
Effects of the Powder from Hoggery Desulfurization Tanks on the Salinity Resistance of Lettuce


Abstract
:1. Introduction
2. Materials and Methods
2.1. Design of Experiment
2.2. Mineral Content
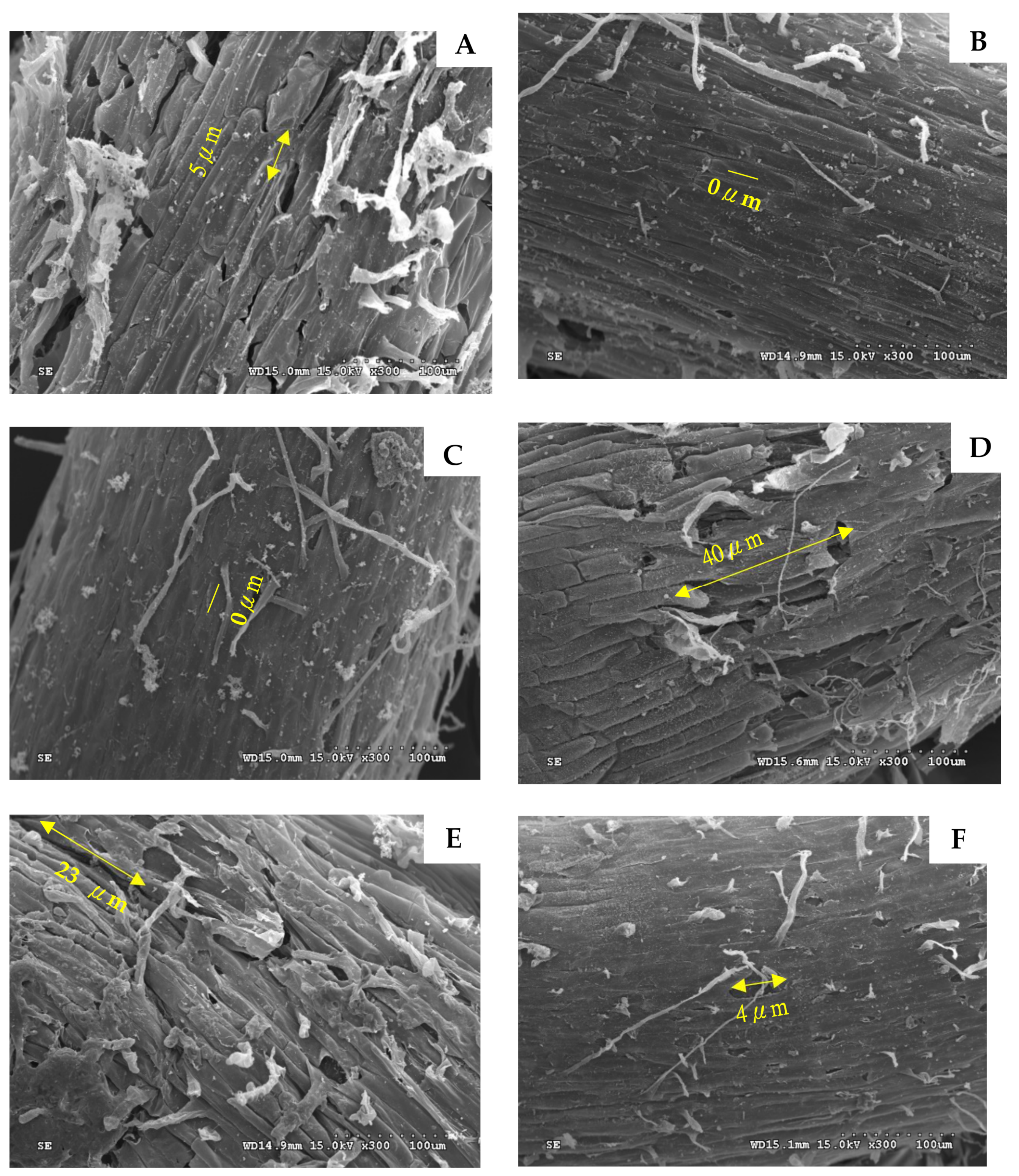
2.3. Cell Morphology of Root Apices in Lettuces
2.4. Statistical Analysis
3. Results
3.1. Growth Analysis
3.2. Mineral Content in Lettuce
3.3. Between-Group Differences in Cell Morphology of Lettuce Root Apices
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Alavi, S.A.; Ghehsareh, A.M.; Soleymani, A.; Panahpour, E. Enhanced nutrient uptake in salt-stressed Mentha piperita using magnetically treated water. Protoplasma 2020, 258, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Andriolo, J.L.; da Luz, G.L.; Witter, M.H.; Godoi, R.D.S.; Barros, G.T. Growth and yield of lettuce plants under salinity. Hortic. Bras. 2004, 23, 931–934. [Google Scholar] [CrossRef] [Green Version]
- Nasri, N.; Saïdi, I.; Kaddour, R.; Lachaâl, M. Effect of Salinity on Germination, Seedling Growth and Acid Phosphatase Activity in Lettuce. Am. J. Plant Sci. 2015, 6, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Maathuis, F.J.M. Sodium in plants: Perception, signalling, and regulation of sodium fluxes. J. Exp. Bot. 2014, 65, 849–858. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef]
- Burow, M.; Wittstock, U.; Gershenzon, J. Role of Sulfur for Plant Production in Agricultural and Natural Ecosystems. Adv. Photosyn. Respir. 2008, 27, 201–216. [Google Scholar] [CrossRef]
- Belikova, H.; Meszaros, M.; Varga, L.; Arvay, J.; Wiśniowska-Kielian, B.; Gondek, K.; Antonkiewicz, J.; Torma, S.; Lazarevic, B.; von Bennewitz, E.; et al. The effect of different forms of sulphur on incidence of apple scab on apple tree (Malus × domestica BORKH) gloster CV. Ecol. Chem. Eng. S. 2019, 26, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Tavakkoli, E.; Fatehi, F.; Coventry, S.; Rengasamy, P.; McDonald, G.K. Additive effects of Na+ and Cl– ions on barley growth under salinity stress. J. Exp. Bot. 2011, 62, 2189–2203. [Google Scholar]
- Nazar, R.; Khan, M.I.R.; Iqbal, N.; Masood, A.; Khan, N.A. Involvement of ethylene in reversal of salt-inhibited photosynthesis by sulfur in mustard. Physiol. Plant. 2014, 152, 331–344. [Google Scholar] [CrossRef]
- Phour, M.; Sindhu, S.S. Amelioration of salinity stress and growth stimulation of mustard (Brassica juncea L.) by salt-tolerant Pseudomonas species. Appl. Soil Ecol. 2020, 149, 103518. [Google Scholar] [CrossRef]
- Freitas, W.E.D.S.; de Oliveira, A.B.; Mesquita, R.O.; de Carvalhob, H.H.; Prisco, J.T.; Gomes-Filho, E. Sulfur-induced salinity tolerance in lettuce is due to a better P and K uptake, lower Na/K ratio and an efficient antioxidative defense system. Sci. Hortic. 2019, 257, 1–11. [Google Scholar] [CrossRef]
- Syed, M.; Soreanu, G.; Falletta, P.; Béland, M. Removal of hydrogen sulfide from gas streams using biological processes-A review. Can. Biosyst. Eng. 2006, 48, 1–14. [Google Scholar]
- Schieder, D.; Quicker, P.; Schneider, R.; Winter, H.; Prechtl, S.; Faulstich, M.; Schieder, D. Microbiological removal of hydrogen sulfide from biogas by means of a separate biofilter system: Experience with technical operation. Water Sci. Technol. 2003, 48, 209–212. [Google Scholar] [CrossRef]
- Andriamanohiarisoamanana, F.J.; Sakamoto, Y.; Yamashiro, T.; Yasui, S.; Iwasaki, M.; Ihara, I.; Tsuji, O.; Umetsu, K. Effects of handling parameters on hydrogen sulfide emission from stored dairy manure. J. Environ. Manag. 2015, 154, 110–116. [Google Scholar] [CrossRef]
- Xiao, C.; Ma, Y.Q.; Ji, D.D.; Zang, L.H. Review of desulfurization process for biogas purification. Earth Environ. Sci. 2017, 100, 1–7. [Google Scholar] [CrossRef]
- Chen, J.H.; Lin, Y.H. Sodium chloride causes variation in organic acids and proteins in tomato root. Afr. J. Biotechnol. 2010, 9, 8161–8167. [Google Scholar] [CrossRef] [Green Version]
- Ugo, N.J.; Ade, A.R.; Joy, A.T. Nutrient Composition of Carica Papaya Leaves Extracts. J. Food Sci. Nutr. Res. 2019, 2, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur assimilation in photosynthetic organisms: Molecular functions and regulations of transporters and assimilatory enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar] [CrossRef]
- Sudhir, P.; Murthy, S.D.S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Fotopoulos, V.; Christou, A.; Antoniou, C.; Manganaris, G.A. Hydrogen sulphide: A versatile tool for the regulation of growth and defence responses in horticultural crops. J. Hortic. Sci. Biotechnol. 2015, 90, 227–234. [Google Scholar] [CrossRef]
- Gong, D.H.; Wang, G.Z.; Si, W.T.; Zhou, Y.; Liu, Z.; Jia, J. Effects of Salt Stress on Photosynthetic Pigments and Activity of Ribulose-1,5-bisphosphate Carboxylase/Oxygenase in Kalidium foliatum. Russ. J. Plant Physiol. 2018, 65, 98–103. [Google Scholar] [CrossRef]
- Wang, J.J.; Dustin, L. Effect of Ammonium, Potassium, and Sodium Cations and Phosphate, Nitrate, and Chloride Anions on Zinc Sorption and Lability in Selected Acid and Calcareous Soils. Soil Sci. Soc. Am. J. 2005, 69, 1036–1046. [Google Scholar] [CrossRef]
- Tian, T.; Chu, X.Y.; Yang, Y.; Zhang, X.; Liu, Y.M.; Gao, J.; Ma, B.G.; Zhang, H.Y. Phosphates as Energy Sources to Expand Metabolic Networks. Life 2019, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Cerozi, B.S.; Fitzsimmons, K. The effect of pH on phosphorus availability and speciation in an aquaponics nutrient solution. Bioresour. Technol. 2016, 219, 778–781. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.A.; Feng, L.Y.; Iqbal, N.; Manaf, A.; Khalid, M.H.B.; Rehman, S.U.; Wasaya, A.; Ansar, M.; Billah, M.; Yang, F.; et al. Effect of sulphur application on photosynthesis and biomass accumulation of sesame varieties under rainfed conditions. Agronomy 2018, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.A.; Khan, M.I.R.; Asgher, M.; Fatma, M.; Masood, A.; Syeed, S. Salinity tolerance in plants: Revisiting the role of sulfur metabolites. Plant Physiol. Biochem. 2014, 2, 120. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, S.; Zuchi, S. Adequate S supply protects barley plants from adverse effects of salinity stress by increasing thiol contents. Acta Physiol. Plant 2013, 35, 175–181. [Google Scholar] [CrossRef]
- Neinhuis, C.; Edelmann, H.G. Methanol as a rapid fixative for the investigation of plant surfaces by SEM. J. Microsc. 1996, 184, 14–16. [Google Scholar] [CrossRef]
- Zhao, Q.; Wu, Y.; Gao, L.; Ma, J.; Li, C.Y.; Xiang, C.B. Sulfur nutrient availability regulates root elongation by affecting root IAA levels and the stem cell niche. J. Integr. Plant Biol. 2014, 56, 1151–1163. [Google Scholar] [CrossRef]
- Hell, R.; Khan, M.S.; Wirtz, M. Cellular biology of sulfur and its functions in plants. In Cell Biology of Metals and Nutrients; Springer: Berlin/Heidelberg, Germany, 2010; pp. 243–279. [Google Scholar] [CrossRef]
- Chan, K.S.; Phua, S.Y.; Breusegem, F.V. Secondary sulfur metabolism in cellular signalling and oxidative stress responses. J. Exp. Bot. 2019, 70, 4237–4250. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Treatment | Plant Height (cm) | Root Length (cm) | FW 1 (g) | N (%) | P (%) | K (%) |
|---|---|---|---|---|---|---|
| S0H0 | 7.9 ± 0.7 a 2 | 18.1 ± 0.8 ab | 51.4 ± 3.1 ab | 3.92 ± 0.4 a | 0.47 ± 0.02 a | 2.99 ± 0.5 a |
| S0H1.5 | 8.3 ± 0.8 a | 18.7 ± 0.7 a | 58.2 ± 3.5 a | 3.85 ± 0.3 a | 0.49 ± 0.03 a | 3.15 ± 0.4 a |
| S0H3.0 | 8.1 ± 0.6 a | 18.7 ± 0.9 a | 62.8 ± 4.1 a | 3.77 ± 0.3 a | 0.52 ± 0.03 a | 3.39 ± 0.4 a |
| S40H0 | 7.1 ± 0.5 b | 17.2 ± 0.5 b | 43.6 ± 3.0 b | 2.83 ± 0.4 b | 0.32 ± 0.04 b | 2.59 ± 0.3 b |
| S40H1.5 | 7.3 ± 0.5 b | 17.6 ± 0.6 ab | 47.8 ± 4.2 b | 2.91 ± 0.3 b | 0.34 ± 0.03 b | 2.99 ± 0.4 a |
| S40H3.0 | 7.8 ± 0.3 a | 18.0 ± 0.4 ab | 49.5 ± 3.7 ab | 3.12 ± 0.2 ab | 0.38 ± 0.02 b | 3.16 ± 0.5 a |
| Treatments | Root | Leaf | ||||
|---|---|---|---|---|---|---|
| K (mg/g DW 1) | Na (mg/g DW) | K/Na | K (mg/g DW) | Na (mg/g DW) | K/Na | |
| S0H0 | 27.85 ± 0.58 a 2 | 5.88 ± 0.14 b | 4.73 ± 0.15 a | 29.91 ± 0.34 ab | 4.71 ± 0.11 b | 6.35 ± 0.24 b |
| S0H1.5 | 27.93 ± 0.35 a | 5.93 ± 0.16 b | 4.71 ± 0.13 a | 31.46 ± 0.22 ab | 4.80 ± 0.13 c | 6.55 ± 0.21 ab |
| S0H3.0 | 28.17 ± 0.56 a | 6.06 ± 0.18 b | 4.65 ± 0.17 a | 33.88 ± 0.18 a | 4.76 ± 0.04 c | 7.12 ± 0.09 a |
| S40H0 | 19.76 ± 0.43 b | 12.05 ± 0.42 a | 1.64 ± 0.08 c | 25.94 ± 0.53 b | 16.11 ± 0.29 a | 1.61 ± 0.03 d |
| S40H1.5 | 23.08 ± 0.94 ab | 11.74 ± 0.15 a | 1.97 ± 0.09 c | 29.92 ± 0.07 ab | 15.23 ± 0.35 a | 1.96 ± 0.05 c |
| S40H3.0 | 24.41 ± 0.29 ab | 10.12 ± 0.08 a | 2.41 ± 0.11 b | 31.60 ± 0.11 ab | 14.85 ± 0.30 ab | 2.12 ± 0.06 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-T.; Lin, Y.-H.; Wang, W.-J. Effects of the Powder from Hoggery Desulfurization Tanks on the Salinity Resistance of Lettuce. Plants 2022, 11, 868. https://doi.org/10.3390/plants11070868
Chang Y-T, Lin Y-H, Wang W-J. Effects of the Powder from Hoggery Desulfurization Tanks on the Salinity Resistance of Lettuce. Plants. 2022; 11(7):868. https://doi.org/10.3390/plants11070868
Chicago/Turabian StyleChang, Yao-Tsung, Yong-Hong Lin, and Wei-Jia Wang. 2022. "Effects of the Powder from Hoggery Desulfurization Tanks on the Salinity Resistance of Lettuce" Plants 11, no. 7: 868. https://doi.org/10.3390/plants11070868
APA StyleChang, Y.-T., Lin, Y.-H., & Wang, W.-J. (2022). Effects of the Powder from Hoggery Desulfurization Tanks on the Salinity Resistance of Lettuce. Plants, 11(7), 868. https://doi.org/10.3390/plants11070868