Scavenging Capacity of Marine Carotenoids against Reactive Oxygen and Nitrogen Species in a Membrane-Mimicking System

Abstract
:1. Introduction
2. Results
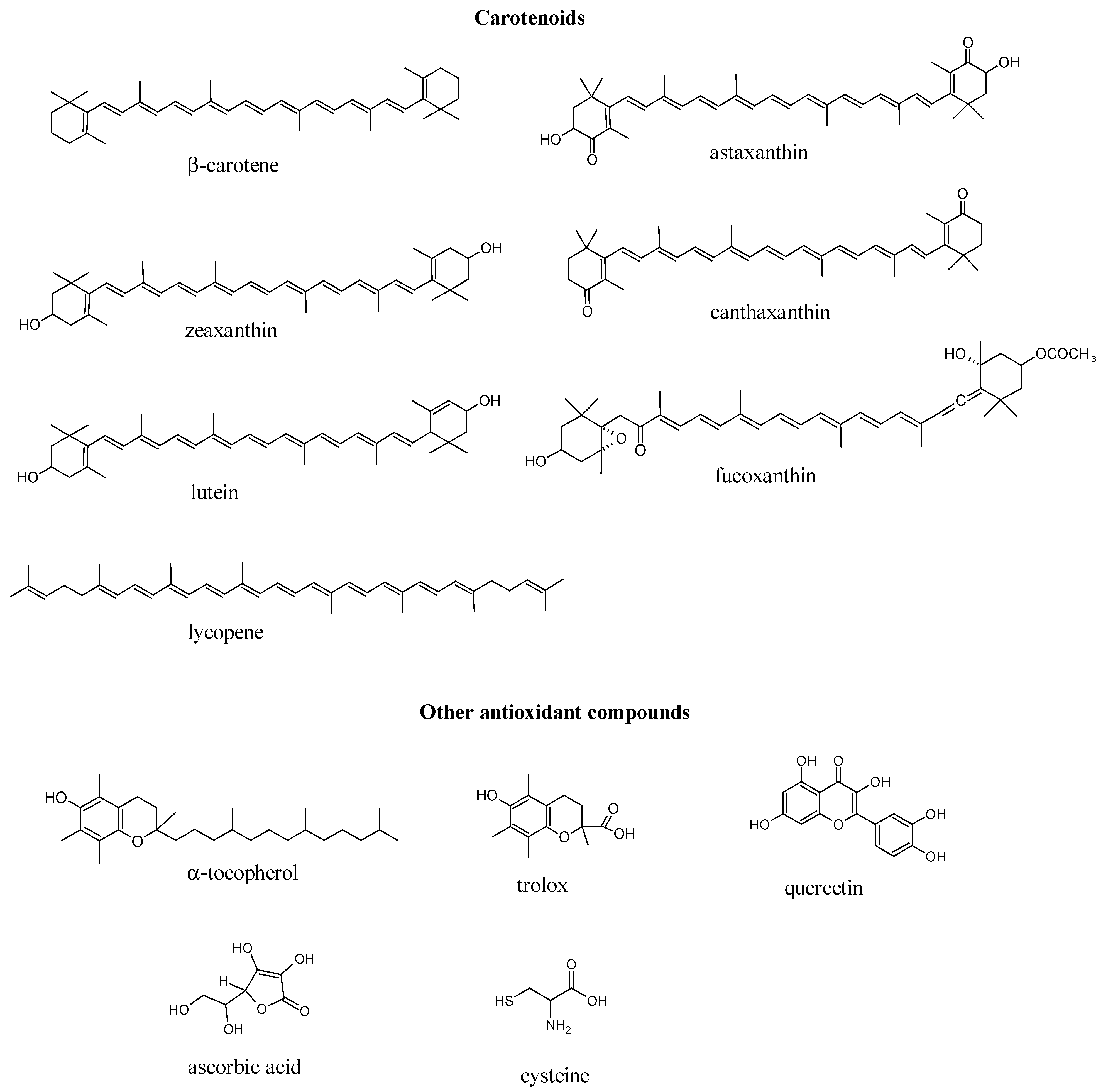

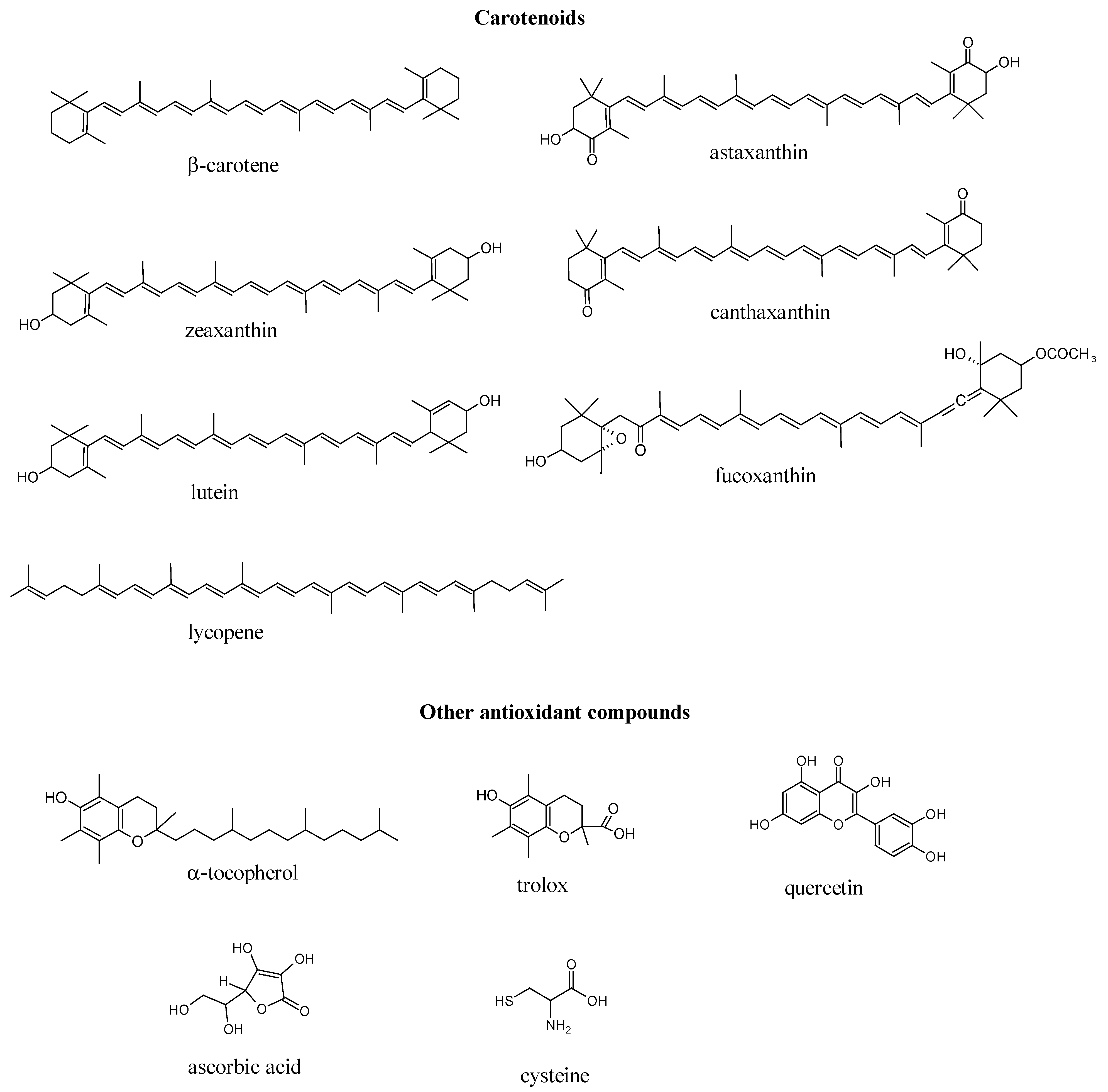{kind=link}
| Compound | Scavenging Capacity a | log P b | |||
|---|---|---|---|---|---|
| ROO• | HO• | HOCl | ONOO− | ||
| β-carotene | 0.14 | 0.71 | NA | 1.02 | 14.76 ± 0.43 |
| zeaxanthin | 0.56 | 1.41 | 3.87 | 0.77 | 10.92 ± 0.45 |
| lutein | 0.60 | 0.97 | 4.81 | 0.78 | 11.52 ± 0.46 |
| lycopene | 0.08 | 0.35 | 0.40 | 0.31 | 14.53 ± 0.45 |
| fucoxanthin | 0.43 | 1.18 | 6.26 | NA | 7.30 ± 0.65 |
| canthaxanthin | 0.04 | 0.28 | 0.10 | NA | 9.53 ± 0.45 |
| astaxanthin | 0.64 | 1.66 | 9.40 | 0.73 | 8.24 ± 0.59 |
| α-tocopherol | 0.48 | 1.77 | NA | 0.37 | 10.96 ± 0.35 |
| trolox | 1.00 | 1.00 | NA | NA | 2.46 ± 0.36 |
| quercetin | 0.84 | 1.42 | 5.63 | 0.97 | 1.99 ± 1.08 |
| ascorbic acid | NA c | NA | 0.41 | 1.00 | −2.78 ± 0.42 |
| cysteine | 0.04 | NA | 1.00 | 0.02 | 0.08 ± 0.32 |
3. Discussion



4. Experimental Section
4.1. Chemicals and Standards
4.2. Preparation and Characterization of the Liposomes
| Reactive species | Linearity range (mol%) | R2 a | Slope b | Intercept b |
|---|---|---|---|---|
| ROO•c | 1.0–8.0 | 0.96 | 20.8 | 4.7 |
| HO•c | 1.0–6.0 | 0.97 | 19.3 | 3.9 |
| HOCl d | 6.0–51.0 | 0.98 | 0.17 | 2.3 |
| ONOO− e | 1.6–12.8 | 0.98 | 3.5 | 9.2 |
4.3. ROS and RNS Scavenging Capacity Assays
4.3.1. Adaptation and Validation of the Methods
4.3.2. Peroxyl Radical Scavenging Assay
4.3.3. Hydroxyl Radical Scavenging Assay
4.3.4. Hypochlorous Acid Scavenging Assay
4.3.5. Peroxynitrite Scavenging Assay
4.3.6. Calculation of the Scavenging Capacity


4.4. Partition Ratio (log P)
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Britton, G. UV/Visible Spectroscopy. In Carotenoids: Spectroscopy; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 1995; Volume 1B, pp. 13–62. [Google Scholar]
- Mandelli, F.; Miranda, V.S.; Rodrigues, E.; Mercadante, A.Z. Identification of carotenoids with high antioxidant capacity produced by extremophile microorganisms. World J. Microbiol. Biotechnol. 2011, 28, 1781–1790. [Google Scholar]
- Kelman, D.; Posner, E.K.; McDermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant activity of Hawaiian marine algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids in marine animals. Mar. Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef]
- Voutilainen, S.; Nurmi, T.; Mursu, J.; Rissanen, T. Carotenoids and cardiovascular health. Am. J. Clin. Nutr. 2006, 83, 1265–1271. [Google Scholar]
- Zhang, J.; Dhakal, I.; Stone, A.; Ning, B.; Greene, G.; Lang, N.P.; Kadlubar, F.F. Plasma carotenoids and prostate cancer: A population-based case-control study in Arkansas. Nutr. Cancer 2007, 59, 46–53. [Google Scholar] [CrossRef]
- Rock, C.L. Carotenoids and cancer. In Carotenoids: Nutrition and Health; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 2009; Volume 5, pp. 269–282. [Google Scholar]
- Serpeloni, J.M.; Barcelos, G.R.M.; Friedmann, A.; José, P.; Mercadante, A.Z.; Bianchi, M.L.P.; Antunes, L.M.G. Dietary carotenoid lutein protects against DNA damage and alterations of the redox status induced by cisplatin in human derived HepG2 cells. Toxicol. Vitro 2011, 26, 288–294. [Google Scholar]
- Halliwell, B.; Gutteridge, J. Radicals in Biology and Medicine, 4th ed; Oxford University Press: New York, NY, USA, 2007; pp. 187–267. [Google Scholar]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Naguib, Y.M.A. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Rodrigues, E.; Mariutti, L.R.B.; Chisté, R.C.; Mercadante, A.Z. Development of a novel micro-assay for evaluation of peroxyl radical scavenger capacity: Application to carotenoids and structure-activity relationship. Food Chem. 2012. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Miller, N.J.; Sampson, J.; Candeias, L.P.; Bramley, P.M.; Rice-Evans, C.A. Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 1996, 384, 240–242. [Google Scholar] [CrossRef]
- Roberts, W.G.; Gordon, M.H. Determination of the total antioxidant activity of fruits and vegetables by a liposome assay. J. Agric. Food Chem. 2003, 51, 1486–1493. [Google Scholar] [CrossRef]
- Woodall, A.A.; Lee, S.W.M.; Weesie, R.J.; Jackson, M.J.; Britton, G. Oxidation of carotenoids by free radicals: Relationship between structure and reactivity. Biochim. Biophys. Acta 1336, 33–42. [Google Scholar]
- Zhang, J.; Stanley, R.A.; Melton, L.D. Lipid peroxidation inhibition capacity assay for antioxidants based on liposomal membranes. Mol. Nutr. Food Res. 2006, 50, 714–724. [Google Scholar] [CrossRef]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef]
- Gruszecki, W.I. Carotenoids in Lipid Membranes. In Carotenoids: Physical, Chemical, and Biological Functions and Properties; Landrum, J.T., Ed.; CRC Press: New York, NY, USA, 2009; pp. 19–30. [Google Scholar]
- Faria, A.F.; Mignone, R.A.; Montenegro, M.A.; Mercadante, A.Z.; Borsarelli, C.D. Characterization and singlet oxygen quenching capacity of spray-dried microcapsules of edible biopolymers containing antioxidant molecules. J. Agric. Food Chem. 2010, 58, 8004–8011. [Google Scholar] [CrossRef]
- Rodrigues, E.; Mariutti, L.R.B.; Faria, A.F.; Mercadante, A.Z. Microcapsules containing antioxidant molecules as scavengers of reactive oxygen and nitrogen species. Food Chem. 2012, 134, 704–711. [Google Scholar] [CrossRef]
- Kiokias, S.; Oreopoulou, V. Antioxidant properties of natural carotenoid extracts against the AAPH-initiated oxidation of food emulsions. Innov. Food Sci. Emerg. Technol. 2006, 7, 132–139. [Google Scholar] [CrossRef]
- Scheidegger, R.; Pande, A.K.; Bounds, P.L.; Koppenol, W.H. The reaction of peroxynitrite with zeaxanthin. Nitric Oxide 1998, 2, 8–16. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar]
- Castro, I.A.; Rogero, M.M.; Junqueira, R.M.; Carrapeiro, M.M. Free radical scavenger and antioxidant capacity correlation of α-tocopherol and Trolox measured by three in vitro methodologies. Int. J. Food Sci. Nutr. 2006, 57, 75–82. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Development and validation of oxygen radical absorbance capacity assay for lipophilic antioxidants using randomly methylated β-cyclodextrin as the solubility enhancer. J. Agric. Food Chem. 2002, 50, 1815–1821. [Google Scholar] [CrossRef]
- Pastor, N.; Weinstein, H.; Jamison, E.; Brenowitz, M. A detailed interpretation of OH radical footprints in a TBP DNA complex reveals the role of dynamics in the mechanism of sequence-specific binding. J. Mol. Biol. 2000, 304, 55–68. [Google Scholar] [CrossRef]
- Pullar, J.M.; Vissers, M.C.; Winterbourn, C.C. Living with a killer: The effects of hypochlorous acid on mammalian cells. IUBMB Life 2000, 50, 259–266. [Google Scholar] [CrossRef]
- Malech, H.L.; Gallin, J.I. Neutrophils in human diseases. N. Engl. J. Med. 1987, 317, 687–694. [Google Scholar] [CrossRef]
- Malle, E.; Buch, T.; Grone, H.J. Myeloperoxidase in kidney disease. Kidney Int. 2003, 64, 956–967. [Google Scholar]
- Pennathur, S.; Maitra, D.; Byun, J.; Sliskovic, I.; Abdulhamid, I.; Saed, G.M.; Diamond, M.P.; Abu-Soud, H.M. Potent antioxidative activity of lycopene: A potential role in scavenging hypochlorous acid. Free Radic. Biol. Med. 2010, 49, 205–213. [Google Scholar]
- Yokota, T.; Ohtake, T.; Ishikawa, H.; Inakuma, T.; Ishiguro, Y.; Terao, J.; Nagao, A.; Etoh, H. Quenching of peroxynitrite by lycopene in vitro. Chem. Lett. 2004, 33, 80–81. [Google Scholar] [CrossRef]
- Yoshioka, R.; Hayakawa, T.; Ishizuka, K.; Kulkarni, A.; Terada, Y.; Maoka, T.; Etoh, H. Nitration reactions of astaxanthin and β-carotene by peroxynitrite. Tetrahedron Lett. 2006, 47, 3637–3640. [Google Scholar]
- Tsuboi, M.; Etoh, H.; Yomoda, Y.; Kato, K.; Kato, H.; Kulkarni, A.; Terada, Y.; Maoka, T.; Mori, H.; Inakuma, T. Nitration reaction of lutein with peroxynitrite. Tetrahedron Lett. 2010, 51, 676–678. [Google Scholar]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a marine carotenoid present in seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- D’Orazio, N.; Gemello, E.; Gammone, M.A.; Girolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A Treasure from the Sea. Mar. Drugs 2012, 10, 604–616. [Google Scholar] [CrossRef]
- Gross, G.J.; Hazen, S.L.; Lockwood, S.F. Seven day oral supplementation with Cardax (disodium disuccinate astaxanthin) provides significant cardioprotection and reduces oxidative stress in rats. Mol. Cell Biochem. 2006, 283, 23–30. [Google Scholar] [CrossRef]
- Lauver, D.A.; Driscoll, E.M.; Lucchesi, B.R. Disodium disuccinate astaxanthin prevents carotid artery rethrombosis and ex vivo platelet activation. Pharmacology 2008, 82, 67–73. [Google Scholar] [CrossRef]
- Holick, C.N.; Michaud, D.S.; Stolzenberg-Solomon, R.; Mayne, S.T.; Pietinen, P.; Taylor, P.R.; Virtamo, J.; Albanes, D. Dietary carotenoids, serum β-carotene, and retinol and risk of lung cancer in the alpha-tocopherol, beta-carotene cohort study. Am. J. Epidemiol. 2002, 156, 536–547. [Google Scholar] [CrossRef]
- Nishino, H.; Murakosh, M.; Ii, T.; Takemura, M.; Kuchide, M.; Kanazawa, M.; Mou, X.Y.; Wada, S.; Masuda, M.; Ohsaka, Y.; et al. Carotenoids in cancer chemoprevention. Cancer Metastasis Rev. 2002, 21, 257–264. [Google Scholar] [CrossRef]
- Davies, B.H. Carotenoids. In Chemistry and Biochemistry of Plant Pigments; Goodwin, T.W., Ed.; Academic Press: London, UK, 1976; Volume 2, pp. 38–122. [Google Scholar]
- Drummen, G.P.; van Liebergen, L.C.; Op Den Kamp, J.A.; Post, J.A. C11-BODIPY(581/591), an oxidation-sensitive fluorescent lipid peroxidation probe: (Micro)spectroscopic characterization and validation of methodology. Free Radic. Biol. Med. 2002, 33, 473–490. [Google Scholar] [CrossRef]
Supplementary Files
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rodrigues, E.; Mariutti, L.R.B.; Mercadante, A.Z. Scavenging Capacity of Marine Carotenoids against Reactive Oxygen and Nitrogen Species in a Membrane-Mimicking System. Mar. Drugs 2012, 10, 1784-1798. https://doi.org/10.3390/md10081784
Rodrigues E, Mariutti LRB, Mercadante AZ. Scavenging Capacity of Marine Carotenoids against Reactive Oxygen and Nitrogen Species in a Membrane-Mimicking System. Marine Drugs. 2012; 10(8):1784-1798. https://doi.org/10.3390/md10081784
Chicago/Turabian StyleRodrigues, Eliseu, Lilian R. B. Mariutti, and Adriana Z. Mercadante. 2012. "Scavenging Capacity of Marine Carotenoids against Reactive Oxygen and Nitrogen Species in a Membrane-Mimicking System" Marine Drugs 10, no. 8: 1784-1798. https://doi.org/10.3390/md10081784
APA StyleRodrigues, E., Mariutti, L. R. B., & Mercadante, A. Z. (2012). Scavenging Capacity of Marine Carotenoids against Reactive Oxygen and Nitrogen Species in a Membrane-Mimicking System. Marine Drugs, 10(8), 1784-1798. https://doi.org/10.3390/md10081784