
MicroRNA-Based and Proteomics Fingerprinting of Avena sativa L. Genotypes †


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Selection of Markers and Primers Design
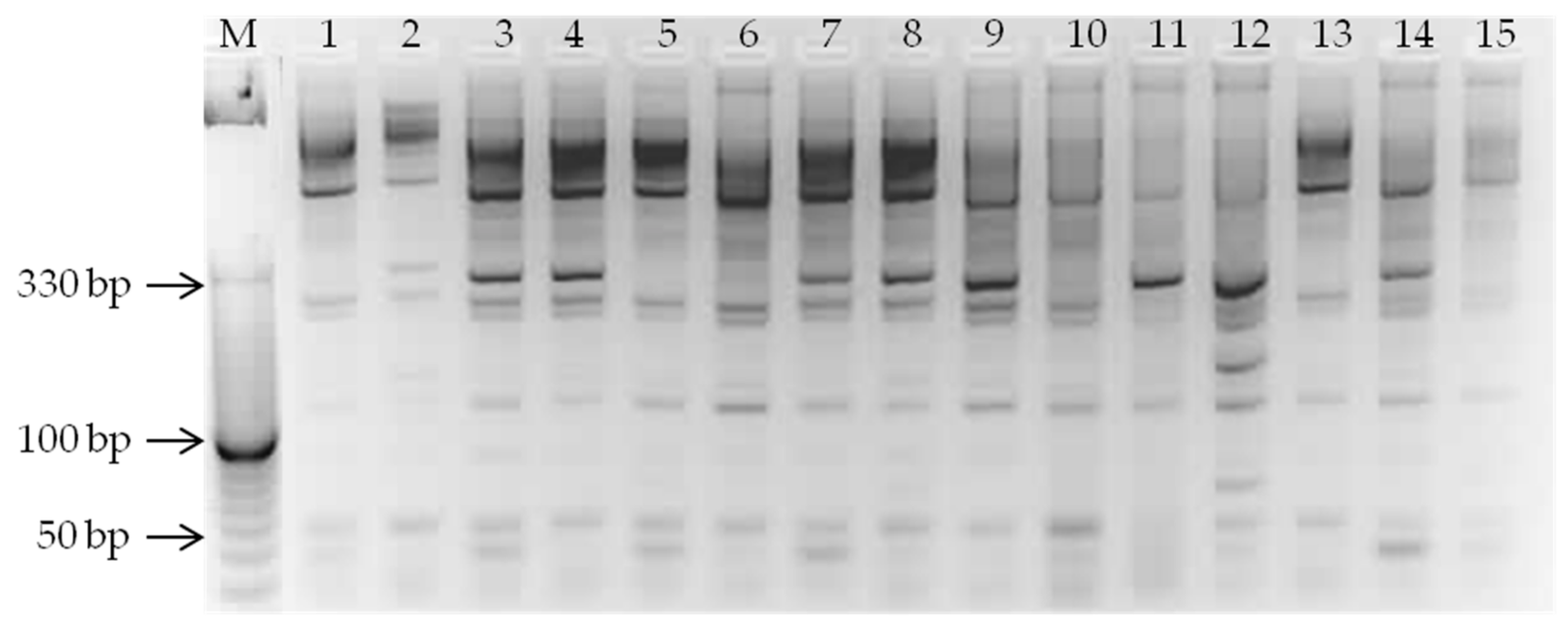
2.3. MicroRNA-Based Marker Assay
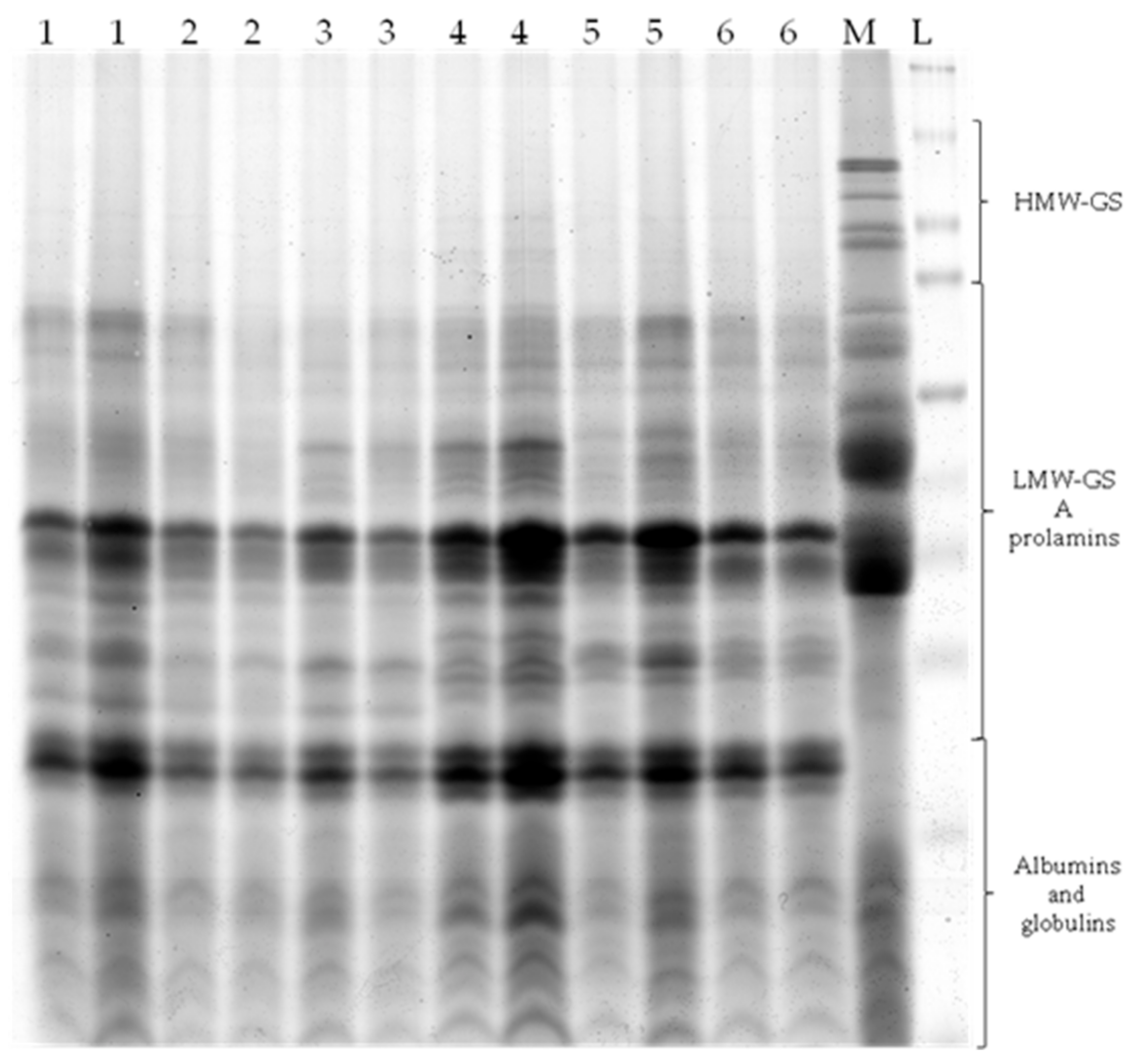
2.4. SDS-PAGE
3. Results and Discussion
3.1. MiRNA-Based Assay
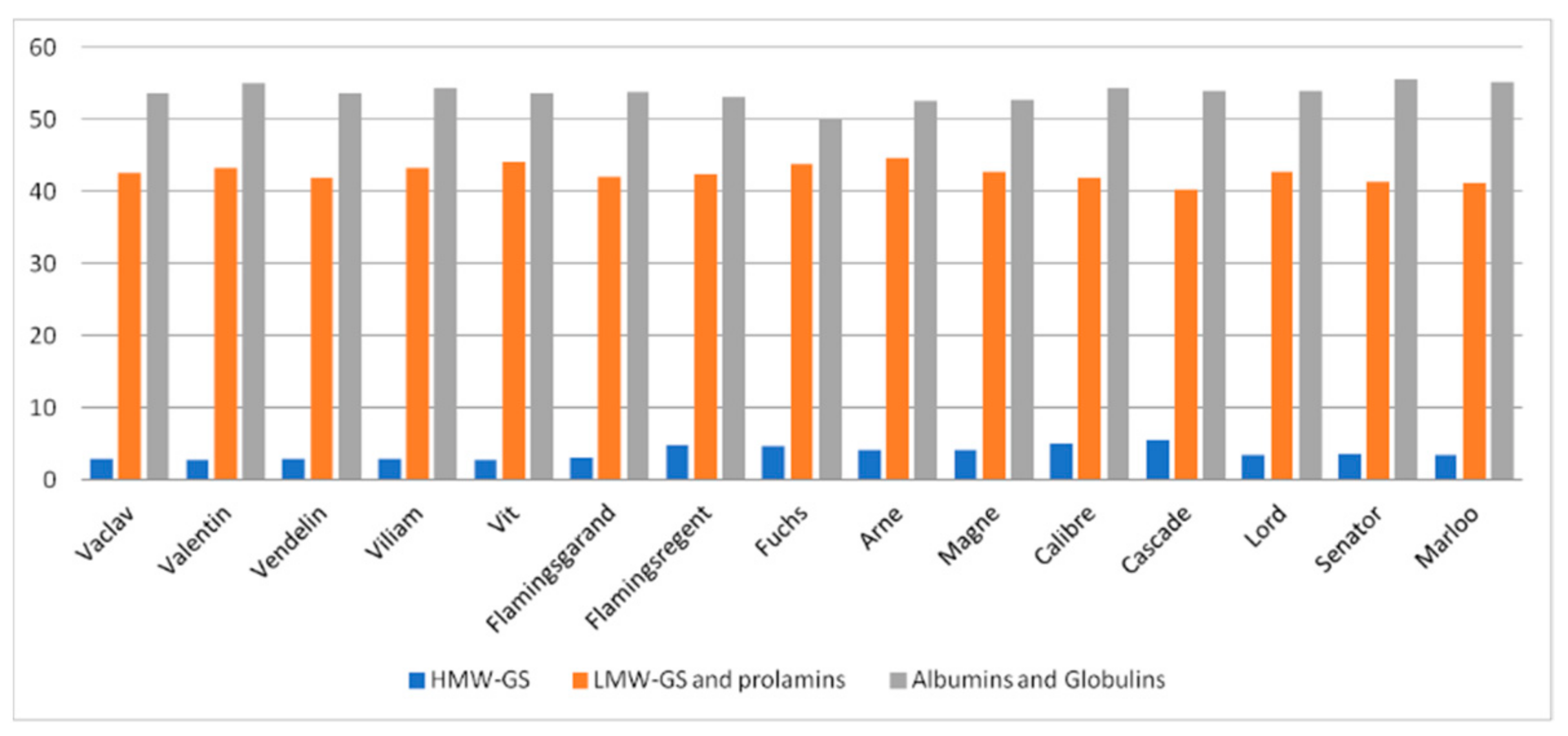
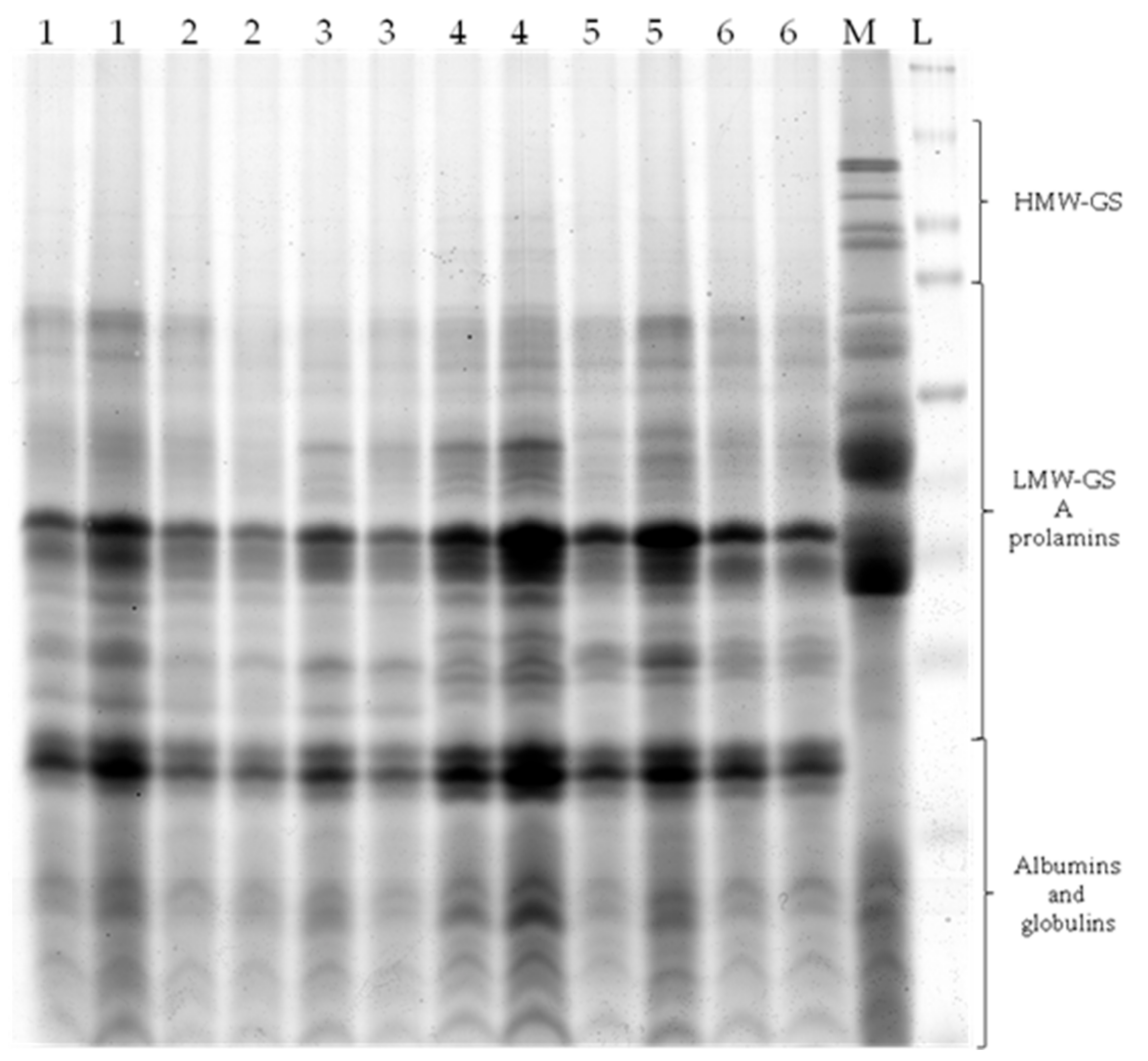
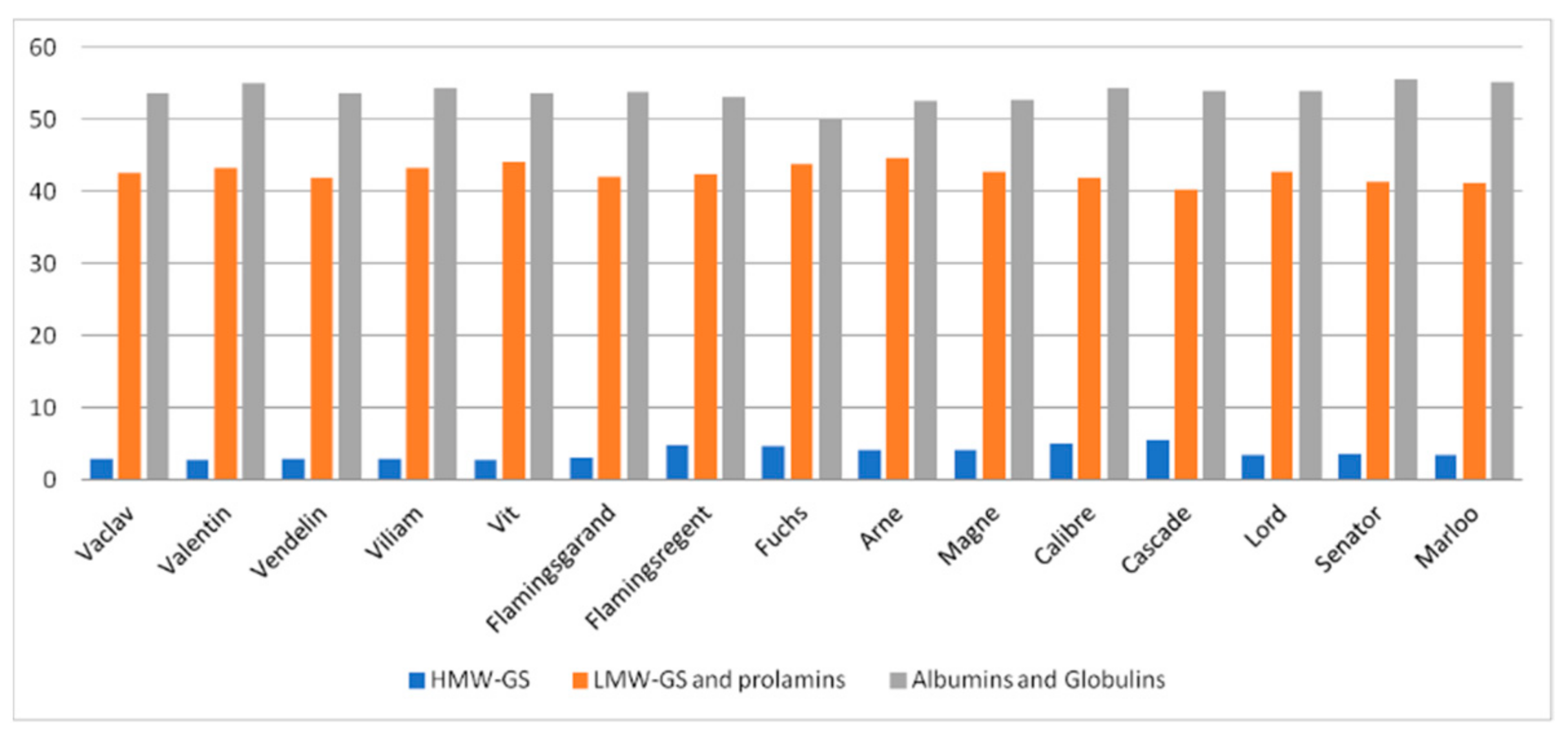
3.2. SDS-PAGE Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhu, C.; Ding, Y.; Liu, H. MiR398 and plant stress responses. Physiol. Plant. 2011, 143, 1–9. [Google Scholar] [CrossRef]
- Bej, S.; Basak, J. MicroRNAs: The potential biomarkers in plant stress response. Am. J. Plant Sci. 2014, 5, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Barciszewska-Pacak, M.; Milanowska, K.; Knop, K.; Bielewicz, D.; Nuc, P.; Plewka, P.; Pacak, A.M.; Vazquez, F.; Karlowski, W.; Jarmolowski, A.; et al. Arabidopsis microRNA expression regulation in a wide range of abiotic stress responses. Front. Plant Sci. 2015, 6, 410. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Ma, B.I.; Mason, A.S.; Xiao, M.; Wei, L.; An, Z. MicroRNA-based molecular markers: A novel PCR-based genotyping technique in Brassica species. Plant Breed. 2013, 132, 375–381. [Google Scholar] [CrossRef]
- Yadav, C.B.Y.; Muthamilarasan, M.; Pandey, G.; Khan, Y.; Prasad, M. Development of novel microRNA-based genetic markers in foxtail millet for genotyping applications in related grass species. Mol. Breed. 2014, 34, 2219–2224. [Google Scholar] [CrossRef]
- Mondal, T.K.; Ganie, S.A. Identification and characterization of salt responsive miRNA-SSR. Gene 2014, 535, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Ganie, S.A.; Mondal, T.K. Genome-wide development of novel miRNA-based microsatellite markers of rice (Oryza sativa) for genotyping applications. Mol. Breed. 2015, 35, 1–12. [Google Scholar] [CrossRef]
- Pegler, J.L.; Grof, C.P.L.; Eamens, A.L. Profiling of the Differential Abundance of Drought and Salt Stress-Responsive MicroRNAs Across Grass Crop and Genetic Model Plant Species. Agronomy 2018, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Li, J.; Liu, F.; Gu, J.; Guo, C.; Xu, L.; Zhang, H.; Xiao, K. Expression pattern of wheat miRNAs under salinity stress and prediction of salt-inducible miRNAs targets. Front. Agric. China 2011, 5, 413–422. [Google Scholar] [CrossRef]
- Kantar, M.; Unver, T.; Budak, H. Regulation of barley miRNAs upon dehydration stress correlated with target gene expression. Funct. Integr. Genom. 2010, 10, 493–507. [Google Scholar] [CrossRef]
- Gálová, Z.; Palenčárová, E.; Chňapek, M.; Balážová, Ž. Využitie Obilnín, Pseudoobilnín a Strukovín v Bezlepkovej Diéte, 1st ed.; [Use of Cereals, Pseudo-Cereals and Legumes in a Gluten-Free Diet]; Slovenská Poľnohospodárska Unierzita: Nitra, Slovakia, 2012; 182p, ISBN 978-80-552-0826-8. [Google Scholar]
- Sunkar, R.; Zhou, X.; Zheng, Y.; Zhang, W.; Zhu, J.K. Identification of novel and candidate miRNAs in rice by high throughput sequencing. BMC Plant Biol. 2008, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Barrera-Figueroa, B.E.; Gao, L.; Wu, Z.; Zhou, X.; Zhu, J.; Jin, H.; Liu, R.; Zhu, J. High throughput sequencing reveals novel and abiotic stress-regulated microRNAs in the inflorescences of rice. BMC Plant Biol. 2012, 12, 132. [Google Scholar] [CrossRef] [Green Version]
- Kehr, L. Systemic regulation of mineral homeostasis by microRNA. Front. Plant Sci. 2013, 4, 145. [Google Scholar] [CrossRef] [Green Version]
- Akdogan, G.; Tufekci, E.D.; Uranbey, S.; Unver, T. MiRNA-based drought regulation in wheat. Funct. Integr. Genom. 2016, 16, 221–233. [Google Scholar] [CrossRef]
- Griffiths-Jones, S. The microRNA Registry. Nucleic Acids Res. 2004, 32, D109–D111. [Google Scholar] [CrossRef]
- Ražná, K.; Hlavačková, L. The applicability of genetic markers based on molecules microRNA in agriculturel research. Open Access J. Agri. Res. 2017, 2, 000125. [Google Scholar] [CrossRef]
- Wrigley, C. Identification of cereal varieties by gel electrophoresis of the grain proteins. In Seed Analysis; Springer: Berlin/Heidelberg, Germany, 1992; pp. 17–41. [Google Scholar]
- Sunkar, R.; Kapoor, A.; Zhu, J.K. Posttranscriptional Induction of Two Cu/Zn Superoxide Dismutase Genes in Arabidopsis Is Mediated by Downregulation of miR398 and Important for Oxidative Stress Tolerance. Plant Cell 2006, 18, 2051–2065. [Google Scholar] [CrossRef] [Green Version]
- Trindade, I.; Capitão, C.; Dalmay, T.; Fevereiro, M.P.; Dos Santos, D.M. miR398 and miR408 are up-regulated in response to water deficit in Medicago truncatula. Planta 2010, 231, 705–716. [Google Scholar] [CrossRef]
- Jia, X.; Wang, W.X.; Ren, L.; Chen, Q.J.; Mendu, V.; Willcut, B.; Dinkins, R.; Tang, X.; Tang, G. Differential and dynamic regulation of miR398 in response to ABA and salt stress in Populus tremula and Arabidopsis thaliana. Plant Mol. Biol. 2009, 71, 51–59. [Google Scholar] [CrossRef]
- Pan, J.; Huang, D.; Guo, Z.; Kuang, Z.; Zhang, H.; Xie, X.; Ma, Z.; Gao, S.; Lerdau, M.T.; Chu, C.; et al. Overexpression of microRNA408 enhances photosynthesis, growth, and seed yield in diverse plants. J. Integr. Plant Biol. 2018, 60, 323–340. [Google Scholar] [CrossRef]
- Barvkar, V.T.; Pardeshi, V.C.; Kale, S.M.; Qiu, S.; Rollins, M.; Datla, R.; Gupta, V.S.; Kadoo, N.Y. Genome-wide identification and characterization of microRNA genes and their targets in flax (Linum usitatissimum): Characterization of flax miRNA genes. Planta 2013, 237, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Shen, J.; Hou, X.; Yao, J.; Li, X.; Xiao, J.; Xiong, L. Gradual increase of miR156 Regulates Temporal Expression Changes of Numerous Genes during Leaf Development in Rice. ASPB 2012, 158, 1382–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, L.C.; Lin, S.; Shih, A.C.; Chen, J.; Lin, W.; Tseng, C.; Li, W.; Chiou, T. Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing. Plant Physiol. 2009, 151, 2120–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, A.W.; Shi, B.J.; Huang, C.Y.; Langridge, P.; Baumann, U. Discovery of barley miRNAs through deep sequencing of short reads. BMC Genom. 2011, 12, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, G.; Ai, Q.; Yu, D. Uncovering miRNAs involved in crosstalk between nutrient deficiencies in Arabidopsis. Sci. Rep. 2015, 5, 11813. [Google Scholar] [CrossRef] [Green Version]
- Rajwanshi, R.; Chakraborty, S.; Jayanandi, K.; Deb, B.; Lightfoot, D.A. Orthologous plant microRNAs: Microregulators with great potential for improving stress tolerance in plants. Theor. Appl. Genet. 2014, 127, 2525–2543. [Google Scholar] [CrossRef]
- Sharma, D.; Tiwari, M.; Pandey, A.; Bhatia, C.; Sharma, A.; Trivedi, P.K. MicroRNA858 Is a Potential Regulator of Phenylpropanoid Pathway and Plant Development. Plant Physiol. 2016, 171, 944–959. [Google Scholar] [CrossRef] [Green Version]
- Gregova, E.; Šliková, S.; Hozlár, P. Seed protein electrophoresis for identification of oat registered cultivars. Potravinarstvo 2015, 9, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Anderson, O.D. The Spectrum of Major Seed Storage Genes and Proteins in Oats (Avena sativa). PLoS ONE 2014, 9, e83569. [Google Scholar] [CrossRef] [Green Version]
- Mickowska, B.; Litwinek, D.; Gambuś, H. Oat raw materials and bakery products–amino acid composition and celiac immunoreactivity. Acta Sci. Pol. Technol. Aliment 2016, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Rajnincová, D.; Gálová, Z.; Chňapek, M. Comparison of selected wheat, oat and buckwheat genotypes on proteomic level. J. Cent. Eur. Agric. 2019, 20, 891–899. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ražná, K.; Gálová, Z.; Balážová, Ž.; Chňapek, M.; Harenčár, Ľ. MicroRNA-Based and Proteomics Fingerprinting of Avena sativa L. Genotypes. Biol. Life Sci. Forum 2022, 11, 5. https://doi.org/10.3390/IECPS2021-11917
Ražná K, Gálová Z, Balážová Ž, Chňapek M, Harenčár Ľ. MicroRNA-Based and Proteomics Fingerprinting of Avena sativa L. Genotypes. Biology and Life Sciences Forum. 2022; 11(1):5. https://doi.org/10.3390/IECPS2021-11917
Chicago/Turabian StyleRažná, Katarína, Zdenka Gálová, Želmíra Balážová, Milan Chňapek, and Ľubomír Harenčár. 2022. "MicroRNA-Based and Proteomics Fingerprinting of Avena sativa L. Genotypes" Biology and Life Sciences Forum 11, no. 1: 5. https://doi.org/10.3390/IECPS2021-11917
APA StyleRažná, K., Gálová, Z., Balážová, Ž., Chňapek, M., & Harenčár, Ľ. (2022). MicroRNA-Based and Proteomics Fingerprinting of Avena sativa L. Genotypes. Biology and Life Sciences Forum, 11(1), 5. https://doi.org/10.3390/IECPS2021-11917