadaic Acid in the North Atlantic. Marine Drugs. 2013; 11(6):1936-1960. https://doi.org/10.3390/md11061936
Chicago/Turabian StyleSilva, Marisa, Aldo Barreiro, Paula Rodriguez, Paz Otero, Joana Azevedo, Amparo Alfonso, Luis M. Botana, and Vitor Vasconcelos. 2013. "New Invertebrate Vectors for PST, Spirolides and Okadaic Acid in the North Atlantic" Marine Drugs 11, no. 6: 1936-1960. https://doi.org/10.3390/md11061936
APA StyleSilva, M., Barreiro, A., Rodriguez, P., Otero, P., Azevedo, J., Alfonso, A., Botana, L. M., & Vasconcelos, V. (2013). New Invertebrate Vectors for PST, Spirolides and Okadaic Acid in the North Atlantic. Marine Drugs, 11(6), 1936-1960. https://doi.org/10.3390/md11061936