Toxin Levels and Profiles in Microalgae from the North-Western Adriatic Sea—15 Years of Studies on Cultured Species


 ,
,  ,
,
Abstract
:
Abbreviations
| AA | 5,8,11,14-eicosatetraenoic acid |
| BTX | Brevetoxin |
| DA | Domoic acid |
| DSP | Diarrhetic shellfish poisoning |
| DTX | Dinophysistoxin |
| EC50 | 50% effect concentration |
| ELA | Erythrocyte lysis assay |
| EPA | 5,8,11,14,17-eicosapentaenoic acid |
| GTX | Gonyautoxin |
| ITS | Internal transcribed spacer |
| LC-MS | Liquid chromatography-mass spectrometry |
| LSU | Large subunit |
| NEO | Neosaxitoxin |
| NMR | Nuclear magnetic resonance |
| OA | Okadaic acid |
| OTA | 6,9,12,15-octadecatetraenoic acid |
| OVTX | Ovatoxin |
| pPLTX | Putative palytoxin |
| PSP | Paralytic shellfish poisoning |
| PUFAs | Polyunsaturated fatty acids |
| ROS | Reactive oxygen species |
| SEM | Scanning electron microscope |
| SSU | Small subunit |
| STX | Saxitoxin |
| YTX | Yessotoxin |
1. Introduction
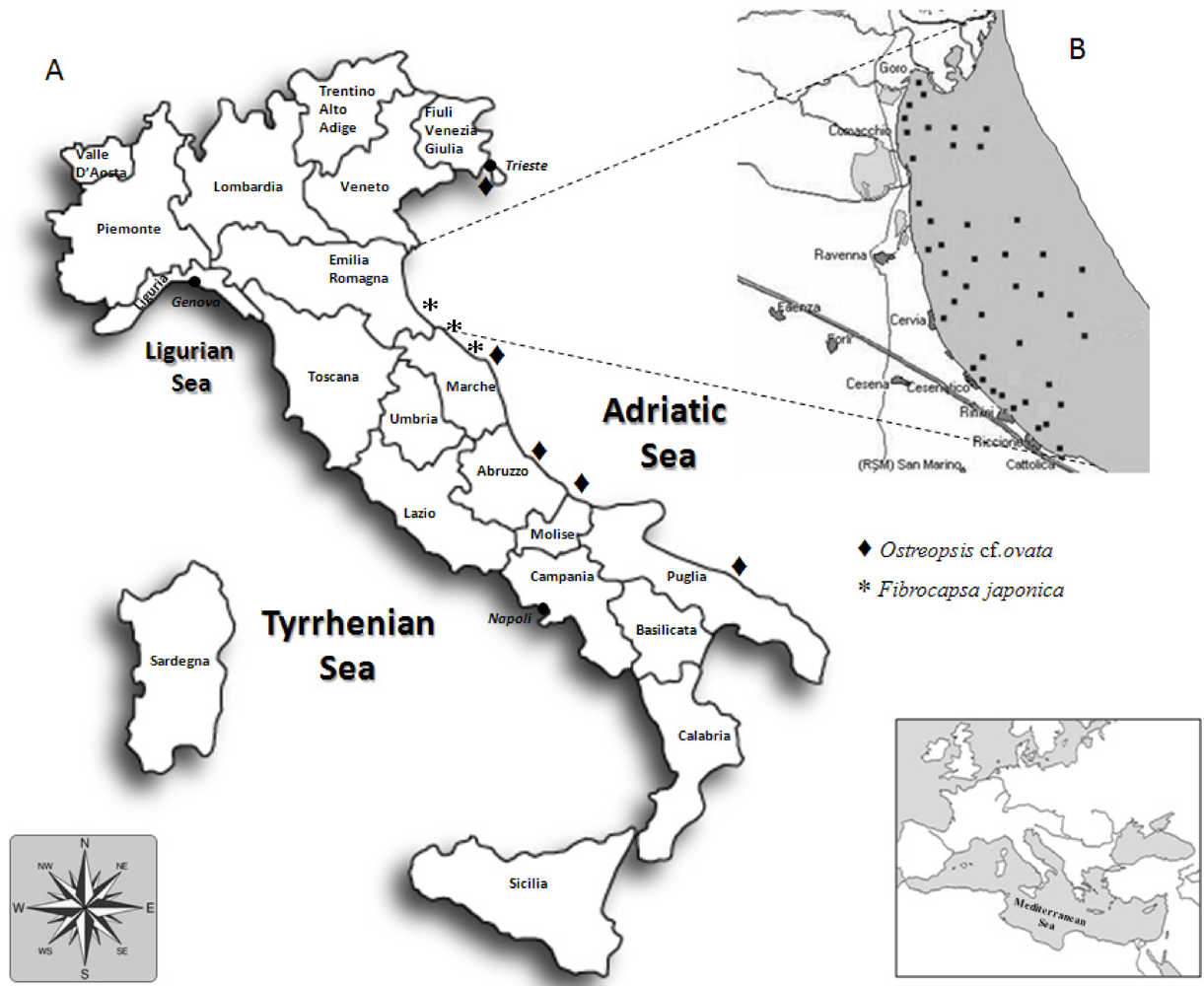
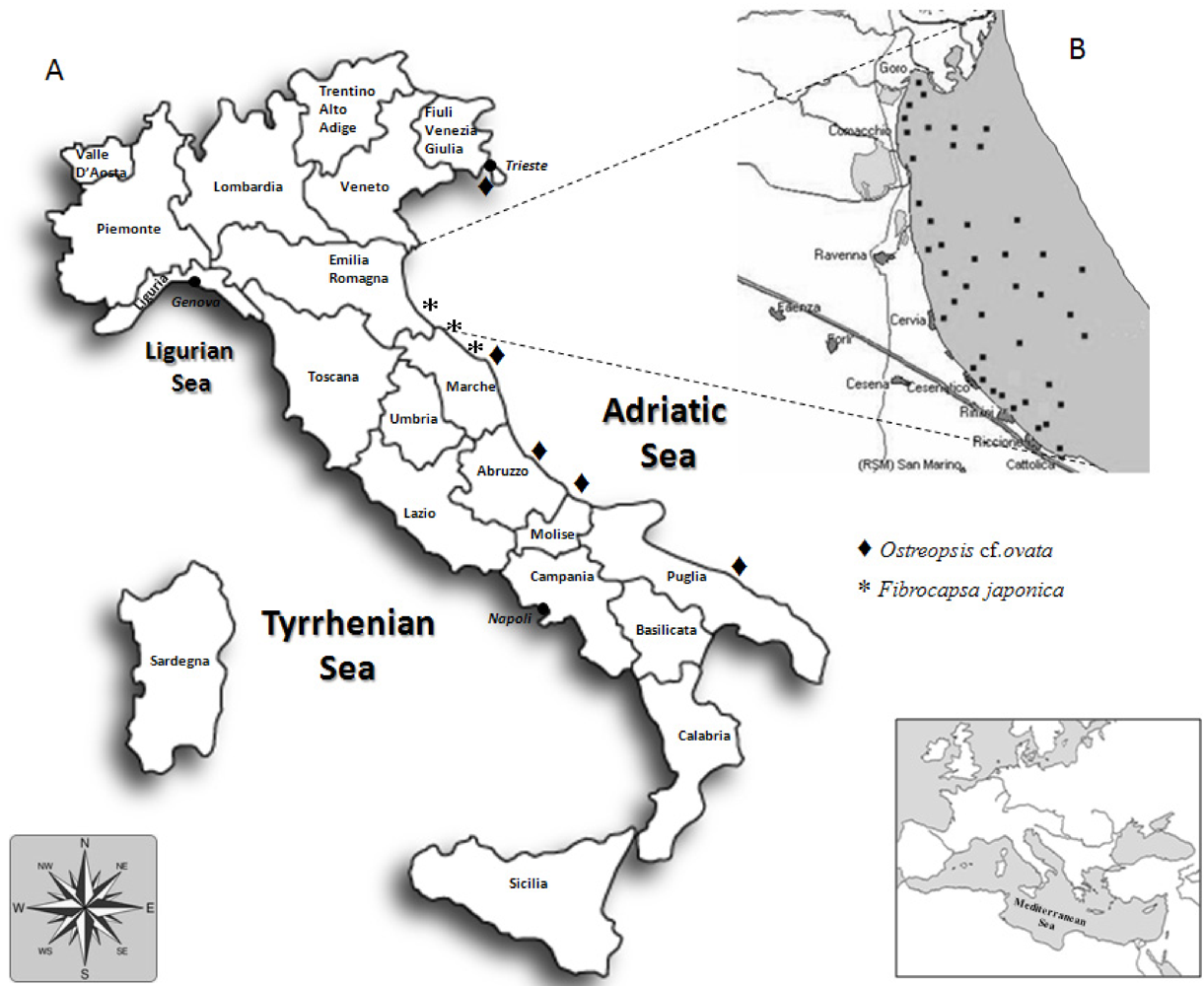
2. Overview of Toxins Detected in Shellfish Farmed along the Coast of the Emilia-Romagna Region from 1980s to Date
2.1. Toxins Affecting Humans Due to the Consumption of Mussels and Responsible for Shellfish Farms Closure
2.2. Toxins Sporadically Detected in Cultivated Mussels
3. Algal Species and Toxins Mostly Affecting the Ecosystem and/or Humans by Aerosol Inhalation
4. Studies on Qualitative and Quantitative Toxin Profiles of Cultured Microalgae
4.1. Yessotoxin Group
4.2. Saxitoxin Group
4.3. Okadaic Acid Group
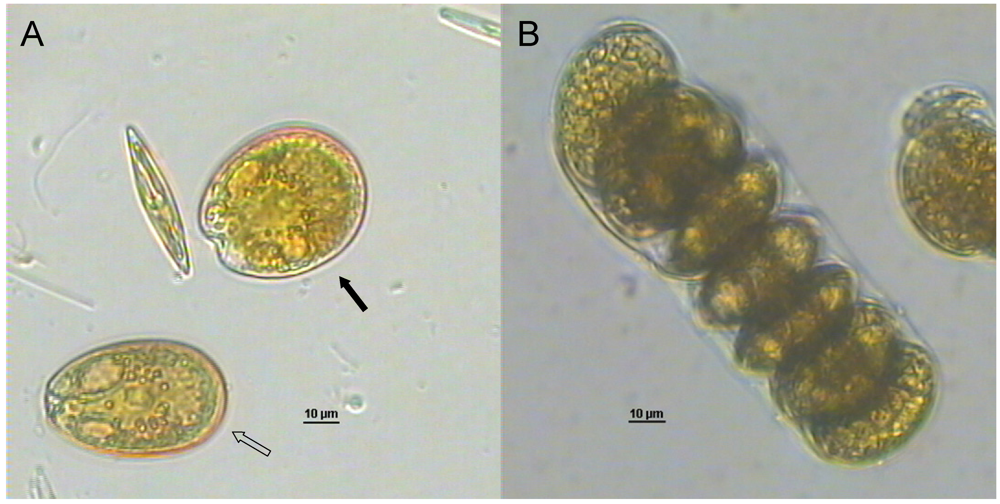
4.4. Cyclic Imine Group
4.5. Domoic Acid Group

{kind=link}
{kind=link}
{kind=link}
{kind=link}
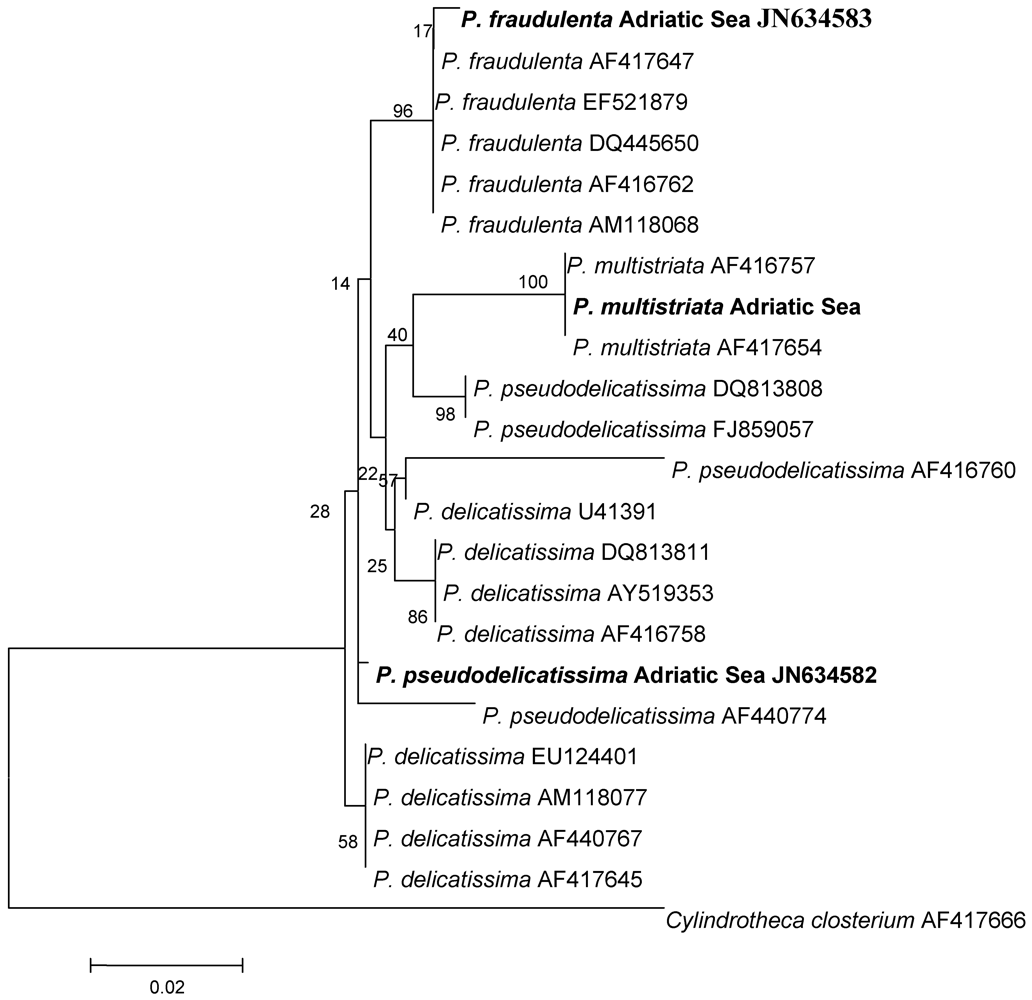
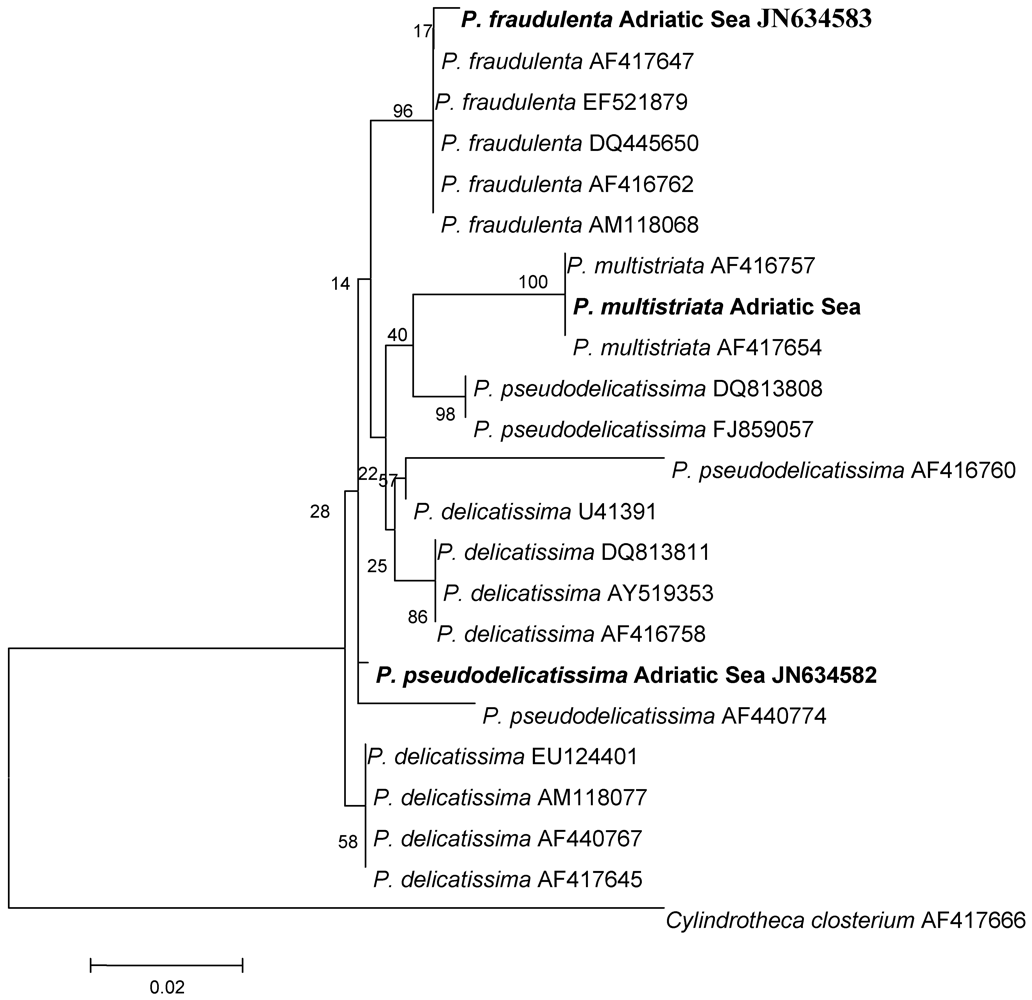{kind=link}
| Apical Axis (μm) | Transapical Axis (μm) | Overlap of Cell Length | ||||
|---|---|---|---|---|---|---|
| Studied Strain | Ref. Value | Studied Strain | Ref. Value | Studied Strain | Ref. Value | |
| P. pseudodelicatissima | 64.2–70.0 | 50–140 a | 2.1–2.9 | 1.5–3.4 a | 1/6 | 1/5–1/6 a |
| P. fraudulenta | 75.6–80.0 | 50–119 a | 5.0–6.7 | 4.5–10 a | 1/5 | / |
| P. multistriata | 40.6–42.8 | 38–65 b | 3.0–3.3 | 2.5–4 b | 1/8 | / |
4.6. Palytoxin Group
4.7. Ichthyotoxins
| EC50 (cell/mL) | ||
|---|---|---|
| Artemia sp. Assay | Erythrocyte Lysis Assay | |
| Heterosigma akashiwo | 3671 ± 59 | 187,263 ± 7152 |
| Chattonella cf. subsalsa | 495 ± 15 | 27,161 ± 8031 |
5. Conclusions
| Species | Toxin | Toxin Levels | Effect | Toxin Presence in Mussels (Year) |
|---|---|---|---|---|
| Dinophysis spp. | OA, DTXs | n.d. | DSP | 1989–1997, 2010 |
| Protoceratium reticulatum | YTX prevalent, homoYTX, 45-OHYTX, carboxyYTX, noroxoYTX | 4.5–65 pg/cell (YTX) | Neurotoxic through i.p. injection | 1995–2006 |
| Lingulodinium polyedrum | Homo-YTX | from none to 0.80 pg/cell | Neurotoxic through i.p. injection | 1996 |
| Gonyaulax spinifera | Homo-YTX prevalent, YTX | 33,4 pg/cell (Homo-YTX) | Neurotoxic through i.p. injection | 2007 |
| Alexandrium minutum | GTX4 prevalent, GTX2, GTX3, GTX1 | 964 fg/cell (GTX4) | PSP | 1993–1994 |
| Prorocentrum lima | OA prevalent, DTX1, DTX2, DTX4, DTX6 | 6.7–8.4 pg/cell (OA) | DSP | 2002 |
| Alexandrium ostenfeldii | 13-desMe C, 13,19-didesMe C e 27-OH-13,19-didesMe C prevalent plusspirolides A,B,C,D, 13-desMe D | 3,7 pg/cell (13-desMeC) | Neurotoxic through i.p. injection | 2003 |
| Pseudo-nitzschia multistriata | Domoic acid | 0.003–0.010 pg/cell | ASP | Sporadic since 2000 |
| Bloom Presence | ||||
| Ostreopsis cf. ovata | Ovatoxin-a prevalent, OVTX-b, c, d-e, pPLTX | 20–130 pg/cell (total) | Respiratory distress | Recurrent since 2001 |
| Fibrocapsa japonica | ROS, PUFA | - | Ichthyotoxic | Recurrent since 1994 |
Acknowledgments
References
- Okaichi, T. Red Tides; Terra Scientific Publishing Company: Tokyo, Japan; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; p. 439. [Google Scholar]
- Granéli, E.; Tuner, J.T. Ecology of Harmful Algae; Springher-Verlag: Berlin Heidelberg, Germany, 2006; p. 413. [Google Scholar]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Physiological Ecology of Harmful Algal Blooms; Springher-Verlag: Berlin Heidelberg, Germany, 1998; p. 662. [Google Scholar]
- Riccardi, M.; Guerrini, F.; Serrazanetti, G.P.; Ventrella, V.; Pagliarani, A.; Pistocchi, R. Lipid and DNA features of Gonyaulax fragilis (Dinophyceae) as potential biomarkers in mucilage genesis. Harmful Algae 2010, 9, 359–366, and references therein.. [Google Scholar]
- Croci, L.; Cozzi, L.; Suffredini, E.; Ciccaglioni, G.; Toti, L.; Milandri, A.; Ceredi, A.; Benzi, M.; Poletti, P. Characterization of microalgae and associated bacteria collected from shellfish harvesting areas. Harmful Algae 2006, 5, 266–274. [Google Scholar]
- France, J.; Mozetič, P. Ecological characterization of toxic phytoplankton species (Dinophysis spp., Dinophyceae) in Slovenian mariculture areas (Gulf of Trieste, Adriatic Sea) and the implications for monitoring. Mar. Poll. Bull. 2006, 52, 1504–1516. [Google Scholar] [CrossRef]
- Penna, A.; Ingarao, C.; Ercolessi, M.; Rocchi, M.; Penna, N. Potentially harmful microalgal distribution in an area of the NW Adriatic coastline: Sampling procedure and correlations with environmental factors. Estuar. Coast. Shelf Sci. 2006, 70, 307–316. [Google Scholar]
- Ninčević-Gladan, Ž.; Skejić, S.; Buzančić, M.; Marasovic, I.; Arapov, J.; Ujević, I.; Bojanić, N.; Grbec, B.; Kušpilić, G.; Vidjak, O. Seasonal variability in Dinophysis spp. abundances and diarrhetic shellfish poisoning outbreaks along the eastern Adriatic coast. Bot. Mar. 2008, 51, 449–463. [Google Scholar]
- Ljubeķšić, Z.; Bosak, S.; Viličić, D.; Borojević, K.K.; Marić, D.; Godrijan, J.; Ujević, I.; Peharec, P.; Ðakovac, T. Ecology and taxonomy of potentially toxic Pseudo-nitzschia species in Lim Bay (north-eastern Adriatic Sea). Harmful Algae 2011, 10, 713–722. [Google Scholar] [CrossRef]
- Viviani, R.; Poletti, R. Tossine acquatiche diffusione e controllo (parte prima). Laguna Reg. Emilia Romagna 1999, 5, 6–15. [Google Scholar]
- Boni, L. Red tides off the coast of Emilia-Romagna (north western Adriatic Sea) from 1975 to 1982. Inf. Bot. Ital. 1983, 15, 18–24. [Google Scholar]
- Boni, L. Toxic marine phytoplankton in Italy. G. Bot. Ital. 1992, 126, 229–236. [Google Scholar]
- Fattorusso, E.; Ciminiello, P.; Costantino, V.; Magno, S.; Mangoni, A.; Milandri, A.; Poletti, R.; Pompei, M.; Viviani, R. Okadaic acid in mussels of Adriatic sea. Mar. Pollut. Bull. 1992, 24, 234–237. [Google Scholar]
- Bonivento, P.; Battistella, S.; Amirante, G.A. Recognition of a 31,000 molecular weight protein synthesised by Mytilus galloprovincialis Lmk. during Dinophysis spp. bloom in the Gulf of Trieste (Adriatic Sea). Toxicon 1997, 35, 1347–1350. [Google Scholar]
- Poletti, R.; Cangini, M.; Ceredi, A.; Magnani, F.; Milandri, A.; Pigozzi, S.; Pompei, M.; Riccardi, E.; Zoffoli, S. Bioaccumulo di tossine algali liposolubili in Mytilus galloprovincialis (Lamark, 1819) nel Nord Adriatico. Biol. Mar. Medit. 2008, 15, 54–57. [Google Scholar]
- Pavela-Vrančič, M.; Meštrović, V.; Marasović, I.; Gillmanc, M.; Fureyc, A.; Jamesc, K.J. DSP toxin profile in the coastal waters of the central Adriatic Sea. Toxicon. 2002, 40, 1601–1607. [Google Scholar]
- Poletti, R. Personal communication. National Reference Laboratory for Marine Biotoxins: Cesenatico, Italy, 2010. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Oshima, Y.; Poletti, R.; Viviani, R.; Yasumoto, T. Determination of PSP toxins in mussels from the Adriatic sea. Mar. Pollut. Bull. 1995, 30, 733–735. [Google Scholar]
- Honsell, G.; Boni, L.; Cabrini, M.; Pompei, M. Toxic or Potentially Toxic Dinoflagellates from the Northern Adriatic Sea. In Marine Coastal Eutrophication Science Total Environment; Vollenweider, R.A., Marchetti, R., Viviani, R., Eds.; Elsevier: Amsterdam, The Netherlands, 1992; pp. 107–114. [Google Scholar]
- Honsell, G. First Report of Alexandrium minutum in the Northern Adriatic Waters (Mediterranean Sea). In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, J., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1993; pp. 127–132. [Google Scholar]
- Boni, L.; Pompei, M.; Reti, M. Maree colorate e fioriture algali lungo la costa dell’Emilia Romagna dal 1982 al 1985 con particolare riguardo alla comparsa di Protogoniaulax tamarensis. Nova Thalass. 1986, 3, 237–245. [Google Scholar]
- Honsell, G.; Poletti, R.; Pompei, M.; Sidari, L.; Milandri, A.; Casadei, C.; Viviani, R. Alexandrium minutum Halim and PSP Contamination in the Northern Adriatic Sea (Mediterranean Sea). In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; UNESCO: Paris, France, 1996; pp. 77–80. [Google Scholar]
- Milandri, A.; Pompei, M.; Poletti, R. Fioriture tossiche in Emilia Romagna e attività del centro nazionale di riferimento per le biotossine marine. In Toxic algal blooms in Italy: risk assessment and epidemiology; Mattei, D., Melchiorre, S., Messineo, V., Bruno, M., Eds.; Rapporti ISTISAN 05/29, Tipografia Facciotti: Roma, Italy, 2005; pp. 123–129. [Google Scholar]
- Ujević, I.; Roje, R.; Ninčević-Gladan, Ž.; Marasović, I. First report of Paralytic Shellfish Poisoning (PSP) in mussels (Mytilus galloprovincialis) from eastern Adriatic Sea (Croatia). Food Control 2011. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, S.; Tartaglione, L.; Quilliam, M.A.; Tubaro, A.; Poletti, R. Hydrophilic interaction liquid chromatography-mass spectrometry (HILIC-MS) for determination of domoic acid in Adriatic shellfish. Rapid Commun. Mass Spectrom. 2005, 19, 2030–2038. [Google Scholar]
- Poletti, R.; Milandri, A.; Pompei, M.; Viviani, R. Controllo delle biotossine ASP nei prodotti ittici. Laguna Reg. Emilia Romagna 1997, 5, 6–17. [Google Scholar]
- Ujević, I.; Ninčević-Gladan, Z.; Roje, R.; Skejić, S.; Arapov, J.; Marasović, I. Domoic acid—A new toxin in the Croatian Adriatic shellfish toxin profile. Molecules 2010, 15, 6835–6849. [Google Scholar]
- Marić, D.; Ljubeķšić, Z.; Godrijan, J.; Viličić, D.; Ujević, I.; Precali, R. Blooms of the potentially toxic diatom Pseudo-nitzschia calliantha Lundholm, Moestrup & Hasle in coastal waters of the northern Adriatic Sea (Croatia). Estuar. Coast. Shelf Sci. 2011, 92, 323–331. [Google Scholar]
- Cucchiari, E.; Guerrini, F.; Penna, A.; Totti, C.; Pistocchi, R. Effect of salinity, temperature, organic and inorganic nutrients on growth of cultured Fibrocapsa japonica (Raphidophyceae) from the northern Adriatic Sea. Harmful Algae 2008, 7, 405–414. [Google Scholar]
- Cucchiari, E.; Pistocchi, R.; Pezzolesi, L.; Penna, A.; Battocchi, C.; Cerino, F.; Totti, C. Resting cysts of Fibrocapsa japonica (Raphidophyceae) from coastal sediments of the northern Adriatic Sea (Mediterranean Sea). Harmful Algae 2010, 10, 81–87. [Google Scholar]
- Pezzolesi, L.; Cucchiari, E.; Guerrini, F.; Pasteris, A.; Galletti, P.; Tagliavini, E.; Totti, C.; Pistocchi, R. Toxicity evaluation of Fibrocapsa japonica from the Northern Adriatic Sea through a chemical and toxicological approach. Harmful Algae 2010, 9, 504–514. [Google Scholar]
- Penna, A.; Vila, M.; Fraga, S.; Giacobbe, M.G.; Andreoni, F.; Riobó, P.; Vernesi, C. Characterization of Ostreopsis and Coolia (Dinophyceae) isolates in the Western Mediterranean Sea based on morphology, toxicity and internal transcribed spacer 5.8S rDNA sequences. J. Phycol. 2005, 41, 212–225. [Google Scholar] [CrossRef]
- Aligizaki, K.; Nikolaidis, G. The presence of the potentially toxic genera Ostreopsis and Coolia (Dinophyceae) in the North Aegean Sea, Greece. Harmful Algae 2006, 5, 717–730. [Google Scholar]
- Mangialajo, L.; Ganzin, N.; Accoroni, S.; Asnaghi, V.; Blanfuné, A.; Cabrini, M.; Cattaneo-Vietti, R.; Chavanon, F.; Chiantore, M.; Cohu, S.; et al. Trends in Ostreopsis proliferation along the Northern Mediterranean coasts. Toxicon 2011, 57, 408–420. [Google Scholar] [PubMed]
- Brescianini, C.; Grillo, C.; Melchiorre, N.; Bertolotto, R.; Ferrari, A.; Vivaldi, B.; Icardi, G.; Gramaccioni, L.; Funari, E.; Scardala, S. Ostreopsis ovata algal blooms affecting human health in Genova, Italy, 2005 and 2006. Euro Surveill. 2006, 11, E060907.3. [Google Scholar]
- Gallitelli, M.; Ungaro, N.; Addante, L.M.; Procacci, V.; Silveri, N.G.; Sabbà, C. Respiratory illness as a reaction to tropical algal blooms occurring in a temperate climate. J. Am. Med. Assoc. 2005, 293, 2599–2600. [Google Scholar]
- Totti, C.; Cucchiari, E.; Romagnoli, T.; Penna, A. Bloom of Ostreopsis ovata in the Conero Riviera (NW Adriatic Sea). Harmful Algae News 2007, 33, 12–13. [Google Scholar]
- Totti, C.; Accoroni, S.; Cerino, F.; Cucchiari, E.; Romagnoli, T. Ostreopsis ovata bloom along the Conero Riviera (northern Adriatic Sea): Relationships with environmental conditions and substrata. Harmful Algae 2010, 9, 233–239. [Google Scholar]
- Ministero Della Salute. Gestione del Rischio Associato alle Fioriture di Ostreopsis Ovata Nelle Coste Italiane. Linee Guida; Ministero Della Salute: Roma, Itlay, 2007. Available online: http://www.salute.gov.it/imgs/C_17_pubblicazioni_641_allegato.pdf (accessed on 16 January 2012).
- Monti, M.; Minocci, M.; Beran, A.; Ivesa, L. First record of Ostreopsis cfr. ovata on macroalgae in the Northern Adriatic Sea. Mar. Pollut. Bull. 2007, 54, 598–601. [Google Scholar]
- Ingarao, C.; Lanciani, G.; Teodori, A.; Pagliani, T. First presence of Ostreopsis cfr. ovata (Dinophyceae) along Abruzzo coasts (W Adriatic Sea). Biol. Mar. Medit. 2009, 16, 172–173. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Magno, S.; Poletti, R.; Satake, M.; Viviani, R.; Yasumoto, T. Yessotoxin in mussels of the northern Adriatic Sea. Toxicon 1997, 35, 177–183. [Google Scholar]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Yasumoto, T. Isolation and structure of yessotoxin, a novel polyether compound implicated in diarrhetic shellfish poisoning. Tetrahedron Lett. 1987, 28, 5869–5872. [Google Scholar]
- Boni, L.; Ceredi, A.; Guerrini, F.; Milandri, A.; Pistocchi, R.; Poletti, R.; Pompei, M. Toxic Protoceratium reticulatum (Peridiniales, Dinophyta) in the North-Western Adriatic Sea (Italy). In Harmful Algal Blooms 2000; Hallegraeff, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2001; pp. 37–40. [Google Scholar]
- Satake, M.; MacKenzie, L.; Yasumoto, T. Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin. Nat. Toxins 1997, 5, 164–167. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, S.; Guerrini, F.; Pistocchi, R.; Boni, L. Complex yessotoxins profile in Protoceratium reticulatum from north-western Adriatic Sea revealed by LC-MS analysis. Toxicon 2003, 42, 7–14. [Google Scholar]
- lvarez, G.; Uribe, E.; Díaz, R.; Braun, M.; Mariño, C.; Blanco, J. Bloom of the Yessotoxin producing dinoflagellate Protoceratium reticulatum (Dinophyceae) in Northern Chile. J. Sea Res. 2011, 65, 427–434. [Google Scholar]
- Dominguez, H.J.; Paz, B.; Daranas, A.H.; Norte, M.; Franco, J.M.; Fernández, J.J. Dinoflagellate polyether within the yessotoxin, pectenotoxin and okadaic acid toxin groups: Characterization, analysis and human health implications. Toxicon 2010, 56, 191–217. [Google Scholar]
- Paz, B.; Riobó, P.; Ramilo, I.; Franco, J.M. Yessotoxins profile in strains of Protoceratium reticulatum from Spain and USA. Toxicon 2007, 50, 1–17. [Google Scholar]
- Suzuki, T.; Horie, Y.; Koike, K.; Satake, M.; Oshima, Y.; Iwataki, M.; Yoshimatsu, S. Yessotoxin analogues in several strains of Protoceratium reticulatum in Japan determined by liquid chromatography–hybrid triple quadrupole/linear ion trap mass spectrometry. J. Chrom. A 2007, 1142, 172–177. [Google Scholar]
- Guerrini, F.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; Fattorusso, E.; Boni, L.; Pistocchi, R. Influence of temperature, salinity and nutrient limitation on yessotoxin production and release by the dinoflagellate Protoceratium reticulatum in batch-cultures. Harmful Algae 2007, 6, 707–717. [Google Scholar]
- Riccardi, M.; Guerrini, F.; Pistocchi, R.; Tinti, F.; Boni, L. Molecular characterization of different strains of Protoceratium reticulatum (Claparède & Lachmann) Buetschli. Biol. Mar. Medit. 2006, 13, 1042–1044. [Google Scholar]
- Riccardi, M. Studio di specie fitoplanctoniche implicate nella produzione di mucillagini e tossine attraverso le tecniche del DNA ricombinante. 2007. [Google Scholar]
- Grzebyk, D.; Sako, Y.; Berland, B. Phylogenetic analysis of nine species of Prorocentrum (Dinophyceae) inferred from 18S ribosomal DNA sequences, morphological comparisons, and description of Prorocentrum panamensis sp. nov. J. Phycol. 1998, 34, 1055–1068. [Google Scholar]
- Daugbjerg, N.; Hansen, G.; Larsen, J.; Moestrup, Ø. Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. Phycologia 2000, 39, 302–317. [Google Scholar] [CrossRef]
- Saldarriaga, J.F.; Taylor, F.J.R.; Keeling, P.J.; Cavalier-Smith, T. Dinoflagellate nuclear SSU rRNA phylogeny suggests multiple plastid losses and replacements. J. Mol. Evol. 2001, 53, 204–213. [Google Scholar]
- Riccardi, M.; Guerrini, F.; Roncarati, F.; Milandri, A.; Cangini, M.; Pigozzi, S.; Riccardi, E.; Ceredi, A.; Ciminiello, P.; Dell’Aversano, C.; et al. Gonyaulax spinifera from the Adriatic sea: Toxin production and phylogenetic analysis. Harmful Algae 2009, 8, 279–290. [Google Scholar] [CrossRef]
- Howard, M.D.A.; Smith, G.J.; Kudela, R.M. Phylogenetic relationships of yessotoxin-producing dinoflagellates based on the large subunit and internal transcribed spacer ribosomal DNA domains. Appl. Environ. Microbiol. 2009, 7, 54–63. [Google Scholar]
- Tubaro, A.; Sidari, L.; Della Loggia, R.; Yasumoto, T. Occurrence of Homoyessotoxin in Phytoplankton and Mussels from Northern Adriatic Sea. In Harmful Algae; Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago de Compostela, Spain, 1998; pp. 470–472. [Google Scholar]
- Paz, B.; Riobó, P.; Fernandez, M.L.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar]
- Stobo, L.A.; Lewis, J.; Quilliam, M.A.; Hardstaff, W.R.; Gallacher, S.; Webster, L.; Smith, E.; McKenzie, M. Detection of Yessotoxin in UK and Canadian Isolates of Phytoplankton and Optimization and Validation of LC-MS Methods. In Gulf Fisheries Centre; Bates, S., Ed.; Moncton: New Brunswick, Canada, 2003; pp. 8–14. [Google Scholar]
- Guerrini, F. 2005; University of Bologna, Italy. Unpublished results.
- Rhodes, L.; McNabb, P.; de Salas, M.; Briggs, L.; Beuzenberg, V.; Gladstone, M. Yessotoxin production in Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar]
- Riccardi, M. 2008; University of Bologna, Italy. Unpublished results.
- Poletti, R.; Cangini, M.; Ceredi, A.; Magnani, F.; Milandri, A.; Pigozzi, S.; Pompei, M.; Riccardi, E.; Zoffoli, S. Bioaccumulo di tossine algali liposolubili in Mytilus Galloprovincialis (Lamarck, 1819) del Nord Adriatico. Biol. Mar. Medit. 2008, 15, 54–57. [Google Scholar]
- Masseret, E.; Grzebyk, D.; Nagai, S.; Genovesi, B.; Lasserre, B.; Laabir, M.; Collos, Y.; Vaquer, A.; Berrebi, P. Unexpected genetic diversity among and within populations of the toxic dinoflagellate Alexandrium catenella as revealed by nuclear microsatellite markers. Appl. Environ. Microbiol. 2009, 75, 2037–2045. [Google Scholar]
- Alpermann, T.J.; Tillmann, U.; Beszteri, B.; Cembella, A.D.; John, U. Phenotypic variation and genotypic diversity in a planktonic population of the toxigenic marine dinoflagellate Alexandrium tamarense (Dinophyceae). J. Phycol. 2010, 46, 18–32. [Google Scholar]
- Beani, L.; Bianchi, C.; Guerrini, F.; Marani, L.; Pistocchi, R.; Tomasini, M.C.; Ceredi, A.; Milandri, A.; Poletti, R.; Boni, L. High sensitivity bioassay of paralytic (PSP) and amnesic (ASP) algal toxins based on the fluorimetric detection of [Ca2+]i in rat cortical primary cultures. Toxicon 2000, 38, 1283–1297. [Google Scholar]
- Franco, J.M.; Fraga, S.; Zapata, M.; Bravo, I.; Fernandez, P.; Ramilo, I. Harmful Marine Algal Bloom; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier, Intercept Ltd.: Paris, France, 1995; pp. 53–58. [Google Scholar]
- Chang, F.H.; Anderson, D.M.; Kulis, D.M.; Till, D.G. Toxin production of Alexandrium minutum (Dinophyceae) from the Bay of Plenty, New Zealand. Toxicon 1997, 35, 393–409. [Google Scholar]
- Nascimento, S.M.; Purdie, D.A.; Lilly, E.L.; Larsen, J.; Morris, S. Toxin profile, pigment composition and large subunit rDNA phylogenetic analysis of an Alexandrium minutum (Dinophyceae) strain isolated from the Fleet Lagoon, United Kingdom. J. Phycol. 2005, 41, 343–353. [Google Scholar]
- Anderson, D.M.; Kulis, D.M.; Sullivan, J.J.; Hall, S.; Lee, C. Dynamics and physiology of saxitoxin production by the dinoflagellates Alexandrium spp. Mar. Biol. 1990, 104, 511–524. [Google Scholar]
- Touzet, N.; Franco, J.M.; Raine, R. Influence of inorganic nutrition on growth and PSP toxin production of Alexandrium minutum (Dinophyceae) from Cork Harbour, Ireland. Toxicon 2007, 50, 106–119. [Google Scholar]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects and their increased occurrence. Environ. Health Persp. 2000, 1081, 133–141. [Google Scholar] [CrossRef]
- Faust, M.A.; Vandersen, M.W.; Kibler, S.R.; Tester, P.A.; Litaker, R.W. Prorocentrum levis, a new benthic species (Dinophyceae) from a mangrove island, Twin Cays, Belize. J. Phycol. 2008, 44, 232–240. [Google Scholar]
- Vanucci, S.; Guerrini, F.; Milandri, A.; Pistocchi, R. Effects of different levels of N- and P-deficiency on cell yield, okadaic acid, DTX-1, protein and carbohydrate dynamics in the benthic dinoflagellate Prorocentrum lima. Harmful Algae 2010, 9, 590–599. [Google Scholar]
- Cembella, A.D.; Lewis, N.I.; Quilliam, M.A. Spirolide composition of micro-extracted pooled cells isolated from natural plankton assemblages and from cultures of dinoflagellate Alexandrium ostenfeldii. Nat. Toxins 1999, 7, 197–206. [Google Scholar]
- Mackenzie, L.; White, D.; Oshima, Y.; Kapa, J. The resting cyst of Alexandrium ostenfeldii (Dinophyceae) in New Zealand. Phycologia 1996, 35, 148–155. [Google Scholar]
- Cembella, A.D.; Quilliam, M.A.; Lewis, N.I. The marine dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the causative organism of spirolide shellfish toxins. Phycologia 2000, 39, 67–74. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, S.; Tartaglione, L.; Cangini, M.; Pompei, M.; Guerrini, F.; Boni, L.; et al. Toxin profile of Alexandrium ostenfeldii (Dinophyceae) from the Northern Adriatic Sea revealed by liquid chromatography-mass spectrometry. Toxicon 2006, 47, 597–604. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pistocchi, R. Spirolide toxin profile of Adriatic Alexandrium ostenfeldii cultures and structure elucidation of 27-hydroxy-13,19-didesmethyl spirolide C. J. Nat. Prod. 2007, 70, 1878–1883. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pezzolesi, L.; Pistocchi, R. Characterization of 27-hydroxy-13-desmethyl spirolide C and 27-oxo-13,19-didesmethyl spirolide C. Further insights into the complex Adriatic Alexandrium ostenfeldii toxin profile. Toxicon 2010, 56, 1327–1333. [Google Scholar]
- Skov, J.; Lundholm, N.; Moestrup, Ø.; Larsen, J. Potentially toxic phytoplankton. 4. The Diatom Genus Pseudo-nitzschia (Diatomophyceae/Bacillariophyceae). In ICES Identification Leaflets for Phytoplankton; Lindley, J.A., Ed.; Leaflet No. 185; International Council for the Exploration of the Sea: Copenhagen, Denmark, 1999; pp. 1–23. [Google Scholar]
- Hasle, G.R.; Syvertsen, E.E. Marine Diatoms. In Identifying Marine Phytoplankton; Tomas, C., Ed.; Academic Press: New York, NY, USA, 1997; pp. 5–385. [Google Scholar]
- Riccardi, M. 2007. University of Bologna, Italy. Unpublished results..
- Lundholm, N.; Daugbjerg, N.; Moestrup, Ø. Phylogeny of the Bacillariaceae with emphasis on the genus Pseudo-nitzschia (Bacillariophyceae) based on partial LSU r DNA. Eur. J. Phycol. 2002, 37, 115–134. [Google Scholar] [CrossRef]
- Orsini, L.; Sarno, D.; Procaccini, G.; Poletti, R.; Dahlmann, J.; Montresor, M. Toxic Pseudo-nitzschia multistriata (Bacillariophyceae) from the Gulf of Naples: Morphology, toxin analysis and phylogenetic relationships with other Pseudo-nitzschia species. Eur. J. Phycol. 2002, 37, 247–257. [Google Scholar]
- Stehr, C.M.; Connell, L.; Baugh, K.A.; Bill, B.D.; Adams, N.G.; Trainer, V.L. Morphological, toxicological, and genetic differences among Pseudo-nitzschia (Bacillariophyceae) species in inland embayments and outer coastal waters of Washington State, USA. J. Phycol. 2002, 38, 55–65. [Google Scholar] [CrossRef]
- Bates, S.S.; Garrison, D.L.; Horner, R.A. Bloom Dynamics and Physiology of Domoic Acid Producing Species of Pseudonitzschia Species. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer-Verlag: Berlin, Germany, 1998; Volume G 41, pp. 267–292. NATO ASI Series. [Google Scholar]
- Zingone, A.; Siano, R.; D’Alelio, D.; Sarno, D. Potentially toxic and harmful microalgae from coastal waters of the Campania region (Tyrrhenian Sea, Mediterranean Sea). Harmful Algae 2006, 5, 321–337. [Google Scholar]
- Rhodes, L.; Adamson, J.; Scholin, C. Pseudo-nitzschia multistriata (Bacillariophyceae) in New Zealand. J. Mar. Fresh. Res. 2000, 34, 463–467. [Google Scholar]
- Guerrini, F. 2008; University of Bologna, Italy. Unpublished results.
- Pistocchi, R.; Pezzolesi, L.; Guerrini, F.; Vanucci, S.; Dell’Aversano, C.; Fattorusso, E. A review of the effect of environmental conditions on growth and toxin production of Ostreopsis ovata. Toxicon 2011, 57, 421–428. [Google Scholar]
- Pezzolesi, L.; Guerrini, F.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Pistocchi, R. Influence of temperature and salinity on Ostreopsis cf. ovata growth and evaluation of toxin content through HR LC-MS and biological assays. Water Res. 2012, 46, 82–92. [Google Scholar]
- Guerrini, F.; Pezzolesi, L.; Feller, A.; Riccardi, M.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; et al. Comparative growth and toxin profile of cultured Ostreopsis ovata from the Tyrrhenian and Adriatic seas. Toxicon 2010, 55, 211–220. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pistocchi, R. Complex palitoxin-like profile of Ostreopsis ovata. Identification of four new ovatoxins by high resolution LC-MS. Rapid Commun. Mass Spectrom. 2010, 24, 2735–2744. [Google Scholar]
- Pezzolesi, L. 2009; University of Bologna, Italy. Unpublished results.
- de Boer, M.K.; Tyl, M.R.; Vrieling, E.G.; van Rijssel, M. Effects of salinity and nutrients on growth and haemolytic activity of Fibrocapsa japonica (Raphidophyceae). Aquat. Microb. Ecol. 2004, 37, 171–181. [Google Scholar]
- Fu, M.; Koulman, A.; van Rijssel, M.; Lützen, A.; de Boer, M.K.; Tyl, M.R.; Liebezeit, G. Chemical characterization of three haemolytic compounds from the microalgal species Fibrocapsa japonica (Raphidophyceae). Toxicon 2004, 43, 355–363. [Google Scholar]
- Oda, T.; Nakamura, A.; Shikayama, M.; Kawano, I.; Ishimatsu, A.; Muramatsu, T. Generation of reactive oxygen species by raphidophycean phytoplankton. Biosci. Biotechnol. Biochem. 1997, 61, 1658–1662. [Google Scholar]
- Marshall, J.A.; Nichols, P.D.; Hamilton, B.; Lewis, R.J.; Hallegraeff, G.M. Ichthyotoxicity of Chattonella marina (Raphidophyceae) to damselfish (Acanthochromis polycanthus): The synergistic role of reactive oxygen species and free fatty acids. Harmful Algae 2003, 2, 273–281. [Google Scholar]
- Khan, S.; Arakawa, O.; Onoue, Y. Neurotoxin production by a chloromonad Fibrocapsa japonica (Raphidophyceae). J. World Aquat. Soc. 1996, 27, 254–263. [Google Scholar]
- Woo, S.P.S.; Liu, W.; Au, D.W.T.; Anderson, D.M.; Wu, R.S.S. Antioxidant responses and lipid peroxidation in gills and erythrocytes of fish (Rhabdosarga sarba) upon exposure to Chattonella marina and hydrogen peroxide: Implication on the cause of fish kills. J. Exp. Mar. Biol. Ecol. 2006, 336, 230–241. [Google Scholar]
- Pezzolesi, L. Harmful Algae and Their Potential Impacts on the Coastal Ecosystem: Growth and Toxin Production Dynamics. Ph.D. Dissertation, University of Bologna, Italy, 2011. [Google Scholar]
- Draisci, R.; Lucentini, L.; Giannetti, L.; Boria, P.; Poletti, R. First report of pectenotoxin-2 (PTX-2) in algae (Dinophysis fortii) related to seafood poisoning in Europe. Toxicon 1996, 34, 923–935. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Morino, M.; Magno, S.; Poletti, R.; Viviani, R. Isolation of adriatoxin, a new analogue of yessotoxin from mussels of the Adriatic Sea. Tetrahedron Lett. 1998, 39, 8897–8900. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Forino, M. Structural elucidation of a new cytotoxin isolated from mussels of the Adriatic Sea. J. Org. Chem. 2001, 66, 578–582. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, S. Toxins from Adriatic blue mussels. A decade of studies. Pure Appl. Chem. 2003, 75, 325–336. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Boschetti, L.; Rubini, S.; Cangini, M.; Pigozzi, S.; Poletti, R. Complex toxin profile of Mytilus galloprovincialis from the Adriatic Sea revealed by LC-MS. Toxicon 2010, 55, 280–288. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pistocchi, R.; Guerrini, F.; Pezzolesi, L.; Riccardi, M.; Vanucci, S.; Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Fattorusso, E.; Tartaglione, L.; et al. Toxin Levels and Profiles in Microalgae from the North-Western Adriatic Sea—15 Years of Studies on Cultured Species. Mar. Drugs 2012, 10, 140-162. https://doi.org/10.3390/md10010140
Pistocchi R, Guerrini F, Pezzolesi L, Riccardi M, Vanucci S, Ciminiello P, Dell’Aversano C, Forino M, Fattorusso E, Tartaglione L, et al. Toxin Levels and Profiles in Microalgae from the North-Western Adriatic Sea—15 Years of Studies on Cultured Species. Marine Drugs. 2012; 10(1):140-162. https://doi.org/10.3390/md10010140
Chicago/Turabian StylePistocchi, Rossella, Franca Guerrini, Laura Pezzolesi, Manuela Riccardi, Silvana Vanucci, Patrizia Ciminiello, Carmela Dell’Aversano, Martino Forino, Ernesto Fattorusso, Luciana Tartaglione, and et al. 2012. "Toxin Levels and Profiles in Microalgae from the North-Western Adriatic Sea—15 Years of Studies on Cultured Species" Marine Drugs 10, no. 1: 140-162. https://doi.org/10.3390/md10010140
APA StylePistocchi, R., Guerrini, F., Pezzolesi, L., Riccardi, M., Vanucci, S., Ciminiello, P., Dell’Aversano, C., Forino, M., Fattorusso, E., Tartaglione, L., Milandri, A., Pompei, M., Cangini, M., Pigozzi, S., & Riccardi, E. (2012). Toxin Levels and Profiles in Microalgae from the North-Western Adriatic Sea—15 Years of Studies on Cultured Species. Marine Drugs, 10(1), 140-162. https://doi.org/10.3390/md10010140