

Functional Analysis of Malus halliana WRKY69 Transcription Factor (TF) Under Iron (Fe) Deficiency Stress

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Bioinformatic Analysis of WRKY69 Gene
2.3. Cloning and Expression Vector Construction of WRKY69 Gene
2.4. Agrobacterium-Mediated Transformation of Tobacco and Apple Calli
2.5. Treatment with Fe Deffciency in Transgenic Tobacco and Apple Calli
2.6. Transgenic Tobacco Plants and Apple Calli Under Fe (Iron) Deficiency Stress and Determination of Related Indicators
2.7. Data Analysis
3. Results
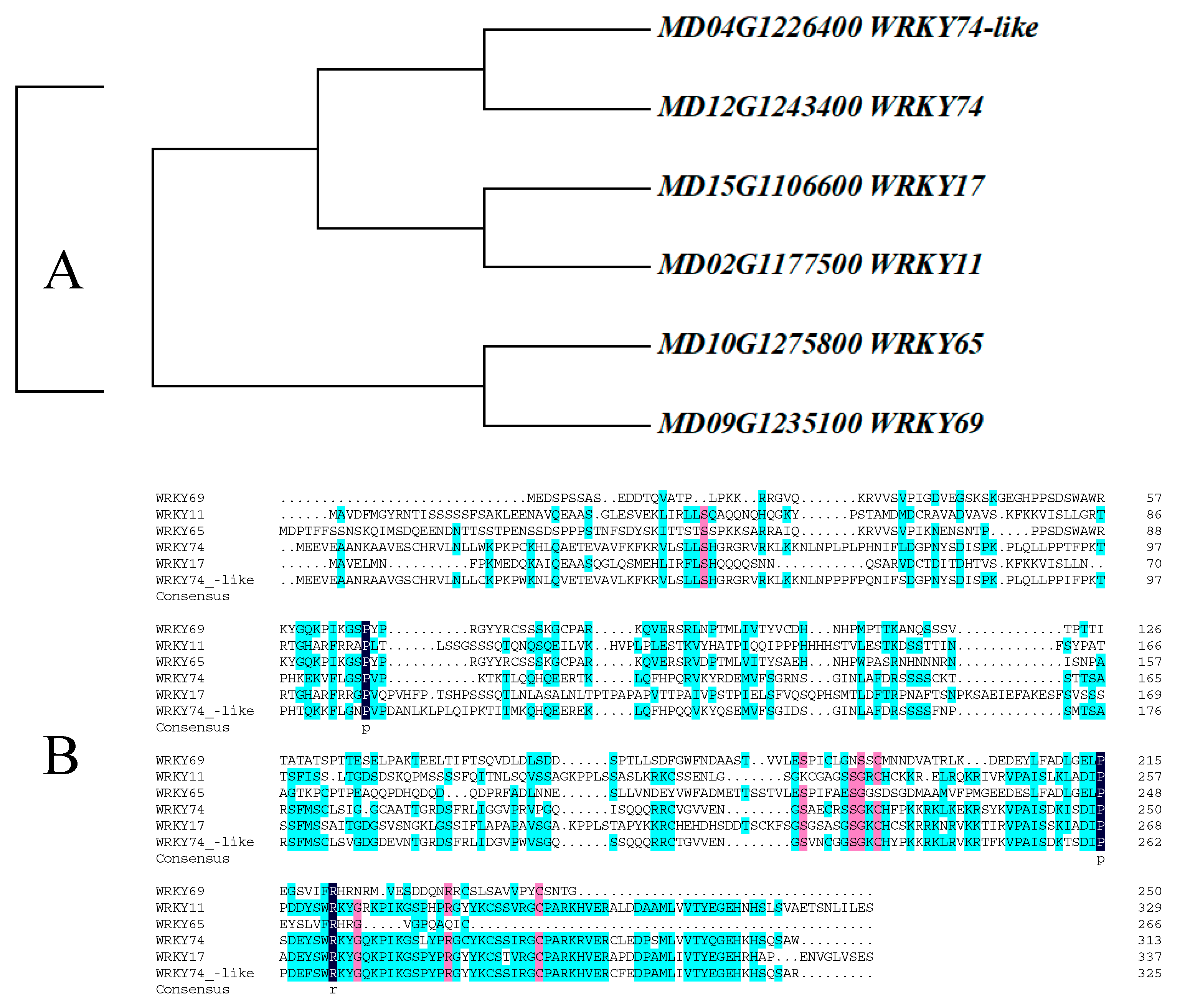
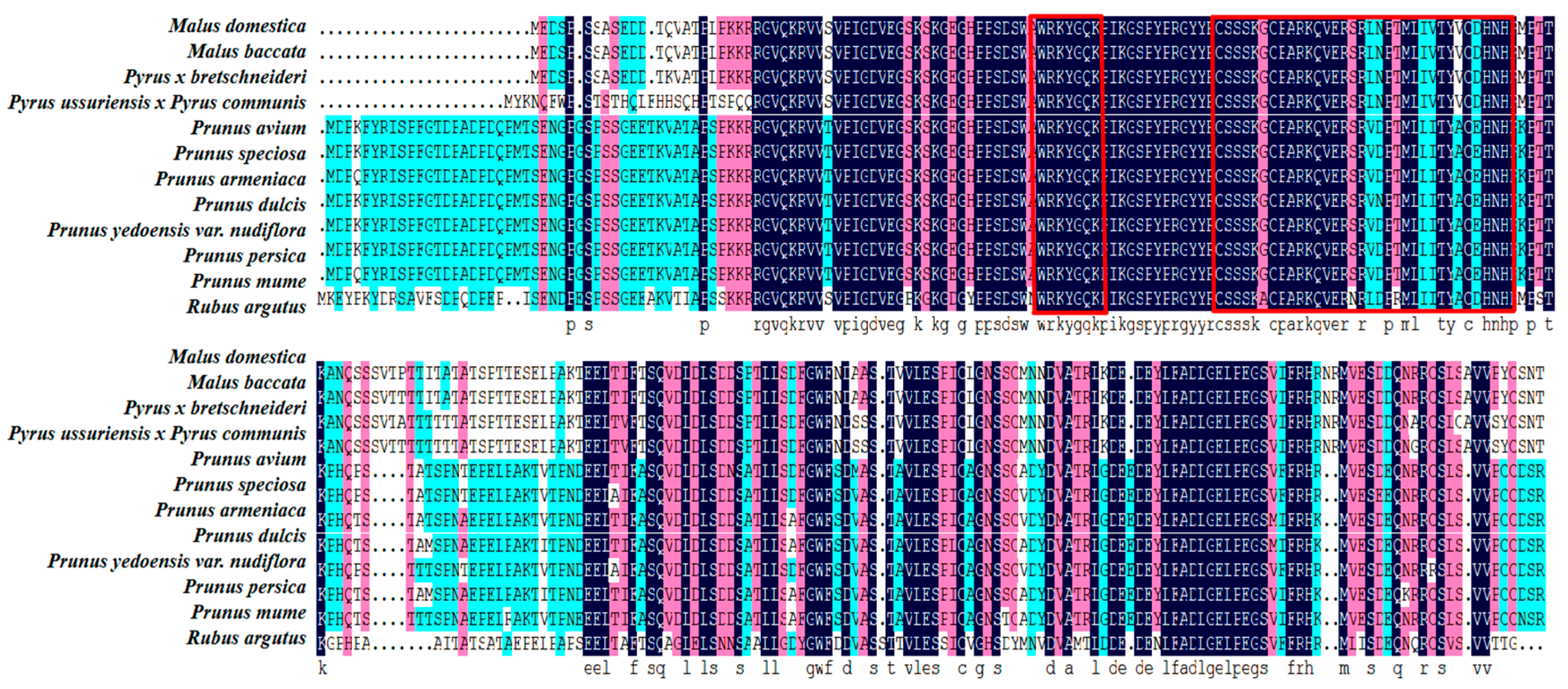
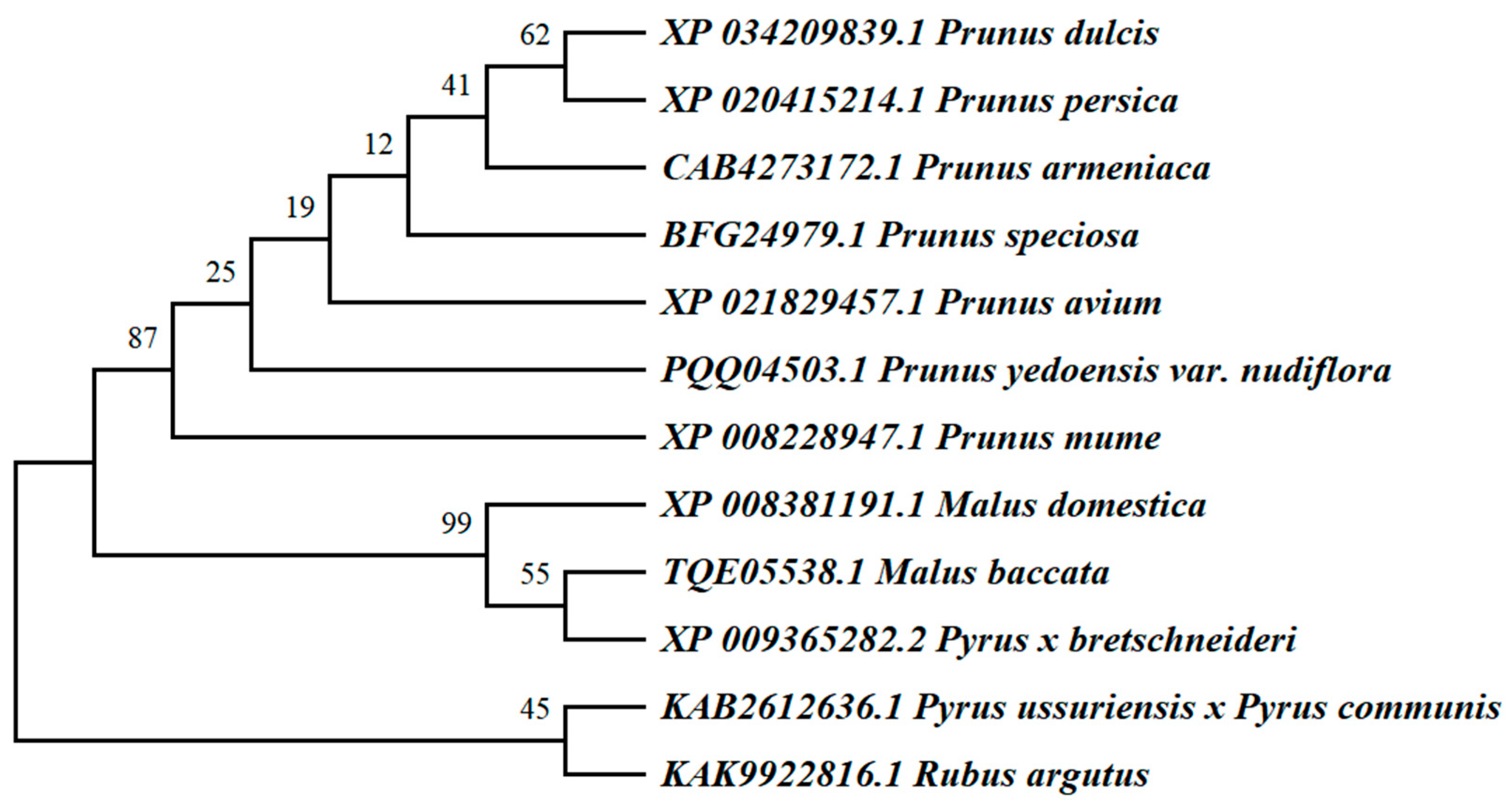
3.1. Protein Sequence Analysis and Evolutionary Relationship Analysis of WRKY69 and Its Homologous Genes
3.2. Analysis of the WRKY69 Gene
3.3. Identiffcation of Transgenic Tobacco and Overexpressed Apple Calli
3.4. Resistance of Transgenic WRKY69 Tobacco to Fe Deffciency Stress
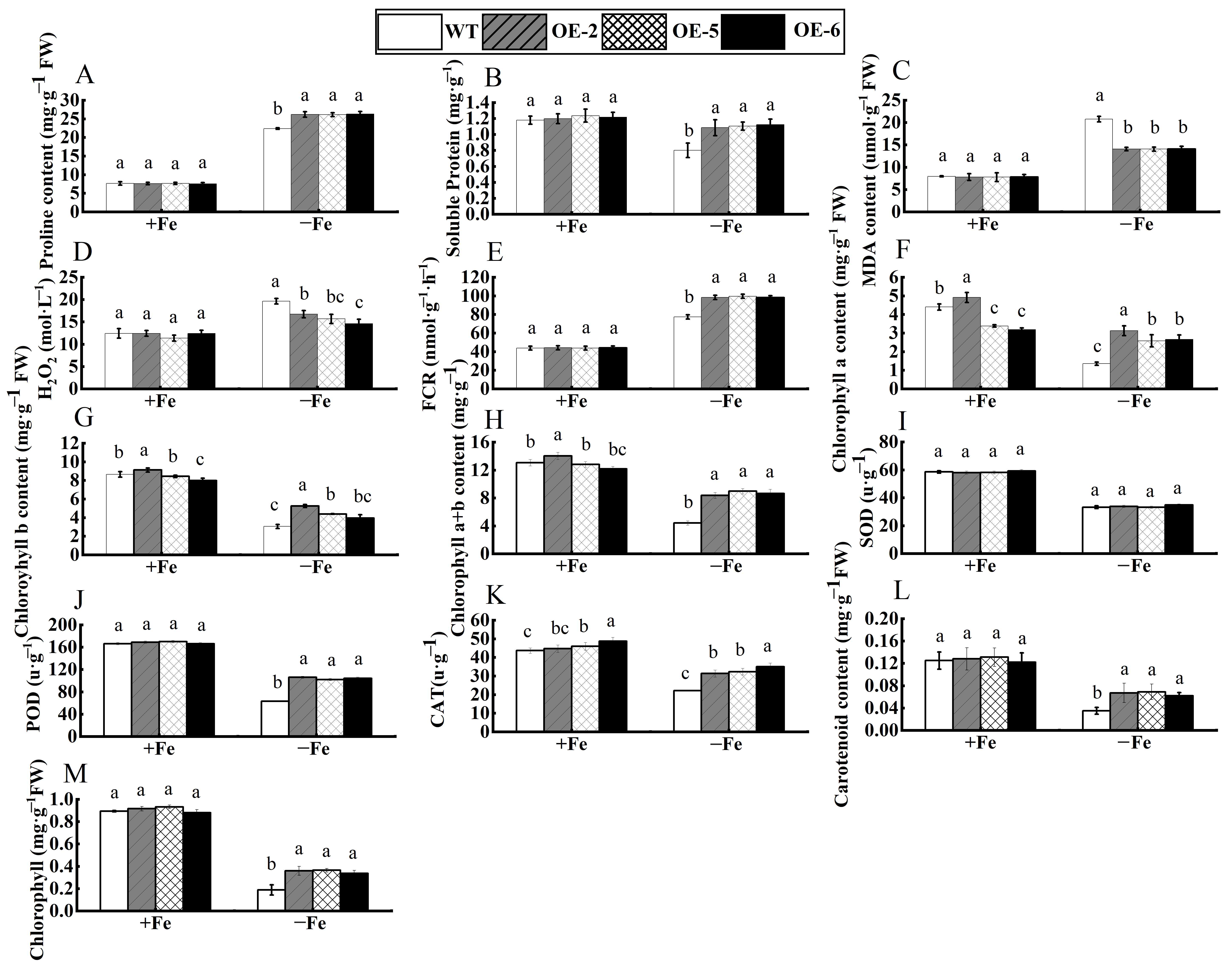
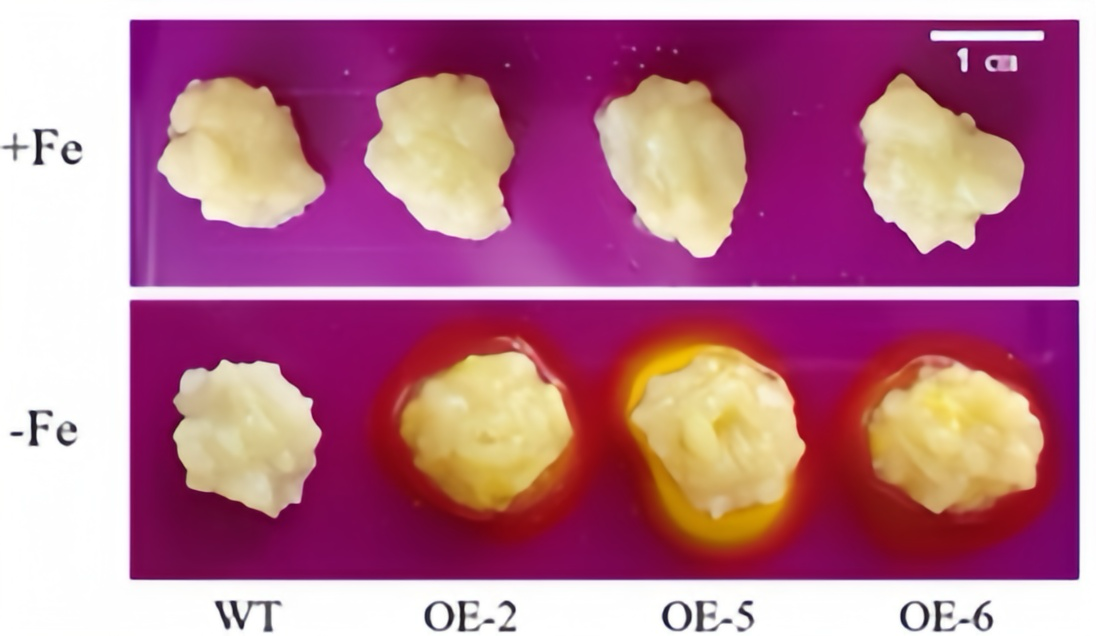
3.5. Functional Analysis of Apple Calli with Overexpression of WRKY69 Gene Under Iron Deficiency Stress
4. Discussion
4.1. Bioinformatics Analysis of WRKY69 TF
4.2. Overexpression of WRKY69-Enhanced Tolerance to Fe Deficiency in Transgenic Plants and Apple Calli
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meena, V.; Kaur, G.; Joon, R.; Shukla, V.; Choudhary, P.; Roy, J.K.; Pandey, A.K. Transcriptome and biochemical analysis in hexaploid wheat with contrasting tolerance to iron deficiency pinpoints multi-layered molecular process. Plant Physiol. Biochem. 2024, 207, 108336. [Google Scholar] [CrossRef] [PubMed]
- Therby-Vale, R.; Lacombe, B.; Rhee, S.Y.; Nussaume, L.; Rouached, H. Mineral nutrient signaling controls photosynthesis: Focus on iron deficiency-induced chlorosis. Trends Plant Sci. 2022, 27, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Briat, J.F.; Fobis-Loisy, I.; Grignon, N.; Lobréaux, S.; Pascal, N.; Savino, G. Cellular and molecular aspects of iron metabolism in plants. Biol. Cell 1995, 84, 69–81. [Google Scholar] [CrossRef]
- Khobra, R.; Ahuja, S.; Singh, B. Chlorophyll biosynthesis as the basis of iron use efficiency under iron deficiency and its relationship with the phytosiderophore synthesis and release in wheat. Indian J. Plant Physiol. 2014, 19, 330–337. [Google Scholar] [CrossRef]
- Walker, E.L.; Waters, B.M. The role of transition metal homeostasis in plant seed development. Curr. Opin. Plant Biol. 2011, 14, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Vigani, G.; Zocchi, G.; Bashir, K.; Philippar, K.; Briat, J.F. Cellular iron homeostasis and metabolism in plant. Front. Plant Sci. 2013, 4, 490. [Google Scholar] [CrossRef]
- Jin, C.W.; Du, S.T.; Chen, W.W.; Li, G.X.; Zhang, Y.S.; Zheng, S.J. Elevated carbon dioxide improves plant iron nutrition through enhancing the iron-deficiency-induced responses under iron-limited conditions in tomato. Plant Physiol. 2009, 15, 272–280. [Google Scholar] [CrossRef]
- Trofimov, K.; Mankotia, S.; Ngigi, M.; Baby, D.; Satbhai, S.B.; Bauer, P. Shedding light on iron nutrition: Exploring intersections of transcription factor cascades in light and iron deficiency signaling. J. Exp. Bot. 2025, 76, 787–802. [Google Scholar] [CrossRef]
- Yang, G.; Li, J.; Liu, W. Molecular cloning and characterization of MxNAS2, a gene encoding nicotianamine synthase in Malus xiaojinensis, with functions in tolerance to iron stress and misshapen flower in transgenic tobacco. Sci. Hortic. 2015, 183, 77–86. [Google Scholar] [CrossRef]
- Darbani, B.; Briat, J.F.; Holm, P.B.; Husted, S.; Noeparvar, S.; Borg, S. Dissecting plant iron homeostasis under short and long-term iron fluctuations. Biotechnol. Adv. 2013, 31, 1292–1307. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Ren, Y.R.; Wang, Q.J. Overexpression of MdbHLH104 gene enhances the tolerance to iron deffciency in apple. Plant Biotechnol. J. 2016, 14, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, E.J.; Waters, B.M. Alkaline stress and iron deficiency regulate iron uptake and riboflavin synthesis gene expression differently in root and leaf tissue: Implications for iron deficiency chlorosis. J. Exp. Bot. 2016, 67, 5671–5685. [Google Scholar] [CrossRef] [PubMed]
- Abadía, J.; Vázquez, S.; Rellán-Álvarez, R.; El-Jendoubi, H.; Abadía, A.; Álvarez-Fernández, A. Towards a knowledge-based correction of iron chlorosis. Plant Physiol. Biochem. 2011, 49, 471–482. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Chen, X.; Wu, X.; Han, C.; Jia, Y.; Wan, X.; Liu, Q.; He, F.; Zhang, F.A. WRKY transcription factor, PyWRKY71, increased the activities of antioxidant enzymes and promoted the accumulation of cadmium in poplar. Plant Physiol. Biochem. 2023, 205, 108163. [Google Scholar] [CrossRef]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stress in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef]
- Han, D.; Wang, Y.; Zhang, Z. Isolation and functional analysis of MxCS3: A gene encoding a citrate synthase in Malus xiaojinensis, with functions in tolerance to iron stress and abnormal flower in transgenic Arabidopsis thaliana. Plant Growth Regul. 2017, 82, 479–489. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.J.; Schmelzer, E. Early nuclear events in plant defense signalling: Rapid gene activation by WRKY transcription factors. Embo J. 1999, 18, 4689–4699. [Google Scholar] [CrossRef]
- Yan, J.Y.; Li, C.X.; Sun, L.; Ren, J.Y.; Li, G.X.; Ding, Z.J.; Zheng, S.J. A WRKY Transcription Factor Regulates Fe Translocation under Fe Deficiency. Plant Physiol. 2016, 171, 2017–2027. [Google Scholar] [CrossRef]
- Dang, F.; Wang, Y.; She, J.; Lei, Y.; Liu, Z.; Eulgem, T.; Lai, Y.; Lin, J.; Yu, L.; Lei, D.; et al. Overexpression of CaWRKY27, a subgroup IIe WRKY transcription factor of Capsicum annuum, positively regulates tobacco resistance to Ralstonia solanacearum infection. Physiol. Plant 2014, 150, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Kou, Y.; Liu, H.; Li, X.; Xiao, J.; Wang, S. OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice. J. Exp. Bot. 2011, 62, 4863–4874. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cheng, X.; Yin, D.; Chen, D.; Luo, C.; Liu, H.; Huang, C. Advances in the research on plant WRKY transcription factors responsive to external stresses. Curr. Issues Mol. Biol. 2023, 45, 2861–2880. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.J.; Yan, J.Y.; Xu, X.Y. WRKY46 functions as a transcriptional repressor of ALMT1, regulating aluminum-induced malate secretion in Arabidopsis. Plant J. 2013, 76, 825–835. [Google Scholar] [CrossRef]
- Huang, Y.; Sun, Z.; Zhou, X. WRKY Transcription Factors in Response to Metal Stress in Plants: A Review. Int. J. Mol. Sci. 2024, 25, 10952. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, H.J.; Yang, X.Y. CsWRKY46, a WRKY transcription factor from cucumber, confers cold resistance in transgenic-plant by regulating a set of cold-stress responsive genes in an ABA-dependent manner. Plant Physiol. Biochem. 2016, 108, 478–487. [Google Scholar] [CrossRef]
- Han, D.; Xu, T.; Han, J.; Liu, W.; Wang, Y.; Li, X.; Yang, G. Overexpression of MxWRKY53 increased iron and high salinity stress tolerance in Arabidopsis thaliana. In Vitro Cell. Dev. Biol.-Plant 2022, 58, 266–278. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473. [Google Scholar] [CrossRef]
- Han, Z.H.; Wang, Q.; Shen, T. Comparison of some physiological and biochemical characteristics between iron-efficient and iron-inefffcient species in the genus malus. J. Plant Nutr. 1994, 17, 1257–1264. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, Z.; Cheng, J.; Wang, S.; Gao, Y.; Xian, X.; Li, C.; Wang, Y. Molecular cloning and functional characterization of MhHEC2-like genes in Malus halliana reveals it enhances Fe(iron) deficiency tolerance. Funct. Integr. Genom. 2022, 22, 1283–1295. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xiang, G.; Sun, Q. Melatonin enhances salt tolerance by promoting MYB108A-mediated ethylene biosynthesis in grapevines. Hortic. Res. 2019, 6, 114. [Google Scholar] [CrossRef]
- Li, H.H.; Liu, X.; An, J.P. Cloning and elucidation of the functional role of apple MdLBD13 in anthocyanin biosynthesis and nitrate assimilation. Plant Cell Tissue Organ Cult. 2017, 130, 47–59. [Google Scholar] [CrossRef]
- Cheng, L.; Zhao, T.; Wu, Y.X.; Wang, H.; Zhang, Z.X.; Zhang, D.; Wang, S.C.; Wang, Y.X. Identification of AP2/ERF genes in apple (Malus×domestica) and demonstration that MdERF017 enhances iron deficiency tolerance. Plant Cell Tissue Organ Cult. 2020, 143, 465–482. [Google Scholar] [CrossRef]
- Ferreira Júnior, D.C.; Gaion, L.A.; Sousa Júnior, G.S.; Santos, D.M.; Carvalho, R.F. Drought-induced proline synthesis depends on root-to-shoot communication mediated by light perception. Acta. Physiol. Plant 2018, 40, 15. [Google Scholar] [CrossRef]
- Xiao, F.; Zhou, H. Plant salt response: Perception, signaling, and tolerance. Front. Plant Sci. 2023, 13, 1053699. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Schikora, A.; Schmidt, W. Iron stress-induced changes in root epidermal cell fate are regulated independently from physiological responses to low iron availability. Plant Physiol. 2001, 125, 1679–1687. [Google Scholar] [CrossRef]
- Zhao, Q.; Ren, Y.R.; Wang, Q.J.; Wang, X.F.; Hao, Y.J. Ubiquitination-related MdBT scaffold proteins target a bHLH transcription factor for iron homeostasis. Plant Physiol. 2016, 172, 1973–1988. [Google Scholar] [CrossRef]
- Shen, T.; Wen, X.; Wen, Z. Genome-wide identiffcation and expression analysis of bHLH transcription factor family in response to cold stress in sweet cherry (Prunus avium L.). Sci. Hortic. 2021, 279, 109905. [Google Scholar] [CrossRef]
- Wang, W.N.; Wei, Y.T.; Zhao, S.T.; Yu, F.H.; Wang, J.W.; Gu, C.Y.; Meng, L.S. ABSCISIC ACID-INSENSITIVE 5-KIP-RELATED PROTEIN 1-SHOOT MERISTEMLESS modulates reproductive development of Arabidopsis. Plant Physiol. 2024, 195, 2309–2322. [Google Scholar] [CrossRef] [PubMed]
- Hell, R.; Stephan, U. Iron uptakem, trafficking and homeostasis in plants. Planta 2003, 216, 541–551. [Google Scholar] [CrossRef]
- Zhang, Z.; Cheng, J.; Wang, W.; Gao, Y.; Li, C.; Wang, Y. Transcription factors dealing with Iron-deficiency stress in plants: Focus on the bHLH transcription factor family. Physiol. Plant. 2023, 175, e14091. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; McIntyre, L.M.; Sherman, L.A. Microarray analysis of the genome-wide response to iron deficiency and iron reconstitution in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2003, 132, 1825–1839. [Google Scholar] [CrossRef] [PubMed]
- Moseley, L.J.; Allinger, T.; Herzog, S.; Hoerth, P.; Wehinger, E.; Merchant, S. Adaptation to Fe-deficiency requires remodeling of the photosynthetic apparatus. EMBO J. 2002, 21, 6709–6720. [Google Scholar] [CrossRef]
- Venkateswarlu, Y.; Jolley, C.C.; Chandramouli, M. Alteration of proteins and pigments inffuence the function of photosystem I under iron deffciency from Chlamydomonas reinhardtii. PLoS ONE 2012, 7, e35084. [Google Scholar]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Tian, A.G.; Zou, H.F.; Lei, G.; Huang, J.; Wang, C.M.; Wang, H.W.; Zhang, J.S.; Chen, S.Y. Soybean WRKY-type transcription factor genes, GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants. Plant Biotechnol. J. 2008, 6, 486–503. [Google Scholar] [CrossRef]
- Liu, Q.; Li, X.; Yan, S. OsWRKY67 positively regulates blast and bacteria blight resistance by direct activation of PR genes in rice. BMC Plant Bio. 2018, 18, 257. [Google Scholar] [CrossRef]
- Wang, C.T.; Ru, J.N.; Liu, Y.W.; Yang, J.F.; Li, M.; Xu, Z.S.; Fu, J.D. The maize WRKY transcription factor ZmWRKY40 confers drought resistance in transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, b2580. [Google Scholar] [CrossRef]
- Qin, Y.; Yu, H.; Cheng, S.; Liu, Z.; Yu, C.; Zhang, X.; Gong, X. Genome-wide analysis of the WRKY gene family in Malus domestica and the role of MdWRKY70L in response to drought and salt stresses. Genes 2022, 13, 1068. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Somssich, I.E.; Ringler, P. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Wu, K.L.; Guo, Z.J.; Wang, H.H. The WRKY family of transcription factors in rice and Arabidopsis and their origins. DNA Res. 2005, 12, 9–26. [Google Scholar] [CrossRef]
- Cai, M.; Qiu, D.; Yuan, T. Identification of novel pathogen-responsive cis-elements and their binding proteins in the promoter of OsWRKY13, a gene regulating rice disease resistance. Plant Cell Environ. 2010, 31, 86–96. [Google Scholar] [CrossRef]
- Jiang, Y.; Duan, Y.; Yin, J.; Ye, S.; Zhu, J.; Zhang, F.; Lu, W.; Fan, D.; Luo, K. Genome-wide identification and characterization of the Populus WRKY transcription factor family and analysis of their expression in response to biotic and abiotic stresses. J. Exp. Bot. 2014, 65, 6629–6644. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Wang, R. Gene cloning and expression analysis of benzoic acid stress responsive transcription factor WRKY7 in Panax ginseng. Zhong Guo Zhong Yao Za Zhi 2017, 42, 1130–1135. [Google Scholar]
- Han, D.G.; Hou, Y.J.; Ding, H.B. Isolation and preliminary functional analysis of MbWRKY4 gene involved in salt tolerance in transgenic tobacco. Int. J. Agric. Biol. 2018, 20, 2045–2052. [Google Scholar]
- Han, D.; Zhou, Z.; Du, M.; Li, T.; Wu, X.; Yu, J.; Zhang, P.; Yang, G. Overexpression of a Malus xiaojinensis WRKY transcription factor gene (MxWRKY55) increased iron and high salinity stress tolerance in Arabidopsis thaliana. In Vitro Cell. Dev. Biol.-Plant 2020, 56, 600–609. [Google Scholar] [CrossRef]
- Nei, M. Phylogenetic analysis in molecular evolutionary genetics. Annu. Rev. Genet. 1996, 30, 371–403. [Google Scholar] [CrossRef]
- Xiu, H.; Nuruzzaman, M.; Guo, X.Q. Molecular cloning and expression analysis of eight PgWRKY genes in panax ginseng responsive to salt and hormones. Int. J. Mol. Sci. 2016, 17, 303–319. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.B.; Xia, Z.L.; Cai, Z.D. GmWRKY16 enhances drought and salt tolerance through an ABA-mediated pathway in Arabidopsis thaliana. Front. Plant Sci. 2019, 9, 1979. [Google Scholar] [CrossRef]
- Shi, W.Y.; Du, Y.T.; Ma, J. The WRKY transcription factor GmWRKY12 confers drought and salt tolerance in soybean. Int. J. Mol. Sci. 2018, 19, 4087. [Google Scholar] [CrossRef]
- Hu, Y.; Zhu, Y.F.; Guo, A.X.; Jia, X.M.; Cheng, L.; Zhao, T.; Wang, Y.X. Transcriptome analysis in Malus halliana roots in response to iron deficiency reveals insight into sugar regulation. Mol. Genet. Genom. 2018, 293, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- Santi, S.; Schmidt, W. Dissecting iron deffciency-induced proton extrusion in Arabidopsis roots. New Phytol. 2009, 183, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.J.; Zhang, C.L.; Zhang, R.F.; Wang, G.L.; Li, Y.Y.; Hao, Y.J. The SUMO E3 ligase MdSIZ1 targets MdbHLH104 to regulate plasma membrane H+-ATPase activity and iron homeostasis. Plant Physiol. 2019, 179, 88-106. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. Interaction of proline, sugars, and anthocyanins during photosynthetic acclimation of Arabidopsis thaliana to drought stress. J. Plant Physiol. 2012, 169, 577–585. [Google Scholar] [CrossRef]
- Zhu, H.; Zhou, Y.; Zhai, H. A novel sweetpotato WRKY transcription factor, IbWRKY2, positively regulates drought and salt tolerance in transgenic Arabidopsis. Biomolecules 2020, 10, 506. [Google Scholar] [CrossRef]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline-alkali stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef]
- Dong, H.; Wu, C.; Luo, C.; Wei, M.; Qu, S.; Wang, S. Overexpression of MdCPK1a gene, a calcium dependent protein kinase in apple, increase tobacco cold tolerance via scavenging ROS accumulation. PLoS ONE 2020, 15, e0242139. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, L.; Wang, B.; Zhang, J.; Nie, J. Dynamic degradation of penconazole and its effect on antioxidant enzyme activity and malondialdehyde content in apple fruit. Sci. Hortic. 2022, 300, 111053. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, L.; Chen, J.; Tao, L.; An, Y.; Cai, H.; Guo, C. Overexpression of the alfalfa WRKY11 gene enhances salt tolerance in soybean. PLoS ONE 2018, 13, e0192382. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, N.N.; Pan, Q. Hydrogen sulphide alleviates iron deffciency by promoting iron availability and plant hormone levels in Glycine max seedlings. BMC Plant Biol. 2020, 20, 383. [Google Scholar] [CrossRef]
- Chen, C.; Cao, Q.; Jiang, Q. Comparative transcriptome analysis reveals gene network regulating cadmium uptake and translocation in peanut roots under iron deffciency. BMC Plant Biol. 2019, 19, 35. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Size/aa | Mw /kDa | Instability Index | pI | Aliphatic Index | GRAVY |
|---|---|---|---|---|---|---|---|
| MD09G1235100 | WRKY69 | 250 | 61.739 | 45.46 | 5.10 | 28.29 | 0.916 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, H.; Liu, W.; Wang, X.; Wang, Y. Functional Analysis of Malus halliana WRKY69 Transcription Factor (TF) Under Iron (Fe) Deficiency Stress. Curr. Issues Mol. Biol. 2025, 47, 576. https://doi.org/10.3390/cimb47070576
Luo H, Liu W, Wang X, Wang Y. Functional Analysis of Malus halliana WRKY69 Transcription Factor (TF) Under Iron (Fe) Deficiency Stress. Current Issues in Molecular Biology. 2025; 47(7):576. https://doi.org/10.3390/cimb47070576
Chicago/Turabian StyleLuo, Hongjia, Wenqing Liu, Xiaoya Wang, and Yanxiu Wang. 2025. "Functional Analysis of Malus halliana WRKY69 Transcription Factor (TF) Under Iron (Fe) Deficiency Stress" Current Issues in Molecular Biology 47, no. 7: 576. https://doi.org/10.3390/cimb47070576
APA StyleLuo, H., Liu, W., Wang, X., & Wang, Y. (2025). Functional Analysis of Malus halliana WRKY69 Transcription Factor (TF) Under Iron (Fe) Deficiency Stress. Current Issues in Molecular Biology, 47(7), 576. https://doi.org/10.3390/cimb47070576