
The Physiology of Postharvest Tea (Camellia sinensis) Leaves, According to Metabolic Phenotypes and Gene Expression Analysis


Abstract
:1. Introduction
2. Results
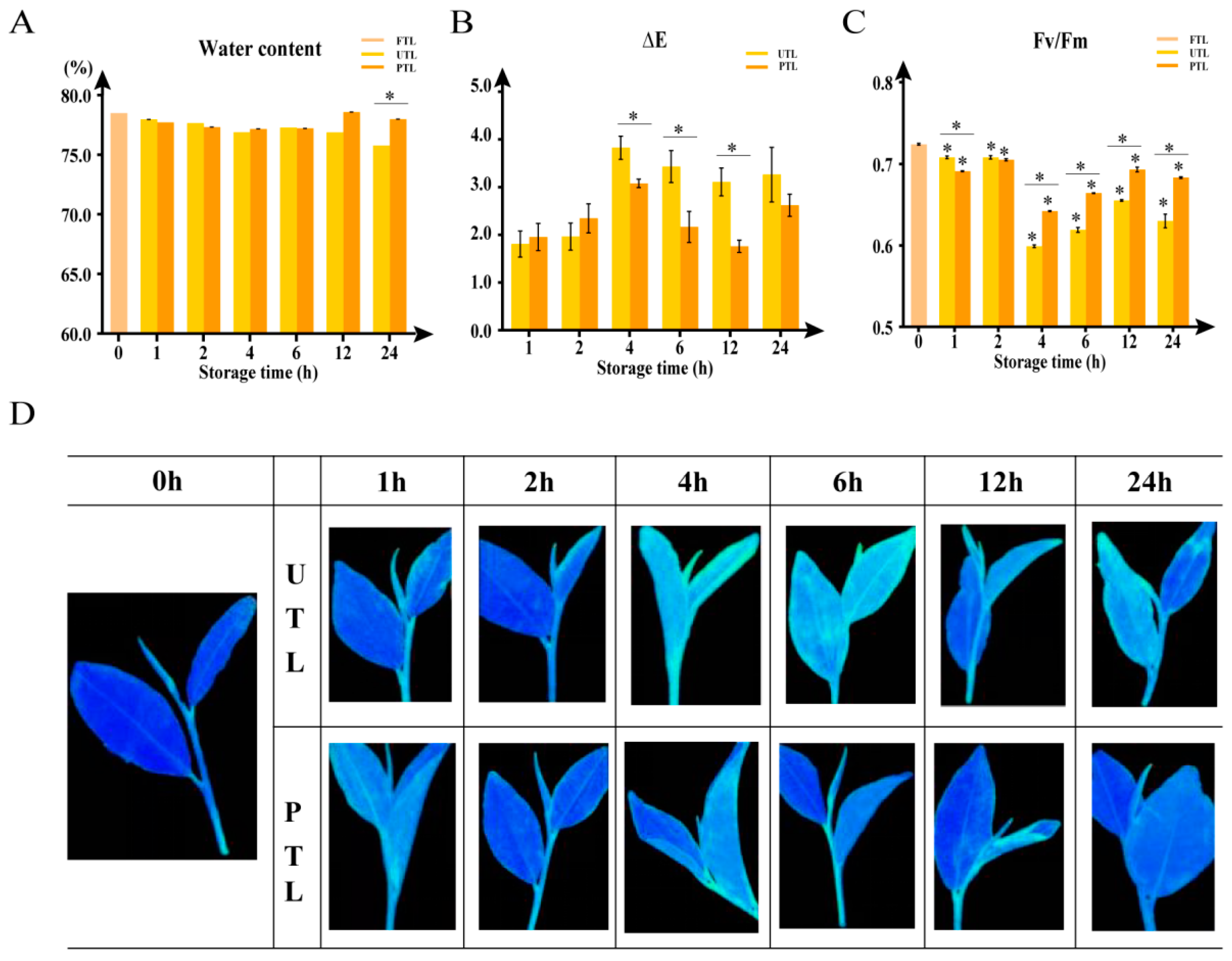
2.1. Changes in Water Content, Color, and Degree of Damage during Storage
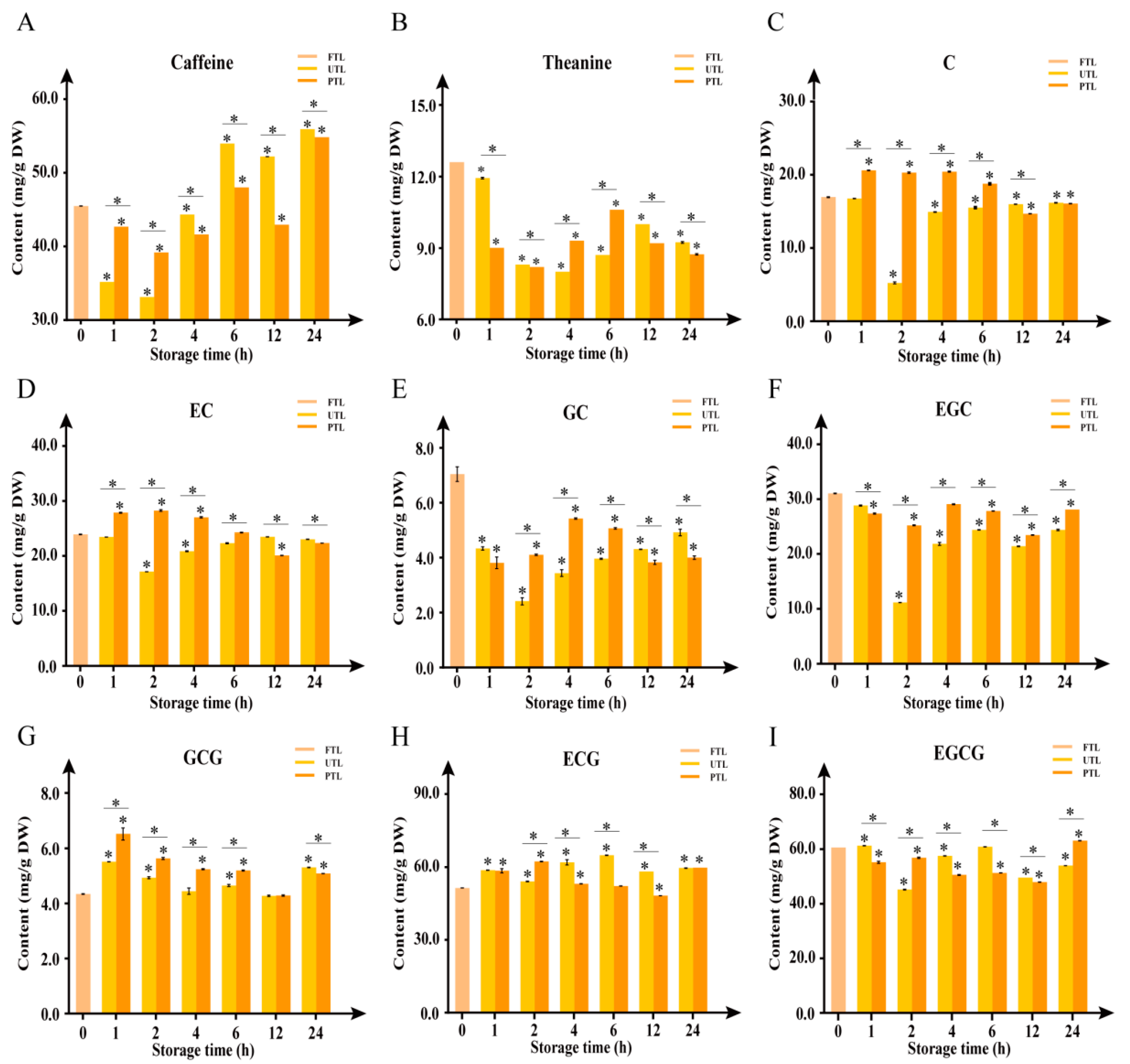
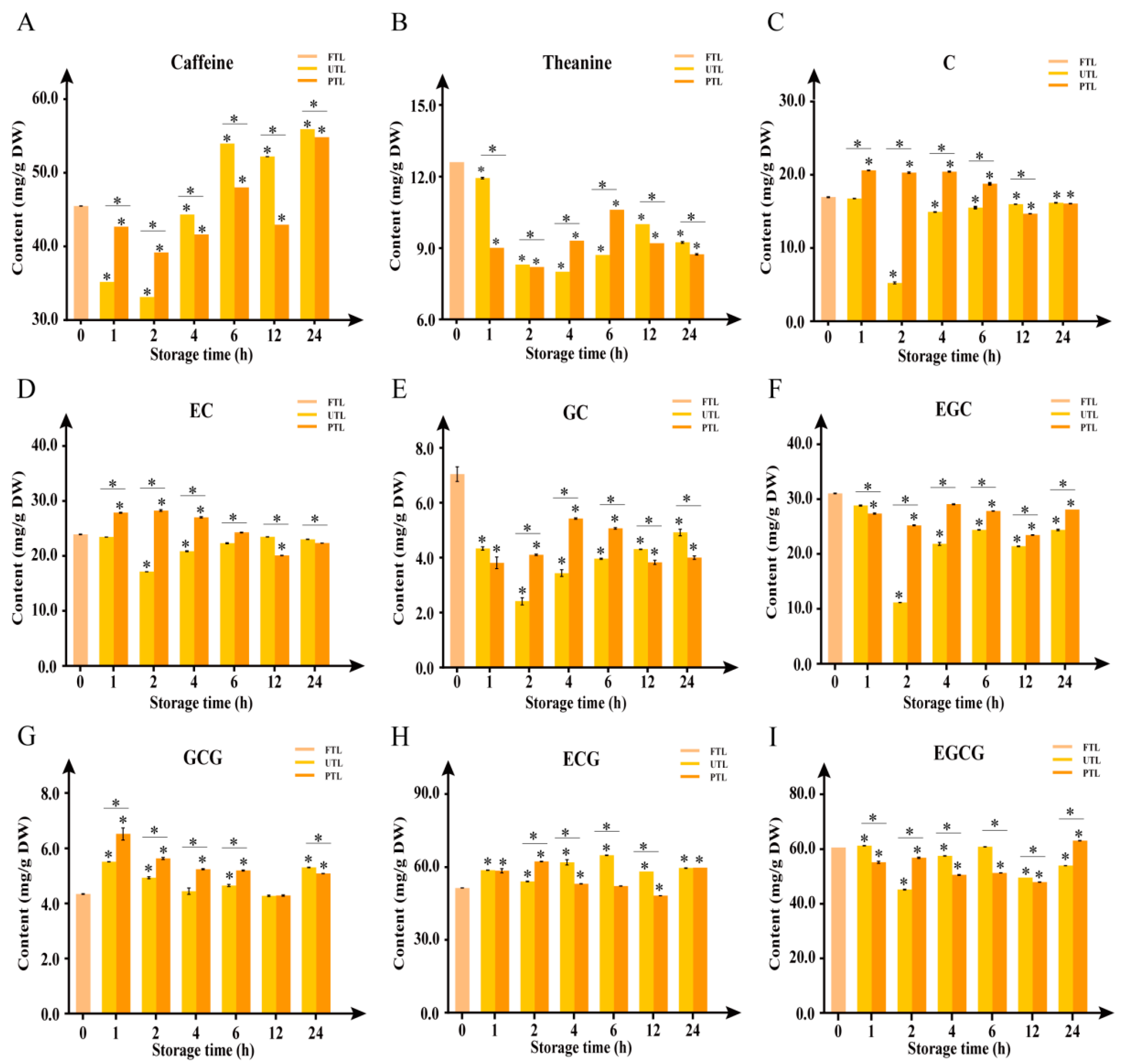
2.2. Changes in Caffeine, Theanine, and Catechins Contents during Storage
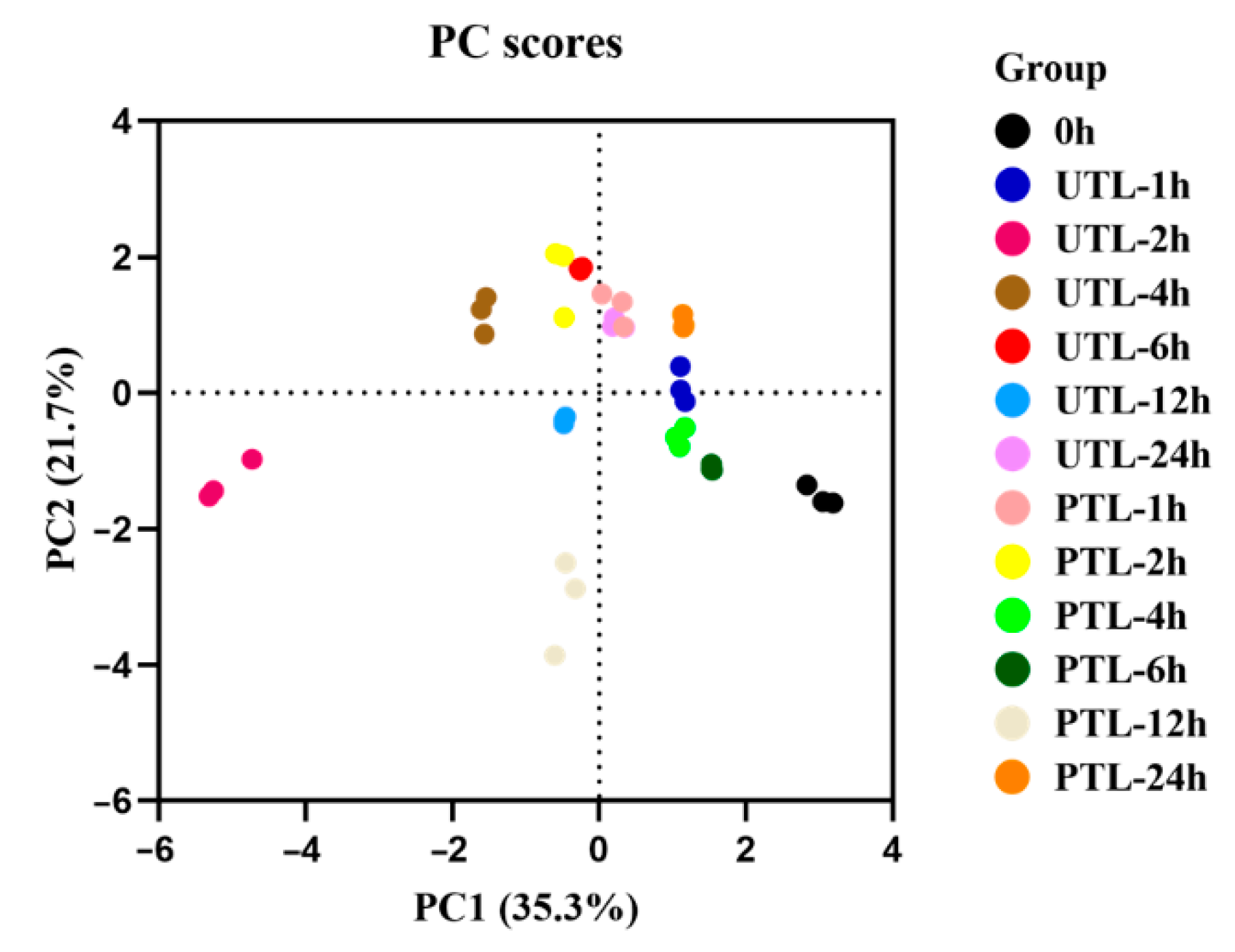
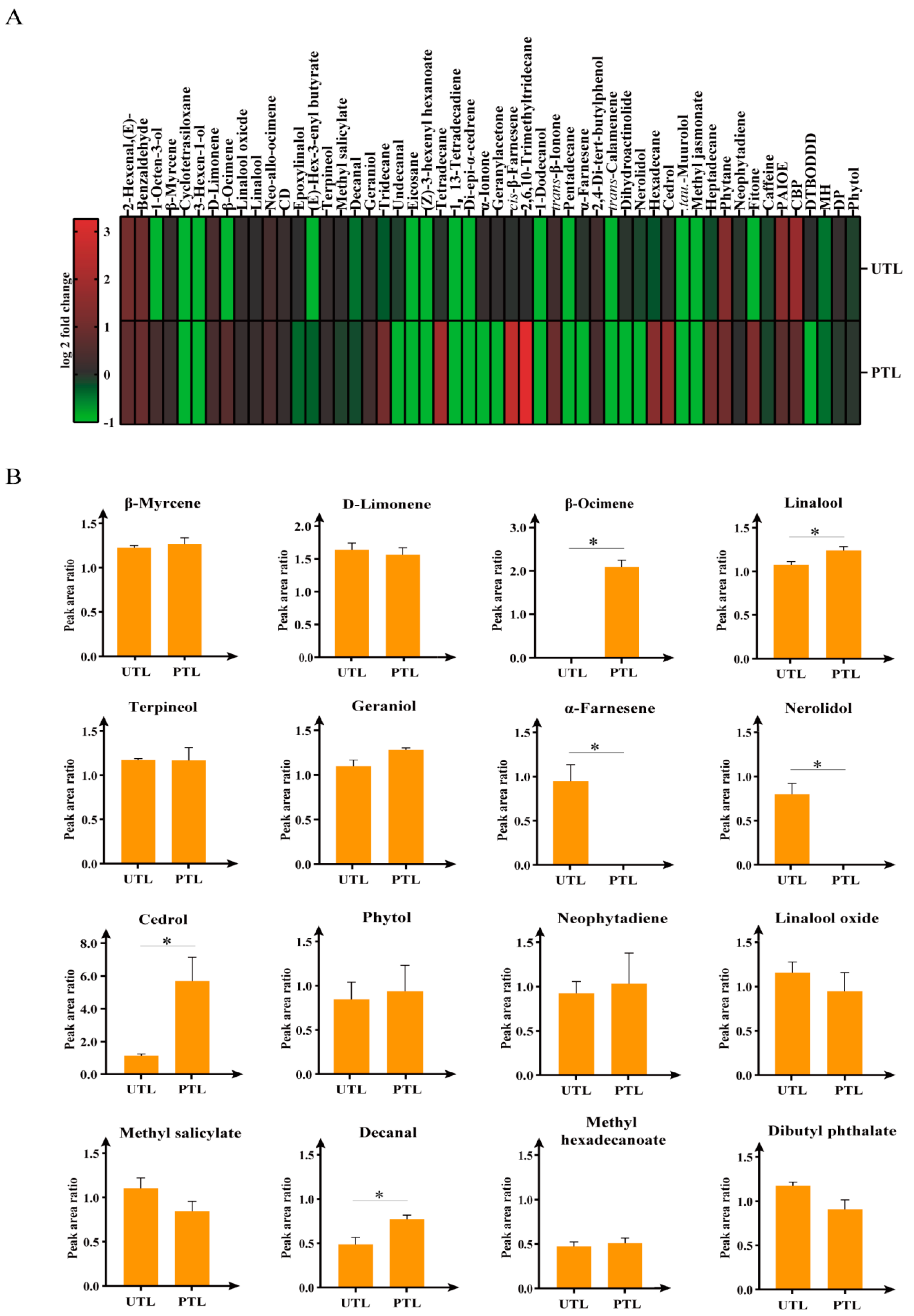
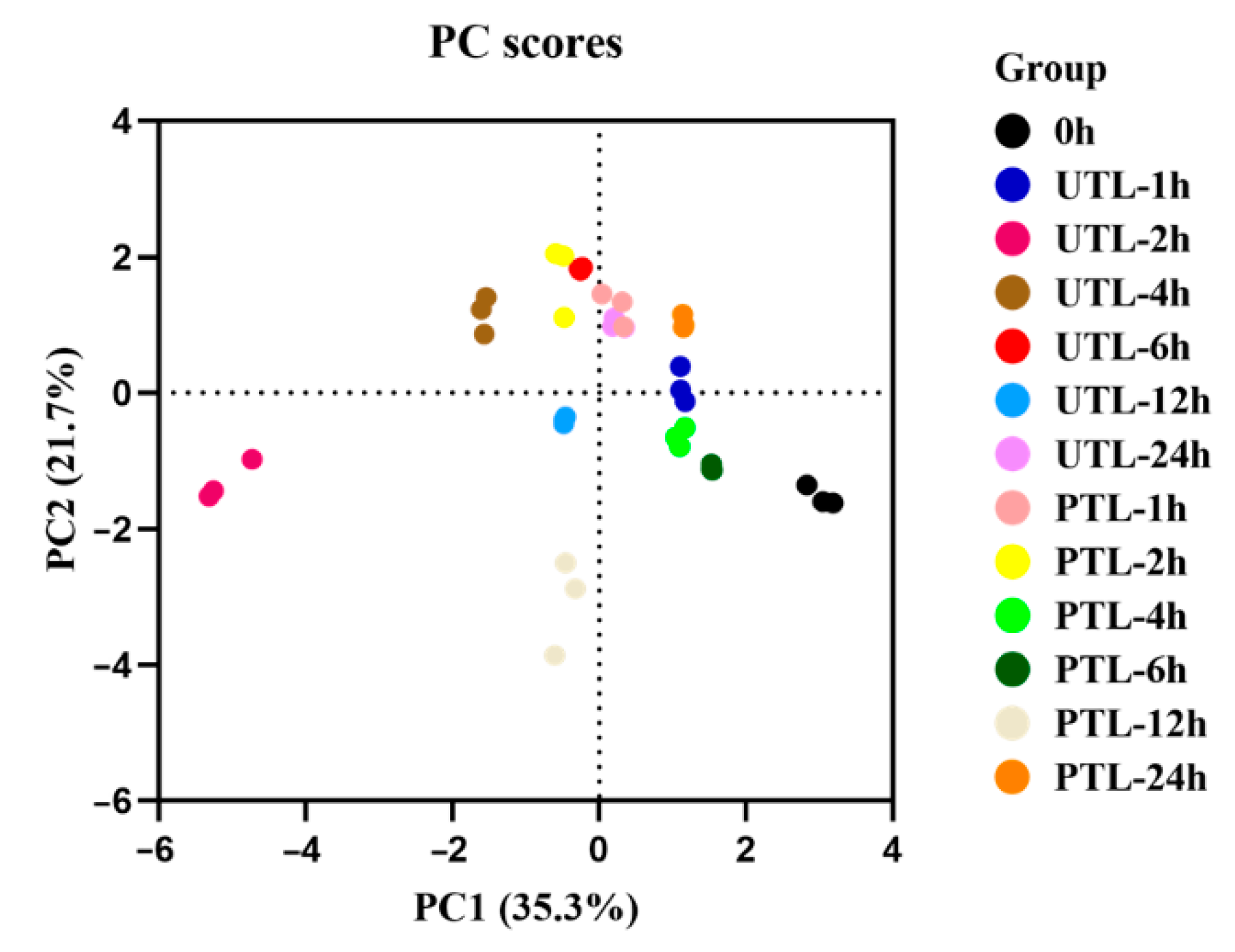
2.3. Comprehensive Analysis of Volatilized Compounds in UTL and PTL
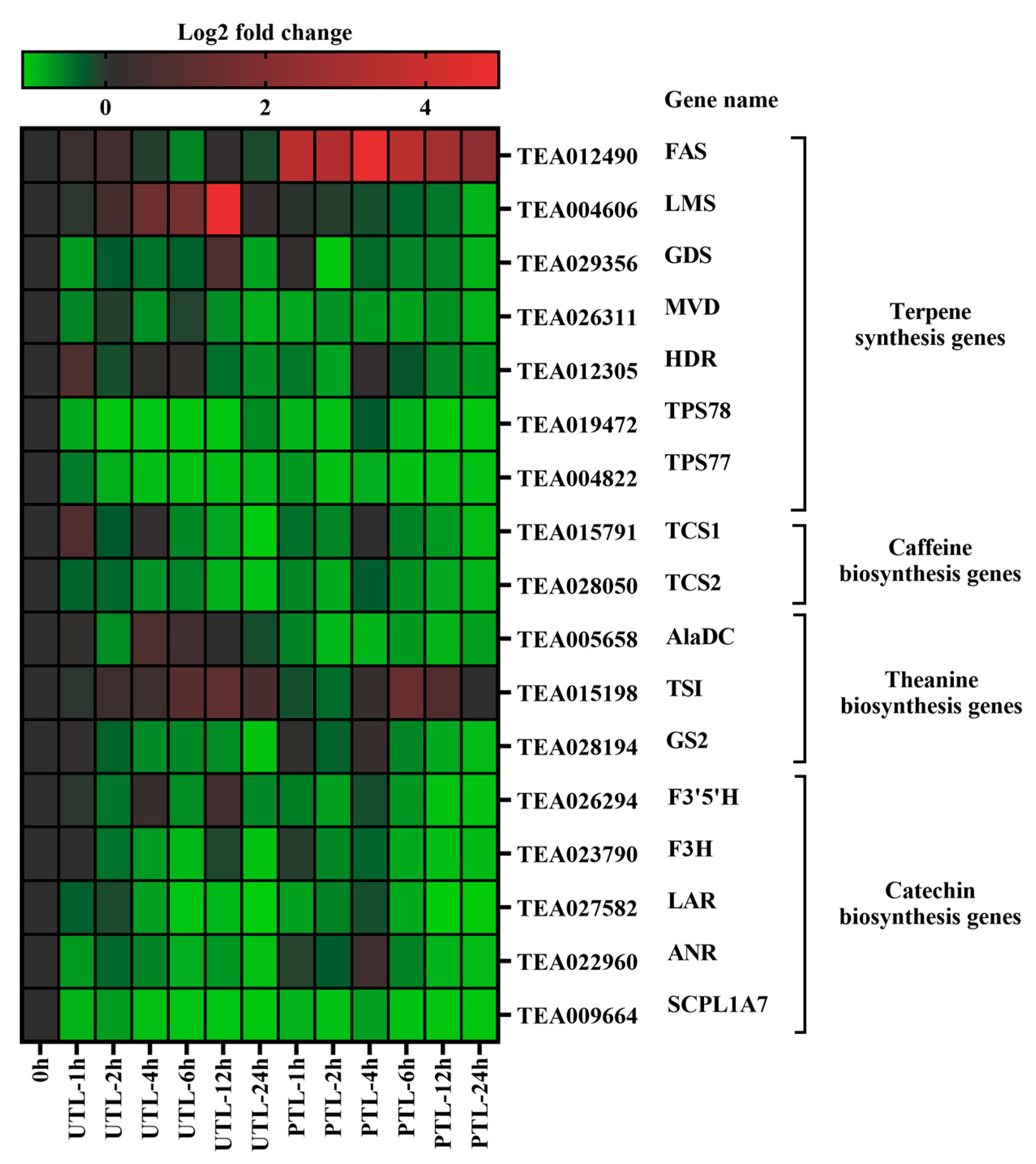
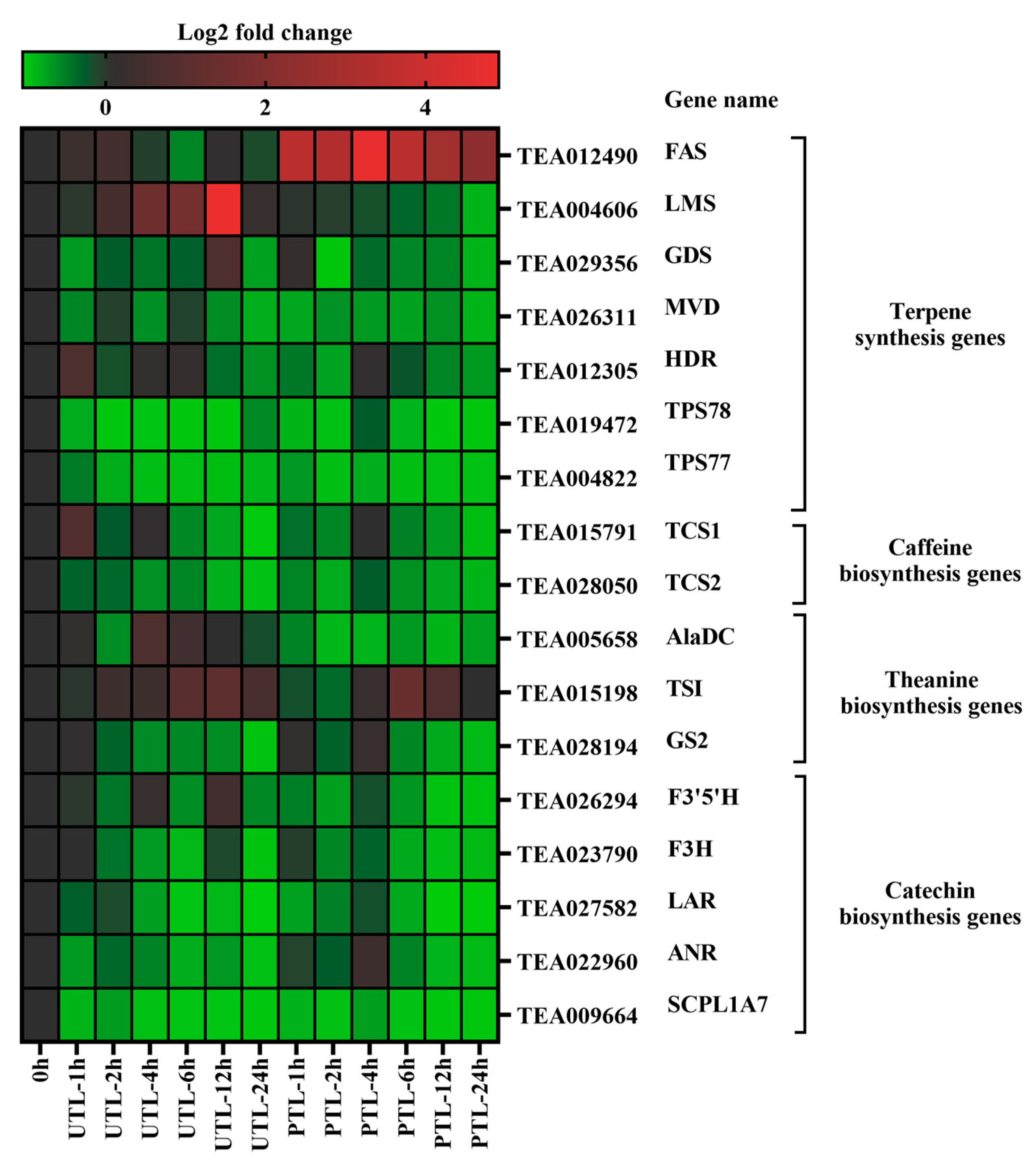
2.4. Expression Levels of Metabolite Biosynthesis Genes during Storage
2.5. A Comprehensive Evaluation of Quality in UTL and PTL
3. Materials and Methods
3.1. Plant Materials and Sampling
3.2. Measurement of Water Content
3.3. Measurement of Color Difference
3.4. Measurement of Chlorophyll Fluorescence (Fv/Fm)
3.5. Quantification of Caffeine Contents
3.6. Quantification of Theanine Contents
3.7. Quantification of Catechin Contents
3.8. Analysis of Microextraction Compounds
3.9. Analysis of Gene Expression
3.10. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| PTL | preserved tea leaves; |
| UTL | unpreserved tea leaves; |
| FTL | fresh tea leaves; |
| GB/T8304-2013 | Tea-Determination of Moisture Content; |
| Fv/Fm | maximum photosynthetic efficiency; |
| Fm | maximum fluorescence; |
| Fo | minimum fluorescence; |
| HPLC | High Performance Liquid Chromatography; |
| GB/T 8312-2013 | Tea-Determination of Caffeine Content; |
| GB/T 8313-2018 | Determination of total polyphenols and catechins content in tea; |
| C | catechin; |
| EC | epicatechin; |
| GC | gallocatechin; |
| EGC | epigallocatechin; |
| ECG | epicatechin gallate; |
| GCG | gallocatechin gallate; |
| EGCG | epigallocatechin gallate; |
| HS-SPME/GC-MS | Headspace Solid Phase Micro Extraction/ Gas Chromatography-Mass Spectrometry; |
| DVB/CAR/PDMS | divinylbenzene/carboxen/polydimethylsiloxane; |
| RI | retention indices; |
| RIs | retention indices with the values provided by the NIST 14 database; |
| MS | mass spectrum comparison using NIST 14 library; |
| qRT-PCR | quantitative real-time PCR; |
| PCA | principal component analysis; |
| CD | Cyclopentasiloxane, decamethyl-; |
| PAIOE | Phthalic acid, isobutyl octyl ester; |
| CBP | Cyclohexyl butyl phthalate; |
| DTBODDD | 7,9-Di-tert-butyl-1-oxaspiro (4,5) deca-6,9-diene-2,8-dione; |
| MH | Methyl hexadecanoate; |
| DP | Dibutyl phthalate; |
| CBGs | catechin biosynthesis genes; |
| FBGs | caffeine biosynthesis genes; |
| TBGs | theanine biosynthesis genes; |
| TPSGs | terpene synthesis genes; |
| FAS | farnesene synthase; |
| LMS | limonene synthase; |
| GDS | germacrene D synthase; |
| MVD | mevalonate-5-pyrophosphate decarboxylase; |
| HDR | 4-hydroxy-3-methylbutenlyl diphosphate reductase; |
| TPS78 | terpene synthase 78; |
| TPS77 | terpene synthase 77; |
| TCS1 | tea caffeine synthase 1; |
| TCS2 | tea caffeine synthase 2; |
| AlaDC | alanine decarboxylase; |
| TSI | theanine synthetase; |
| GS2 | glutamine synthetase 2; |
| F3’5’H | flavonoid 3’,5’-hydroxylase; |
| F3H | flavonoid 3-hydroxylase; |
| LAR | leucoanthocyanidin reductase; |
| ANR | anthocyanidin reductase; |
| SCPL1A7 | serine carboxypeptidase-like acyltransferases 7; |
| PC1 | the first Principal Component; |
| PC2 | the second Principal Component. |
References
- Yu, X.; Li, Y.; He, C.; Zhou, J.; Chen, Y.; Yu, Z.; Wang, P.; Ni, D. Nonvolatile Metabolism in Postharvest Tea (Camellia sinensis L.) Leaves: Effects of Different Withering Treatments on Nonvolatile Metabolites, Gene Expression Levels, and Enzyme Activity. Food Chem. 2020, 327, 126992. [Google Scholar] [CrossRef]
- Gong, A.-D.; Lian, S.-B.; Wu, N.-N.; Zhou, Y.-J.; Zhao, S.-Q.; Zhang, L.-M.; Cheng, L.; Yuan, H.-Y. Integrated Transcriptomics and Metabolomics Analysis of Catechins, Caffeine and Theanine Biosynthesis in Tea Plant (Camellia sinensis) over the Course of Seasons. BMC Plant Biol. 2020, 20, 294. [Google Scholar] [CrossRef]
- Rawat, R.; Gulati, A.; Kiran Babu, G.D.; Acharya, R.; Kaul, V.K.; Singh, B. Characterization of Volatile Components of Kangra Orthodox Black Tea by Gas Chromatography-Mass Spectrometry. Food Chem. 2007, 105, 229–235. [Google Scholar] [CrossRef]
- Brasil, I.M.; Siddiqui, M.W. Chapter 1—Postharvest quality of fruits and vegetables: An overview. In Preharvest Modulation of Postharvest Fruit and Vegetable Quality; Siddiqui, M.W., Ed.; Academic Press: Basel, Switzerland, 2018; pp. 1–40. ISBN 978-0-12-809807-3. [Google Scholar]
- Krahe, J.; Krahe, M.A.; Naumovski, N. The Implications of Post-Harvest Storage Time and Temperature on the Phytochemical Composition and Quality of Japanese-Styled Green Tea Grown in Australia: A Food Loss and Waste Recovery Opportunity. Beverages 2021, 7, 25. [Google Scholar] [CrossRef]
- Guo, J.; Chen, J.; Zhang, M.; Lv, E. Research Status and Prospect of Storage and Transportation Technology of Fresh Tea Leaves. China Tea 2020, 42, 11–15. [Google Scholar]
- Katsuno, T.; Kasuga, H.; Kusano, Y.; Yaguchi, Y.; Tomomura, M.; Cui, J.; Yang, Z.; Baldermann, S.; Nakamura, Y.; Ohnishi, T.; et al. Characterisation of Odorant Compounds and Their Biochemical Formation in Green Tea with a Low Temperature Storage Process. Food Chem. 2014, 148, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; He, C.; Li, Y.; Zhou, J.; Chen, Y.; Yu, Z.; Wang, Y.; Ni, D.; Yu, X.; He, C.; et al. Effects of Different Spreading Treatments on the Formation of Aroma Quality in Green Tea. Beverage Plant Res. 2021, 1, 1–11. [Google Scholar] [CrossRef]
- Cui, J.; Zhai, X.; Guo, D.; Du, W.; Gao, T.; Zhou, J.; Schwab, W.G.; Song, C. Characterization of Key Odorants in Xinyang Maojian Green Tea and Their Changes During the Manufacturing Process. J. Agric. Food Chem. 2022, 70, 279–288. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, F.; Sun, J.; Ni, L. Dynamic Changes of Volatile and Phenolic Components during the Whole Manufacturing Process of Wuyi Rock Tea (Rougui). Food Chem. 2022, 367, 130624. [Google Scholar] [CrossRef]
- Hu, D.; Chen, W.; Ma, C.; Lv, E.; Miao, A.; Qiao, X. Changes in Contents of Quality Related Components of Tea Leaves under Different Preservation. Modern Food Sci. Technol. 2021, 95, 110–115. [Google Scholar] [CrossRef]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft Genome Sequence of Camellia Sinensis Var. Sinensis Provides Insights into the Evolution of the Tea Genome and Tea Quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Li, Y.; Li, M.; Wang, Y.; Zhang, L.; Wan, X.; Yang, X. Tea Aroma Formation from Six Model Manufacturing Processes. Food Chem. 2019, 285, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ho, C.-T.; Schwab, W.; Wan, X. Effect of the Roasting Degree on Flavor Quality of Large-Leaf Yellow Tea. Food Chem. 2021, 347, 129016. [Google Scholar] [CrossRef]
- Ho, C.-T.; Zheng, X.; Li, S. Tea Aroma Formation. Food Sci. Hum. Wellness 2015, 4, 9–27. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Li, J.; Chen, W.; Wang, W.; Qi, D.; Pang, S.; Miao, A. Study of the Aroma Formation and Transformation during the Manufacturing Process of Oolong Tea by Solid-Phase Micro-Extraction and Gas Chromatography–Mass Spectrometry Combined with Chemometrics. Food Res. Int. 2018, 108, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Qiao, D.; Mi, X.; An, Y.; Xie, H.; Cao, K.; Chen, H.; Chen, M.; Liu, S.; Chen, J.; Wei, C. Integrated Metabolic Phenotypes and Gene Expression Profiles Revealed the Effect of Spreading on Aroma Volatiles Formation in Postharvest Leaves of Green Tea. Food Res. Int. 2021, 149, 110680. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Wang, H.; Niu, Y.; Liu, Q.; Zhu, J.; Chen, H.; Ma, N. Characterization of Aroma Compositions in Different Chinese Congou Black Teas Using GC–MS and GC–O Combined with Partial Least Squares Regression. Flavour Fragr. J. 2017, 32, 265–276. [Google Scholar] [CrossRef]
- Yang, Y.; Hua, J.; Deng, Y.; Jiang, Y.; Qian, M.C.; Wang, J.; Li, J.; Zhang, M.; Dong, C.; Yuan, H. Aroma Dynamic Characteristics during the Process of Variable-Temperature Final Firing of Congou Black Tea by Electronic Nose and Comprehensive Two-Dimensional Gas Chromatography Coupled to Time-of-Flight Mass Spectrometry. Food Res. Int. 2020, 137, 109656. [Google Scholar] [CrossRef]
- Ma, W.; Kang, X.; Liu, P.; She, K.; Zhang, Y.; Lin, X.; Li, B.; Chen, Z. The NAC-like Transcription Factor CsNAC7 Positively Regulates the Caffeine Biosynthesis-Related Gene YhNMT1 in Camellia sinensis. Hortic. Res. 2022, 9, uhab046. [Google Scholar] [CrossRef]
- Hangzhou Tea Research Institute, China Coop; National Center for Tea Quality Inspection and Testing; Xiamen Empereur Tea Co., LTD. Tea-Determination of Moisture Content; China Standards Press: Beijing, China, 2013. [Google Scholar]
- Shen, T.; Han, M.; Liu, Q.; Yang, C.; Meng, J.; Li, H. Pigment Profile and Gene Analysis Revealed the Reasons of Petal Color Difference of Crabapples. Braz. J. Bot. 2021, 44, 287–296. [Google Scholar] [CrossRef]
- Huang, D.; Qiu, Q.; Wang, Y.; Wang, Y.; Lu, Y.; Fan, D.; Wang, X. Rapid Identification of Different Grades of Huangshan Maofeng Tea Using Ultraviolet Spectrum and Color Difference. Molecules 2020, 25, 4665. [Google Scholar] [CrossRef] [PubMed]
- Biró, B.; Fodor, R.; Szedljak, I.; Pásztor-Huszár, K.; Gere, A. Buckwheat-Pasta Enriched with Silkworm Powder: Technological Analysis and Sensory Evaluation. LWT 2019, 116, 108542. [Google Scholar] [CrossRef]
- Yu, B.; Ming, F.; Liang, Y.; Wang, Y.; Gan, Y.; Qiu, Z.; Yan, S.; Cao, B. Heat Stress Resistance Mechanisms of Two Cucumber Varieties from Different Regions. Int. J. Mol. Sci. 2022, 23, 1817. [Google Scholar] [CrossRef] [PubMed]
- Hangzhou Tea Research Institute, China Coop; National Center for Tea Quality Inspection and Testing. Tea-Determination of Caffeine Content; China Standards Press: Beijing, China, 2013. [Google Scholar]
- Hangzhou Tea Research Institute, China Coop; National Center for Tea Quality Inspection and Testing; National Food Quality Supervision and Inspection Center; Quanzhou Entry-Exit Inspection and Quarantine Bureau and Comprehensive Technical Service Center; Fujian Richun Industrial Co., LTD. Determination of Theanine in Tea-Using High Performance Liquid Chromatography; China Standards Press: Beijing, China, 2017.
- Li, J.; Liu, S.; Chen, P.; Cai, J.; Tang, S.; Yang, W.; Cao, F.; Zheng, P.; Sun, B. Systematic Analysis of the R2R3-MYB Family in Camellia sinensis: Evidence for Galloylated Catechins Biosynthesis Regulation. Front. Plant Sci. 2022, 12, 782220. [Google Scholar] [CrossRef]
- Hangzhou Tea Research Institute, China Coop; National Center for Tea Quality Inspection and Testing; Quanzhou Entry-Exit Inspection and Quarantine Bureau and Comprehensive Technical Service Center; Fujian Richun Industrial Co., LTD. Determination of Total Polyphenols and Catechins Content in Tea; China Standards Press: Beijing, China, 2018.
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ning, J.; Li, D.; Luo, X.; Ding, D.; Song, Y.; Zhang, Z.; Wan, X. Stepwise Identification of Six Tea (Camellia sinensis (L.)) Categories Based on Catechins, Caffeine, and Theanine Contents Combined with Fisher Discriminant Analysis. Food Anal. Methods 2016, 9, 3242–3250. [Google Scholar] [CrossRef]
- Han, Z.-X.; Rana, M.M.; Liu, G.-F.; Gao, M.-J.; Li, D.-X.; Wu, F.-G.; Li, X.-B.; Wan, X.-C.; Wei, S. Green Tea Flavour Determinants and Their Changes over Manufacturing Processes. Food Chem. 2016, 212, 739–748. [Google Scholar] [CrossRef]
- Rawson, H.M.; Begg, J.E.; Woodward, R.G. The Effect of Atmospheric Humidity on Photosynthesis, Transpiration and Water Use Efficiency of Leaves of Several Plant Species. Planta 1977, 134, 5–10. [Google Scholar] [CrossRef]
- GRANTZ, D.A. Plant Response to Atmospheric Humidity. Plant Cell Environ. 1990, 13, 667–679. [Google Scholar] [CrossRef]
- Xia, E.-H.; Zhang, H.-B.; Sheng, J.; Li, K.; Zhang, Q.-J.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Li, W.; et al. The Tea Tree Genome Provides Insights into Tea Flavor and Independent Evolution of Caffeine Biosynthesis. Mol. Plant 2017, 10, 866–877. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Zhang, C.; Lv, Z.; Shen, C. Pre- and Post-Harvest Exposure to Stress Influence Quality-Related Metabolites in Fresh Tea Leaves (Camellia sinensis). Sci. Hortic. 2021, 281, 109984. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Contents | ||||||
|---|---|---|---|---|---|---|
| No. | Compound a | RI b | RT c | Aroma Description d | UTL | PTL |
| 1 | 2-Hexenal, (E)- e | 854 | Green, leafy, fruity | 3.312 ± 0.507 | 2.423 ± 0.237 | |
| 2 | Benzaldehyde e | 967 | 962 | Almond, burnt sugar | 2.957 ± 0.398 | 2.706 ± 0.764 |
| 3 | 1-Octen-3-ol e | 981 | 982 | Sweet, earthy, mushroom-like | n.d. | 1.535 ± 0.176 |
| 4 | β-Myrcene e | 994 | 993 | Woody, resinous, musty | 1.225 ± 0.024 | 1.269 ± 0.069 |
| 5 | Cyclotetrasiloxane, octamethyl- | 1000 | — | n.d. | n.d. | |
| 6 | 3-Hexen-1-ol, acetate, (Z)- e | 1009 | Grass | n.d. | n.d. | |
| 7 | D-Limonene e | 1031 | 1030 | Citrus, lemon, orange-like, green | 1.639 ± 0.103 | 1.566 ±0.105 |
| 8 | β-Ocimene e | 1039 | 1044 | Sweet, herb | n.d. | 2.092±0.159 |
| 9 | Linalool oxide e | 1083 | Flower | 1.155 ± 0.122 | 1.181 ± 0.180 | |
| 10 | Linalool e | 1104 | 1100 | Floral, sweet, grape-like, woody | 1.076 ± 0.035 | 1.238 ± 0.046 |
| 11 | Neo-allo-ocimene e | 1132 | 1131 | Sweet, floral, nutty, herbal, peppery | 1.492 ± 0.054 | 1.606 ± 0.089 |
| 12 | Cyclopentasiloxane, decamethyl e | 1157 | — | 1.226 ± 0.038 | 1.177 ± 0.051 | |
| 13 | Epoxylinalol e | 1176 | 1183 | Floral | 1.055 ± 0.035 | 0.548 ± 0.028 |
| 14 | (E)-Hex-3-enyl butyrate e | 1188 | 1185 | Fruity, green, vanilla, cream | n.d. | 0.612 ± 0.095 |
| 15 | Terpineol e | 1194 | 1190 | Pleasant, floral | 1.173 ± 0.014 | 1.167 ± 0.143 |
| 16 | Methyl salicylate e | 1197 | 1191 | Minty, fresh, sweet | 1.102 ± 0.120 | 0.845 ± 0.110 |
| 17 | Decanal e | 1207 | 1200 | Soap, orange peel, tallow | 0.487 ± 0.079 | 0.768 ± 0.049 |
| 18 | Geraniol e | 1231 | 1250 | Rose-like, sweet, honey-like | 1.133 ± 0.072 | 1.453 ± 0.021 |
| 19 | Tridecane e | 1300 | Alkane | 0.666 ± 0.032 | 3.259 ± 0.312 | |
| 20 | Undecanal e | 1308 | 1308 | Rose, waxy, oily | 0.865 ± 0.150 | n.d. |
| 21 | Eicosane e | 1326 | Alkane | n.d. | n.d. | |
| 22 | (Z)-3-hexenyl hexanoate e | 1383 | Fruity, waxy, green, fatty, winey | n.d. | n.d. | |
| 23 | Tetradecane e | 1400 | Alkane | 0.938 ± 0.072 | 6.995 ± 0.752 | |
| 24 | 1,13-Tetradecadiene | 1410 | — | n.d. | n.d. | |
| 25 | Di-epi-α-cedrene e | 1419 | — | n.d. | n.d. | |
| 26 | α-Ionone e | 1431 | 1433 | Floral, violet-like, powdery, berry-like | 1.153 ± 0.110 | n.d. |
| 27 | Geranylacetone e | 1455 | 1454 | Fresh floral, sweet-rosy | 1.106 ± 0.042 | n.d. |
| 28 | cis-β-Farnesene e | 1458 | 1457 | Citrus, green | 0.984 ± 0.096 | 11.089 ± 0.956 |
| 29 | 2,6,10- Trimethyltridecane | 1462 | 1461 | — | 1.053 ± 0.097 | 18.202 ± 3.610 |
| 30 | 1-Dodecanol e | 1475 | 1480 | Sweet, fatty | n.d. | n.d. |
| 31 | trans-β-Ionone e | 1490 | 1490 | Violet-like, raspberry, floral | 1.395 ± 0.137 | 2.891 ± 0.461 |
| 32 | Pentadecane e | 1500 | Alkane | n.d. | n.d. | |
| 33 | α-Farnesene e | 1510 | 1508 | Woody, green, floral, herbal | 0.944 ± 0.228 | n.d. |
| 34 | 2,4-Di-tert-butylphenol | 1515 | 1518 | — | 1.386 ± 0.123 | 1.579 ± 0.128 |
| 35 | trans-Calamenene e | 1529 | 1529 | Herbal, spicy | n.d. | n.d. |
| 36 | Dihydroactinolide e | 1537 | 1538 | Musky, coumarin-like | 0.941 ± 0.059 | n.d. |
| 37 | Nerolidol e | 1567 | 1567 | Wood, flower, wax | 0.797 ± 0.124 | n.d. |
| 38 | Hexadecane e | 1600 | Alkane | 0.704 ± 0.052 | 4.483 ± 0.112 | |
| 39 | Cedrol e | 1610 | 1609 | Sweet, fruity, cedar-like | 1.147 ± 0.090 | 5.691 ± 1.452 |
| 40 | .tau.-Muurolol e | 1649 | 1648 | Herb, weak spice | n.d. | n.d. |
| 41 | Methyl jasmonate e | 1653 | 1655.4 | Jasmine | n.d. | n.d. |
| 42 | Heptadecane e | 1700 | Alkane | 0.764 ± 0.072 | 3.221 ± 0.309 | |
| 43 | Phytane | 1809 | 1795 | — | 4.193 ± 0.358 | 3.035 ± 0.912 |
| 44 | Neophytadiene e | 1840 | 1837 | Fresh | 0.924 ± 0.132 | 1.031 ± 0.349 |
| 45 | Fitone e | 1847 | 1847 | — | n.d. | 3.681 ± 1.311 |
| 46 | Caffeine | 1854 | 1842 | — | 0.801 ± 0.087 | 0.808 ± 0.142 |
| 47 | Phthalic acid, isobutyl octyl ester | 1872 | — | 4.368 ± 0.364 | 2.104 ± 0.446 | |
| 48 | Cyclohexyl butyl phthalate e | 1919 | 1892 | Mild | 5.801 ± 0.552 | 2.391 ± 0.837 |
| 49 | 7,9-Di-tert-butyl-1-oxaspiro (4,5) deca-6,9-diene-2,8-dione | 1924 | 1916.8 | — | 1.005 ± 0.081 | n.d. |
| 50 | Methyl hexadecanoate e | 1928 | 1925 | Oily, waxy, fatty | 0.471 ± 0.052 | 0.508 ± 0.058 |
| 51 | Dibutyl phthalate e | 1967 | 1969 | Slight, aromatic | 1.173 ± 0.043 | 0.904 ± 0.110 |
| 52 | Phytol e | 2115 | 2116 | Floral, balsam, powdery, waxy | 0.844 ± 0.196 | 0.935 ± 0.295 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, S.; Yu, Z.; Chen, J.; Zheng, P.; Sun, B.; Guo, J.; Liu, S. The Physiology of Postharvest Tea (Camellia sinensis) Leaves, According to Metabolic Phenotypes and Gene Expression Analysis. Molecules 2022, 27, 1708. https://doi.org/10.3390/molecules27051708
Mei S, Yu Z, Chen J, Zheng P, Sun B, Guo J, Liu S. The Physiology of Postharvest Tea (Camellia sinensis) Leaves, According to Metabolic Phenotypes and Gene Expression Analysis. Molecules. 2022; 27(5):1708. https://doi.org/10.3390/molecules27051708
Chicago/Turabian StyleMei, Shuang, Zizi Yu, Jiahao Chen, Peng Zheng, Binmei Sun, Jiaming Guo, and Shaoqun Liu. 2022. "The Physiology of Postharvest Tea (Camellia sinensis) Leaves, According to Metabolic Phenotypes and Gene Expression Analysis" Molecules 27, no. 5: 1708. https://doi.org/10.3390/molecules27051708
APA StyleMei, S., Yu, Z., Chen, J., Zheng, P., Sun, B., Guo, J., & Liu, S. (2022). The Physiology of Postharvest Tea (Camellia sinensis) Leaves, According to Metabolic Phenotypes and Gene Expression Analysis. Molecules, 27(5), 1708. https://doi.org/10.3390/molecules27051708