
Resveratrol and Coumarin: Novel Agricultural Antibacterial Agent against Ralstonia solanacearum In Vitro and In Vivo

Abstract
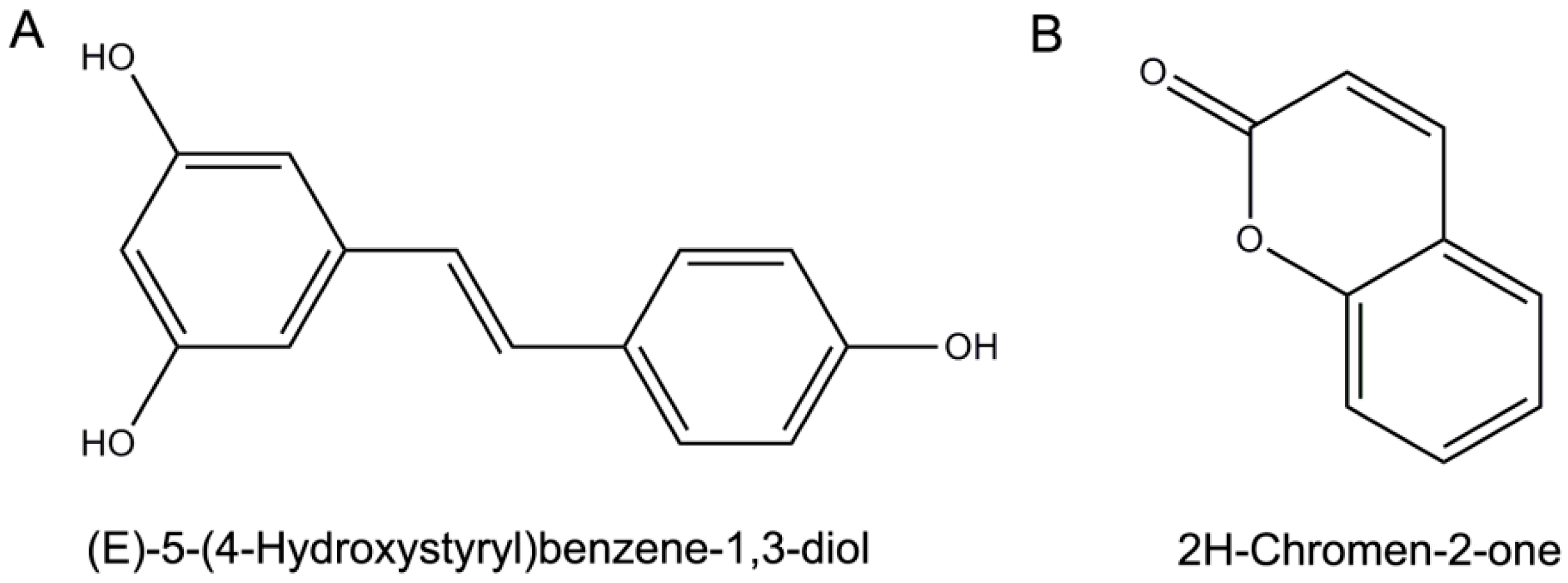
:1. Introduction
2. Results
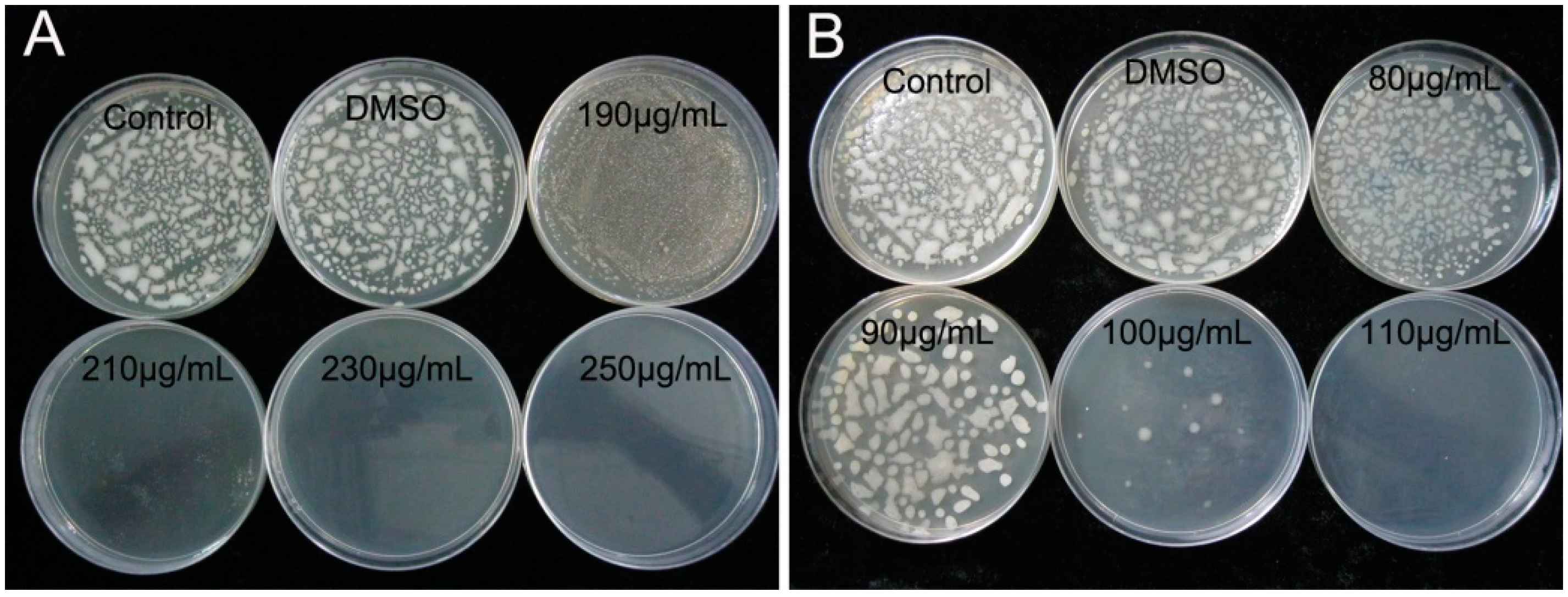
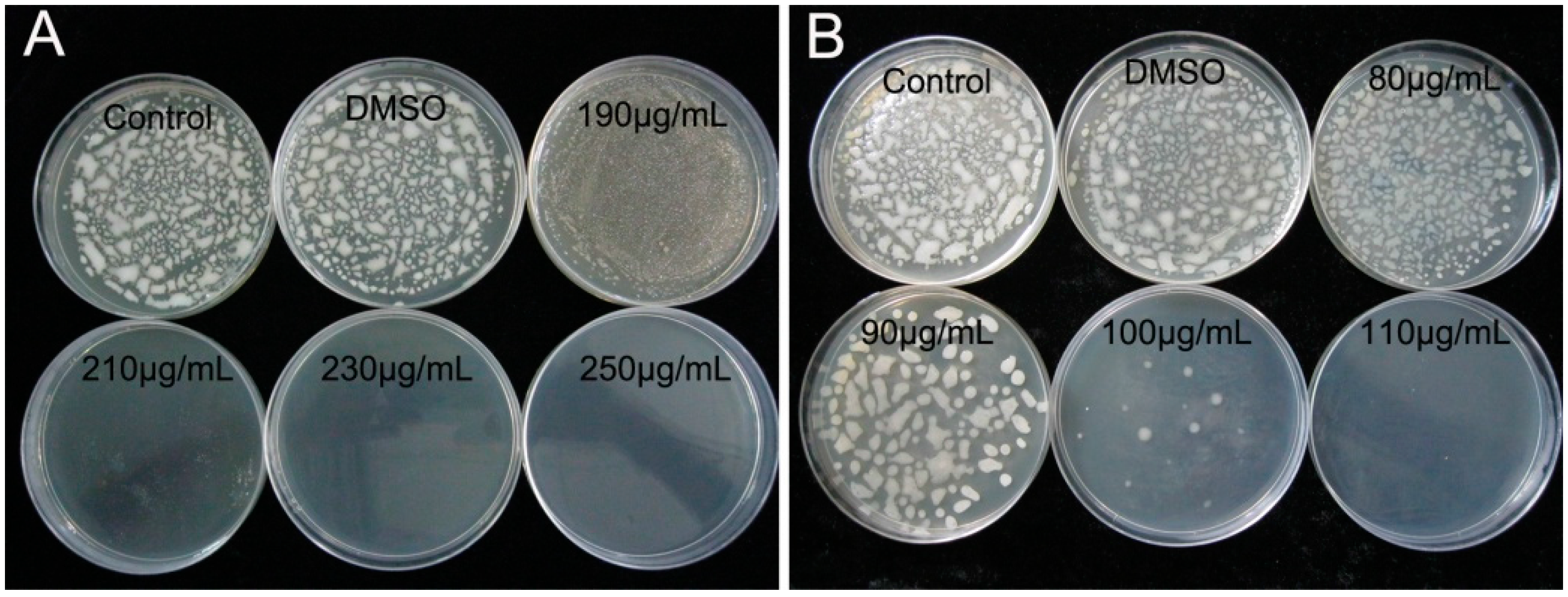
2.1. Determination of MIC and MBC
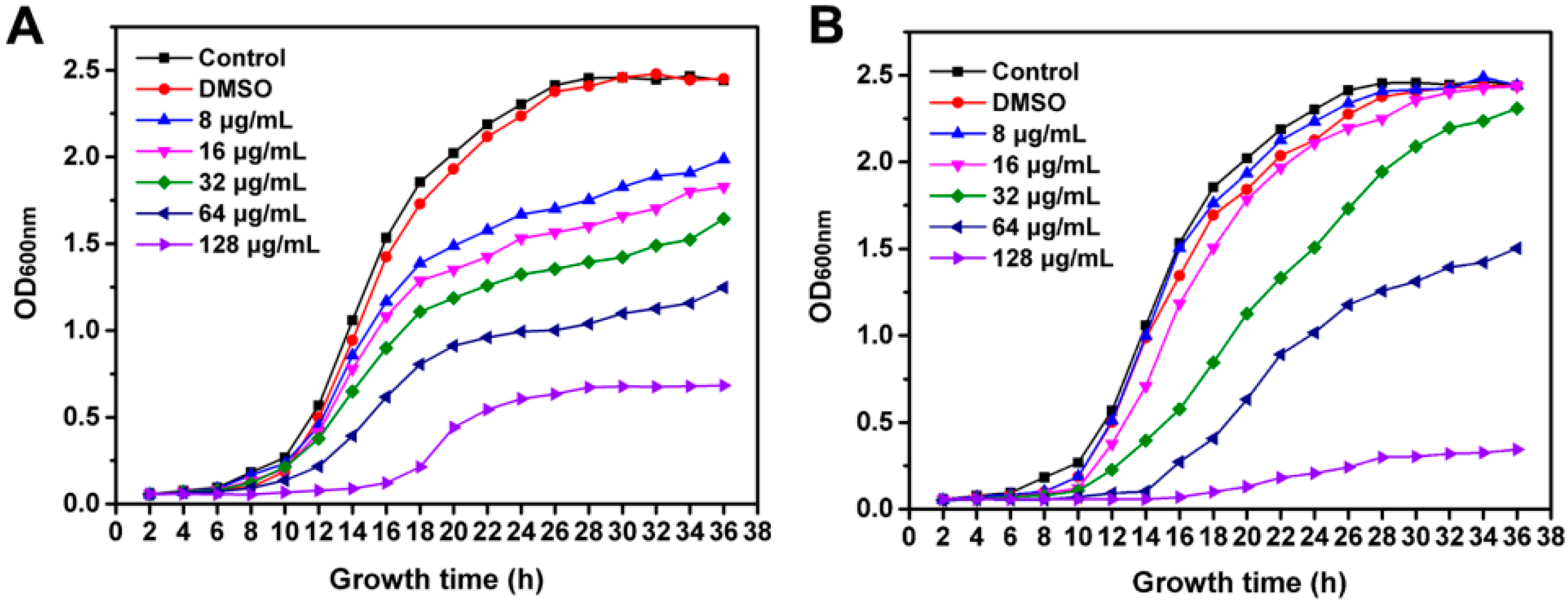
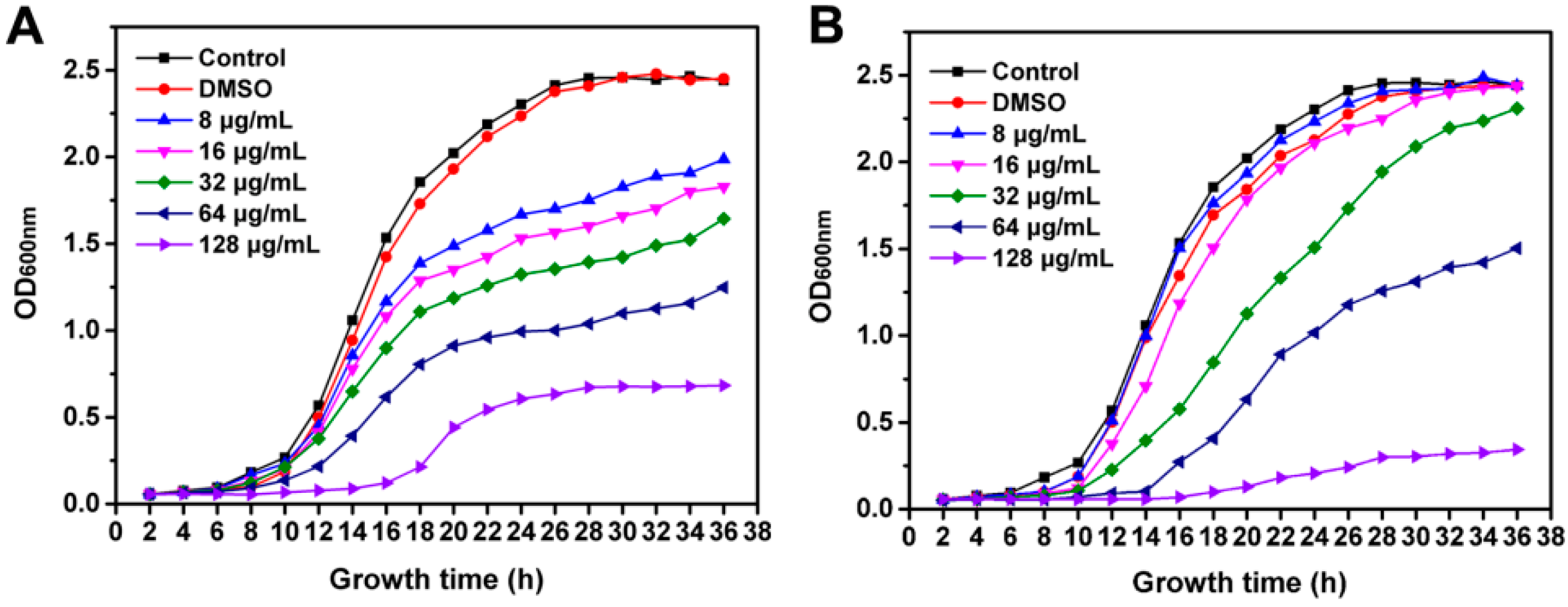
2.2. Inhibition of R. solanacearum Growth
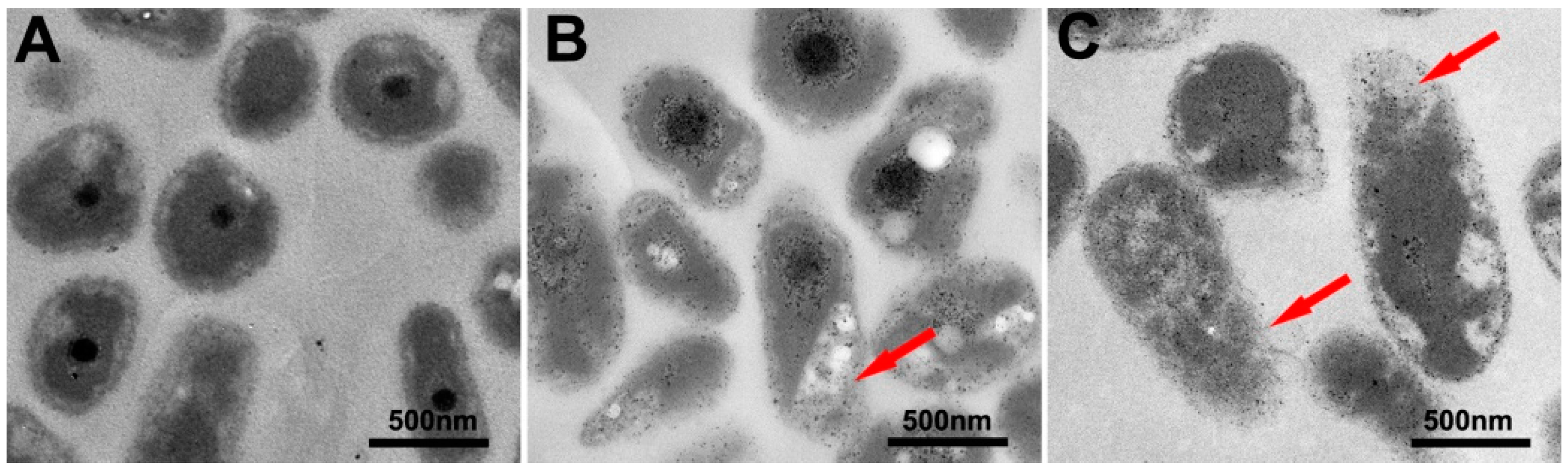
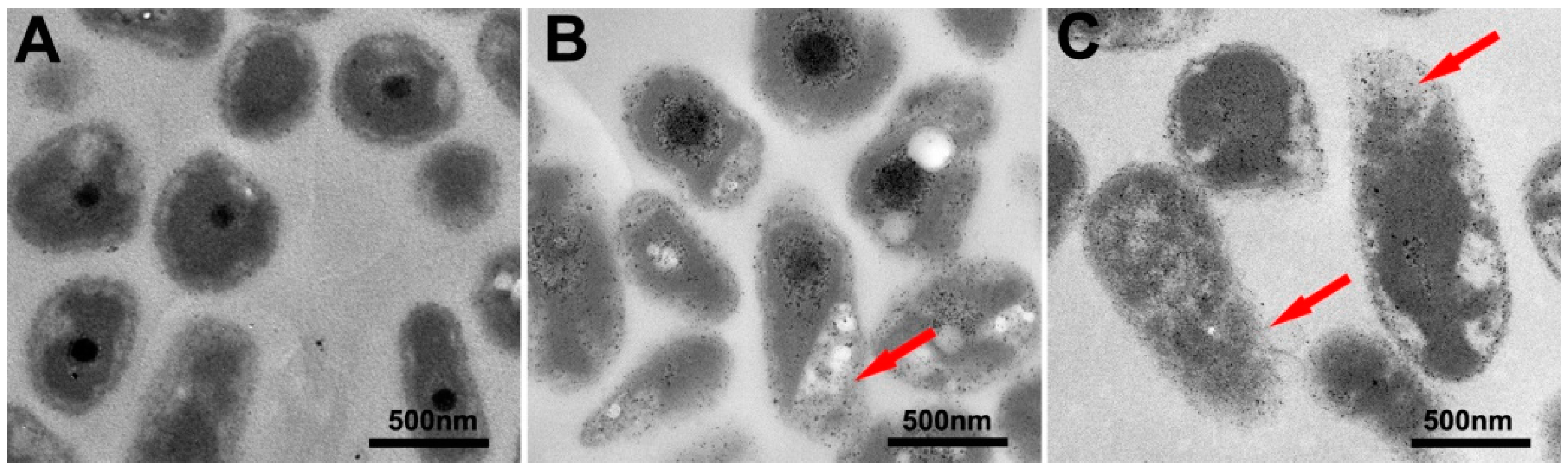
2.3. Bacterial Cell Morphology Observations
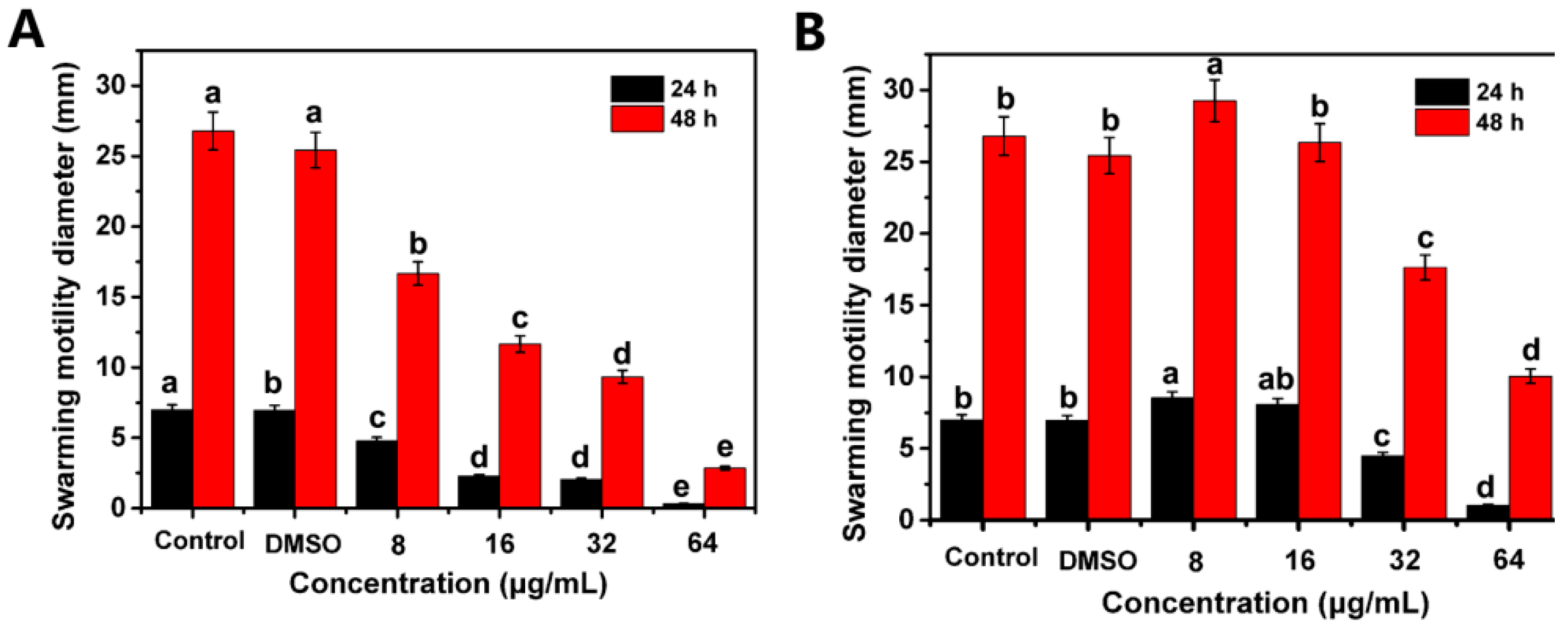
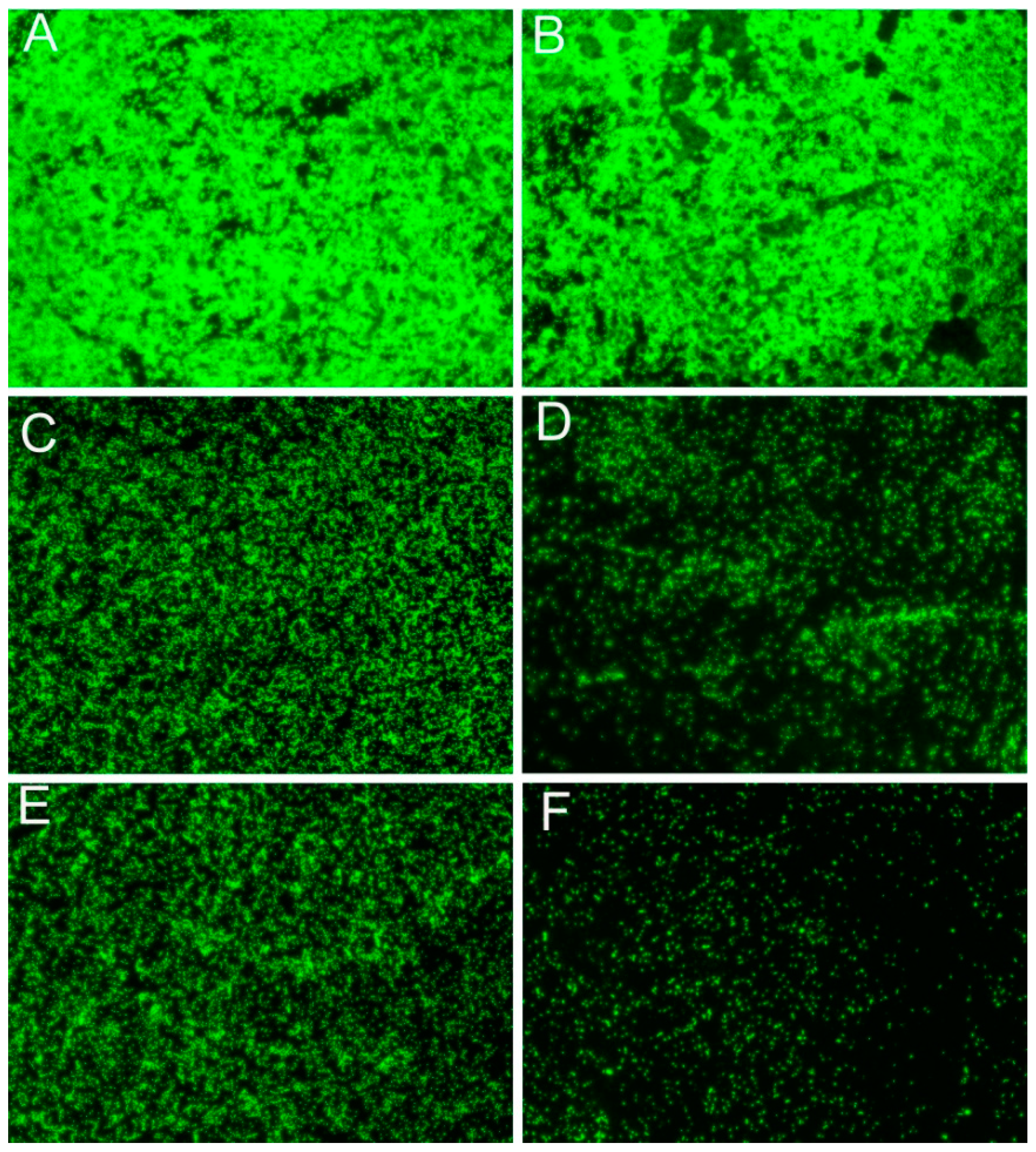
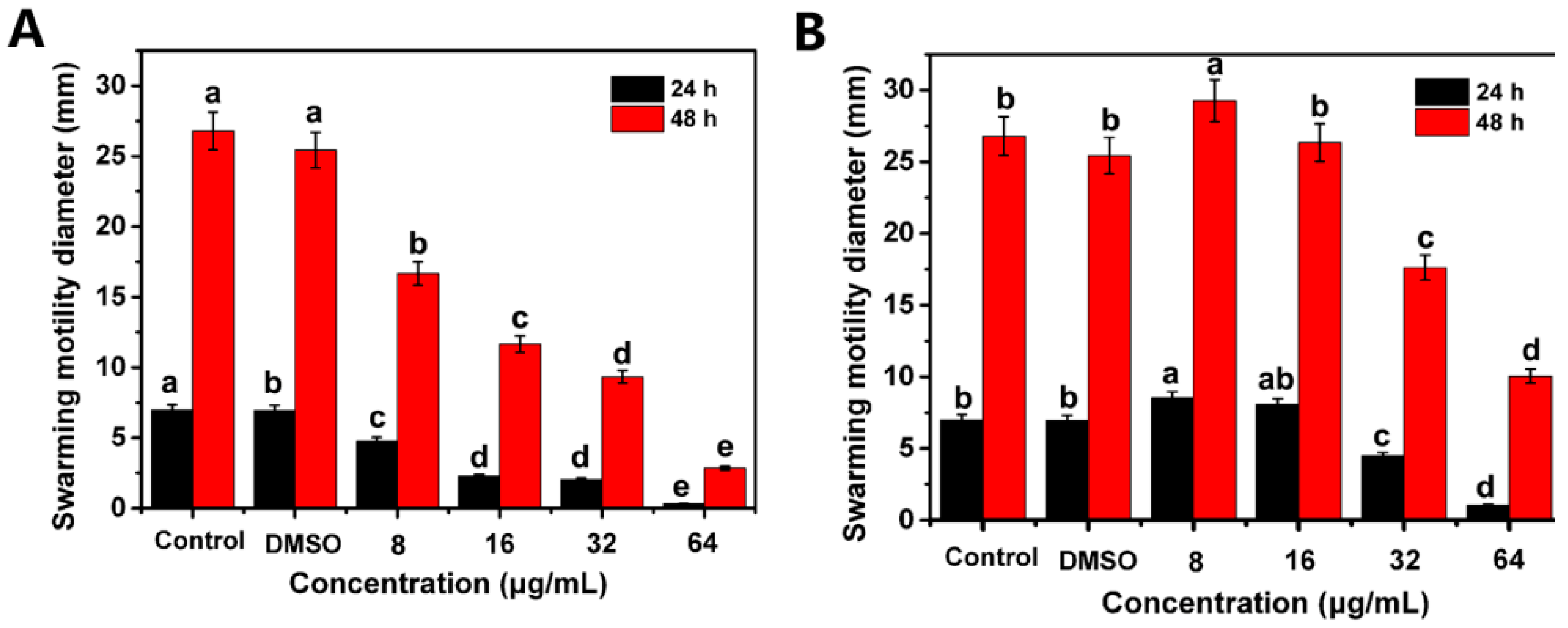
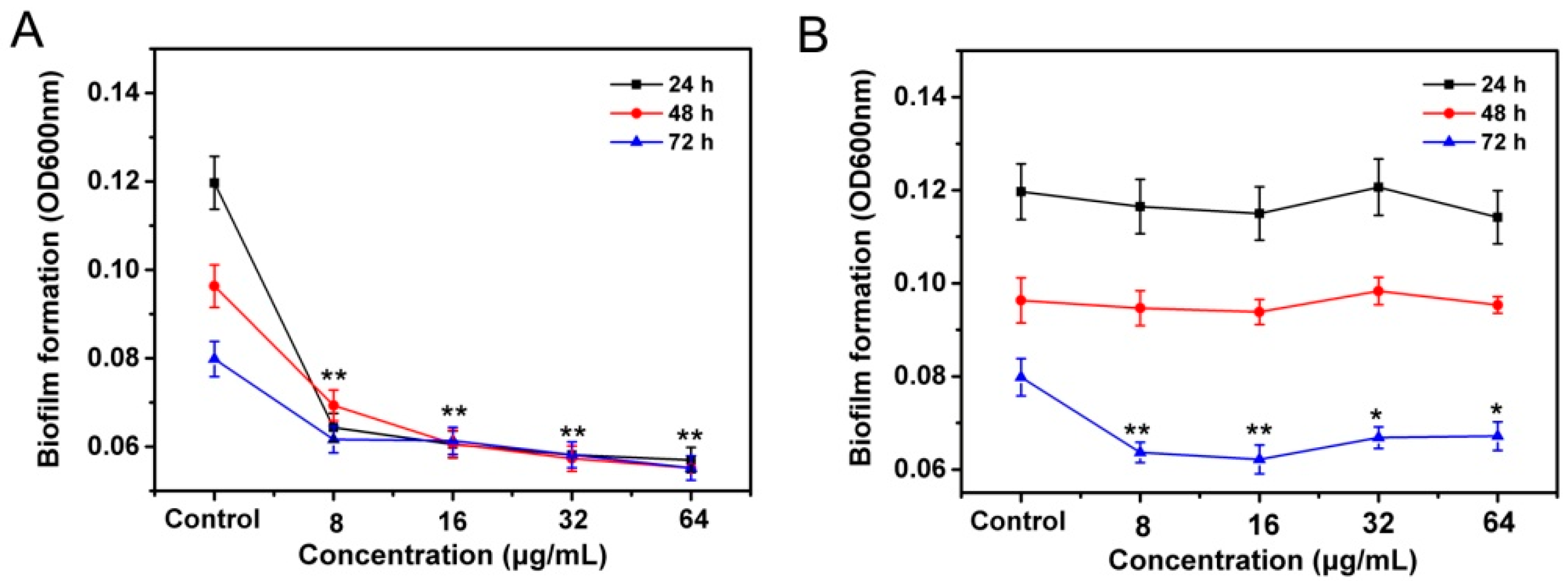
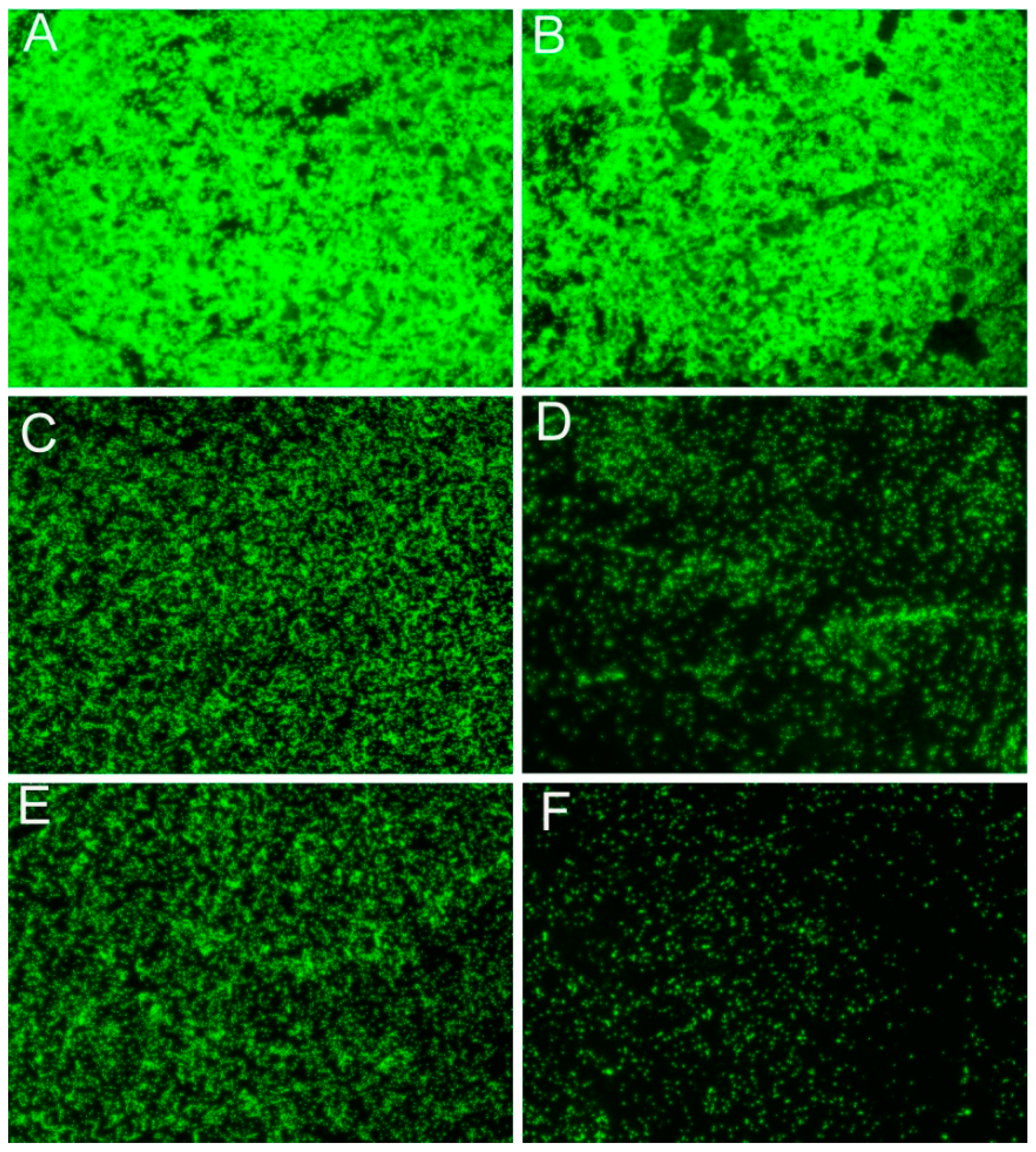
2.4. Suppress the Swarming Motility and Biofilm Formation of R. solanacearum
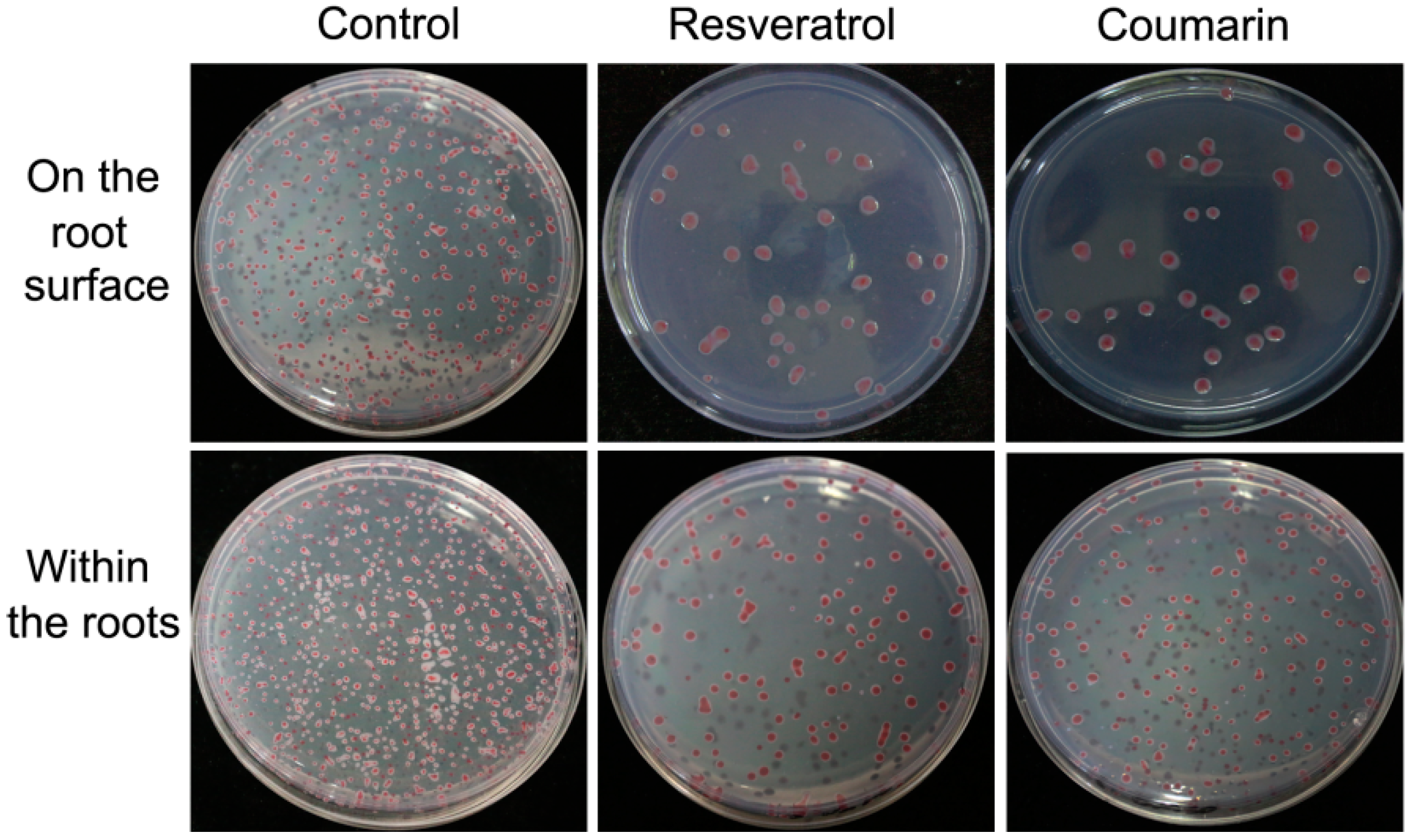
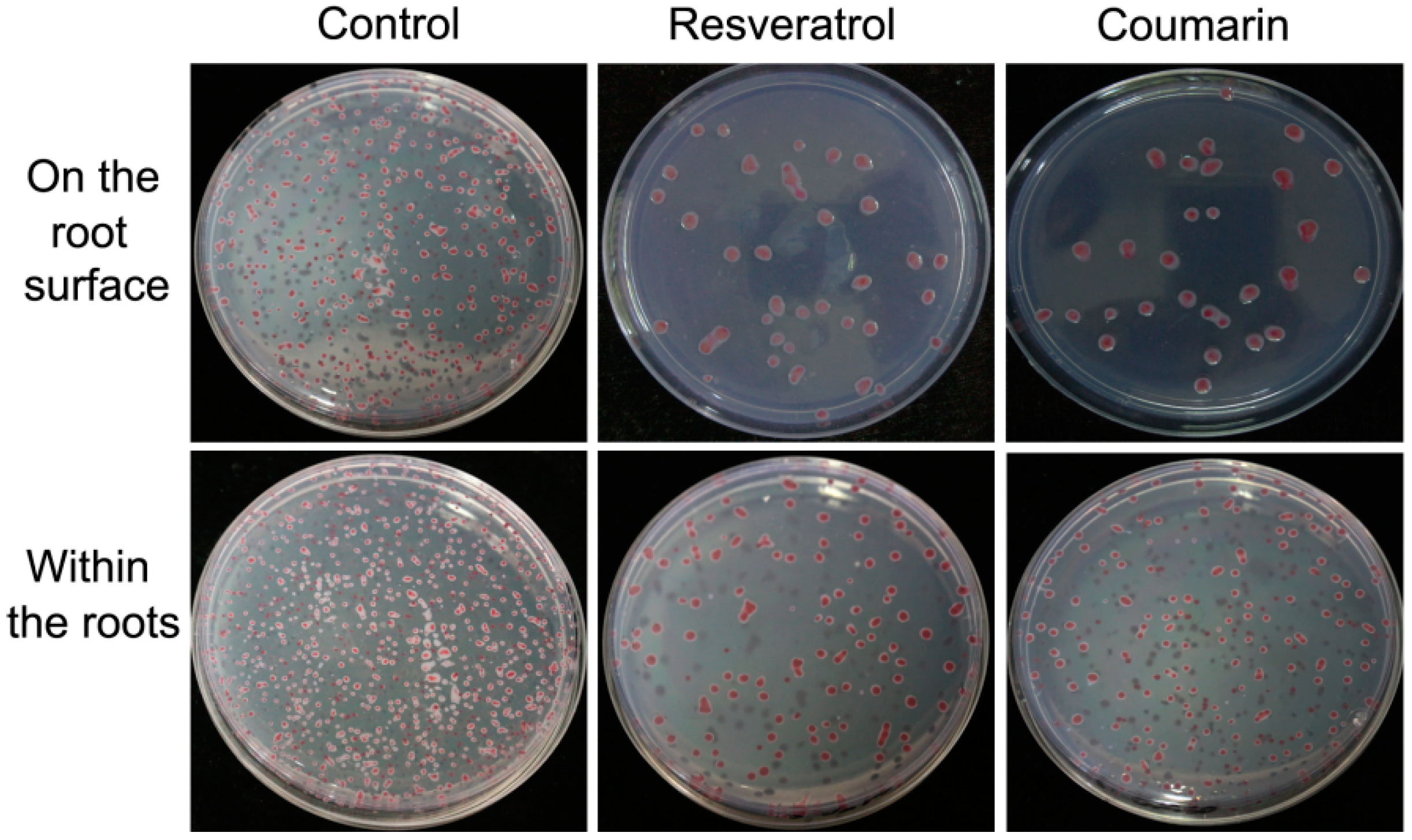
2.5. Effects of Phytochemicals on Bacterial Adhesion and Colonization
2.6. Control Efficacy of Phytochemicals to Bacterial Wilt of Tobacco
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Conditions
4.2. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Assays
4.3. Growth Curve Assay
4.4. Observation of Bacterial Cell Morphology by TEM
4.5. Bacterial Motility Assay
4.6. Bacterial Biofilm Formation Assay
4.7. Biofilm Observations by Fluorescence Microscopy
4.8. Bacterial Adhesion Test
4.9. Controlling Tobacco Bacterial Wilt In Vivo under Greenhouse Conditions
4.10. Statistic Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hayward, A.C. Biology and epidemiology of bacterial wilt caused by Pseudomonas-solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Buddenhagen, I.; Kelman, A. Biological and physiological aspects of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 1964, 2, 203–230. [Google Scholar] [CrossRef]
- Yi, Y.J.; Liu, R.S.; Yin, H.Q.; Luo, K.; Liu, E.M.; Liu, X.D. Isolation, identification and field control efficacy of an endophytic strain against tobacco bacterial wilt (Ralstonia solanacarum). Chin. J. Appl. Ecol. 2007, 18, 554–558. (in Chinese). [Google Scholar]
- Zhang, S.; Raza, W.; Yang, X.; Hu, J.; Huang, Q.; Xu, Y.; Liu, X.; Ran, W.; Shen, Q. Control of Fusarium wilt disease of cucumber plants with the application of a bioorganic fertilizer. Biol. Fert. Soils 2008, 44, 1073–1080. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Liu, W.; Levi, A. Grafting for disease resistance. Hort Sci. 2008, 43, 1673–1676. [Google Scholar]
- Zhang, N.; Wu, K.; He, X.; Li, S.Q.; Zhang, Z.H.; Shen, B.; Yang, X.M.; Zhang, R.F.; Huang, Q.W.; Shen, Q.R. A new bioorganic fertilizer can effectively control banana wilt by strong colonization with Bacillus subtilis N11. Plant Soil 2011, 344, 87–97. [Google Scholar] [CrossRef]
- Ren, X.; Zhang, N.; Cao, M.; Wu, K.; Shen, Q.; Huang, Q. Biological control of tobacco black shank and colonization of tobacco roots by a Paenibacillus polymyxa strain C5. Biol. Fert. Soils 2012, 48, 613–620. [Google Scholar] [CrossRef]
- Zheng, S.; Zhou, X.; Xu, S.; Zhu, R.; Bai, H.; Zhang, J. Synthesis and antimicrobial characterization of half-calycanthaceous alkaloid derivatives. Molecules 2016, 21, 1207. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.Y.; Cha, B.; Kim, J.C. Recent trends in studies on botanical fungicides in agriculture. Plant Pathol. J. 2013, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pradhanang, P.; Momol, M.; Olson, S.; Jones, J. Effects of plant essential oils on Ralstonia solanacearum population density and bacterial wilt incidence in tomato. Plant Dis. 2003, 87, 423–427. [Google Scholar] [CrossRef]
- Deberdt, P.; Perrin, B.; Coranson-Beaudu, R.; Duyck, P.F.; Wicker, E. Effect of Allium fistulosum extract on Ralstonia solanacearum populations and tomato bacterial wilt. Plant Dis. 2012, 96, 687–692. [Google Scholar] [CrossRef]
- Feng, J.; Yuan, F.H.; Gao, Y.; Liang, C.G.; Xu, J.; Zhang, C.L.; He, L.Y. A novel antimicrobial protein isolated from potato (Solanum tuberosum) shares homology with an acid phosphatase. Biochem. J. 2003, 376, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.; Ye, Y.; Wei, Z.; Wang, G.; Yang, H. Study on antimicrobial activities of α-(l)-terpineol against plant pathogens. Technol. Dev. Chem. Ind. 2007, 12, 5–7. [Google Scholar]
- Dorman, H.; Deans, S. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Shafi, P.M.; Nambiar, M.G.; Clery, R.A.; Sarma, Y.; Veena, S. Composition and antifungal activity of the oil of Artemisia nilagirica (Clarke) Pamp. J. Essent. Oil Res. 2004, 16, 377–379. [Google Scholar] [CrossRef]
- Paret, M.L.; Cabos, R.; Kratky, B.; Alvarez, A.M. Effect of plant essential oils on Ralstonia solanacearum race 4 and bacterial wilt of edible ginger. Plant Dis. 2010, 94, 521–527. [Google Scholar] [CrossRef]
- Ji, P.; Momol, M.; Olson, S.; Pradhanang, P.; Jones, J. Evaluation of thymol as biofumigant for control of bacterial wilt of tomato under field conditions. Plant Dis. 2005, 89, 497–500. [Google Scholar] [CrossRef]
- Fan, W.W.; Yuan, G.Q.; Li, Q.Q.; Lin, W. Antibacterial mechanisms of methyl gallate against Ralstonia solanacearum. Australas. Plant Path. 2014, 43, 1–7. [Google Scholar] [CrossRef]
- Li, L.; Feng, X.; Tang, M.; Hao, W.; Han, Y.; Zhang, G.; Wan, S. Antibacterial activity of Lansiumamide B to tobacco bacterial wilt (Ralstonia solanacearum). Microbiol. Res. 2014, 169, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Choi, C.W.; Kim, S.H.; Yun, J.G.; Chang, S.W.; Kim, Y.S.; Hong, J.K. Chemical pesticides and plant essential oils for disease control of tomato bacterial wilt. Plant Pathol. J. 2012, 28, 32–39. [Google Scholar] [CrossRef]
- Li, S.; Yu, Y.; Chen, J.; Guo, B.; Yang, L.; Ding, W. Evaluation of the Antibacterial Effects and Mechanism of Action of Protocatechualdehyde against Ralstonia solanacearum. Molecules 2016, 21, 754. [Google Scholar] [CrossRef] [PubMed]
- Tiew, P.; Ioset, J.; Kokpol, U.; Chavasiri, W.; Hostettmann, K. Antifungal, antioxidant and larvicidal activities of compounds isolated from the heartwood of Mansonia gagei. Phytother. Res. 2003, 17, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Yasunaka, K.; Abe, F.; Nagayama, A.; Okabe, H.; Lozada-Pérez, L.; López-Villafranco, E.; Muñiz, E.E.; Aguilar, A.; Reyes-Chilpa, R. Antibacterial activity of crude extracts from Mexican medicinal plants and purified coumarins and xanthones. J. Ethnopharmacol. 2005, 97, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wang, J.; Hou, Z.; Li, H.; Bi, Y. Inhibitory activity of resveratrol and its derivatives against phytopathogenic fungi. Chin. J. Pestic. Sci. 2012, 14, 283–290. [Google Scholar]
- Paulo, L.; Oleastro, M.; Gallardo, E.; Queiroz, J.; Domingues, F. Antimicrobial properties of resveratrol: A review. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Formatex Research Center: Badajoz, Spain, 2001; Volume 1, pp. 1225–1235. [Google Scholar]
- Houillé, B.; Papon, N.; Boudesocque, L.; Bourdeaud, E.; Besseau, S.; Courdavault, V.; Enguehard-Gueiffier, C.; Delanoue, G.; Guérin, L.; Bouchara, J.P. Antifungal activity of resveratrol derivatives against Candida species. J. Nat. Prod. 2014, 77, 1658–1662. [Google Scholar] [CrossRef] [PubMed]
- Nawrot-Modranka, J.; Nawrot, E.; Graczyk, J. In vivo antitumor, in vitro antibacterial activity and alkylating properties of phosphorohydrazine derivatives of coumarin and chromone. Eur. J. Med. Chem. 2006, 41, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, S.; Maggio, A.; Bellone, G.; Formisano, C.; Basile, A.; Cicala, C.; Alfieri, A.; Mascolo, N.; Bruno, M. Antibacterial and anticoagulant activities of coumarins isolated from the flowers of Magydaris tomentosa. Planta Med. 2007, 73, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Nguefack, J.; Budde, B.B.; Jakobsen, M. Five essential oils from aromatic plants of Cameroon: Their antibacterial activity and ability to permeabilize the cytoplasmic membrane of Listeria innocua examined by flow cytometry. Lett. Appl. Microbiol. 2004, 39, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [PubMed]
- Yang, L.; Ding, W.; Xu, Y.; Wu, D.; Li, S.; Chen, J.; Guo, B. New insights into the antibacterial activity of hydroxycoumarins against Ralstonia solanacearum. Molecules 2016, 21, 468. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Allen, C. The plant pathogen Ralstonia solanacearum needs aerotaxis for normal biofilm formation and interactions with its tomato host. J. Bacteriol. 2007, 189, 6415–6424. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.E.; Monier, J.M. The ecological significance of biofilm formation by plant-associated bacteria. Annu. Rev. Phytopathol. 2003, 41, 429–453. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, F.; Genin, S.; van Dijk, I.; Valls, M. A luminescent reporter evidences active expression of Ralstonia solanacearum type III secretion system genes throughout plant infection. Microbiology 2012, 158, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Sashidhara, K.V.; Kumar, A.; Kumar, M.; Sarkar, J.; Sinha, S. Synthesis and in vitro evaluation of novel coumarin-chalcone hybrids as potential anticancer agents. Bioorg. Med. Chem. Lett. 2010, 20, 7205–7211. [Google Scholar] [CrossRef] [PubMed]
- Riveiro, M.; De Kimpe, N.; Moglioni, A.; Vazquez, R.; Monczor, F.; Shayo, C.; Davio, C. Coumarins: Old compounds with novel promising therapeutic perspectives. Curr. Med. Chem. 2010, 17, 1325–1338. [Google Scholar] [CrossRef] [PubMed]
- Thati, B.; Noble, A.; Rowan, R.; Creaven, B.S.; Walsh, M.; McCann, M.; Egan, D.; Kavanagh, K. Mechanism of action of coumarin and silver(I)-coumarin complexes against the pathogenic yeast Candida albicans. Toxicol. In Vitro 2007, 21, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Paulo, L.; Ferreira, S.; Gallardo, E.; Queiroz, J.A.; Domingues, F. Antimicrobial activity and effects of resveratrol on human pathogenic bacteria. World J. Microbiol. Biotechnol. 2010, 26, 1533–1538. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Kets, E.; Smid, E. Mechanisms of action of carvacrol on the food-borne pathogen Bacillus cereus. Appl. Environ. Microbiol. 1999, 65, 4606–4610. [Google Scholar] [PubMed]
- La Storia, A.; Ercolini, D.; Marinello, F.; Di Pasqua, R.; Villani, F.; Mauriello, G. Atomic force microscopy analysis shows surface structure changes in carvacrol-treated bacterial cells. Res. Microbiol. 2011, 162, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.T.; Chu, S.D.; Hagerman, A.E.; Lorigan, G.A. Probing the interaction of polyphenols with lipid bilayers by solid-State NMR spectroscopy. J. Agric. Food Chem. 2011, 12, 6783–6789. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.D.; Hawes, J.W.; Lorigan, G.A. Solid-state NMR spectroscopic studies on the interaction of sorbic acid with phospholipid membranes at different pH levels. Magn. Reson. Chem. 2009, 8, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Vasse, J.; Frey, P.; Trigalet, A. Microscopic studies of intercellular infection and protoxylem invasion of tomato roots by Pseudomonas-solanacearum. Mol. Plant Microbe Interact. 1995, 8, 241–251. [Google Scholar] [CrossRef]
- Genin, S.; Denny, T.P. Pathogenomics of the Ralstonia solanacearum species complex. Annu. Rev. Phytopathol. 2012, 50, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, S.; Schell, M.A.; Denny, T.P. Pyramiding unmarked deletions in Ralstonia solanacearum shows that secreted proteins in addition to plant cell-wall-degrading enzymes contribute to virulence. Mol. Plant Microbe Interact. 2005, 18, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Tans-Kersten, J.; Brown, D.; Allen, C. Swimming motility, a virulence trait of Ralstonia solanacearum, is regulated by FlhDC and the plant host environment. Mol. Plant Microbe Interact. 2004, 17, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.M.; Macintyre, A.; Hawes, M.; Allen, C. Escaping underground nets: Extracellular DNases degrade plant extracellular traps and contribute to virulence of the plant pathogenic bacterium Ralstonia solanacearum. PLoS Pathog. 2016, 12, e1005686. [Google Scholar] [CrossRef] [PubMed]
- Tans-Kersten, J.; Huang, H.; Allen, C. Ralstonia solanacearum needs motility for invasive virulence on tomato. J. Bacteriol. 2001, 183, 3597–3605. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Kornberg, A. Inorganic polyphosphate is needed for swimming, swarming, and twitching motilities of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 4885–4890. [Google Scholar] [CrossRef] [PubMed]
- O’May, C.; Tufenkji, N. The swarming motility of Pseudomonas aeruginosa is blocked by cranberry proanthocyanidins and other tannin-containing materials. Appl. Environ. Microbiol. 2011, 77, 3061–3067. [Google Scholar] [CrossRef] [PubMed]
- Bogino, P.C.; Oliva, M.d.L.; Sorroche, F.G.; Giordano, W. The role of bacterial biofilms and surface components in plant-bacterial associations. Int. J. Mol. Sci. 2013, 14, 15838–15859. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, S.; Shams-Bakhsh, M.; Hosseinzadeh, E. Effects of sub-bactericidal concentration of plant essential oils on pathogenicity factors of Ralstonia solanacearum. Arch. Phytopathol. Plant Prot. 2013, 46, 643–655. [Google Scholar] [CrossRef]
- Chen, J.; Peng, H.; Wang, X.; Shao, F.; Yuan, Z.; Han, H. Graphene oxide exhibits broad-spectrum antimicrobial activity against bacterial phytopathogens and fungal conidia by intertwining and membrane perturbation. Nanoscale 2014, 6, 1879–1889. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yan, Z.; Xu, J. Quantitative variation of biofilms among strains in natural populations of Candida albicans. Microbiology 2003, 149, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Holden, C.; Trounson, A. Staining of the inner acrosomal membrane of human spermatozoa with concanavalin A lectin as an indicator of potential egg penetration ability. Fertil. Steril. 1991, 56, 967–974. [Google Scholar] [CrossRef]
- China State Tobacco Monopoly Administration. YC/T39−1996, Grade and Investigating Method of Tobacco Disease; China Standard Press: Beijing, China, 1996.
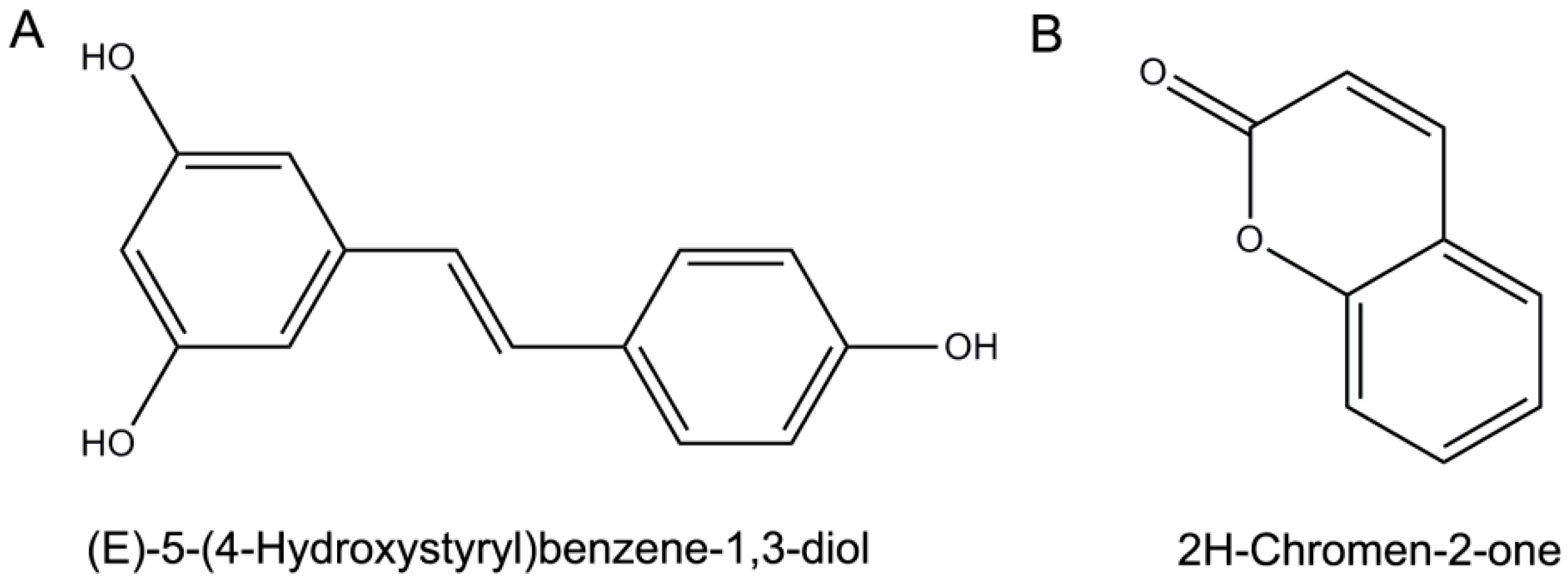
- Sample Availability: Samples of the compounds (Resveratrol and Coumarin) are available from Shanghai Yuanye Bio-technology Co., Ltd. (Shanghai, China).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Plant Species | Active Ingredients |
|---|---|---|
| Poaceae | Phyllostachys pubescens Mazel | Bamboo tar |
| Cymbopogon martinii | Palmarosa Oil | |
| Lamiaceae | Thymus serpyllum L. | Essential oil |
| Origanum vulgare “Rogeukuppel” | ||
| Saluia plebeia R. Br. | Ethanol extract | |
| Leucas aspera (Willd.) Link | ||
| Apocynaceae | Herba Catharanthi rosei | Ethanol extract |
| Hunteria zeylanica | ||
| Rutaceae | Atalantia buxifolia | Ethanol extract |
| Micromelum falcatum | ||
| Clausena lansium (Lour.) Skeels | Seed extracts | |
| Liliaceae | Allium sativum L. | Root exudates |
| Asteraceae | Amaranthus tricolor L. | Ethanol extract |
| Eupatorium adenophorum Spreng | Leaf exudates | |
| Elephantopus tomentosus L | Ethanol extract | |
| Magnoliaceae | Magnolia officinalis Rehd. et Wils. | Phenolic compounds |
| Others | Stemona sessilifolia (Miq.) Miq | Petrol ether extract |
| Casuarinaequisetifolia L | Ethanol extract | |
| Sophoraflavescens | Alkaloid | |
| Lithospermum erythrorhizon | Chloroform extract | |
| Toxicodendron sylvestre | Trimethylgallic acid | |
| Litsea cubeba | Ethanol extract |
| Compounds | Concentration (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| 128 | 64 | 32 | 16 | 8 | 4 | 2 | 1 | |
| resveratrol | + | + | + | + | + | + | + | - |
| coumarin | + | + | + | + | + | - | - | - |
| Treatments | On the Root Surface | Within the Root | ||||
|---|---|---|---|---|---|---|
| One Day after Inoculation (cfu/g) | Three Days after Inoculation (cfu/g) | Five Days after Inoculation (cfu/g) | One Day after Inoculation (cfu/g) | Three Days after Inoculation (cfu/g) | Five Days after Inoculation (cfu/g) | |
| Control | 1.03 × 106 b | 6.26 × 106 a | 1.79 × 108 a | 0.00 | 1.50 × 105 b | 5.2 × 106 a |
| Coumarin | 9.30 × 105 b | 4.31 × 106 a,b | 1.83 × 107 c | 0.00 | 4.12 × 104 c | 2.44 × 106 b |
| Resveratrol | 3.44 × 105 c | 3.58 × 106 b | 2.64 × 107 c | 0.00 | 1.41 × 105 b | 2.41 × 106 b |
| Days after Inoculation | Control | Coumarin Treatment | Resveratrol Treatment | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Disease Incidence (%) | Disease Index | Control Efficacy (%) | Disease Incidence (%) | Disease Index | Control Efficacy (%) | Disease Incidence (%) | Disease Index | Control Efficacy (%) | |
| 7 | 0 | 0 | - | 0 | 0 | 100 | 0 | 0 | 100 |
| 9 | 13 | 3 | - | 0.3 | 0 | 99 | 0 | 0 | 100 |
| 13 | 67 | 47 | - | 28 | 15 | 68 | 22 | 7 | 85 |
| 19 | 100 | 92 | - | 80 | 58 | 37 | 86 | 74 | 20 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Yu, Y.; Li, S.; Ding, W. Resveratrol and Coumarin: Novel Agricultural Antibacterial Agent against Ralstonia solanacearum In Vitro and In Vivo. Molecules 2016, 21, 1501. https://doi.org/10.3390/molecules21111501
Chen J, Yu Y, Li S, Ding W. Resveratrol and Coumarin: Novel Agricultural Antibacterial Agent against Ralstonia solanacearum In Vitro and In Vivo. Molecules. 2016; 21(11):1501. https://doi.org/10.3390/molecules21111501
Chicago/Turabian StyleChen, Juanni, Yanmei Yu, Shili Li, and Wei Ding. 2016. "Resveratrol and Coumarin: Novel Agricultural Antibacterial Agent against Ralstonia solanacearum In Vitro and In Vivo" Molecules 21, no. 11: 1501. https://doi.org/10.3390/molecules21111501
APA StyleChen, J., Yu, Y., Li, S., & Ding, W. (2016). Resveratrol and Coumarin: Novel Agricultural Antibacterial Agent against Ralstonia solanacearum In Vitro and In Vivo. Molecules, 21(11), 1501. https://doi.org/10.3390/molecules21111501