Drugs from the Sea - Opportunities and Obstacles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction
The Supply Problem
Opportunities of Invertebrate Mariculture
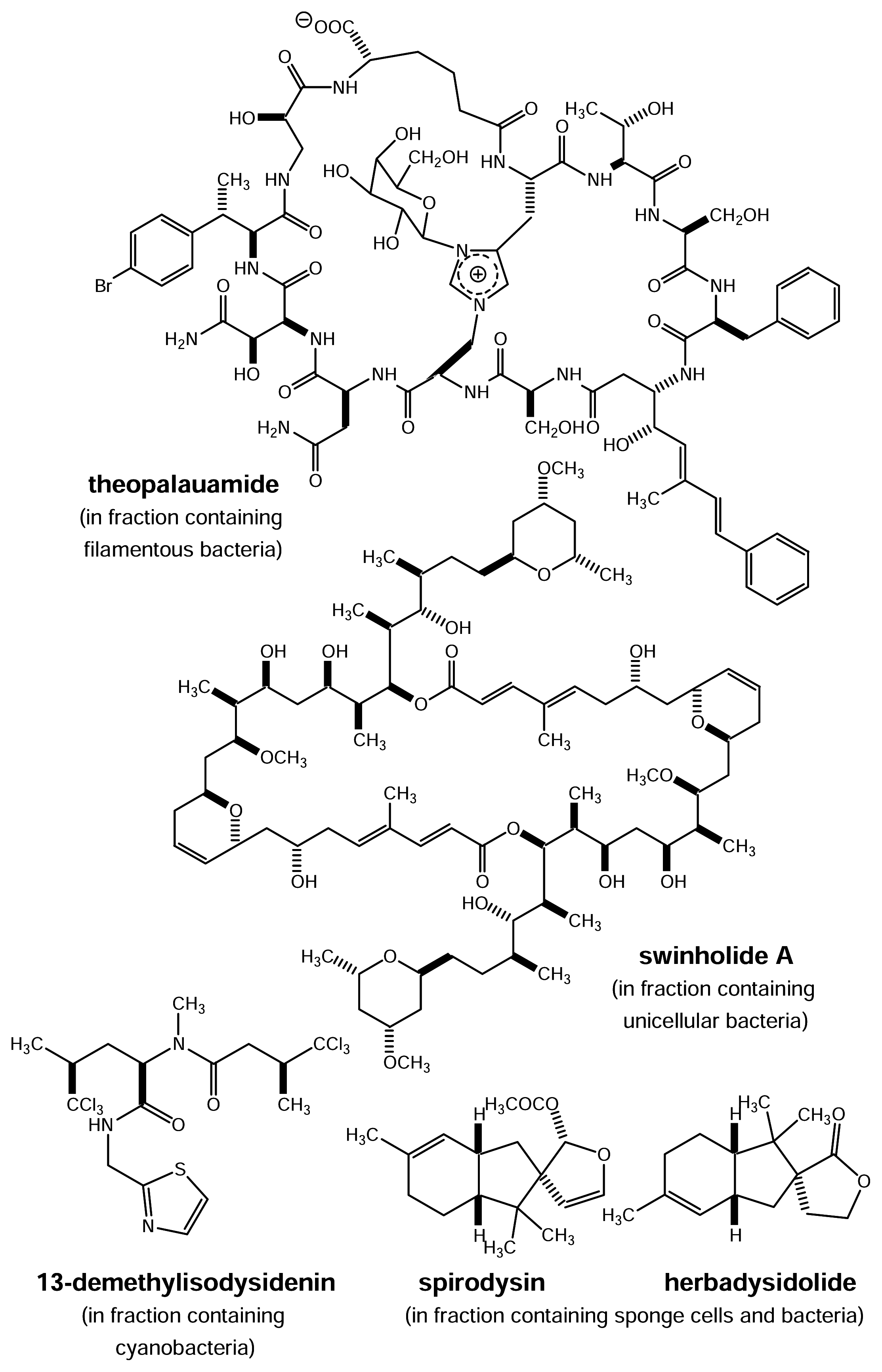
Arguments for a Microbial Origin of Bioactive Natural Products from Marine Invertebrates
Can the putative microbial producers of bioactive invertebrate metabolites provide a solution to the supply problem?





Acknowlegements
References and Notes
- Newman, D. J.; Cragg, G. M.; Snader, K. M. The influence of natural products upon drug discovery. Nat. Prod. Rep 2000, 17, 215–234. [Google Scholar]
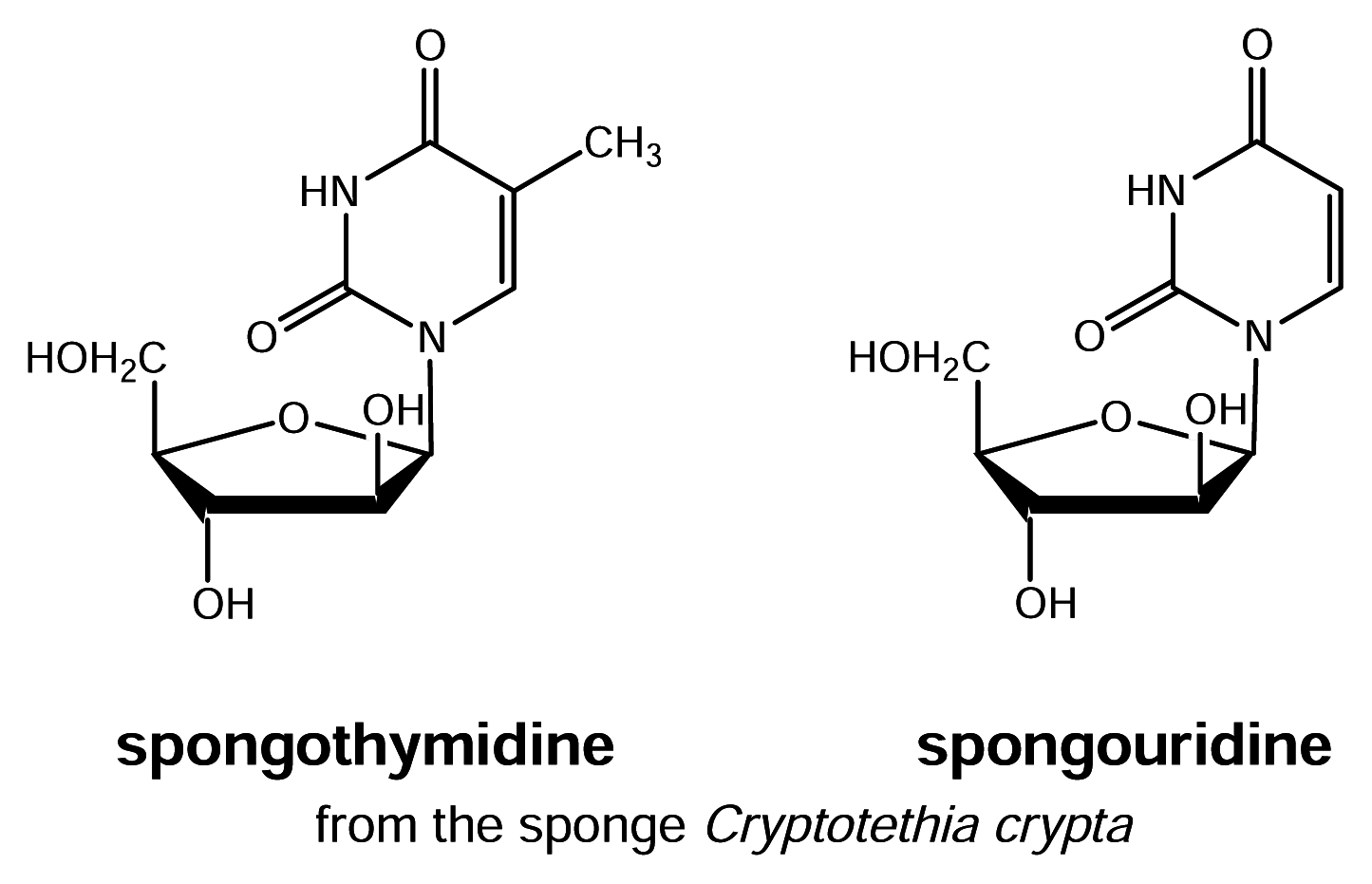
- Bergmann, W.; Feeney, R. Contribution to the study of marine sponges. 32. The nucleosides of sponges. J. Org. Chem 1951, 16, 981–987. [Google Scholar]
- MarinLit, Version September 2003. A marine literature database produced and maintained by the Department of Chemistry, University of Canterbury, New Zealand.
- Kerr, R. G.; Kerr, S. S. Marine natural products as therapeutic agents. Expert Opin. Ther. Pat 1999, 9, 1207–1222. [Google Scholar]
- Proksch, P.; Edrada, R. A.; Ebel, R. Drugs from the Seas – Current Status and Microbiological Implications. Appl. Microbiol. Biotech 2002, 59, 125–134. [Google Scholar]
- Cimino, G.; De Rosa, S.; De Stefano, S. Antiviral agents from a gorgonian Eunicella cavolini; one previously found in the sponge Cryptotehya crypta. Experientia 1984, 40, 339–340. [Google Scholar]
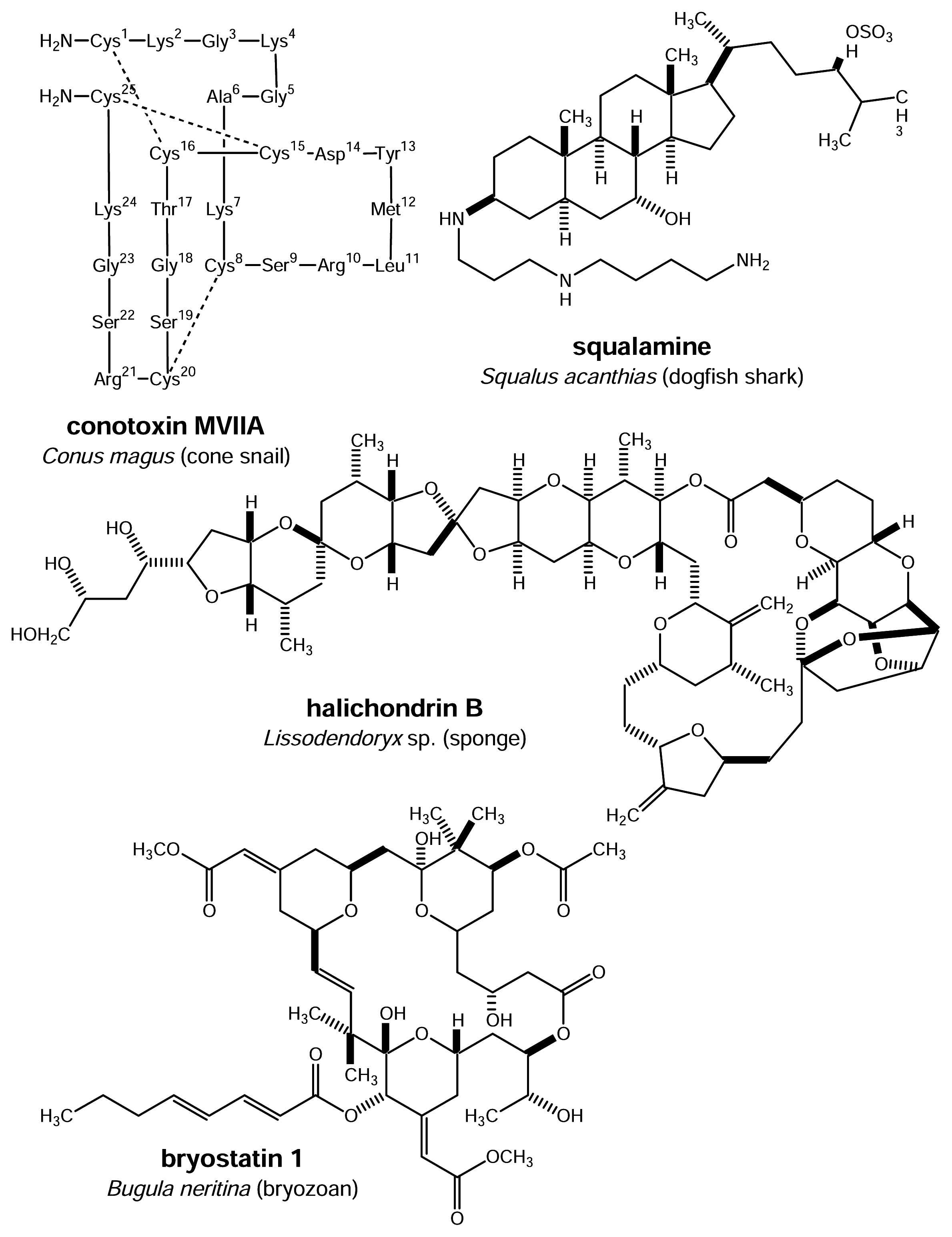
- Mendola, D. Aquacultural production of bryostatin 1 and ecteinascidin 743. In Drugs from the sea; Fusetani, N., Ed.; Karger: Basel, 2000; pp. 120–133. [Google Scholar]
- Hart, J. B.; Lill, R. E.; Hickford, S. J. H.; Blunt, J. W.; Munro, M. H. G. The halichondrins: Chemistry, biology, supply and delivery. In Drugs from the sea; Fusetani, N., Ed.; Karger: Basel, 2000; pp. 134–153. [Google Scholar]
- Olivera, B. M. ω-conotoxin MVIIA: From marine snail venom to analgesic drug. In Drugs from the sea; Fusetani, N., Ed.; Karger: Basel, 2000; pp. 74–85. [Google Scholar]
- Mutter, R.; Wills, M. Chemistry and clinical biology of the bryostatins. Bioorg. Med. Chem 2000, 8, 1841–1860. [Google Scholar]
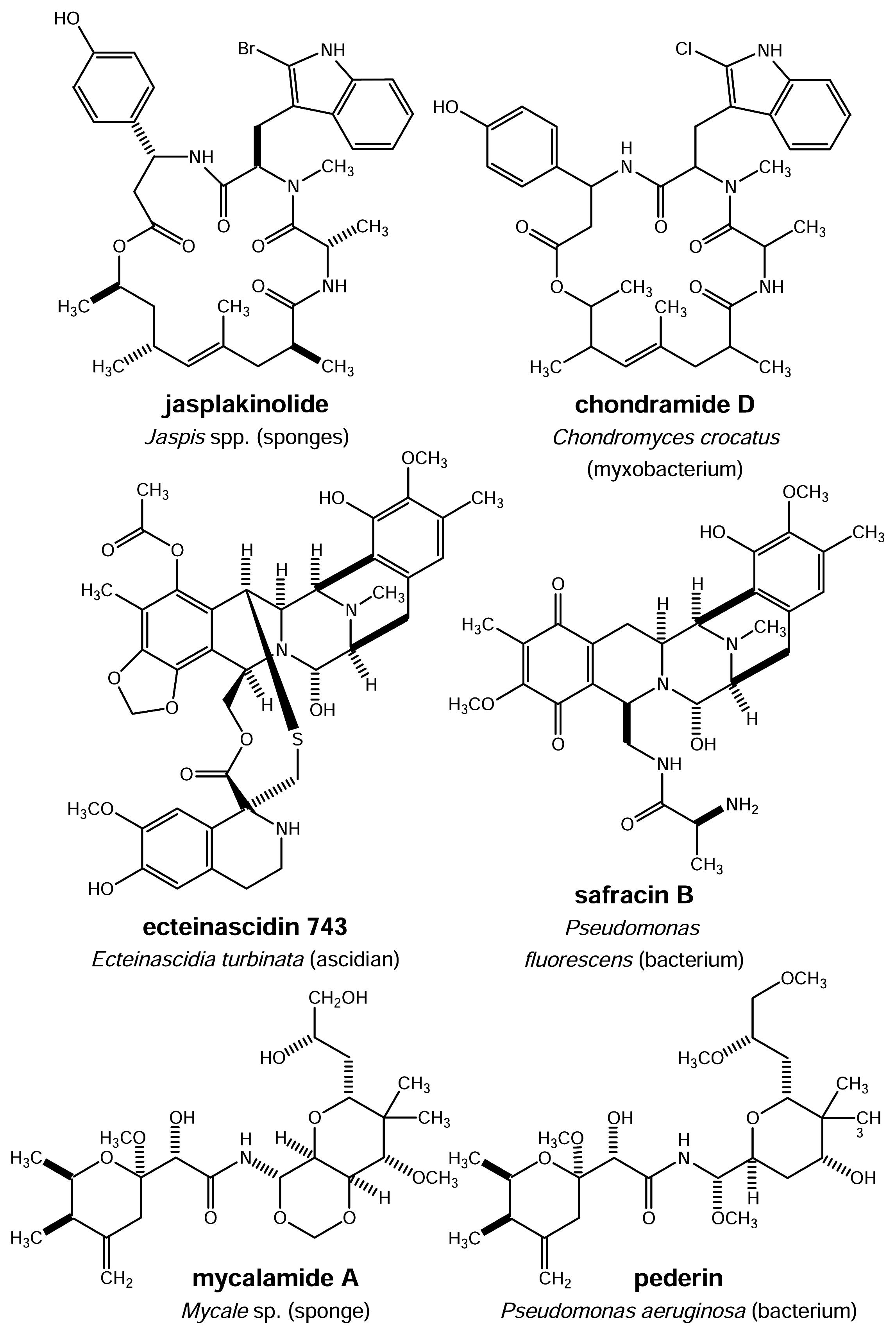
- Bubb, M. R.; Senderowicz, A. M.; Sausville, E. A.; Duncan, K. L.; Korn, E. D. Jasplakinolide, a cytotoxic natural product, induces actin polymerization and competitively inhibits the binding of phalloidin to F-actin. J. Biol. Chem 1994, 269, 14869–14871. [Google Scholar]
- Jansen, R.; Kunze, B.; Reichenbach, H.; Höfle, G. Chondramides A – D, new cytostatic and antifungal cyclodepsipeptides from Chondromyces crocatus (Myxobacteria): Isolation and structure elucidation. Liebigs Ann 1996, 2, 285–290. [Google Scholar]
- Ikeda, Y.; Matsuki, H.; Ogawa, T.; Munakata, T. Safracins, new antitumor antibiotics. II. Physicochemical properties and chemical structures. J. Antibiot 1983, 36, 1284–1289. [Google Scholar]
- Cuevas, C.; Pérez, M.; Martin, M. J.; Chicharro, J. L.; Fernández-Rivas, C.; Flores, M.; Francesch, A.; Gallego, P.; Zarzuelo, M.; de la Calle, F.; Garcia, J.; Polanco, C.; Rodriguez, I.; Manzanares, I. Synthesis of ecteinascidin ET-743 and phthalascidin Pt-650 from cyanosafracin B. Org. Lett 2000, 2, 2545–2548. [Google Scholar]
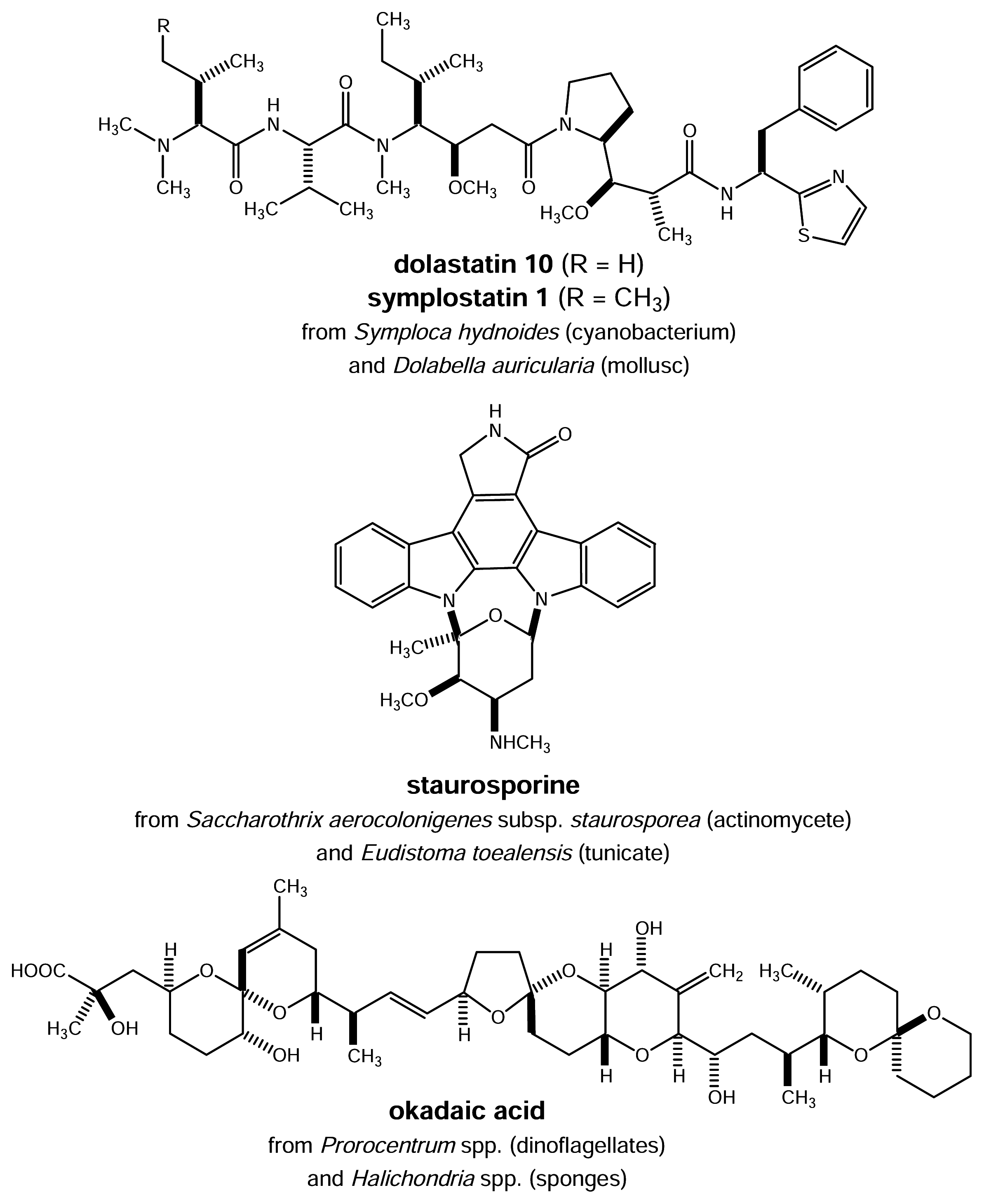
- Harrigan, G. G.; Luesch, H.; Yoshida, W. Y.; Moore, R. E.; Nagle, D. G.; Paul, V. J.; Mooberry, S. L.; Corbett, T. H.; Valeriote, F. A. Symplostatin 1. A dolastatin 10 analogue from marine cyanobacterium Symploca hydnoides. J. Nat. Prod 1998, 61, 1075–1077. [Google Scholar]
- Luesch, H.; Moore, R. E.; Paul, V. J.; Mooberry, S. L.; Corbett, T. H. Isolation of dolastatin 10 from the marine cyanobacterium Symploca species VP642 and total stereochemistry and biological evaluation of its analogue symplostatin 1. J. Nat. Prod 2001, 64, 907–910. [Google Scholar]
- Luesch, H.; Harrigan, G. G.; Goetz, G.; Horgen, F. D. The cyanobacterial origin of potent anticancer agents originally isolated from sea hares. Curr. Med. Chem 2002, 9, 1791–1806. [Google Scholar]
- Schupp, P.; Wray, V.; Eder, C.; Schneider, P.; Herderich, M.; Paul, V.; Proksch, P. Staurosporine derivatives from the ascidican Eudistoma toealensis and its predatory flatworm Pseudoceros sp. J. Nat. Prod 1999, 62, 959–962. [Google Scholar]
- Murakami, Y.; Oshima, Y.; Yasumoto, T. Identification of okadaic acid as a toxic component of a marine dinoflagellate Prorocentrum lima. Bull. Jp. Soc. Fish Sci 1982, 48, 69–72. [Google Scholar]
- Bewley, C. A.; Holland, N. D.; Faulkner, D. J. Two classes of metabolites from Theonella swinhoei are localized in distinct populations of bacterial symbionts. Experientia 1996, 52, 716–722. [Google Scholar]
- Bewley, C. A.; Faulkner, D. J. Lithistid sponges: Star performers or hosts to the stars? Angew. Chem. Int. Ed 1998, 37, 2162–2178. [Google Scholar]
- Schmidt, E. W.; Obraztsova, A. Y.; Davidson, S. K.; Faulkner, D. J.; Haygood, M. G. Identification of the antifungal peptide-containing symbiont of the marine sponge Theonella swinhoei as a novel δ-proteobacterium, “Candidatus Entotheonella palauensis”. Mar. Biol 2000, 136, 969–977. [Google Scholar]
- Unson, M. D.; Faulkner, D. J. Cyanobacterial symbiont biosynthesis of chlorinated metabolites from Dysidea herbacea (Porifera). Experientia 1993, 49, 349–353. [Google Scholar]
- Flowers, A. E.; Garson, M. J.; Webb, R. I.; Dumdei, E. J.; Charan, R. D. Cellular origin of chlorinated diketopiperazines in the dictyoceratid sponge Dysidea herbacea (Keller). Cell Tissue Res 1998, 292, 597–607. [Google Scholar]
- Friedrich, A. B.; Fischer, I.; Proksch, P.; Hacker, J.; Hentschel, U. Temporal variation of the microbial community associated with the Mediterranean sponge Aplysina aerophoba. FEMS Microbiol. Ecol 2001, 1301, 1–9. [Google Scholar]
- Piel, J. A polyketide synthase-peptide synthetase gene cluster from an uncultured bacterial symbiont of Paederus beetles. Proc. Natl. Acad. Sci. USA 2002, 99, 14002–14007. [Google Scholar]
- Narquizian, R.; Kocienski, P. J. The pederin family of antitumor agents: structures, synthesis and biological activity. Ernst Schering Res. Found. Workshop 2000, 32, 25–56. [Google Scholar]
- Perry, N. B.; Blunt, J. W.; Munro, M. H. G.; Pannell, L. K. Mycalamide A, an antiviral compound from a New Zealand sponge of genus Mycale. J. Am. Chem. Soc 1988, 110, 4850–4851. [Google Scholar]
- Hood, K. A.; West, L. M.; Northcote, P. T.; Berridge, M. V.M.; Miller, J. H. Induction of apoptosis by the marine sponge (Mycale) metabolites, mycalamide A and pateamine. Apoptosis 2001, 6, 207–219. [Google Scholar]
- Kellner, R. L. Molecular identification of an endosymbiotic bacterium associated with pederin biosynthesis in Paederus sabaeus (Coleoptera: Staphylinidae). Insect Biochem. Mol. Biol 2002, 32, 389–395. [Google Scholar]
- Kerr, R. G. Biosynthesis of bioactive marine natural products. In Studies in Natural Products Chemistry; Volume 21, Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, 2000; pp. 293–328. [Google Scholar]
- Garson, M. J.; Simpson, J. S.; Flowers, A. E.; Dumdei, E. J. Cyanide and thiocyanate-derived functionality in marine organisms -structures, biosynthesis and ecology. In Studies in Natural Products Chemistry; Volume 21, Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, 2000; pp. 329–372. [Google Scholar]
© 2003 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Proksch, P.; Edrada-Ebel, R.; Ebel, R. Drugs from the Sea - Opportunities and Obstacles. Mar. Drugs 2003, 1, 5-17. https://doi.org/10.3390/md101005
Proksch P, Edrada-Ebel R, Ebel R. Drugs from the Sea - Opportunities and Obstacles. Marine Drugs. 2003; 1(1):5-17. https://doi.org/10.3390/md101005
Chicago/Turabian StyleProksch, Peter, RuAngelie Edrada-Ebel, and Rainer Ebel. 2003. "Drugs from the Sea - Opportunities and Obstacles" Marine Drugs 1, no. 1: 5-17. https://doi.org/10.3390/md101005
APA StyleProksch, P., Edrada-Ebel, R., & Ebel, R. (2003). Drugs from the Sea - Opportunities and Obstacles. Marine Drugs, 1(1), 5-17. https://doi.org/10.3390/md101005