Abstract
Obesity is a chronic metabolic disorder and a growing global public health challenge, affecting hundreds of millions of individuals worldwide. While diet and physical activity are well-established contributors, increasing evidence underscores the critical role of epigenetic mechanisms in mediating obesity-related processes. Epigenetic modifications—such as DNA methylation, RNA methylation (particularly N6-methyladenosine), histone modifications, non-coding RNAs, and chromatin remodeling—modulate gene expression without altering the DNA sequence. This review aims to provide an overview of the epigenetic mechanisms involved in obesity, with an emphasis on their molecular functions and regulatory networks. Integrating findings from relevant studies, we discuss how these modifications influence obesity-related outcomes through regulating key processes such as adipocyte differentiation and energy metabolism. Advancing our understanding of epigenetic regulation may pave the way for novel, targeted strategies in the prevention and treatment of obesity.
1. Introduction
Obesity is an increasingly severe global public health issue. According to a 2022 report by the World Health Organization (WHO), obesity affects approximately one-eighth of the worldwide population [1]. Beyond its impact on individual quality of life, obesity is closely associated with various metabolic diseases, including type 2 diabetes, metabolic-associated fatty liver disease, hypertension, and multiple malignancies such as endometrial and colorectal cancer [2,3,4]. Fundamentally, obesity results from a chronic imbalance between energy intake and expenditure [5]. A key factor in this process is the heterogeneity of adipose tissue: brown adipose tissue (BAT) and beige adipose tissue contribute to energy expenditure through thermogenesis, whereas white adipose tissue (WAT) regulates energy homeostasis by storing and releasing fatty acids [6]. These adipose tissue types affect the metabolic process of excessive fat accumulation by regulating the dynamic balance between energy consumption and storage [7].
Obesity is a complex condition influenced by genetic and environmental factors [8]. Genetic factors establish the basis for an individual’s susceptibility to obesity, while environmental elements such as diet and physical activity directly impact the development of obesity [9]. Epigenetics—defined as heritable changes in gene function that occur without alterations to the DNA sequence—serves as a critical interface between environmental inputs and gene regulation [10,11,12]. The field of epigenetics has gained increasing prominence for its role in modulating gene expression through mechanisms such as DNA methylation, RNA methylation, and histone modifications [11]. Growing evidence supports the involvement of epigenetic dysregulation in obesity [12,13]. In obese individuals, abnormal histone modifications and alterations in DNA methylation levels are closely linked to key pathological mechanisms such as adipose tissue dysfunction, insulin resistance (IR), and chronic inflammation [14,15]. Moreover, interventions like bariatric surgery have been shown to reverse certain epigenetic modifications, suggesting their potential as therapeutic targets [16].
Among the various epigenetic mechanisms, DNA methylation, RNA methylation (particularly N6-methyladenosine (m6A)), histone modifications, non-coding RNAs, and chromatin remodeling have emerged as crucial regulators of adipose tissue development and energy metabolism. This review summarizes their regulatory mechanisms on adipose tissue development and energy metabolism. A deeper understanding of these mechanisms may facilitate the identification of early diagnostic biomarkers and the development of innovative epigenetic therapies for obesity.
2. Epigenetic Regulation of Obesity and Underlying Mechanisms
2.1. DNA Methylation and Obesity
DNA methylation is an epigenetic mechanism involving the transfer of a methyl group at the C5 position of the cytosine to form 5-methylcytosine (5mC) [17,18]. Within the genome, cytosine-phosphate-guanine (CpG) sites often cluster into regions known as CpG islands, which are frequently located in gene promoter regions [19]. Extensive research has demonstrated that DNA methylation modifications related to obesity are mostly concentrated at specific CpG sites. Through comparing obese and non-obese humans, researchers have identified 114 differentially methylated positions (DMPs) in CpG islands [20]. Furthermore, genome-wide DNA methylation analyses of adipocytes from obese individuals versus non-obese individuals have revealed that most altered CpG sites are significantly associated with obesity-related gene expression [21]. Notably, lifestyle interventions such as caloric restriction and bariatric surgery have demonstrated the capacity to partially reverse these methylation alterations, highlighting their potential plasticity and therapeutic relevance [22,23].
DNA methylation is catalyzed by DNA methyltransferases (DNMTs), primarily DNMT1, DNMT3A, and DNMT3B. Three conserved DNMTs are responsible for the de novo establishment and maintenance of DNA methylation patterns [24,25]. Conversely, active DNA demethylation is mediated by the ten-eleven translocation (TET) family of methylcytosine dioxygenases [26]. These methylation marks are interpreted by methyl-binding proteins (MBPs), which recognize methylated CpG sites and recruit chromatin remodeling complexes to activate or repress transcription, depending on the cellular context [27]. In obesity research, overexpression of DNMTs results in excessive methylation of promoter regions in genes regulating lipid metabolism, leading to gene silencing and excessive fat accumulation [28,29]. In contrast, DNMT deficiency can alleviate aberrant methylation in obesity-related genes, restoring their transcriptional activity and mitigating pathological fat deposition [30] (Table 1).

Table 1.
The link between DNA methylation and obesity.
2.1.1. Regulation of DNA Methylation in Adipose Tissue Development and Differentiation
DNMTs play a pivotal role in the regulation of adipose tissue development and differentiation by modulating the methylation status of genes central to adipogenesis and metabolic function. Studies have shown that DNMT1 exhibits stage-specific regulation during the differentiation of 3T3-L1 preadipocytes. In early differentiation, inhibition of DNMT1 reduces DNA methylation at the promoter region of wingless-type MMTV integration site family, member 10a (Wnt10a)—a known anti-adipogenic gene—thereby promoting its expression. Overexpression of WNT10A impairs adipogenesis by inhibiting the differentiation of preadipocytes [41]. Conversely, during the late stage of differentiation, DNMT1 inhibition decreases methylation at the sterol regulatory element-binding protein 1c (Srebp-1c) promoter, leading to upregulation of SREBP-1C (a pro-lipogenic transcription factor), which enhances lipid synthesis and adipocyte maturation [42,43,44]. Intriguingly, DNMT3A also exhibits tissue-specific roles in adipose tissue. DNMT3A acts as an epigenetic suppressor of insulin sensitivity by methylating the promoter of fibroblast growth factor 21 (Fgf21), thereby reducing its expression and promoting IR [45,46]. However, liver-specific knockdown of DNMT3A in mice attenuates fibroblast growth factor 19 (FGF19)-mediated DNA methylation at the fatty acid synthase (Fasn) promoter and weakens the inhibition of hepatic lipogenesis [47]. Furthermore, the impact of DNMT3A on adipocyte differentiation is linked to the epithelial–mesenchymal transition (EMT) process. Experiments have shown that DNMT3A deficiency or dysfunction leads to the enrichment of EMT-related genes in adipocytes and reduces the differentiation of adipocytes [48].
DNA methylation modulates adipogenesis by regulating transcription factors. Notably, peroxisome proliferator-activated receptor γ (PPARγ) is a key transcriptional regulator that governs adipocyte differentiation and the expression of lipid metabolism-related genes [49,50]. Treatment with the DNA demethylating agent 5′-azacytidine induces the differentiation of fibroblast NIH-3T3 cells into adipocytes by targeting zinc finger protein 423 (Zfp423). Specifically, 5′-azacytidine reduces methylation within the Zfp423 promoter, which initiates chromatin remodeling into a transcriptionally active conformation, thereby promoting Zfp423 expression [51,52]. The upregulation of Zfp423 further enhances the expression of PPARγ2 and differentiation markers, facilitating adipogenesis in NIH-3T3 cells [53].
DNA methylation plays an essential role in regulating brown and beige adipocytes. Uncoupling protein 1 (UCP1) is a mitochondrial membrane protein and is involved in adipocyte thermogenesis [54,55]. Obesogenic diets specifically regulate visceral adipose tissue-derived serine protease inhibitor (VASPIN) expression in BAT, a process associated with DNA methylation changes. VASPIN expression correlates positively with Ucp1 expression [56]. However, the specific molecular mechanism by which VASPIN regulates BAT function remains unclear. Beyond DNMT, recent studies have revealed additional roles of DNA-modifying enzymes. The DNA demethylase ten-eleven translocation 1 (TET1) overexpression impairs the thermogenic capacity of beige adipocytes and contributes to obesity development. Interestingly, this effect is largely DNA demethylase-independent. Instead, TET1 interacts with histone deacetylase 1 (HDAC1) to mediate epigenetic changes that suppress the expression of key thermogenic genes (such as Ucp1 and Ppargc1a) [57]. In summary, DNA methylation regulates multiple stages of adipogenesis, but further exploration is needed to fully elucidate the complex interactions and tissue-specific effects.
2.1.2. DNA Methylation and Obesity-Related Metabolism
DNA methylation plays a pivotal role in regulating metabolic homeostasis and energy balance. Adiponectin, an adipocyte-derived hormone, enhances metabolic homeostasis through AMP-activated protein kinase (AMPK) and stimulates fatty acid oxidation via peroxisome proliferator-activated receptor α (PPARα) [58,59]. In contrast, resistin is an adipose-derived hormone that promotes inflammation and IR, ultimately disrupting energy homeostasis [60]. It has been demonstrated that antibiotic exposure alters the gut microbiota in obese mice, resulting in reduced expression of DNMT1 and DNMT3A. These modifications increase mRNA expression of adiponectin and resistin while activating key metabolic pathways and suppressing lipid accumulation [61]. Mechanistically, DNMT1 overexpression in adipocytes induces hypermethylation of a specific region (R2) within the adiponectin promoter, thereby suppressing adiponectin expression and promoting adipogenesis [62,63]. Moreover, a high-fat diet (HFD) augments the stability of DNMT1 through a ubiquitination-mediated mechanism. Liver-specific deletion of either Dnmt1 or Dnmt3a upregulates Beta-klotho expression and fatty acid oxidation, ultimately ameliorating HFD-induced hepatic steatosis [64].
DNA methylation also directly impacts glucose metabolism through the regulation of insulin signaling. The insulin receptor (INSR) is a critical mediator of insulin-dependent glucose uptake. Hypermethylation of the Insr promoter has been shown to repress gene expression, impairing the signaling pathway and consequently disrupting glucose uptake and utilization [65,66]. Clinical investigations have demonstrated a notable inverse correlation between the levels of INSR in adipose tissue and the body mass index (BMI) [67,68]. In addition, MBPs critically modulate metabolic outcomes. Methyl-CpG binding domain protein 2 (MBD2) acts as a reader of DNA methylation and influences the transcription of genes involved in energy expenditure and storage. Consequently, Mbd2-knockout mice exhibit enhanced glucose utilization, increased metabolic flexibility, and resistance to the deleterious effects of HFD, highlighting the physiological significance of methylation-dependent transcriptional regulation [69]. These studies have identified DNA methylation markers associated with metabolic regulation. Nevertheless, emerging evidence suggests that certain methylation changes may be reversible through lifestyle interventions such as dietary modification and physical activity [70,71]. However, the underlying mechanisms mediating these effects remain poorly understood.
2.1.3. DNA Methylation in the Adipose Tissue Microenvironment
DNA methylation plays a key role in regulating pathological processes within the adipose tissue microenvironment. Recent evidence indicates that obesity-derived macrophages can induce apoptosis of glial cells by upregulating tumor necrosis factor α (TNF-α) and activating the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB)/PH domain leucine-rich repeat protein phosphatase 1 (PHLPP1) axis, thereby contributing to adipose tissue inflammation [72]. Among CD4+ T cells, the methylation levels of 79 CpG sites are strongly correlated with the amount of visceral adipose tissue, including 4 CpG sites in the promoter region of calsyntenin 1 (Clstn1) [73]. Moreover, in subcutaneous adipose tissue and circulating leukocytes, the methylation levels of fibroblast growth factor receptor-like 1 (Fgfrl1), non-SMC condensin II complex subunit H2 (Ncaph2), paroxysmal nonkinesigenic dyskinesia (Pnkd), and SMAD family member 3 (Smad3) exhibit strong and statistically significant efficiencies in distinguishing obesity from non-obese status, as well as a great correlation between both tissues [74]. In macrophages, obesity-related factors alter DNA methylation of the Pparγ1 promoter, suppressing PPARγ1 expression and contributing to the establishment of a pro-inflammatory phenotype [75].
2.2. RNA Methylation and Obesity
To date, over 170 RNA modifications have been identified across all classes of RNA molecules [76]. Among these, RNA methylation has emerged as a fundamental post-transcriptional regulatory mechanism, critically involved in diverse aspects of RNA metabolism, including splicing, stability, nuclear export, localization, and translation efficiency [77,78]. Among the various types of RNA modifications, N6-methyladenosine (m6A) has been the most extensively studied [79,80,81]. Additionally, modifications like N1-methyladenosine (m1A), 5-methylcytosine (m5C), and 7-methylguanosine (m7G) have also gained increasing attention in recent years. Given the central role of m6A in previous research, this review exclusively focuses on m6A modification.
The m6A modification is a dynamic and reversible co-transcriptional process involving three key components: methyltransferases, demethylases, and m6A-binding proteins [82] (Table 2). Methyltransferases act as “Writers” by catalyzing RNA methylation to generate m6A modification [83]. The m6A methyltransferase complex comprises functionally distinct components organized into two subcomplexes: the m6A-Methyltransferase-like (METTL) complex (MAC) and the m6A–METTL–associated complex (MACOM) [84,85]. More specifically, METTL3 and METTL14 are the core components of MAC [86]. METTL3 provides methyltransferase activity [87,88], while METTL14 plays a structural role in stabilizing the complex and facilitating RNA binding [89]. MACOM includes proteins such as Wilms tumor 1-associated protein (WTAP), E3 ubiquitin ligase Hakai, and vir-like m6A methyltransferase-associated protein (VIRMA/KIAA1429) [90,91]. Although MACOM lacks intrinsic catalytic activity, its components interact with the METTL3-METTL14 heterodimer to ensure the efficient and precise m6A methylation [92,93]. Demethylases, termed “Erasers,” reverse m6A modifications and thereby restore RNA to its unmethylated state [94]. Among them, the fat mass and obesity-associated protein (FTO) is a well-characterized m6A demethylase that exhibits a preference for N6,2′-O-dimethyladenosine (m6Am) located at the 5′mRNA cap structure [95,96]. Compared to m6A, m6Am contains an additional 2′-O-methyl group on the ribose moiety, and FTO demethylates m6Am with higher efficiency, which leads to decreased stability of m6Am mRNAs [97,98]. Another critical demethylase is AlkB homolog 5 (ALKBH5), which specifically catalyzes the demethylation of m6A to adenosine, demonstrating greater substrate specificity and catalytic efficiency [99]. In addition to its demethylase activity, ALKBH5 also plays a distinct role in regulating mRNA export within nuclear speckles [100]. m6A-binding proteins, known as “Readers”, interpret m6A modifications by recognizing methylated mRNAs and mediate downstream biological processes [101]. YTH domain family proteins influence the fate of modified RNAs by modulating processes such as splicing, stability, and degradation [102]. For example, YTH domain-containing family protein 1 (YTHDF1) promotes the translation of m6A-modified mRNA, whereas YTHDF2 facilitates their degradation [103]. In contrast to the degradation-promoting function of YTHDF2, insulin-like growth factor 2 mRNA-binding proteins (IGF2BP1–3) promote the stabilization of target mRNAs in an m6A-dependent manner, thereby affecting gene expression output under normal and heat shock conditions [104].
In a differential gene expression analysis comparing lean and obese individuals, WTAP and VIRMA are identified as significantly upregulated in adipose tissues. In addition to these MACOM components, other m6A regulatory proteins have been implicated in obesity. For example, the expression levels of ALKBH5 and YTHDF3 in peripheral blood mononuclear cells (PBMCs) correlate with obesity [105]. Notably, Fto mRNA expression is markedly elevated in obese individuals and shows a strong positive association with BMI and other obesity-related indicators [106,107].
2.2.1. Regulation of RNA Methylation in Adipose Tissue Development and Differentiation
m6A modification regulates adipogenesis through methyltransferases, demethylases, and m6A-binding proteins. Notably, METTL3 has been extensively linked to obesity. In male mice subjected to a long-term diet-induced obesity model, Mettl3 mRNA expression is reduced [108]. Similarly, during the differentiation of porcine intramuscular adipocytes, dynamic changes in m6A levels are primarily attributed to altered expression of METTL3 and METTL14 [109]. Functionally, METTL3 facilitates WAT beiging by enhancing the stability of thermogenic mRNAs, including Krüppel-like factor 9 (Klf9) [110]. In BAT, prostaglandin E receptor 3 (EP3) has been shown to maintain WTAP expression via activation of the protein kinase A (PKA)–extracellular signal-regulated kinase 1/2 (Erk1/2) signaling pathway. Loss of Ep3 inhibits WTAP-mediated m6A methylation, resulting in downregulation of zinc finger protein 410 (Zfp410), ultimately impeding the BAT differentiation program [111]. In parallel, demethylation pathways are involved in adipogenesis. Genes regulated by the nicotinamide adenine dinucleotide phosphate (NADP)/FTO axis are mainly involved in the modulation of white adipocyte differentiation and chromatin remodeling enzymes. The knockdown of Fto blocks NADP-enhanced adipogenesis in 3T3-L1 preadipocytes [112]. Moreover, m6A-binding proteins play a pivotal role in post-transcriptional regulation during adipose tissue remodeling. For instance, YTHDF1 enhances the translation of bone morphogenetic protein 8b (Bmp8b) mRNA by specifically binding to its 3′ untranslated region (3′UTR), thereby facilitating inguinal WAT beiging and alleviating obesity [113].
These three categories of proteins can act in concert to regulate adipogenesis, especially through the FTO-YTHDF2 axis. At the cellular cycle level, FTO promotes cyclin-dependent kinase 2 (CDK2) and cyclin A2 (CCNA2) expression via its m6A demethylation activity, thus driving cell cycle progression [114]. Conversely, YTHDF2 recognizes and promotes the decay of methylated Cdk2 and Ccna2 mRNAs, blocking the mitotic clonal expansion and inhibiting adipogenesis [115,116]. In terms of cellular differentiation, silencing of Fto leads to enhanced methylation of Janus kinase 2 (Jak2) mRNA, which in turn promotes YTHDF2-mediated degradation of Jak2 transcripts. This results in the inactivation of JAK2–signal transducer and activator of transcription 3 (STAT3) signaling and subsequent downregulation of CCAAT/enhancer-binding protein β (C/EBPβ), ultimately inhibiting adipogenesis [117]. Notably, this regulatory axis also influences autophagy. Upon Fto silencing, YTHDF2 targets highly methylated autophagy-related 5 (Atg5) and autophagy-related 7 (Atg7) mRNA, causing their degradation and reduced protein expression, thereby impairing autophagic activity and further suppressing adipogenesis [118].
In addition to the aforementioned m6A modification, modifications of m5C on mRNAs have been identified in adipogenesis-related studies. NOP2/Sun domain family, member 2 (NSUN2) catalyzes the m5C modification of cyclin-dependent kinase inhibitor 1a (Cdkn1a) mRNA. Aly/REF export factor (ALYREF) is an m5C reader protein that binds to the modified Cdkn1a and facilitates its transportation from the nucleus to the cytoplasm. This enhancement in cytoplasmic localization facilitates increased translation efficiency of Cdkn1a, leading to increased protein expression, which subsequently suppresses the cell cycle and reduces adipogenesis [119]. Similarly, ALYREF recognizes and exports Y-box binding protein 2 (Ybx2) and smoothened, frizzled class receptor (Smo) mRNAs with m5C modifications, resulting in elevated protein expression of YBX2 and SMO, which inhibit adipogenesis and promote myogenesis, respectively [120]. Moreover, recent evidence indicates that YBX1 regulates the stability of m5C-modified mRNAs involved in autophagy and ubiquitination pathways, such as unc-51-like kinase 2 (Ulk2) and Ulk1, thus enhancing autophagy and adipogenesis [121,122]. Collectively, these findings suggest that RNA methylation regulates adipogenesis by orchestrating key biological processes such as cell cycle progression, cellular differentiation, and autophagy.
2.2.2. RNA Methylation and Obesity-Related Metabolism
The m6A modification plays a vital role in the regulation of lipid metabolism. Recent studies have demonstrated that mice with liver-specific knockout of Mettl3 exhibit significant hepatic lipid accumulation and elevated serum total cholesterol levels after four weeks. Gene set enrichment analysis further reveals abnormal activation of lipid metabolism pathways [123], indicating that METTL3 functions as a key regulator of lipid homeostasis. Specifically, METTL3 overexpression increases the methylated level of Fasn mRNA, subsequently facilitating fatty acid metabolism [124]. In addition, METTL3 directly binds to Rubicon mRNA and mediates its m6A modification, which in turn modulates lipid metabolism by regulating autophagic activity [125].
The role of m6A modification in glucose metabolism is indispensable. Several studies have demonstrated that m6A modification influences the stability and translation of key glycolytic enzyme mRNAs to modulate glycolytic activity [126,127]. In beige adipocytes, METTL3 and IGF2BP2 cooperatively enhance the stability of glycolysis-related mRNAs, promoting glucose uptake and metabolic activity. This process also increases lactate production, which in turn facilitates preadipocyte proliferation [128]. Moreover, m6A modification intersects with key metabolic signaling pathways. Leptin activates the JAK-STAT signaling pathway in the hypothalamus to regulate insulin sensitivity [129]. However, overexpression of FTO in liver cells impairs the leptin-induced phosphorylation process of STAT3, resulting in the disruption of glucose metabolism [130]. Conversely, ALKBH5 enhances the stability of glucagon receptor (Gcgr) mRNA through its demethylase activity, leading to elevated blood glucose levels [131]. Mechanistically, GCGR, located on the surface of hepatocytes, binds to glucagon and promotes hepatic gluconeogenesis by promoting cyclic adenosine monophosphate (cAMP)-PKA-dependent activation of the cAMP response element-binding protein (CREB)-CREB-binding protein (CBP)-CREB-regulated transcription coactivator 2 (CRTC2) complex [132,133]. Taken together, these findings highlight that m6A modification exerts a crucial role in energy metabolism through the regulation of key metabolic genes and signaling pathways.
Table 2.
The functions of m6A-related protein.
Table 2.
The functions of m6A-related protein.
| Key Components | Enzymes Included | Functions | Target RNAs | Obesity-Related Functions | References |
|---|---|---|---|---|---|
| Methyltransferases (Writer) | METTL3, METTL14, etc. | Catalyze the methylation of specific adenosines in RNA | Fasn, Rubicon, Klf9, glycolysis-related mRNAs | Regulate lipid metabolism, promote WAT beiging, modulate autophagy, enhance glycolysis and glucose uptake | [108,109,110,111,124,125,128] |
| Demethylases (Eraser) | FTO, ALKBH5, etc. | Removal of m6A methylation modification on RNA | Gcgr, Jak2, Atg5, Atg7, Cdk2, Ccna2 | Modulate gluconeogenesis, autophagy, insulin sensitivity, cell cycle, adipocyte differentiation | [112,114,117,118,130,131] |
| m6A-binding proteins (Reader) | YTH domain family proteins, IGF2BP1–3, etc. | Recognizes and binds RNAs with m6A and participates in the regulation of RNA metabolism | Bmp8b, Jak2, Atg5, Atg7, Cdk2, Ccna2, glycolytic enzyme mRNAs | Promote adipogenesis, regulate cell cycle, glucose metabolism, RNA stability, thermogenesis in adipose tissues | [113,115,116,117,118,128] |
The columns represent the following: Key Components, functional categories of m6A regulators (writers, erasers, readers); Enzymes Included, representative enzymes in each category; Functions, main roles in RNA methylation; Target RNAs, example transcripts affected by these enzymes; Obesity-related Functions, related roles in adipose tissue biology and obesity; References, citation number corresponding to each study, as listed in the reference section. Abbreviations: Fasn, fatty acid synthase; Klf9, Krüppel-like factor 9; Gcgr, glucagon receptor; Jak2, janus kinase 2; Atg5, autophagy-related 5; Cdk2, cyclin-dependent kinase 2; Ccna2, cyclin A2.
Although direct involvement of m1A modification in adipocyte development and metabolic regulation has not yet been established, it is broadly distributed across various types of RNA. Its enrichment on tRNAs and the impact they exert on tRNA structural dynamics suggest a possible role in modulating adiposity-related biological processes [85,134,135]. Supporting this possibility, hepatic transcriptomic m1A levels were found to be elevated in a mouse model of non-alcoholic steatohepatitis (NASH), accompanied by increased expression of YTH family reader proteins [136]. Additionally, the m1A demethylase ALKBH3 has been shown to enhance glycolysis in cancer cells by modulating the expression of m1A-modified ATP synthase subunit delta (Atp5d) mRNA [137]. In addition, NOP2/Sun RNA methyltransferase family member 3 (NSUN3)-dependent RNA modifications—m5C and its derivative 5-formylcytosine (f5C)—enhance mitochondrial mRNA translation [138]. When mitochondrial m5C levels are reduced, oxidative phosphorylation declines, and glycolysis is upregulated to compensate for the energy deficit [139]. In another mechanism, ALYREF stabilizes pyruvate kinase M2 (Pkm2) by binding to its m5C sites in the 3′UTR, leading to enhanced glycolysis [140]. These findings imply that RNA modifications like m1A and m5C regulate energy metabolism, though their roles in obesity and metabolic diseases remain to be clarified.
2.2.3. RNA Methylation in the Adipose Tissue Microenvironment
Beyond adipocytes, m6A modification also plays pivotal roles in other cell types within the adipose tissue microenvironment. Emerging studies demonstrate that myeloid lineage–restricted deletion of METTL3 prevents obesity in mice with improved inflammatory and metabolic phenotypes. At the mechanistic level, loss of METTL3 results in a marked upregulation of DNA damage inducible transcript 4 (Ddit4) mRNA, a key regulator of metabolic adaptation in macrophages. Elevated DDIT4 expression inhibits mammalian target of rapamycin (mTOR) signaling and modulates effector functions of macrophages, promoting an anti-inflammatory phenotype [141]. Consistently, IGF2BP2 switches M1 macrophages to M2 activation by targeting tuberous sclerosis 1 via an m6A-dependent manner [142]. Furthermore, FTO-related m6A modification regulates microglia-induced inflammatory responses by stabilizing a disintegrin and metalloprotease 17 (Adam17) mRNA expression [143]. These findings provide new insights into the multicellular regulatory mechanisms of m6A modification within the adipose tissue microenvironment.
2.3. Histone Modifications and Obesity
The histone family consists of H1, H2A, H2B, H3, and H4. Among them, two copies each of the core histones—H2A, H2B, H3, and H4—assemble to form the nucleosome core particle [144,145]. In contrast, H1 binds to the DNA entry and exit sites on the nucleosome, thereby contributing to the stabilization of higher-order chromatin structure [146,147]. Histone modifications refer to biochemical processes in which histone-modifying enzymes add or remove chemical groups to specific amino acid residues on histone and non-histone proteins [148]. These modifications can influence gene expression in two main ways: by altering chromatin structure and by recruiting regulatory factors [149,150]. Common types of histone modifications include phosphorylation, acetylation, SUMOylation, methylation, and ubiquitination [151]. While each histone modification operates through a distinct mechanism, they also exhibit interdependence and collectively participate in the regulation of multiple biological processes [152,153].
Obesity can alter adipose histone modifications, affecting adipocyte development and function [154]. Numerous studies have established a strong association between histone modifications and obesity, as well as their involvement in intergenerational epigenetic inheritance. Compared with the control group, the histone methylation level (H3K4me3) of lipid metabolism genes is dramatically elevated in obese Caenorhabditis elegans [155]. Similarly, in mammals, obese mice exhibit significant differences in histone modifications compared to normal-weight counterparts, including alterations in arginine methylation and lysine methylation [156]. Furthermore, histone modifications are a potential mechanism for intergenerational and transgenerational epigenetic effects [157]. In HFD-induced obese mice, altered levels of histone methylation and acetylation have been observed in germ cells of both sexes [158]. Further studies have identified that maternal mice fed an HFD alter epigenetic marks (H3K27me3 and H3K27ac) of numerous genes in the bone tissue of their offspring [159].
2.3.1. Regulation of Histone Modifications in Adipose Tissue Development and Differentiation
Histone modifications interact with adipogenic transcription factors to establish an adipogenic regulatory network. C/EBPβ is expressed during the early stages of adipogenesis and participates in the induction of PPARγ [160]. Histone-modifying enzymes can regulate C/EBPβ expression through diverse mechanisms. Silent mating-type information regulation 2 homolog 1 (SIRT1) suppresses the acetylation of C/EBPβ at lysine 39, as well as histone H3 at lysines 27 (H3K27) and 9 (H3K9), resulting in impaired transcription activity of C/EBPβ [161]. In contrast, protein arginine N-methyltransferase 1 (PRMT1) reduces the expression of Smad ubiquitination regulatory factor 2 (SMURF2), preventing the ubiquitination and degradation of C/EBPβ [162]. At later stages of differentiation, PPARγ and CCAAT/enhancer binding protein α (C/EBPα) are co-expressed and act synergistically to activate numerous key metabolic adipocyte genes [163]. However, this synergistic effect is subject to modulation by histone modifications. The knockdown of nuclear receptor-binding SET domain 2 (Nsd2) increases trimethylation of H3K27 (H3K27me3) levels in adipocytes. This elevation of H3K27me3 significantly inhibits C/EBPα and other targets of PPARγ, thus hindering adipogenesis [164]. In addition, phosphorylation of histone H2B at serine 36 (H2B-Ser36p), an AMPK-mediated epigenetic mark, promotes adipocyte precursor differentiation, as evidenced by increased H2B-Ser36p levels during the peak expression of proadipogenic genes [165].
Histone modifications play a critical role in thermogenic adipocyte differentiation and thermogenesis. During the differentiation process of BAT adipocytes, lysine methyltransferase PR domain-containing protein 9 (PRDM9) expression is markedly upregulated [166]. PRDM9 catalyzes the trimethylation of lysine residues 4 and 36 on histone H3 (H3K4me3 and H3K36me3) via its PR/SET domain [167]. Suppression of PRDM9 reduces the binding of H3K4me3 to pivotal genes (such as Ppargc1b, Prdm16, Cidea, and Ucp1), which impedes the differentiation of BAT adipocytes [166]. Additionally, through interaction with the transcription factor zinc finger protein 516 (ZFP516), lysine-specific demethylase 1 (LSD1) is recruited to BAT-enriched genes, where it removes methyl groups from H3K9, thus initiating the BAT program and thermogenesis [168]. Furthermore, the histone methyltransferase suppressor of variegation 4-20 homolog 2 (SUV420H2) is essential for brown and beige adipocyte development. It catalyzes trimethylation of lysine 20 on histone H4 (H4K20me3) at the promoter of 4E-binding protein 1 (4e-bp1), repressing its expression and thereby increasing peroxisome proliferator-activated receptor-γ coactivator 1α (PGC1α) protein levels, which enhances the thermogenic program [169]. These findings underscore the intricate interplay between histone modifications and transcription factors in regulating adipogenesis and thermogenesis (Figure 1).
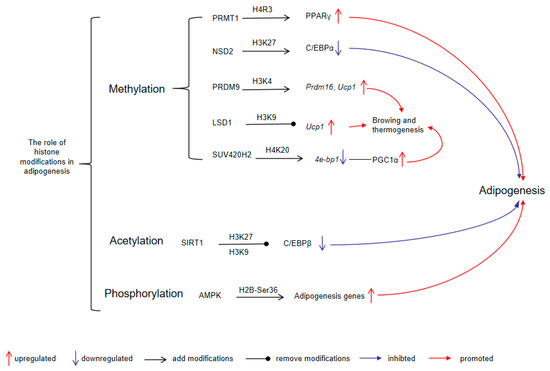
Figure 1.
The role of histone modifications in adipogenesis. PRMT1-mediated H4R3 methylation promotes PPARγ expression and adipogenesis; NSD2-mediated H3K27 methylation suppresses C/EBPα expression and inhibits adipogenesis; PRDM9 methylates H3K4 to activate Prdm16 and Ucp1, thereby enhancing browning and thermogenesis; LSD1, a demethylase, removes H3K9 methylation, facilitating Ucp1 expression; SUV420H2 catalyzes H4K20 methylation, suppressing 4e-bp1 and thereby activating PGC1α, contributing to thermogenic activation and adipogenesis; SIRT1, by deacetylating H3K27 and H3K9, represses C/EBPβ, negatively regulating adipogenesis; AMPK phosphorylates H2B at Ser36, resulting in increased transcription of adipogenesis-related genes. Abbreviations: PRMT1, protein arginine N-methyltransferase 1; NSD2, nuclear receptor-binding SET domain 2; PRDM9, PR domain-containing protein 9; LSD1, lysine-specific demethylase 1; SUV420H2, suppressor of variegation 4-20 homolog 2; SIRT1, silent mating-type information regulation 2 homolog 1; AMPK, AMP-activated protein kinase; PPARγ, peroxisome proliferator-activated receptor γ; C/EBPα, CCAAT/enhancer binding protein α; Prdm16, PR domain-containing protein 16; Ucp1, uncoupling protein 1; 4e-bp1, 4E-binding protein 1; PGC1α, peroxisome proliferator-activated receptor-γ coactivator 1α; C/EBPβ, CCAAT/enhancer-binding protein β.
2.3.2. Histone Modifications and Obesity-Related Metabolism
Histone modifications are an indispensable regulatory mechanism in energy metabolism, particularly through methylation, acetylation, phosphorylation, and ubiquitination. Concerning methylation, jumonji domain-containing protein 3 (JMJD3), as a histone demethylase, can demethylate H3K27me3. Upon fasting-activated PKA signaling, JMJD3, together with SIRT1 and PPARα, epigenetically activates β-oxidation genes to maintain energy balance [170]. Another JmjC domain-containing demethylase, jumonji histone demethylase 2a (JHDM2A), directly binds to the PPAR responsive element (PPRE) of the Ucp1 gene. It is noteworthy that β-Adrenergic activation-induced binding of Jhdm2a to the PPRE of the Ucp1 gene not only reduces the dimethylation of H3K9 (H3K9me2) at the PPRE but also facilitates the recruitment of transcription factors (Rxrα and Pparγ) and their co-activators (SRC1, CBP/P300, and PGC-1α) to the PPRE, thus enhancing Ucp1 expression and regulating fatty acid oxidation and thermogenesis in lipid metabolism [171]. In the context of acetylation, male absence on the first (MOF) acts as a lysine acetyltransferase to participate in the acetylation of lysine 16 on histone H4 (H4K16ac) [172]. MOF-mediated H4K16ac regulates glucose uptake and lipid storage in adipocytes by controlling the expression of Pparg and its entire downstream transcriptional network [173]. Moreover, deletion of HDAC11 activates the adiponectin-adiponectin receptor (AdipoR)-AMPK pathway in the liver, which effectively alleviates obesity-related metabolic diseases [174]. Phosphorylation of histones also contributes to metabolic regulation. Carbohydrate response element binding protein (ChREBP) binds to the carbohydrate response element (ChoRE) to upregulate FASN expression through histone acetylation, methylation, and phosphorylation of histone H3 at serine 10 (H3S10). In contrast, protein phosphatase 2A (PP2A) counteracts PKA by dephosphorylating histone H3 serine 28 phosphorylation (H3S28ph), thereby repressing transcription and reducing the expression of gluconeogenic genes. With regard to ubiquitination, ring finger protein 20 (RNF20), an E3 ligase critical for the monoubiquitination of histone H2B at lysine 120 (H2Bub), is essential for adipose tissue homeostasis, as its adipocyte-specific deletion in mice results in progressive fat loss, organomegaly, and hyperinsulinemia [175]. Mechanistically, Rnf20 knockdown in adipocytes reduces H3K4me3 occupancy at the solute carrier family 2 member 4 (Slc2a4) gene locus, thus suppressing glucose transporter type 4 (GLUT4) expression [176].
In addition to these classical modifications, emerging modifications such as histone lactylation and palmitoylation have recently garnered increasing attention. Histone lactylation, derived from intracellular lactate [177], has been shown to impact various metabolic processes. The genome-wide localization of histone H3 lysine 18 lactylation (H3K18la) was investigated in a broad panel of in vitro and in vivo samples, revealing that histone lactylation is present in tissues representing diverse metabolic states, with H3K18la marking active promoters [178]. Notably, Lactate-induced histone H4 lysine 12 lactylation (H4K12la) enhances the transcriptional level of peptidylglycine α-amidating monooxygenase (Pam), thereby promoting the synthesis of α-melanocyte-stimulating hormone (α-MSH), which in turn suppresses appetite and improves metabolic homeostasis [179]. Similarly, recent findings have revealed a regulatory role of palmitoylation in metabolic processes. The zinc finger DHHC-type palmitoyltransferase 23 (ZDHHC23) catalyzes the palmitoylation of plant homeodomain finger protein 2 (PHF2), thereby facilitating its ubiquitin-dependent degradation. Since PHF2 directly destabilizes SREBP-1C, this modification ultimately suppresses SREBP-1C-dependent lipogenesis [180]. Although palmitoylation has been identified on histone H3 and H4 [181,182], its specific role in adipocyte-related epigenetic regulation remains to be investigated.
2.3.3. Histone Modifications in the Adipose Tissue Microenvironment
Histone modifications regulate the adipose tissue microenvironment through multiple mechanisms. The acute treatment of hepatocytes with lipopolysaccharide (LPS) induces increased histone acetylation at H3K9 and lysine 18 (H3K9ac and H3K18ac) at the loci of inflammatory genes such as Tnf-α and chemokine (C-C motif) ligand 2 (Ccl2) [183]. Moreover, obese children with IR exhibit reduced expression levels of both SIRT1 and SIRT2 in PBMCs [184]. In addition, the loss of HDAC6 alters the composition of the gut microbiota, characterized by an increase in Bacteroides and Parabacteroides and a decrease in the S24-7 family and Lactobacillus. These alterations may exacerbate obesity by impairing the functionality of regulatory T cells [185]. Overall, histone modifications shape the inflammatory and metabolic landscape of obese adipose tissue.
2.4. Additional Epigenetic Mechanisms Implicated in Obesity
2.4.1. Non-Coding RNAs
Non-coding RNAs (ncRNAs) are RNA molecules that do not code for proteins, but instead play vital regulatory roles in gene expression [186,187,188]. Among them, microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs) have emerged as key players in the regulation of adipose tissue development. In morbidly obese adolescents, at least ten circulating miRNAs show strong associations with BMI, waist-to-height ratio (WHtR), and adipokine levels [189]; among them, miR-122 and miR-34a are upregulated in children with obesity accompanied by non-alcoholic fatty liver disease (NAFLD) and/or IR [190], and miR-486 is implicated in accelerating preadipocyte proliferation [191]. Furthermore, miR-27 has been shown to inhibit adipocyte differentiation by targeting PPARγ [192], miR-375 prevents HFD–induced obesity by targeting the aryl hydrocarbon receptor and the bacterial tryptophanase gene [193], whereas miR-22 drives thermogenic function in BAT by regulating glycolysis and thermogenic genes via the hypoxia-inducible factor 1α (HIF1α) and mechanistic target of rapamycin complex 1 (mTORC1) signaling pathways [194]. CircRNAs modulate gene expression indirectly by acting as miRNA sponges. CircSAMD4A regulates preadipocyte differentiation by acting as a miR-138-5p sponge, and thus increasing enhancer of zeste homolog 2 (EZH2) expression [195]. EZH2, a histone methyltransferase targeting H3K27, directly represses Wnt genes to promote adipogenesis [196]. In contrast, the knockdown of circRNA CDR1as promotes osteogenesis and suppresses adipogenesis of bone marrow-derived mesenchymal stem cells (BMSCs) via the circCDR1as-miR-7-5p-WNT family member 5B (WNT5B) axis [197]. In addition, several lncRNAs exert profound regulatory effects on adipogenesis and thermogenesis. MSTRG4710 promotes adipogenesis by upregulating PPARγ and C/EBPα expression, accompanied by an increased number of lipid droplets [198]. Through transcriptome assembly and meta-analysis of public datasets, several lncRNAs specifically expressed in BAT and WAT have been identified [199,200]. Among these, zinc finger and BTB domain-containing protein 7b (ZBTB7B), a transcriptional factor, is essential for the differentiation of brown and beige adipocytes in vitro. Functionally, ZBTB7B promotes thermogenic gene expression by recruiting the heterogeneous nuclear ribonucleoprotein U (hnRNPU)/Blnc1 ribonucleoprotein complex, thereby enhancing pathways related to fuel oxidation and thermogenesis [201]. Another lncRNA, XIST, is expressed at significantly higher levels in female than in male adipose tissues; its knockdown impairs the differentiation of brown preadipocytes, at least in part by disrupting its interaction with C/EBPα [202]. Moreover, lncRNA βFaar inhibits lipid droplet swelling by binding to Ras-related protein Rab-18 (RAB18) and promoting interferon regulatory factor 4 (IRF4) nuclear translocation, increases Ucp1 transcription, and further induces inguinal WAT browning by binding to karyopherin subunit alpha 6 (KPNA6) [203]. Together, these findings highlight the intricate involvement of ncRNAs in orchestrating adipocyte fate decisions through epigenetic and post-transcriptional regulation.
Growing evidence indicates that ncRNAs play crucial regulatory roles in energy metabolism [204,205]. Numerous studies have identified distinct miRNAs associated with metabolic regulation and NAFLD pathogenesis, including hsa-MIR-6236 linked to insulin sensitivity [206], miR-32-5p and miR-339-3p correlated with adipose tissue macrophage signatures [207], and miR-122-5p, miR-1343-5p, miR-193a-5p, miR-193b-5p, and miR-7845-5p positively associated with key histological features of NAFLD [208]. Among these, miR-802 inhibits hepatic AMPK expression by binding to the 3′UTR of mouse protein kinase, AMP-activated, beta 1 non-catalytic subunit (Prkab1) or human protein kinase AMP-activated catalytic subunit alpha 1 (PRKAA1), disrupting metabolic homeostasis [209], while miR-690 regulates macrophage inflammatory responses and insulin signaling through targeting nicotinamide adenine dinucleotide kinase (Nadk) [210]. Furthermore, circRNF111 sponges miR-143-3p, activating the insulin-like growth factor 2 receptor (IGF2R)-mediated insulin signaling pathway (IRS1–PI3K–AKT) and ultimately promoting glucose uptake while inhibiting lipid synthesis [211]. LncRNAs are also involved in metabolic regulation. A negative correlation between lncRNA HOTAIR expression in subcutaneous fat depots from the arm or abdomen and regional adiposity has been observed in individuals with severe obesity or uremia [212]. Conversely, in hepatic tissue, HOTAIR is significantly upregulated in patients with type 2 diabetes mellitus and mice fed with an HFD [213]. These findings suggest that ncRNAs may serve as potential therapeutic targets for metabolic diseases such as obesity, although their regulatory mechanisms require further investigation in a tissue- and context-specific manner.
2.4.2. Chromatin Remodeling
Chromatin remodeling represents a critical epigenetic mechanism that regulates gene expression by altering the structure and accessibility of chromatin to transcription factors [214]. ATP-dependent chromatin remodeling complexes, such as switch/sucrose nonfermentable (SWI/SNF) and inositol requiring 80 (INO80), alter nucleosome positioning to regulate numerous DNA-templated processes [215]. Notably, chromatin modification and remodeling genes are markedly enriched in tumor tissues of obese patients with prostate cancer compared to those from individuals with a healthy weight [216]. This observation suggests that chromatin dynamics may be influenced by obesity-related metabolic alterations in disease contexts. Consistently, chromatin remodeling complexes are also known to play essential roles in adipocyte differentiation and metabolic regulation. Dynamic changes in chromatin accessibility during forskolin (FSK)-induced cAMP stimulation induce adipocyte browning [217]. Among chromatin remodeling factors, BRG1/BRM-associated factor 60b (BAF60b) has been identified as a key chromatin-remodeling factor that modulates hepatic lipid metabolism and the progression of NAFLD by interacting with C/EBPβ to regulate PPARγ transcription [218]. In addition, myeloid-specific BAF60a knockout promotes adipose tissue macrophage pro-inflammatory activation, exacerbating diet-induced obesity, IR, and metabolic dysfunction [219].
The proliferative expansion of β-cells correlates with weight gain and the extent of IR in HFD-induced models [220,221]. Numerous chromatin remodeling complexes play critical roles in maintaining β-cell function in vivo. In particular, β-cell–specific inactivation of the SWI/SNF complex in mice impaired the ability of the pancreatic transcription factor pancreatic and duodenal homeobox 1 (Pdx1) to bind the insulin gene enhancer, thereby reducing insulin gene expression and ultimately disrupting whole-body glucose homeostasis [222]. Similarly, Chromodomain helicase DNA-binding protein 4 (CHD4), a subunit of the nucleosome remodeling and deacetylase (NuRD) complex, is required in mature β-cells, as its loss leads to glucose intolerance and impaired insulin secretion [223]. Further investigation is needed to clarify the roles of chromatin remodeling in obesity and metabolic regulation, especially its interactions with other epigenetic mechanisms.
2.4.3. Post-Translational Modifications
Post-translational modifications (PTMs) such as phosphorylation, ubiquitination, acetylation, and lactylation critically influence adipocyte differentiation by modulating transcription factor activity. Among these, acetylation and phosphorylation regulate essential signaling cascades and transcriptional programs involved in adipogenesis [224,225]. PR domain-containing protein 16 (PRDM16) acetylation at lysine 915 disrupts its interaction with PPARγ, thereby inhibiting WAT browning [226]. HDAC6-mediated deacetylation of cell death-inducing dffa-like effector c (CIDEC) reduces its stability and impairs lipid droplet fusion [227]. Increased acetylation of SREBP-1c at lysines 289 and 309 by CBP and E1A binding protein p300 (p300) during hepatic lipogenesis upregulates lipogenic genes [228]. Phosphorylation of PPARγ at serine 273 promotes IR by upregulating growth differentiation factor 3 (GDF3) in adipose tissue macrophages (ATMs) and skeletal muscle [229]. Furthermore, phosphorylation of AMPKα at serine 496 impairs AMPK-mitochondrial fission factor (MFF) signaling and disrupts mitochondrial dynamics [230]. SUMOylation, the covalent attachment of small ubiquitin-like modifier (SUMO) proteins, modulates adipogenic gene expression networks. Ubiquitin-specific protease 1 (USP1) promotes adipogenesis by deubiquitinating and stabilizing C/EBPβ [231]. Likewise, the knockdown of SUMO-specific protease 1 (Senp1) reduces the expression of key adipogenic regulators, which impairs the migratory and proliferative capacities of human adipose-derived stem cells, and ultimately inhibits adipocyte differentiation [232,233]. In contrast, SUMOylated C/EBPβ recruits death domain-associated protein (DAXX) to repress homeobox C10 (HOXC10), thereby promoting adipose tissue browning [234]. Additionally, Iroquois homeobox 3 (Irx3) acts as an upstream regulator of SUMOylation, and its repression of SUMOylation is identified as a key mediator of its pro-adipogenic effects [235]. Beyond these direct effects, PTMs also interact with other epigenetic mechanisms to influence chromatin structure and transcriptional regulation in obesity. Notably, SUMOylation may contribute to the stable silencing of pre-adipocyte-specific genes in mature adipocytes by promoting the recruitment or activation of chromatin repressors, such as polycomb group complexes and DNMTs [236,237,238].
PTMs are significant in regulating energy metabolism by modifying key enzymes or proteins [239]. In mouse hepatocytes, increased acetylation of phosphoenolpyruvate carboxykinase (PEPCK) blocks its proteasomal degradation when accompanied by decreased ubiquitination, thus enhancing hepatic gluconeogenesis [240]. Conversely, under normal conditions, acetylation of PEPCK promotes its interaction with E3 ubiquitin ligases, leading to proteasome-dependent degradation and suppression of glucose production [241]. Phosphorylated succinyl-CoA synthetase (SUCLA2) in ATMs is negatively associated with obesity in humans, suggesting a regulatory role in metabolic balance. Phosphorylation also modulates glucose and lipid metabolism. AMPK and forkhead box O1 (FOXO1) phosphorylation suppresses hepatic gluconeogenesis by downregulating key enzymes such as PEPCK and glucose-6-phosphatase (G6PASE) [242], while mTOR-mediated phosphorylation of CRTC2 relieves its inhibition of coat protein complex II (COPII)-dependent SREBP1 maturation to modulate hepatic lipid metabolism [243]. SUMOylation plays a parallel role in metabolic regulation. Previous studies have shown that cytokine stimulation induces endoplasmic reticulum stress, accompanied by alterations in the SUMOylation profile in mouse and human pancreatic β-cells [244]. The loss of SUMOylation at lysine 76 of endoplasmic reticulum protein 44 (ERP44) enhances its degradation and disrupts binding to endoplasmic reticulum oxidoreductase 1 alpha (ERO1A), protecting mice from HFD-induced metabolic disorders [245]. Additionally, SUMO-specific protease 2 (SENP2)-mediated deSUMOylation of PPARα promotes its degradation, suppressing FGF21 expression and fatty acid oxidation [246]. Lactylation, a newly recognized PTM, is elevated in skeletal muscle of obese females and correlates with IR [247]. In the liver, lactylation at lysine 673 of FASN suppresses its enzymatic activity, thereby reducing hepatic lipid accumulation via mitochondrial pyruvate carrier 1 (MPC1)-mediated pathways [248]. Collectively, these diverse PTMs finely regulate adipogenesis and metabolic homeostasis.
3. Conclusions and Future Perspectives
Recent advances in epigenetic research have significantly deepened our understanding of obesity pathogenesis. Specifically, epigenetic modifications regulate obesity-associated gene expression by altering chromatin accessibility, recruiting transcriptional regulators, and shaping complex regulatory networks, ultimately influencing adipocyte differentiation and metabolic homeostasis. About DNA methylation, studies have not only pinpointed the CpG sites associated with obesity but also clarified the regulatory mechanisms of DNMTs from multiple dimensions. Regarding RNA methylation, m6A indicates the role of various key enzymes and related proteins in the obesity process. In histone modifications, several key enzymes have been identified as regulators of lipid metabolic reprogramming and insulin signal transduction. In addition, non-coding RNAs, chromatin remodeling complexes, and post-translational modifications act as crucial epigenetic regulators in obesity by modulating adipocyte differentiation, metabolic pathways, and gene expression through distinct layers of epigenetic regulation.
Epigenetic mechanisms interact in various ways to establish a complex regulatory network, exerting multidimensional control over cellular metabolism. During the BAT-myocyte remodeling process, the histone demethylase ubiquitously transcribed tetratricopeptide repeat, X chromosome-encoded (UTX) reduces H3K27me3 levels in the promoter of Prdm16, thus promoting its high expression. PRDM16 subsequently recruits DNMT1 to the myogenic differentiation 1 (Myod1) promoter, inducing hypermethylation of the Myod1 promoter and inhibiting Myod1 expression. This process supports the maintenance of the brown adipocyte character and prevents its transition into myocytes [249]. Additionally, in the DNMT1-mediated METTL16/m6A-YTHDC2/SCD1 axis, DNA and RNA methylation cooperate to regulate lipid metabolism [250]. Moreover, RNA methylation influences the function of lncRNAs in adipogenesis; METTL14 enhances the m6A modification of LINC00278, facilitating its interaction with brahma-related gene 1 (BRG1) and subsequent activation of the PPARγ2 pathway [251]. Furthermore, RNA modification and protein phosphorylation jointly contribute to an integrated regulatory axis that governs metabolic homeostasis and tumor progression, as exemplified by the ALKBH5–GCGR/mTORC1, FTO–PPARγ/AMPK, and METTL3-large tumor suppressor kinase 1 (LATS1)/Hippo signaling pathways [131,252,253]. Despite these advances, the interactions between epigenetic mechanisms in obesity and related metabolic diseases are still not clear and require more research to provide a basis for clinical treatment and prevention.
Interestingly, epigenetic mechanisms not only affect adipogenesis and its associated metabolism but also influence the maintenance of weight loss through epigenetic memory. Current studies suggest that obesity-induced alterations in histone modifications (H3K27me3, H3K4me3) and chromatin accessibility in adipocytes do not fully revert to normal levels after weight loss. When adipocytes are re-exposed to an obesogenic environment, these persistent epigenetic marks facilitate the reactivation of genes involved in inflammation and metabolic dysregulation [254], which is a key factor in weight regain. In addition, the transgenerational epigenetics of obesity has garnered increasing attention. Maternal obesity can induce epigenetic modifications like DNA methylation in oocytes, which are transmitted to subsequent generations and influence the risk of metabolic disorders in offspring [255]. Nevertheless, the underlying mechanisms of these effects remain unclear.
Research on the epigenetics of obesity still has limitations. (1) Most animal experiments only focus on a single tissue, such as the liver or adipose tissue, lacking a synchronous epigenetic analysis across multiple tissues; (2) future research should aim to delineate the heterogeneity of epigenetic modifications within and across metabolic tissues; (3) the synergistic or antagonistic effects among epigenetic modifications in the regulation of obesity occurrence remain unclear and require further exploration; (4) integrating longitudinal human cohort studies with multi-omics approaches is essential for elucidating the role of epigenetic alterations in obesity; and (5) translating these epigenetic findings into clinical applications poses significant challenges.
Author Contributions
Conceptualization, S.Y. and Z.M.; validation, S.Y. and Z.M.; writing—original draft preparation, W.W. and S.Y.; review and editing, W.W., S.Y., S.G., F.G., Z.C., and Z.M.; supervision, S.Y. and Z.M.; project administration, S.Y.; funding acquisition, S.Y., S.G., and Z.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32400980 to S.Y.), Science and Technology Project of Zhejiang Provincial Department of Water Resources (RC2242 to Z.C.), and Zhejiang Provincial Natural Science Foundation of China (LQ22H260003 to S.G.).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Abbreviation | Full Name |
| 4e-bp1 | 4E-binding protein 1 |
| 5mC | 5-methylcytosine |
| ALYREF | Aly/REF export factor |
| AMPK | AMP-activated protein kinase |
| Atg5 | Autophagy-related 5 |
| Bmp8b | Bone morphogenetic protein 8b |
| C/EBPα | CCAAT/enhancer binding protein α |
| CCNA2 | Cyclin A2 |
| CDK2 | Cyclin-dependent kinase 2 |
| Cdkn1a | Cyclin-Dependent Kinase Inhibitor 1A |
| DMPs | Differentially methylated positions |
| DNMTs | DNA methyltransferases |
| EMT | Epithelial–mesenchymal transition |
| EP3 | Prostaglandin E receptor 3 |
| FTO | Fat mass and obesity-associated |
| Fasn | Fatty acid synthase |
| FGF19 | Fibroblast growth factor 19 |
| H4K16ac | Acetylation of lysine 16 on histone H4 |
| H3K9me2 | Dimethylation of histone H3 at lysine 9 |
| HDAC1 | Histone deacetylase 1 |
| IGF2BPs | Insulin-like growth factor 2 mRNA-binding proteins |
| INSR | Insulin receptor |
| Jak2 | Janus kinase 2 |
| JMJD3 | Jumonji domain-containing protein 3 |
| JHDM2A | Jumonji histone demethylase 2a |
| Klf9 | Krüppel-like factor 9 |
| LSD1 | Lysine-specific demethylase 1 |
| m6A | N6-methyladenosine |
| MOF | Male absence on the first |
| METTL | Methyltransferase-like |
| MBPs | Methyl-binding proteins |
| Myod1 | Myogenic differentiation 1 |
| NADP | Nicotinamide adenine dinucleotide phosphate |
| NSUN2 | NOP2/Sun domain family, member 2 |
| Nsd2 | Nuclear receptor-binding SET domain 2 |
| PPARγ | Peroxisome proliferator-activated receptor γ |
| PGC1α | Peroxisome proliferator-activated receptor-γ coactivator 1α |
| Pik3r1 | Phosphoinositide-3-kinase regulatory subunit 1 |
| Prdm16 | PR domain-containing protein 16 |
| PRMT1 | Protein arginine N-methyltransferase 1 |
| PKA | Protein kinase A |
| Srebp-1c | Sterol regulatory element-binding protein 1c |
| SIRT1 | Silent mating-type information regulation 2 homolog 1 |
| STAT3 | Sgnal transducer and activator of transcription 3 |
| SMURF2 | Smad ubiquitination regulatory factor 2 |
| SUV420H2 | Suppressor of variegation 4-20 homolog 2 |
| TNF-α | Tumor necrosis factor α |
| TET1 | Ten-eleven translocation 1 |
| UTX | Ubiquitously transcribed tetratricopeptide repeat, X chromosome-encoded |
| UCP1 | Uncoupling protein 1 |
| VASPIN | Visceral adipose tissue-derived serine protease inhibitor |
| VIRMA/KIAA1429 | Vir-like m6A methyltransferase associated |
| WAT | White adipose tissue |
| WTAP | Wilms tumor 1-associated protein |
| Wnt10a | Wingless-type MMTV integration site family, member 10a |
| YTHDF1 | YTH domain-containing family protein 1 |
| Zfp410 | Zinc finger protein 410 |
References
- Phelps, N.H.; Singleton, R.K.; Zhou, B.; Heap, R.A.; Mishra, A.; Bennett, J.E.; Paciorek, C.J.; Lhoste, V.P.; Carrillo-Larco, R.M.; Stevens, G.A.; et al. Worldwide Trends in Underweight and Obesity from 1990 to 2022: A Pooled Analysis of 3663 Population-Representative Studies with 222 Million Children, Adolescents, and Adults. Lancet 2024, 403, 1027–1050. [Google Scholar] [CrossRef] [PubMed]
- Pi-Sunyer, X. The Medical Risks of Obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef]
- Mahmoud, A.M. An Overview of Epigenetics in Obesity: The Role of Lifestyle and Therapeutic Interventions. Int. J. Mol. Sci. 2022, 23, 1341. [Google Scholar] [CrossRef]
- Lu, S.; Zhao, H.; Zhou, Y.; Xu, F. Curcumin Affects Leptin-Induced Expression of Methionine Adenosyltransferase 2A in Hepatic Stellate Cells by Inhibition of JNK Signaling. Pharmacology 2021, 106, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.-S. Obesity and Its Metabolic Complications: The Role of Adipokines and the Relationship between Obesity, Inflammation, Insulin Resistance, Dyslipidemia and Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Dragoo, J.L.; Shapiro, S.A.; Bradsell, H.; Frank, R.M. The Essential Roles of Human Adipose Tissue: Metabolic, Thermoregulatory, Cellular, and Paracrine Effects. J. Cartil. Jt. Preserv. 2021, 1, 100023. [Google Scholar] [CrossRef]
- Sakaguchi, M. Adipose Tissue Dynamics, Thermogenesis, and Interorgan Connections for Preventing Obesity and Metabolic Disorders. JMA J. 2024, 7, 172–177. [Google Scholar] [CrossRef]
- Shang, Y.; Wang, X.; Su, S.; Ji, F.; Shao, D.; Duan, C.; Chen, T.; Liang, C.; Zhang, D.; Lu, H. Identifying of Immune-Associated Genes for Assessing the Obesity-Associated Risk to the Offspring in Maternal Obesity: A Bioinformatics and Machine Learning. CNS Neurosci. Ther. 2024, 30, e14700. [Google Scholar] [CrossRef]
- Górczyńska-Kosiorz, S.; Kosiorz, M.; Dzięgielewska-Gęsiak, S. Exploring the Interplay of Genetics and Nutrition in the Rising Epidemic of Obesity and Metabolic Diseases. Nutrients 2024, 16, 3562. [Google Scholar] [CrossRef]
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An Operational Definition of Epigenetics. Genes Dev. 2009, 23, 781–783. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, K.; Guan, K.; Khan, F.A.; Pandupuspitasari, N.S.; Negara, W.; Sun, F.; Huang, C. Future in the Past: Paternal Reprogramming of Offspring Phenotype and the Epigenetic Mechanisms. Arch. Toxicol. 2024, 98, 1685–1703. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Mao, C.; Liu, S.; Tao, Y.; Xiao, D. Epigenetic Modifications in Obesity-Associated Diseases. MedComm 2024, 5, e496. [Google Scholar] [CrossRef]
- Vohl, M.-C.; Malagón, M.M.; Ramos-Molina, B. Editorial: Dietary Factors, Epigenetics and Their Implications for Human Obesity. Front. Endocrinol. 2020, 11, 601. [Google Scholar] [CrossRef]
- Park, Y.J.; Han, S.M.; Huh, J.Y.; Kim, J.B. Emerging Roles of Epigenetic Regulation in Obesity and Metabolic Disease. J. Biol. Chem. 2021, 297, 101296. [Google Scholar] [CrossRef]
- Izquierdo, A.G.; Crujeiras, A.B. Obesity-Related Epigenetic Changes after Bariatric Surgery. Front. Endocrinol. 2019, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Aghaei-Zarch, S.M. From Molecular Pathogenesis to Therapy: Unraveling Non-Coding RNAs/DNMT3A Axis in Human Cancers. Biochem. Pharmacol. 2024, 222, 116107. [Google Scholar] [CrossRef]
- Bird, A. DNA Methylation Patterns and Epigenetic Memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef]
- Wang, C.; Wang, M.; Ma, J. Analysis of Genome-Wide DNA Methylation Patterns in Obesity. Endocr. J. 2021, 68, 1439–1453. [Google Scholar] [CrossRef]
- McAllan, L.; Baranasic, D.; Villicaña, S.; Brown, S.; Zhang, W.; Lehne, B.; Adamo, M.; Jenkinson, A.; Elkalaawy, M.; Mohammadi, B.; et al. Integrative Genomic Analyses in Adipocytes Implicate DNA Methylation in Human Obesity and Diabetes. Nat. Commun. 2023, 14, 2784. [Google Scholar] [CrossRef]
- Nicoletti, C.F.; Cortes-Oliveira, C.; Noronha, N.Y.; Pinhel, M.A.S.; Dantas, W.S.; Jácome, A.; Marchini, J.S.; Gualano, B.; Crujeiras, A.B.; Nonino, C.B. DNA Methylation Pattern Changes Following a Short-Term Hypocaloric Diet in Women with Obesity. Eur. J. Clin. Nutr. 2020, 74, 1345–1353. [Google Scholar] [CrossRef]
- Pinhel, M.A.S.; Noronha, N.Y.; Nicoletti, C.F.; Pereira, V.A.; de Oliveira, B.A.; Cortes-Oliveira, C.; Salgado, W.; Barbosa, F.; Marchini, J.S.; Souza, D.R.; et al. Changes in DNA Methylation and Gene Expression of Insulin and Obesity-Related Gene PIK3R1 after Roux-En-Y Gastric Bypass. Int. J. Mol. Sci. 2020, 21, 4476. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.; Krassowska, A.; Gilbert, N.; Chevassut, T.; Forrester, L.; Ansell, J.; Ramsahoye, B. Severe Global DNA Hypomethylation Blocks Differentiation and Induces Histone Hyperacetylation in Embryonic Stem Cells. Mol. Cell. Biol. 2004, 24, 8862–8871. [Google Scholar] [CrossRef]
- Yagi, M.; Kabata, M.; Tanaka, A.; Ukai, T.; Ohta, S.; Nakabayashi, K.; Shimizu, M.; Hata, K.; Meissner, A.; Yamamoto, T.; et al. Identification of Distinct Loci for de Novo DNA Methylation by DNMT3A and DNMT3B during Mammalian Development. Nat. Commun. 2020, 11, 3199. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-F.; Li, B.-Z.; Li, Z.; Liu, P.; Wang, Y.; Tang, Q.; Ding, J.; Jia, Y.; Chen, Z.; Li, L.; et al. Tet-Mediated Formation of 5-Carboxylcytosine and Its Excision by TDG in Mammalian DNA. Science 2011, 333, 1303–1307. [Google Scholar] [CrossRef]
- Mahmood, N.; Rabbani, S.A. DNA Methylation Readers and Cancer: Mechanistic and Therapeutic Applications. Front. Oncol. 2019, 9, 489. [Google Scholar] [CrossRef] [PubMed]
- Kamei, Y.; Suganami, T.; Ehara, T.; Kanai, S.; Hayashi, K.; Yamamoto, Y.; Miura, S.; Ezaki, O.; Okano, M.; Ogawa, Y. Increased Expression of DNA Methyltransferase 3a in Obese Adipose Tissue: Studies with Transgenic Mice. Obesity 2010, 18, 314–321. [Google Scholar] [CrossRef]
- Wu, F.-Y.; Yin, R.-X. Recent Progress in Epigenetics of Obesity. Diabetol. Metab. Syndr. 2022, 14, 171. [Google Scholar] [CrossRef]
- Bruggeman, E.C.; Garretson, J.T.; Wu, R.; Shi, H.; Xue, B. Neuronal Dnmt1 Deficiency Attenuates Diet-Induced Obesity in Mice. Endocrinology 2018, 159, 145–162. [Google Scholar] [CrossRef]
- Shinozaki, H.; Kawai, S.; Gamo-Kawasaki, M.; Takei, A.; Tsujikado, K.; Fukuda, K.; Yamauchi, M.; Hara, K.; Tsuchiya, T.; Takebayashi, K.; et al. Analysis of Serum Levels and DNA Methylation of Fibroblast Growth Factor 21 Using Peripheral Blood-Derived Genomes in Patients with Obesity. Endocr. J. 2024, 71, 907–924. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Vilarrubla, A.; Mas-Parés, B.; Carreras-Badosa, G.; Bonmatí-Santané, A.; Martínez-Calcerrada, J.-M.; Niubó-Pallàs, M.; de Zegher, F.; Ibáñez, L.; López-Bermejo, A.; Bassols, J. DNA Methylation Signatures in Paired Placenta and Umbilical Cord Samples: Relationship with Maternal Pregestational Body Mass Index and Offspring Metabolic Outcomes. Biomedicines 2024, 12, 301. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Huang, P.; Huang, X.; Zhang, L.; Liu, L.; Xiang, W.; Liu, L.; He, X. Alterations of DNA Methylation Profile in Peripheral Blood of Children with Simple Obesity. Health Inf. Sci. Syst. 2024, 12, 26. [Google Scholar] [CrossRef]
- Sehgal, R.; Perfilyev, A.; Männistö, V.; Ågren, J.; Nilsson, E.; Käkelä, P.; Ling, C.; de Mello, V.D.; Pihlajamäki, J. Liver Saturated Fat Content Associates with Hepatic DNA Methylation in Obese Individuals. Clin. Epigenetics 2023, 15, 21. [Google Scholar] [CrossRef]
- Kvaløy, K.; Page, C.M.; Holmen, T.L. Epigenome-Wide Methylation Differences in a Group of Lean and Obese Women—A HUNT Study. Sci. Rep. 2018, 8, 16330. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Berg, A.; Imamura Kawasawa, Y.; Bixler, E.O.; Fernandez-Mendoza, J.; Whitsel, E.A.; Liao, D. Association between DNA Methylation in Obesity-Related Genes and Body Mass Index Percentile in Adolescents. Sci. Rep. 2019, 9, 2079. [Google Scholar] [CrossRef]
- Salas-Pérez, F.; Ramos-Lopez, O.; Mansego, M.L.; Milagro, F.I.; Santos, J.L.; Riezu-Boj, J.I.; Martínez, J.A. DNA Methylation in Genes of Longevity-Regulating Pathways: Association with Obesity and Metabolic Complications. Aging 2019, 11, 1874–1899. [Google Scholar] [CrossRef]
- Keyhan, S.; Burke, E.; Schrott, R.; Huang, Z.; Grenier, C.; Price, T.; Raburn, D.; Corcoran, D.L.; Soubry, A.; Hoyo, C.; et al. Male Obesity Impacts DNA Methylation Reprogramming in Sperm. Clin. Epigenetics 2021, 13, 17. [Google Scholar] [CrossRef]
- Rushing, A.; Sommer, E.C.; Zhao, S.; Po’e, E.K.; Barkin, S.L. Salivary Epigenetic Biomarkers as Predictors of Emerging Childhood Obesity. BMC Med. Genet. 2020, 21, 34. [Google Scholar] [CrossRef]
- Nishida, H.; Onishi, K.; Kurose, S.; Tsutsumi, H.; Miyauchi, T.; Takao, N.; Yoshiuchi, S.; Fujii, A.; Kimura, Y. Changes in Body Composition and FTO Whole Blood DNA Methylation among Japanese Women: A Randomized Clinical Trial of Weight-Loss Program. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 2157–2167. [Google Scholar] [CrossRef]
- Jing, J.; Li, F.; Zha, L.; Yang, X.; Wu, R.; Wang, S.; Xue, B.; Shi, H. The Histone Methyltransferase Suv39h Regulates 3T3-L1 Adipogenesis. Adipocyte 2020, 9, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, R.; Shan, W.; Yu, L.; Xue, B.; Shi, H. DNA Methylation Biphasically Regulates 3T3-L1 Preadipocyte Differentiation. Mol. Endocrinol. 2016, 30, 677–687. [Google Scholar] [CrossRef]
- Ju, U.-I.; Jeong, D.-W.; Seo, J.; Park, J.B.; Park, J.-W.; Suh, K.-S.; Kim, J.B.; Chun, Y.-S. Neddylation of Sterol Regulatory Element-Binding Protein 1c Is a Potential Therapeutic Target for Nonalcoholic Fatty Liver Treatment. Cell Death Dis. 2020, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, L.; Pandak, W.M.; Heuman, D.; Hylemon, P.B.; Ren, S. High Glucose Induces Lipid Accumulation via 25-Hydroxycholesterol DNA-CpG Methylation. iScience 2020, 23, 101102. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Tsujimoto, K.; Hashimoto, K.; Kawahori, K.; Hanzawa, N.; Hamaguchi, M.; Seki, T.; Nawa, M.; Ehara, T.; Kitamura, Y.; et al. Epigenetic Modulation of Fgf21 in the Perinatal Mouse Liver Ameliorates Diet-Induced Obesity in Adulthood. Nat. Commun. 2018, 9, 636. [Google Scholar] [CrossRef]
- Xiao, M.; Tang, Y.; Wang, S.; Wang, J.; Wang, J.; Guo, Y.; Zhang, J.; Gu, J. The Role of Fibroblast Growth Factor 21 in Diabetic Cardiovascular Complications and Related Epigenetic Mechanisms. Front. Endocrinol. 2021, 12, 598008. [Google Scholar] [CrossRef]
- Kim, Y.-C.; Seok, S.; Zhang, Y.; Ma, J.; Kong, B.; Guo, G.; Kemper, B.; Kemper, J.K. Intestinal FGF15/19 Physiologically Repress Hepatic Lipogenesis in the Late Fed-State by Activating SHP and DNMT3A. Nat. Commun. 2020, 11, 5969. [Google Scholar] [CrossRef]
- Tovy, A.; Reyes, J.M.; Zhang, L.; Huang, Y.-H.; Rosas, C.; Daquinag, A.C.; Guzman, A.; Ramabadran, R.; Chen, C.-W.; Gu, T.; et al. Constitutive Loss of DNMT3A Causes Morbid Obesity through Misregulation of Adipogenesis. eLife 2022, 11, e72359. [Google Scholar] [CrossRef]
- Ma, L.; Dai, X.; Wu, C.; Li, M.; Sheng, H.; Mao, W. Tanyu Tongzhi Formula Delays Atherosclerotic Plaque Progression by Promoting Alternative Macrophage Activation via PPARγ and AKT/ERK Signal Pathway in ApoE Knock-out Mice. Front. Pharmacol. 2021, 12, 734589. [Google Scholar] [CrossRef]
- Wu, H.; Li, X.; Shen, C. Peroxisome Proliferator-Activated Receptor γ in White and Brown Adipocyte Regulation and Differentiation. Physiol. Res. 2020, 69, 759–773. [Google Scholar] [CrossRef]
- Longo, M.; Raciti, G.A.; Zatterale, F.; Parrillo, L.; Desiderio, A.; Spinelli, R.; Hammarstedt, A.; Hedjazifar, S.; Hoffmann, J.M.; Nigro, C.; et al. Epigenetic Modifications of the Zfp/ZNF423 Gene Control Murine Adipogenic Commitment and Are Dysregulated in Human Hypertrophic Obesity. Diabetologia 2018, 61, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-L.; Kang, S. Rewriting Cellular Fate: Epigenetic Interventions in Obesity and Cellular Programming. Mol. Med. Camb. Mass 2024, 30, 169. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Schmidt, H.; Lai, B.; Ge, K. Transcriptional and Epigenomic Regulation of Adipogenesis. Mol. Cell. Biol. 2019, 39, e00601-18. [Google Scholar] [CrossRef]
- Ricquier, D. Uncoupling Protein 1 of Brown Adipocytes, the Only Uncoupler: A Historical Perspective. Front. Endocrinol. 2011, 2, 85. [Google Scholar] [CrossRef]
- Liang, X.; Guan, H.; Sun, J.; Qi, Y.; Yao, W. Comparative Proteomic Analysis of tPVAT during Ang II Infusion. Biomedicines 2021, 9, 1820. [Google Scholar] [CrossRef]
- Weiner, J.; Rohde, K.; Krause, K.; Zieger, K.; Klöting, N.; Kralisch, S.; Kovacs, P.; Stumvoll, M.; Blüher, M.; Böttcher, Y.; et al. Brown Adipose Tissue (BAT) Specific Vaspin Expression Is Increased after Obesogenic Diets and Cold Exposure and Linked to Acute Changes in DNA-Methylation. Mol. Metab. 2017, 6, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Damal Villivalam, S.; You, D.; Kim, J.; Lim, H.W.; Xiao, H.; Zushin, P.-J.H.; Oguri, Y.; Amin, P.; Kang, S. TET1 Is a Beige Adipocyte-Selective Epigenetic Suppressor of Thermogenesis. Nat. Commun. 2020, 11, 4313. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New Insight into Adiponectin Role in Obesity and Obesity-Related Diseases. Biomed Res. Int. 2014, 2014, 658913. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef]
- Benomar, Y.; Gertler, A.; De Lacy, P.; Crépin, D.; Ould Hamouda, H.; Riffault, L.; Taouis, M. Central Resistin Overexposure Induces Insulin Resistance through Toll-like Receptor 4. Diabetes 2013, 62, 102–114. [Google Scholar] [CrossRef]
- Yao, H.; Fan, C.; Lu, Y.; Fan, X.; Xia, L.; Li, P.; Wang, R.; Tang, T.; Wang, Y.; Qi, K. Alteration of Gut Microbiota Affects Expression of Adiponectin and Resistin through Modifying DNA Methylation in High-Fat Diet-Induced Obese Mice. Genes Nutr. 2020, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.Y.; Park, Y.J.; Pan, X.; Shin, K.C.; Kwak, S.-H.; Bassas, A.F.; Sallam, R.M.; Park, K.S.; Alfadda, A.A.; Xu, A.; et al. Obesity-Induced DNA Hypermethylation of the Adiponectin Gene Mediates Insulin Resistance. Nat. Commun. 2015, 6, 7585. [Google Scholar] [CrossRef]
- Un Nisa, K.; Reza, M.I. Key Relevance of Epigenetic Programming of Adiponectin Gene in Pathogenesis of Metabolic Disorders. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 506–517. [Google Scholar] [CrossRef]
- Wang, S.; Zha, L.; Cui, X.; Yeh, Y.-T.; Liu, R.; Jing, J.; Shi, H.; Chen, W.; Hanover, J.; Yin, J.; et al. Epigenetic Regulation of Hepatic Lipid Metabolism by DNA Methylation. Adv. Sci. Weinh. Baden-Wurtt. Ger. 2023, 10, e2206068. [Google Scholar] [CrossRef]
- Zhong, X.; Jin, F.; Huang, C.; Du, M.; Gao, M.; Wei, X. DNA Methylation of AMHRII and INSR Gene Is Associated with the Pathogenesis of Polycystic Ovary Syndrome (PCOS). Technol. Health Care 2021, 29, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Payankaulam, S.; Raicu, A.-M.; Arnosti, D.N. Transcriptional Regulation of INSR, the Insulin Receptor Gene. Genes 2019, 10, 984. [Google Scholar] [CrossRef]
- Kaminska, D.; Hämäläinen, M.; Cederberg, H.; Käkelä, P.; Venesmaa, S.; Miettinen, P.; Ilves, I.; Herzig, K.-H.; Kolehmainen, M.; Karhunen, L.; et al. Adipose Tissue INSR Splicing in Humans Associates with Fasting Insulin Level and Is Regulated by Weight Loss. Diabetologia 2014, 57, 347–351. [Google Scholar] [CrossRef]
- Calcaterra, V.; Zuccotti, G.; Mari, A.; Iafusco, F.; Maione, G.; Iafusco, D.; Tinto, N. Can Obesity Exacerbate Hyperinsulinaemia in the Presence of the Mutation of an Insulin Receptor Gene? Clin. Obes. 2023, 13, e12619. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Song, J.; He, X.; Zhang, M.; Hu, S.; Zhang, S.; Yu, Q.; Yang, P.; Xiong, F.; Wang, D.W.; et al. Loss of Mbd2 Protects Mice against High-Fat Diet-Induced Obesity and Insulin Resistance by Regulating the Homeostasis of Energy Storage and Expenditure. Diabetes 2016, 65, 3384–3395. [Google Scholar] [CrossRef]
- Mirza, I.; Mohamed, A.; Deen, H.; Balaji, S.; Elsabbahi, D.; Munasser, A.; Naquiallah, D.; Abdulbaseer, U.; Hassan, C.; Masrur, M.; et al. Obesity-Associated Vitamin D Deficiency Correlates with Adipose Tissue DNA Hypomethylation, Inflammation, and Vascular Dysfunction. Int. J. Mol. Sci. 2022, 23, 14377. [Google Scholar] [CrossRef]
- Fitzgerald, K.N.; Hodges, R.; Hanes, D.; Stack, E.; Cheishvili, D.; Szyf, M.; Henkel, J.; Twedt, M.W.; Giannopoulou, D.; Herdell, J.; et al. Potential Reversal of Epigenetic Age Using a Diet and Lifestyle Intervention: A Pilot Randomized Clinical Trial. Aging 2021, 13, 9419–9432. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Lu, Z.; Jiang, X.; Zhang, S.; Tian, X.; Wang, Q. Obesity-Derived Macrophages Upregulate TNF-α to Induce Apoptosis in Glial Cell via the NF-κB/PHLPP1 Axis. Int. Immunopharmacol. 2024, 141, 112962. [Google Scholar] [CrossRef]
- Hohos, N.M.; Smith, A.K.; Kilaru, V.; Park, H.J.; Hausman, D.B.; Bailey, L.B.; Lewis, R.D.; Phillips, B.G.; Meagher, R.B. CD4+ and CD8+ T-Cell-Specific DNA Cytosine Methylation Differences Associated with Obesity. Obesity 2018, 26, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Crujeiras, A.B.; Diaz-Lagares, A.; Sandoval, J.; Milagro, F.I.; Navas-Carretero, S.; Carreira, M.C.; Gomez, A.; Hervas, D.; Monteiro, M.P.; Casanueva, F.F.; et al. DNA Methylation Map in Circulating Leukocytes Mirrors Subcutaneous Adipose Tissue Methylation Pattern: A Genome-Wide Analysis from Non-Obese and Obese Patients. Sci. Rep. 2017, 7, 41903. [Google Scholar] [CrossRef]
- Wang, X.; Cao, Q.; Yu, L.; Shi, H.; Xue, B.; Shi, H. Epigenetic Regulation of Macrophage Polarization and Inflammation by DNA Methylation in Obesity. JCI Insight 2016, 1, e87748. [Google Scholar] [CrossRef] [PubMed]
- Cappannini, A.; Ray, A.; Purta, E.; Mukherjee, S.; Boccaletto, P.; Moafinejad, S.N.; Lechner, A.; Barchet, C.; Klaholz, B.P.; Stefaniak, F.; et al. MODOMICS: A Database of RNA Modifications and Related Information. 2023 Update. Nucleic Acids Res. 2024, 52, D239–D244. [Google Scholar] [CrossRef]
- Han, B.; Xi, W.; Hong, Y.; Gu, L.; Chao, Y.; Li, L.; Liu, C.; Yang, L.; Chao, J.; Yao, H. Mutual Regulation of Noncoding RNAs and RNA Modifications in Psychopathology: Potential Therapeutic Targets for Psychiatric Disorders? Pharmacol. Ther. 2022, 237, 108254. [Google Scholar] [CrossRef]
- Wilkinson, E.; Cui, Y.-H.; He, Y.-Y. Roles of RNA Modifications in Diverse Cellular Functions. Front. Cell Dev. Biol. 2022, 10, 828683. [Google Scholar] [CrossRef]
- Zhang, B.; Hao, Y.; Liu, H.; Wu, J.; Lu, L.; Wang, X.; Bajpai, A.K.; Yang, X. Interplay of RNA m6A Modification-Related Geneset in Pan-Cancer. Biomedicines 2024, 12, 2211. [Google Scholar] [CrossRef]
- Bai, Y.; Chang, D.; Ren, H.; Ju, M.; Wang, Y.; Chen, B.; Li, H.; Liu, X.; Li, D.; Huo, X.; et al. Engagement of N6-Methyladenisine Methylation of Gng4 mRNA in Astrocyte Dysfunction Regulated by CircHECW2. Acta Pharm. Sin. B 2024, 14, 1644–1660. [Google Scholar] [CrossRef]
- Liu, H.; Gu, J.; Jin, Y.; Yuan, Q.; Ma, G.; Du, M.; Ge, Y.; Qin, C.; Lv, Q.; Fu, G.; et al. Genetic Variants in N6-Methyladenosine Are Associated with Bladder Cancer Risk in the Chinese Population. Arch. Toxicol. 2021, 95, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hu, Y.; Zhou, B.; Bao, Y.; Li, Z.; Gong, C.; Yang, H.; Wang, S.; Xiao, Y. The Role of m6A Modification in Physiology and Disease. Cell Death Dis. 2020, 11, 960. [Google Scholar] [CrossRef]
- Xie, L.; Zhang, X.; Xie, J.; Xu, Y.; Li, X.-J.; Lin, L. Emerging Roles for DNA 6mA and RNA m6A Methylation in Mammalian Genome. Int. J. Mol. Sci. 2023, 24, 13897. [Google Scholar] [CrossRef] [PubMed]
- Knuckles, P.; Lence, T.; Haussmann, I.U.; Jacob, D.; Kreim, N.; Carl, S.H.; Masiello, I.; Hares, T.; Villaseñor, R.; Hess, D.; et al. Zc3h13/Flacc Is Required for Adenosine Methylation by Bridging the mRNA-Binding Factor Rbm15/Spenito to the m6A Machinery Component Wtap/Fl(2)d. Genes Dev. 2018, 32, 415–429. [Google Scholar] [CrossRef]
- Wang, X.; Gan, M.; Wang, Y.; Wang, S.; Lei, Y.; Wang, K.; Zhang, X.; Chen, L.; Zhao, Y.; Niu, L.; et al. Comprehensive Review on Lipid Metabolism and RNA Methylation: Biological Mechanisms, Perspectives and Challenges. Int. J. Biol. Macromol. 2024, 270, 132057. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 Complex Mediates Mammalian Nuclear RNA N6-Adenosine Methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Feng, J.; Xue, Y.; Guan, Z.; Zhang, D.; Liu, Z.; Gong, Z.; Wang, Q.; Huang, J.; Tang, C.; et al. Structural Basis of N(6)-Adenosine Methylation by the METTL3-METTL14 Complex. Nature 2016, 534, 575–578. [Google Scholar] [CrossRef]
- Chen, W.-W.; Qi, J.-W.; Hang, Y.; Wu, J.-X.; Zhou, X.-X.; Chen, J.-Z.; Wang, J.; Wang, H.-H. Simvastatin Is Beneficial to Lung Cancer Progression by Inducing METTL3-Induced m6A Modification on EZH2 mRNA. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4263–4270. [Google Scholar] [CrossRef]
- Corbeski, I.; Vargas-Rosales, P.A.; Bedi, R.K.; Deng, J.; Coelho, D.; Braud, E.; Iannazzo, L.; Li, Y.; Huang, D.; Ethève-Quelquejeu, M.; et al. The Catalytic Mechanism of the RNA Methyltransferase METTL3. eLife 2024, 12, RP92537. [Google Scholar] [CrossRef]
- Su, S.; Li, S.; Deng, T.; Gao, M.; Yin, Y.; Wu, B.; Peng, C.; Liu, J.; Ma, J.; Zhang, K. Cryo-EM Structures of Human m6A Writer Complexes. Cell Res. 2022, 32, 982–994. [Google Scholar] [CrossRef]
- Peng, C.; Xiong, F.; Pu, X.; Hu, Z.; Yang, Y.; Qiao, X.; Jiang, Y.; Han, M.; Wang, D.; Li, X. m6A Methylation Modification and Immune Cell Infiltration: Implications for Targeting the Catalytic Subunit m6A-METTL Complex in Gastrointestinal Cancer Immunotherapy. Front. Immunol. 2023, 14, 1326031. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, C.; Yin, W.; Tao, M.; Niu, Z.; Cui, Y.; Wu, D.; Gao, F. From Bone Marrow Mesenchymal Stem Cells to Diseases: The Crucial Role of m6A Methylation in Orthopedics. Stem Cell Res. Ther. 2025, 16, 228. [Google Scholar] [CrossRef] [PubMed]
- Lence, T.; Paolantoni, C.; Worpenberg, L.; Roignant, J.-Y. Mechanistic Insights into m6A RNA Enzymes. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 222–229. [Google Scholar] [CrossRef]
- Berlivet, S.; Scutenaire, J.; Deragon, J.-M.; Bousquet-Antonelli, C. Readers of the m6A Epitranscriptomic Code. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 329–342. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, L.-H.; Wang, Y.; Xiao, Y.; Liu, J.; Zhang, W.; Yan, N.; Amu, G.; Tang, X.; Zhang, L.; et al. Structural Insights into FTO’s Catalytic Mechanism for the Demethylation of Multiple RNA Substrates. Proc. Natl. Acad. Sci. USA 2019, 116, 2919–2924. [Google Scholar] [CrossRef]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.-J.; Chen, Q.; et al. Reversible Methylation of m6Am in the 5′ Cap Controls mRNA Stability. Nature 2017, 541, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Du, K.; Zhang, L.; Lee, T.; Sun, T. m6A RNA Methylation Controls Neural Development and Is Involved in Human Diseases. Mol. Neurobiol. 2019, 56, 1596–1606. [Google Scholar] [CrossRef]
- Yan, S.; Wen, W.; Mo, Z.; Gu, S.; Chen, Z. Epitranscriptomic Role of m6A in Obesity-Associated Disorders and Cancer Metabolic Reprogramming. Genes 2025, 16, 498. [Google Scholar] [CrossRef]
- Kaur, S.; Tam, N.Y.; McDonough, M.A.; Schofield, C.J.; Aik, W.S. Mechanisms of Substrate Recognition and N6-Methyladenosine Demethylation Revealed by Crystal Structures of ALKBH5–RNA Complexes. Nucleic Acids Res. 2022, 50, 4148–4160. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.-M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.-L.; Song, S.-H.; et al. ALKBH5 Is a Mammalian RNA Demethylase That Impacts RNA Metabolism and Mouse Fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef]
- Zhang, W.; Qian, Y.; Jia, G. The Detection and Functions of RNA Modification m6A Based on m6A Writers and Erasers. J. Biol. Chem. 2021, 297, 100973. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zeng, C.; Liu, J.; Wang, L.; Yuan, X.; Yuan, L.; Xia, X.; Huang, W. The YTH Domain-Containing Protein Family: Emerging Players in Immunomodulation and Tumour Immunotherapy Targets. Clin. Transl. Med. 2024, 14, e1784. [Google Scholar] [CrossRef] [PubMed]
- Zaccara, S.; Jaffrey, S.R. A Unified Model for the Function of YTHDF Proteins in Regulating m6A-Modified mRNA. Cell 2020, 181, 1582–1595.e18. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-Methyladenosine by IGF2BP Proteins Enhances mRNA Stability and Translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Rønningen, T.; Dahl, M.B.; Valderhaug, T.G.; Cayir, A.; Keller, M.; Tönjes, A.; Blüher, M.; Böttcher, Y. m6A Regulators in Human Adipose Tissue—Depot-Specificity and Correlation with Obesity. Front. Endocrinol. 2021, 12, 778875. [Google Scholar] [CrossRef]
- Onalan, E.; Yakar, B.; Onalan, E.E.; Karakulak, K.; Kaymaz, T.; Donder, E. m6A RNA, FTO, ALKBH5 Expression in Type 2 Diabetic and Obesity Patients. J. Coll. Physicians Surg.--Pak. JCPSP 2022, 32, 1143–1148. [Google Scholar] [CrossRef]
- Czogała, W.; Czogała, M.; Strojny, W.; Wątor, G.; Wołkow, P.; Wójcik, M.; Bik Multanowski, M.; Tomasik, P.; Wędrychowicz, A.; Kowalczyk, W.; et al. Methylation and Expression of FTO and PLAG1 Genes in Childhood Obesity: Insight into Anthropometric Parameters and Glucose-Lipid Metabolism. Nutrients 2021, 13, 1683. [Google Scholar] [CrossRef] [PubMed]
- Croft, A.J.; Kelly, C.; Chen, D.; Haw, T.J.; Balachandran, L.; Murtha, L.A.; Boyle, A.J.; Sverdlov, A.L.; Ngo, D.T.M. Sex-Based Differences in Short- and Longer-Term Diet-Induced Metabolic Heart Disease. Am. J. Physiol. Heart Circ. Physiol. 2024, 326, H1219–H1251. [Google Scholar] [CrossRef]
- Chao, M.; Wang, M.; Han, H.; Liu, Y.; Sun, X.; Tian, T.; Pang, W.; Cai, R. Profiling of m6A Methylation in Porcine Intramuscular Adipocytes and Unravelling PHKG1 Represses Porcine Intramuscular Lipid Deposition in an m6A-Dependent Manner. Int. J. Biol. Macromol. 2024, 272, 132728. [Google Scholar] [CrossRef]
- Xie, R.; Yan, S.; Zhou, X.; Gao, Y.; Qian, Y.; Hou, J.; Chen, Z.; Lai, K.; Gao, X.; Wei, S. Activation of METTL3 Promotes White Adipose Tissue Beiging and Combats Obesity. Diabetes 2023, 72, 1083–1094. [Google Scholar] [CrossRef]
- Tao, X.; Du, R.; Guo, S.; Feng, X.; Yu, T.; OuYang, Q.; Chen, Q.; Fan, X.; Wang, X.; Guo, C.; et al. PGE2 -EP3 Axis Promotes Brown Adipose Tissue Formation through Stabilization of WTAP RNA Methyltransferase. EMBO J. 2022, 41, e110439. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, C.; Wang, N.; Li, S.; Liu, Q.; Sun, Z.; Wang, K.; Yu, S.-C.; Yang, Q. NADP Modulates RNA m6A Methylation and Adipogenesis via Enhancing FTO Activity. Nat. Chem. Biol. 2020, 16, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhou, X.; Wu, C.; Gao, Y.; Qian, Y.; Hou, J.; Xie, R.; Han, B.; Chen, Z.; Wei, S.; et al. Adipocyte YTH N(6)-Methyladenosine RNA-Binding Protein 1 Protects against Obesity by Promoting White Adipose Tissue Beiging in Male Mice. Nat. Commun. 2023, 14, 1379. [Google Scholar] [CrossRef]
- Wu, R.; Liu, Y.; Yao, Y.; Zhao, Y.; Bi, Z.; Jiang, Q.; Liu, Q.; Cai, M.; Wang, F.; Wang, Y.; et al. FTO Regulates Adipogenesis by Controlling Cell Cycle Progression via m6A-YTHDF2 Dependent Mechanism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1323–1330. [Google Scholar] [CrossRef]
- Li, J.; Xie, H.; Ying, Y.; Chen, H.; Yan, H.; He, L.; Xu, M.; Xu, X.; Liang, Z.; Liu, B.; et al. YTHDF2 Mediates the mRNA Degradation of the Tumor Suppressors to Induce AKT Phosphorylation in N6-Methyladenosine-Dependent Way in Prostate Cancer. Mol. Cancer 2020, 19, 152. [Google Scholar] [CrossRef]
- Sato, S.; Dacher, M.; Kurumizaka, H. Nucleosome Structures Built from Highly Divergent Histones: Parasites and Giant DNA Viruses. Epigenomes 2022, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Guo, G.; Bi, Z.; Liu, Y.; Zhao, Y.; Chen, N.; Wang, F.; Wang, Y.; Wang, X. m6A Methylation Modulates Adipogenesis through JAK2-STAT3-C/EBPβ Signaling. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 796–806. [Google Scholar] [CrossRef]
- Wang, X.; Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Shi, H.; Wang, F.; Wang, Y. m6A mRNA Methylation Controls Autophagy and Adipogenesis by Targeting Atg5 and Atg7. Autophagy 2020, 16, 1221–1235. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Y.; Wu, R.; Chen, Y.; Chen, W.; Liu, Y.; Luo, Y.; Huang, C.; Zeng, B.; Liao, X.; et al. mRNA m5C Controls Adipogenesis by Promoting CDKN1A mRNA Export and Translation. RNA Biol. 2021, 18, 711–721. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Y.; Wu, R.; Gao, C.-C.; Liao, X.; Han, X.; Zeng, B.; Huang, C.; Luo, Y.; Liu, Y.; et al. mRNA m5C Inhibits Adipogenesis and Promotes Myogenesis by Respectively Facilitating YBX2 and SMO mRNA Export in ALYREF-m5C Manner. Cell. Mol. Life Sci. CMLS 2022, 79, 481. [Google Scholar] [CrossRef]
- Wu, R.; Feng, S.; Li, F.; Shu, G.; Wang, L.; Gao, P.; Zhu, X.; Zhu, C.; Wang, S.; Jiang, Q. Transcriptional and Post-Transcriptional Control of Autophagy and Adipogenesis by YBX1. Cell Death Dis. 2023, 14, 29. [Google Scholar] [CrossRef]
- Wu, R.; Cao, S.; Li, F.; Feng, S.; Shu, G.; Wang, L.; Gao, P.; Zhu, X.; Zhu, C.; Wang, S.; et al. RNA-Binding Protein YBX1 Promotes Brown Adipogenesis and Thermogenesis via PINK1/PRKN-Mediated Mitophagy. FASEB J. 2022, 36, e22219. [Google Scholar] [CrossRef] [PubMed]
- Dai, G.; Huang, S.; Li, Y.; Tu, X.; Xia, J.; Zhou, Z.; Chen, W.; Zhang, A.; Lin, J.; Li, Y.; et al. Mettl3-Mediated m6A Modification Plays a Role in Lipid Metabolism Disorders and Progressive Liver Damage in Mice by Regulating Lipid Metabolism-Related Gene Expression. Aging 2023, 15, 5550–5568. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Ma, L.L.; Xu, Y.Q.; Wang, B.H.; Li, S.M. METTL3 Inhibits Hepatic Insulin Sensitivity via N6-Methyladenosine Modification of Fasn mRNA and Promoting Fatty Acid Metabolism. Biochem. Biophys. Res. Commun. 2019, 518, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Gong, Y.; Wang, X.; He, W.; Wu, L.; Zhang, L.; Xiong, L.; Huang, Y.; Su, L.; Shi, P.; et al. METTL3-m6A-Rubicon Axis Inhibits Autophagy in Nonalcoholic Fatty Liver Disease. Mol. Ther. J. Am. Soc. Gene Ther. 2022, 30, 932–946. [Google Scholar] [CrossRef]
- Yue, S.-W.; Liu, H.-L.; Su, H.-F.; Luo, C.; Liang, H.-F.; Zhang, B.-X.; Zhang, W. m6A-Regulated Tumor Glycolysis: New Advances in Epigenetics and Metabolism. Mol. Cancer 2023, 22, 137. [Google Scholar] [CrossRef]
- Liu, W.-W.; Zheng, S.-Q.; Li, T.; Fei, Y.-F.; Wang, C.; Zhang, S.; Wang, F.; Jiang, G.-M.; Wang, H. RNA Modifications in Cellular Metabolism: Implications for Metabolism-Targeted Therapy and Immunotherapy. Signal Transduct. Target. Ther. 2024, 9, 70. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Zhang, T.; Ping, X.; Wang, D.; Chen, Y.; Yu, J.; Liu, C.; Liu, Z.; Zheng, Y.; et al. Rna M6 a Methylation Regulates Glycolysis of Beige Fat and Contributes to Systemic Metabolic Homeostasis. Adv. Sci. Weinh. Baden-Wurtt. Ger. 2023, 10, e2300436. [Google Scholar] [CrossRef]
- D’souza, A.M.; Neumann, U.H.; Glavas, M.M.; Kieffer, T.J. The Glucoregulatory Actions of Leptin. Mol. Metab. 2017, 6, 1052–1065. [Google Scholar] [CrossRef]
- Bravard, A.; Vial, G.; Chauvin, M.-A.; Rouillé, Y.; Bailleul, B.; Vidal, H.; Rieusset, J. FTO Contributes to Hepatic Metabolism Regulation through Regulation of Leptin Action and STAT3 Signalling in Liver. Cell Commun. Signal. CCS 2014, 12, 1–13. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, Z.; Han, Z.; Shi, L.; Li, X.; Liu, Y.; Li, Z.; Zhao, C.; Cui, Y.; Zhou, L.; et al. Liver ALKBH5 Regulates Glucose and Lipid Homeostasis Independently through GCGR and mTORC1 Signaling. Science 2025, 387, eadp4120. [Google Scholar] [CrossRef]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of Hepatic Glucose Metabolism in Health and Disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef]
- He, L.; Sabet, A.; Djedjos, S.; Miller, R.; Sun, X.; Hussain, M.A.; Radovick, S.; Wondisford, F.E. Metformin and Insulin Suppress Hepatic Gluconeogenesis through Phosphorylation of CREB Binding Protein. Cell 2009, 137, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Monshaugen, I.; Wilson, B.; Wang, F.; Klungland, A.; Ougland, R.; Dutta, A. TRMT6/61A-Dependent Base Methylation of tRNA-Derived Fragments Regulates Gene-Silencing Activity and the Unfolded Protein Response in Bladder Cancer. Nat. Commun. 2022, 13, 2165. [Google Scholar] [CrossRef] [PubMed]
- Tomikawa, C. 7-Methylguanosine Modifications in Transfer RNA (tRNA). Int. J. Mol. Sci. 2018, 19, 4080. [Google Scholar] [CrossRef]
- Klinge, C.M.; Piell, K.M.; Petri, B.J.; He, L.; Zhang, X.; Pan, J.; Rai, S.N.; Andreeva, K.; Rouchka, E.C.; Wahlang, B.; et al. Combined Exposure to Polychlorinated Biphenyls and High-Fat Diet Modifies the Global Epitranscriptomic Landscape in Mouse Liver. Environ. Epigenetics 2021, 7, dvab008. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Z.; Xie, G.; Zhang, H.; Wang, Z.; Zhou, J.; Chen, F.; Li, J.; Chen, L.; Niu, H.; et al. RNA m1A Methylation Regulates Glycolysis of Cancer Cells through Modulating ATP5D. Proc. Natl. Acad. Sci. USA 2022, 119, e2119038119. [Google Scholar] [CrossRef]
- Delaunay, S.; Pascual, G.; Feng, B.; Klann, K.; Behm, M.; Hotz-Wagenblatt, A.; Richter, K.; Zaoui, K.; Herpel, E.; Münch, C.; et al. Mitochondrial RNA Modifications Shape Metabolic Plasticity in Metastasis. Nature 2022, 607, 593–603. [Google Scholar] [CrossRef]
- Park, K.-S.; Wiederkehr, A.; Kirkpatrick, C.; Mattenberger, Y.; Martinou, J.-C.; Marchetti, P.; Demaurex, N.; Wollheim, C.B. Selective Actions of Mitochondrial Fission/Fusion Genes on Metabolism-Secretion Coupling in Insulin-Releasing Cells. J. Biol. Chem. 2008, 283, 33347–33356. [Google Scholar] [CrossRef]
- Wang, J.-Z.; Zhu, W.; Han, J.; Yang, X.; Zhou, R.; Lu, H.-C.; Yu, H.; Yuan, W.-B.; Li, P.-C.; Tao, J.; et al. The Role of the HIF-1α/ALYREF/PKM2 Axis in Glycolysis and Tumorigenesis of Bladder Cancer. Cancer Commun. 2021, 41, 560–575. [Google Scholar] [CrossRef]
- Qin, Y.; Li, B.; Arumugam, S.; Lu, Q.; Mankash, S.M.; Li, J.; Sun, B.; Li, J.; Flavell, R.A.; Li, H.-B.; et al. m6A mRNA Methylation-Directed Myeloid Cell Activation Controls Progression of NAFLD and Obesity. Cell Rep. 2021, 37, 109968. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ji, Y.; Feng, P.; Liu, R.; Li, G.; Zheng, J.; Xue, Y.; Wei, Y.; Ji, C.; Chen, D.; et al. The m6A Reader IGF2BP2 Regulates Macrophage Phenotypic Activation and Inflammatory Diseases by Stabilizing TSC1 and PPARγ. Adv. Sci. 2021, 8, 2100209. [Google Scholar] [CrossRef]
- Chen, X.; Lai, J.; Wu, Z.; Chen, J.; Yang, B.; Chen, C.; Ding, C. Fat Mass and Obesity-Mediated N 6 -Methyladenosine Modification Modulates Neuroinflammatory Responses after Traumatic Brain Injury. Neural Regen. Res. 2024, 21, 730–741. [Google Scholar] [CrossRef]
- Harp, J.M.; Hanson, B.L.; Bunick, G.J. The Core Particle of the Nucleosome. In New Comprehensive Biochemistry; Chromatin Structure and Dynamics: State-of-the-Art; Elsevier: Amsterdam, The Netherlands, 2004; Volume 39, pp. 13–44. [Google Scholar]
- Jiang, C.; Pugh, B.F. Nucleosome Positioning and Gene Regulation: Advances through Genomics. Nat. Rev. Genet. 2009, 10, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Stillman, B. Histone Modifications: Insights into Their Influence on Gene Expression. Cell 2018, 175, 6–9. [Google Scholar] [CrossRef]
- Tian, Y.; Feng, T.; Zhang, J.; Meng, Q.; Zhan, W.; Tang, M.; Liu, C.; Li, M.; Tao, W.; Shu, Y.; et al. Histone H1 Deamidation Facilitates Chromatin Relaxation for DNA Repair. Nature 2025, 641, 779–787. [Google Scholar] [CrossRef]
- Marmorstein, R.; Trievel, R.C. Histone Modifying Enzymes: Structures, Mechanisms, and Specificities. Biochim. Biophys. Acta 2009, 1789, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Morgan, M.A.J.; Shilatifard, A. Reevaluating the Roles of Histone-Modifying Enzymes and Their Associated Chromatin Modifications in Transcriptional Regulation. Nat. Genet. 2020, 52, 1271–1281. [Google Scholar] [CrossRef]
- Allis, C.D.; Jenuwein, T. The Molecular Hallmarks of Epigenetic Control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wu, J.; Guo, H.; Yao, W.; Li, S.; Lu, Y.; Jia, Y.; Liang, X.; Tang, J.; Zhang, H. Post-translational Modifications of Histones: Mechanisms, Biological Functions, and Therapeutic Targets. MedComm 2023, 4, e292. [Google Scholar] [CrossRef]
- Castellano-Castillo, D.; Ramos-Molina, B.; Oliva-Olivera, W.; Ocaña-Wilhelmi, L.; Queipo-Ortuño, M.I.; Cardona, F. Genome Profiling of H3k4me3 Histone Modification in Human Adipose Tissue during Obesity and Insulin Resistance. Biomedicines 2021, 9, 1363. [Google Scholar] [CrossRef]
- Wan, Q.-L.; Meng, X.; Wang, C.; Dai, W.; Luo, Z.; Yin, Z.; Ju, Z.; Fu, X.; Yang, J.; Ye, Q.; et al. Histone H3K4me3 Modification Is a Transgenerational Epigenetic Signal for Lipid Metabolism in Caenorhabditis Elegans. Nat. Commun. 2022, 13, 768. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Shuai, L.; Zhu, M.; Liu, P.; Xie, Z.-F.; Jiang, S.; Jiang, H.-W.; Li, J.; Zhao, Y.; Li, J.-Y.; et al. The Landscape of Histone Modifications in a High-Fat Diet-Induced Obese (DIO) Mouse Model. Mol. Cell. Proteom. MCP 2017, 16, 1324–1334. [Google Scholar] [CrossRef]
- Mørkve Knudsen, T.; Rezwan, F.I.; Jiang, Y.; Karmaus, W.; Svanes, C.; Holloway, J.W. Transgenerational and Intergenerational Epigenetic Inheritance in Allergic Diseases. J. Allergy Clin. Immunol. 2018, 142, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.-H.; Zhu, C.-C.; Sun, S.-C. Effects of Obesity and Diabetes on the Epigenetic Modification of Mammalian Gametes. J. Cell. Physiol. 2019, 234, 7847–7855. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Caviness, P.C.; Zhao, H.; Belcher, B.; Wankhade, U.D.; Shankar, K.; Blackburn, M.L.; Lazarenko, O.P. Maternal High-Fat Diet Modifies Epigenetic Marks H3K27me3 and H3K27ac in Bone to Regulate Offspring Osteoblastogenesis in Mice. Epigenetics 2022, 17, 2209–2222. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J.; Liu, X.S.; et al. PPARγ and C/EBP Factors Orchestrate Adipocyte Biology via Adjacent Binding on a Genome-Wide Scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Liu, J.; Han, F.; Shi, J.; Wu, G.; Wang, K.; Shen, K.; Zhao, M.; Gao, X.; et al. Adiponectin Ameliorates Hypertrophic Scar by Inhibiting Yes-Associated Protein Transcription through SIRT1-Mediated Deacetylation of C/EBPβ and Histone H3. iScience 2022, 25, 105236. [Google Scholar] [CrossRef]
- Zhu, Q.; Wang, D.; Liang, F.; Tong, X.; Liang, Z.; Wang, X.; Chen, Y.; Mo, D. Protein Arginine Methyltransferase PRMT1 Promotes Adipogenesis by Modulating Transcription Factors C/EBPβ and PPARγ. J. Biol. Chem. 2022, 298, 102309. [Google Scholar] [CrossRef]
- Madsen, M.S.; Siersbæk, R.; Boergesen, M.; Nielsen, R.; Mandrup, S. Peroxisome Proliferator-Activated Receptor γ and C/EBPα Synergistically Activate Key Metabolic Adipocyte Genes by Assisted Loading. Mol. Cell. Biol. 2014, 34, 939–954. [Google Scholar] [CrossRef]
- Zhuang, L.; Jang, Y.; Park, Y.-K.; Lee, J.-E.; Jain, S.; Froimchuk, E.; Broun, A.; Liu, C.; Gavrilova, O.; Ge, K. Depletion of Nsd2-Mediated Histone H3K36 Methylation Impairs Adipose Tissue Development and Function. Nat. Commun. 2018, 9, 1796. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Camacho, C.V.; Setlem, R.; Ryu, K.W.; Parameswaran, B.; Gupta, R.K.; Kraus, W.L. Functional Interplay between Histone H2B ADP-Ribosylation and Phosphorylation Controls Adipogenesis. Mol. Cell 2020, 79, 934–949.e14. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Ye, L.; Guo, X.; Wang, W.; Zhang, Z.; Wang, Q.; Huang, J.; Xu, J.; Cai, Y.; Shou, X.; et al. Glutamine Production by Glul Promotes Thermogenic Adipocyte Differentiation through Prdm9-Mediated H3K4me3 and Transcriptional Reprogramming. Diabetes 2023, 72, 1574–1596. [Google Scholar] [CrossRef]
- Powers, N.R.; Parvanov, E.D.; Baker, C.L.; Walker, M.; Petkov, P.M.; Paigen, K. The Meiotic Recombination Activator PRDM9 Trimethylates Both H3K36 and H3K4 at Recombination Hotspots in Vivo. PLoS Genet. 2016, 12, e1006146. [Google Scholar] [CrossRef] [PubMed]
- Sambeat, A.; Gulyaeva, O.; Dempersmier, J.; Tharp, K.M.; Stahl, A.; Paul, S.M.; Sul, H.S. LSD1 Interacts with Zfp516 to Promote UCP1 Transcription and Brown Fat Program. Cell Rep. 2016, 15, 2536–2549. [Google Scholar] [CrossRef]
- Cui, X.; Cao, Q.; Li, F.; Jing, J.; Liu, Z.; Yang, X.; Schwartz, G.J.; Yu, L.; Shi, H.; Shi, H.; et al. The Histone Methyltransferase SUV420H2 Regulates Brown and Beige Adipocyte Thermogenesis. JCI Insight 2024, 9, e164771. [Google Scholar] [CrossRef]
- Seok, S.; Kim, Y.-C.; Byun, S.; Choi, S.; Xiao, Z.; Iwamori, N.; Zhang, Y.; Wang, C.; Ma, J.; Ge, K.; et al. Fasting-Induced JMJD3 Histone Demethylase Epigenetically Activates Mitochondrial Fatty Acid β-Oxidation. J. Clin. Invest. 2018, 128, 3144–3159. [Google Scholar] [CrossRef]
- Tateishi, K.; Okada, Y.; Kallin, E.M.; Zhang, Y. Role of Jhdm2a in Regulating Metabolic Gene Expression and Obesity Resistance. Nature 2009, 458, 757–761. [Google Scholar] [CrossRef]
- Rea, S.; Xouri, G.; Akhtar, A. Males Absent on the First (MOF): From Flies to Humans. Oncogene 2007, 26, 5385–5394. [Google Scholar] [CrossRef] [PubMed]
- Pessoa Rodrigues, C.; Chatterjee, A.; Wiese, M.; Stehle, T.; Szymanski, W.; Shvedunova, M.; Akhtar, A. Histone H4 Lysine 16 Acetylation Controls Central Carbon Metabolism and Diet-Induced Obesity in Mice. Nat. Commun. 2021, 12, 6212. [Google Scholar] [CrossRef]
- Sun, L.; Marin de Evsikova, C.; Bian, K.; Achille, A.; Telles, E.; Pei, H.; Seto, E. Programming and Regulation of Metabolic Homeostasis by HDAC11. EBioMedicine 2018, 33, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Tao, C.; Pan, J.; Zhang, L.; Liu, L.; Zhao, Y.; Fan, Y.; Cao, C.; Liu, J.; Zhang, J.; et al. Rnf20 Deficiency in Adipocyte Impairs Adipose Tissue Development and Thermogenesis. Protein Cell 2021, 12, 475–492. [Google Scholar] [CrossRef]
- Zhao, Y.; Liang, X.; Tang, J.; Cao, C.; Yang, C.; Yang, S.; Zhao, J.; Yuan, J.; Zhang, M.; Wang, Y. Adipocyte RNF20 Knockout Leads to Hyperinsulinemia via the H2Bub-H3K4me3-Slc2a4 Axis. J. Cell. Mol. Med. 2025, 29, e70649. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic Regulation of Gene Expression by Histone Lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Galle, E.; Wong, C.-W.; Ghosh, A.; Desgeorges, T.; Melrose, K.; Hinte, L.C.; Castellano-Castillo, D.; Engl, M.; de Sousa, J.A.; Ruiz-Ojeda, F.J.; et al. H3K18 Lactylation Marks Tissue-Specific Active Enhancers. Genome Biol. 2022, 23, 207. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wan, B.; Zhang, H.; Zhang, L.; Zhang, R.; Li, L.; Zhang, Y.; Hu, C. Histone Lactylation Mediated by Fam172a in POMC Neurons Regulates Energy Balance. Nat. Commun. 2024, 15, 10111. [Google Scholar] [CrossRef]
- Jeong, D.-W.; Park, J.-W.; Kim, K.S.; Kim, J.; Huh, J.; Seo, J.; Kim, Y.L.; Cho, J.-Y.; Lee, K.-W.; Fukuda, J.; et al. Palmitoylation-Driven PHF2 Ubiquitination Remodels Lipid Metabolism through the SREBP1c Axis in Hepatocellular Carcinoma. Nat. Commun. 2023, 14, 6370. [Google Scholar] [CrossRef]
- Wilson, J.P.; Raghavan, A.S.; Yang, Y.-Y.; Charron, G.; Hang, H.C. Proteomic Analysis of Fatty-Acylated Proteins in Mammalian Cells with Chemical Reporters Reveals S-Acylation of Histone H3 Variants. Mol. Cell. Proteom. MCP 2011, 10, M110.001198. [Google Scholar] [CrossRef]
- Thompson, H.M. Add Lipidation to Histone Modifications. Sci. Signal. 2011, 4, ec226. [Google Scholar] [CrossRef]
- Mikula, M.; Majewska, A.; Ledwon, J.K.; Dzwonek, A.; Ostrowski, J. Obesity Increases Histone H3 Lysine 9 and 18 Acetylation at Tnfa and Ccl2 Genes in Mouse Liver. Int. J. Mol. Med. 2014, 34, 1647–1654. [Google Scholar] [CrossRef] [PubMed]
- Arab Sadeghabadi, Z.; Nourbakhsh, M.; Pasalar, P.; Emamgholipour, S.; Golestani, A.; Larijani, B.; Razzaghy-Azar, M. Reduced Gene Expression of Sirtuins and Active AMPK Levels in Children and Adolescents with Obesity and Insulin Resistance. Obes. Res. Clin. Pract. 2018, 12, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Lieber, A.D.; Beier, U.H.; Xiao, H.; Wilkins, B.J.; Jiao, J.; Li, X.S.; Schugar, R.C.; Strauch, C.M.; Wang, Z.; Brown, J.M.; et al. Loss of HDAC6 Alters Gut Microbiota and Worsens Obesity. FASEB J. 2019, 33, 1098–1109. [Google Scholar] [CrossRef]
- Uppaluri, K.R.; Challa, H.J.; Gaur, A.; Jain, R.; Krishna Vardhani, K.; Geddam, A.; Natya, K.; Aswini, K.; Palasamudram, K.; K, S.M. Unlocking the Potential of Non-Coding RNAs in Cancer Research and Therapy. Transl. Oncol. 2023, 35, 101730. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, Z.; Song, M.; Huan, H.; Cai, Z.; Wu, B.; Shen, J.; Zhou, Y.L.; Shi, J. CircIQGAP1 Regulates RCAN1 and RCAN2 through the Mechanism of ceRNA and Promotes the Growth of Malignant Glioma. Pharmacol. Res. 2023, 197, 106979. [Google Scholar] [CrossRef]
- Peng, Q.; Deng, Y.; Xu, Z.; Duan, R.; Wang, W.; Wang, S.; Hong, Y.; Wang, Q.; Zhang, Y. Fat Mass and Obesity-Associated Protein Alleviates Cerebral Ischemia/Reperfusion Injury by Inhibiting Ferroptosis via miR-320-3p/SLC7A11 Axis. Biochem. Pharmacol. 2024, 230, 116603. [Google Scholar] [CrossRef]
- Al-Rawaf, H.A. Circulating microRNAs and Adipokines as Markers of Metabolic Syndrome in Adolescents with Obesity. Clin. Nutr. Edinb. Scotl. 2019, 38, 2231–2238. [Google Scholar] [CrossRef]
- Oses, M.; Margareto Sanchez, J.; Portillo, M.P.; Aguilera, C.M.; Labayen, I. Circulating miRNAs as Biomarkers of Obesity and Obesity-Associated Comorbidities in Children and Adolescents: A Systematic Review. Nutrients 2019, 11, 2890. [Google Scholar] [CrossRef]
- Cui, X.; You, L.; Zhu, L.; Wang, X.; Zhou, Y.; Li, Y.; Wen, J.; Xia, Y.; Wang, X.; Ji, C.; et al. Change in Circulating microRNA Profile of Obese Children Indicates Future Risk of Adult Diabetes. Metabolism 2018, 78, 95–105. [Google Scholar] [CrossRef]
- Karbiener, M.; Fischer, C.; Nowitsch, S.; Opriessnig, P.; Papak, C.; Ailhaud, G.; Dani, C.; Amri, E.-Z.; Scheideler, M. microRNA miR-27b Impairs Human Adipocyte Differentiation and Targets PPARgamma. Biochem. Biophys. Res. Commun. 2009, 390, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ren, Y.; Sundaram, K.; Mu, J.; Sriwastva, M.K.; Dryden, G.W.; Lei, C.; Zhang, L.; Yan, J.; Zhang, X.; et al. miR-375 Prevents High-Fat Diet-Induced Insulin Resistance and Obesity by Targeting the Aryl Hydrocarbon Receptor and Bacterial Tryptophanase (tnaA) Gene. Theranostics 2021, 11, 4061–4077. [Google Scholar] [CrossRef]
- Lou, P.; Bi, X.; Tian, Y.; Li, G.; Kang, Q.; Lv, C.; Song, Y.; Xu, J.; Sheng, X.; Yang, X.; et al. MiR-22 Modulates Brown Adipocyte Thermogenesis by Synergistically Activating the Glycolytic and mTORC1 Signaling Pathways. Theranostics 2021, 11, 3607–3623. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, H.; Li, Y.; Mao, R.; Yang, H.; Zhang, Y.; Zhang, Y.; Guo, P.; Zhan, D.; Zhang, T. Circular RNA SAMD4A Controls Adipogenesis in Obesity through the miR-138-5p/EZH2 Axis. Theranostics 2020, 10, 4705–4719. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jin, Q.; Lee, J.-E.; Su, I.-h.; Ge, K. Histone H3K27 Methyltransferase EzH2 Represses Wnt Genes to Facilitate Adipogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 7317–7322. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Q.; Li, Z.; Yang, Q.; Liu, Y.; Du, Z.; Zhang, G.; Song, Y. Circular RNA CDR1as Promotes Adipogenic and Suppresses Osteogenic Differentiation of BMSCs in Steroid-Induced Osteonecrosis of the Femoral Head. Bone 2020, 133, 115258. [Google Scholar] [CrossRef]
- Tang, T.; Jiang, G.; Shao, J.; Wang, M.; Zhang, X.; Xia, S.; Sun, W.; Jia, X.; Wang, J.; Lai, S. lncRNA MSTRG4710 Promotes the Proliferation and Differentiation of Preadipocytes through miR-29b-3p/IGF1 Axis. Int. J. Mol. Sci. 2023, 24, 15715. [Google Scholar] [CrossRef]
- Gao, W.; Liu, J.-L.; Lu, X.; Yang, Q. Epigenetic Regulation of Energy Metabolism in Obesity. J. Mol. Cell Biol. 2021, 13, 480–499. [Google Scholar] [CrossRef]
- Xiong, Y.; Yue, F.; Jia, Z.; Gao, Y.; Jin, W.; Hu, K.; Zhang, Y.; Zhu, D.; Yang, G.; Kuang, S. A Novel Brown Adipocyte-Enriched Long Non-Coding RNA That Is Required for Brown Adipocyte Differentiation and Sufficient to Drive Thermogenic Gene Program in White Adipocytes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 409–419. [Google Scholar] [CrossRef]
- Li, S.; Mi, L.; Yu, L.; Yu, Q.; Liu, T.; Wang, G.-X.; Zhao, X.-Y.; Wu, J.; Lin, J.D. Zbtb7b Engages the Long Noncoding RNA Blnc1 to Drive Brown and Beige Fat Development and Thermogenesis. Proc. Natl. Acad. Sci. USA 2017, 114, E7111–E7120. [Google Scholar] [CrossRef]
- Wu, C.; Fang, S.; Zhang, H.; Li, X.; Du, Y.; Zhang, Y.; Lin, X.; Wang, L.; Ma, X.; Xue, Y.; et al. Long Noncoding RNA XIST Regulates Brown Preadipocytes Differentiation and Combats High-Fat Diet Induced Obesity by Targeting C/EBPα. Mol. Med. Camb. Mass 2022, 28, 6. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Huang, B.; Sha, B.; Gao, D.; Qin, Y.; Li, Z.; Chen, X.; Jin, Y.; Pan, Y.; Zhang, Y.; et al. The Long Noncoding RNA ΒFaar Promotes White Adipose Tissue Browning and Prevents Diet-Induced Obesity. Adv. Sci. Weinh. Baden-Wurtt. Ger. 2025, 12, e05545. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, X.; Li, Z.; Wang, S.; Fan, J. Functions of Circular RNAs in Regulating Adipogenesis of Mesenchymal Stem Cells. Stem Cells Int. 2020, 2020, 3763069. [Google Scholar] [CrossRef]
- Zong, Y.; Wang, X.; Cui, B.; Xiong, X.; Wu, A.; Lin, C.; Zhang, Y. Decoding the Regulatory Roles of Non-Coding RNAs in Cellular Metabolism and Disease. Mol. Ther. J. Am. Soc. Gene Ther. 2023, 31, 1562–1576. [Google Scholar] [CrossRef]
- Paneru, B.D.; Chini, J.; McCright, S.J.; DeMarco, N.; Miller, J.; Joannas, L.D.; Henao-Mejia, J.; Titchenell, P.M.; Merrick, D.M.; Lim, H.-W.; et al. Myeloid-Derived miR-6236 Potentiates Adipocyte Insulin Signaling and Prevents Hyperglycemia during Obesity. Nat. Commun. 2024, 15, 5394. [Google Scholar] [CrossRef]
- Doncheva, A.I.; Romero, S.; Ramirez-Garrastacho, M.; Lee, S.; Kolnes, K.J.; Tangen, D.S.; Olsen, T.; Drevon, C.A.; Llorente, A.; Dalen, K.T.; et al. Extracellular Vesicles and microRNAs Are Altered in Response to Exercise, Insulin Sensitivity and Overweight. Acta Physiol. 2022, 236, e13862. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-J.; Baumert, B.O.; Stratakis, N.; Goodrich, J.A.; Wu, H.-T.; He, J.-X.; Zhao, Y.-Q.; Aung, M.T.; Wang, H.-X.; Eckel, S.P.; et al. Circulating microRNA Expression and Nonalcoholic Fatty Liver Disease in Adolescents with Severe Obesity. World J. Gastroenterol. 2024, 30, 332–345. [Google Scholar] [CrossRef]
- Li, B.; Xiao, Q.; Zhao, H.; Zhang, J.; Yang, C.; Zou, Y.; Zhang, B.; Liu, J.; Sun, H.; Liu, H. Schisanhenol Ameliorates Non-Alcoholic Fatty Liver Disease via Inhibiting miR-802 Activation of AMPK-Mediated Modulation of Hepatic Lipid Metabolism. Acta Pharm. Sin. B 2024, 14, 3949–3963. [Google Scholar] [CrossRef]
- Ying, W.; Gao, H.; Dos Reis, F.C.G.; Bandyopadhyay, G.; Ofrecio, J.M.; Luo, Z.; Ji, Y.; Jin, Z.; Ly, C.; Olefsky, J.M. MiR-690, an Exosomal-Derived miRNA from M2-Polarized Macrophages, Improves Insulin Sensitivity in Obese Mice. Cell Metab. 2021, 33, 781–790.e5. [Google Scholar] [CrossRef]
- Lin, X.; Du, Y.; Lu, W.; Gui, W.; Sun, S.; Zhu, Y.; Wang, G.; Eserberg, D.T.; Zheng, F.; Zhou, J.; et al. CircRNF111 Protects against Insulin Resistance and Lipid Deposition via Regulating miR-143-3p/IGF2R Axis in Metabolic Syndrome. Front. Cell Dev. Biol. 2021, 9, 663148. [Google Scholar] [CrossRef]
- Kuo, F.-C.; Huang, Y.-C.; Yen, M.-R.; Lee, C.-H.; Hsu, K.-F.; Yang, H.-Y.; Wu, L.-W.; Lu, C.-H.; Hsu, Y.-J.; Chen, P.-Y. Aberrant Overexpression of HOTAIR Inhibits Abdominal Adipogenesis through Remodelling of Genome-Wide DNA Methylation and Transcription. Mol. Metab. 2022, 60, 101473. [Google Scholar] [CrossRef]
- Li, M.; Guo, Y.; Wang, X.-J.; Duan, B.-H.; Li, L. HOTAIR Participates in Hepatic Insulin Resistance via Regulating SIRT1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7883–7890. [Google Scholar] [CrossRef]
- Becker, P.B.; Workman, J.L. Nucleosome Remodeling and Epigenetics. Cold Spring Harb. Perspect. Biol. 2013, 5, a017905. [Google Scholar] [CrossRef]
- Morrison, A.J. Chromatin-Remodeling Links Metabolic Signaling to Gene Expression. Mol. Metab. 2020, 38, 100973. [Google Scholar] [CrossRef]
- Ebot, E.M.; Gerke, T.; Labbé, D.P.; Sinnott, J.A.; Zadra, G.; Rider, J.R.; Tyekucheva, S.; Wilson, K.M.; Kelly, R.S.; Shui, I.M.; et al. Gene Expression Profiling of Prostate Tissue Identifies Chromatin Regulation as a Potential Link between Obesity and Lethal Prostate Cancer. Cancer 2017, 123, 4130–4138. [Google Scholar] [CrossRef]
- So, J.; Taleb, S.; Wann, J.; Strobel, O.; Kim, K.; Roh, H.C. Chronic cAMP Activation Induces Adipocyte Browning through Discordant Biphasic Remodeling of Transcriptome and Chromatin Accessibility. Mol. Metab. 2022, 66, 101619. [Google Scholar] [CrossRef]
- Zhong, J.; Ji, X.; Zhao, Y.; Jia, Y.; Song, C.; Lv, J.; Chen, Y.; Zhou, Y.; Lv, X.; Yang, Z.; et al. Identification of BAF60b as a Chromatin-Remodeling Checkpoint of Diet-Induced Fatty Liver Disease. Diabetes 2024, 73, 1615–1630. [Google Scholar] [CrossRef]
- Kong, Q.; Zou, J.; Zhang, Z.; Pan, R.; Zhang, Z.Y.; Han, S.; Xu, Y.; Gao, Y.; Meng, Z.-X. BAF60a Deficiency in Macrophage Promotes Diet-Induced Obesity and Metabolic Inflammation. Diabetes 2022, 71, 2136–2152. [Google Scholar] [CrossRef]
- Linnemann, A.K.; Baan, M.; Davis, D.B. Pancreatic β-Cell Proliferation in Obesity. Adv. Nutr. 2014, 5, 278–288. [Google Scholar] [CrossRef]
- Inaishi, J.; Saisho, Y. Beta-Cell Mass in Obesity and Type 2 Diabetes, and Its Relation to Pancreas Fat: A Mini-Review. Nutrients 2020, 12, 3846. [Google Scholar] [CrossRef]
- Spaeth, J.M.; Liu, J.-H.; Peters, D.; Guo, M.; Osipovich, A.B.; Mohammadi, F.; Roy, N.; Bhushan, A.; Magnuson, M.A.; Hebrok, M.; et al. The Pdx1-Bound Swi/Snf Chromatin Remodeling Complex Regulates Pancreatic Progenitor Cell Proliferation and Mature Islet β-Cell Function. Diabetes 2019, 68, 1806–1818. [Google Scholar] [CrossRef]
- Davidson, R.K.; Kanojia, S.; Wu, W.; Kono, T.; Xu, J.; Osmulski, M.; Bone, R.N.; Casey, N.; Evans-Molina, C.; Sims, E.K.; et al. The Chd4 Helicase Regulates Chromatin Accessibility and Gene Expression Critical for β-Cell Function in Vivo. Diabetes 2023, 72, 746–757. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.; Liu, X.; Gu, H.; Li, Y.; Sun, C. Protein Acetylation: A Novel Modus of Obesity Regulation. J. Mol. Med. 2021, 99, 1221–1235. [Google Scholar] [CrossRef]
- Zhong, Q.; Xiao, X.; Qiu, Y.; Xu, Z.; Chen, C.; Chong, B.; Zhao, X.; Hai, S.; Li, S.; An, Z.; et al. Protein Posttranslational Modifications in Health and Diseases: Functions, Regulatory Mechanisms, and Therapeutic Implications. MedComm 2023, 4, e261. [Google Scholar] [CrossRef]
- Ma, Q.-X.; Zhu, W.-Y.; Lu, X.-C.; Jiang, D.; Xu, F.; Li, J.-T.; Zhang, L.; Wu, Y.-L.; Chen, Z.-J.; Yin, M.; et al. BCAA-BCKA Axis Regulates WAT Browning through Acetylation of PRDM16. Nat. Metab. 2022, 4, 106–122. [Google Scholar] [CrossRef]
- Qian, H.; Chen, Y.; Nian, Z.; Su, L.; Yu, H.; Chen, F.-J.; Zhang, X.; Xu, W.; Zhou, L.; Liu, J.; et al. HDAC6-Mediated Acetylation of Lipid Droplet-Binding Protein CIDEC Regulates Fat-Induced Lipid Storage. J. Clin. Invest. 2017, 127, 1353–1369. [Google Scholar] [CrossRef]
- Kim, M.-K.; Kim, E.J.; Kim, J.E.; Lee, D.H.; Chung, J.H. Anacardic Acid Reduces Lipogenesis in Human Differentiated Adipocytes via Inhibition of Histone Acetylation. J. Dermatol. Sci. 2018, 89, 94–97. [Google Scholar] [CrossRef]
- Hall, J.A.; Ramachandran, D.; Roh, H.C.; DiSpirito, J.R.; Belchior, T.; Zushin, P.-J.H.; Palmer, C.; Hong, S.; Mina, A.I.; Liu, B.; et al. Obesity-Linked PPARγ S273 Phosphorylation Promotes Insulin Resistance through Growth Differentiation Factor 3. Cell Metab. 2020, 32, 665–675.e6. [Google Scholar] [CrossRef]
- Pearah, A.; Ramatchandirin, B.; Liu, T.; Wolf, R.M.; Ikeda, A.; Radovick, S.; Sesaki, H.; Wondisford, F.E.; O’Rourke, B.; He, L. Blocking AMPKαS496 Phosphorylation Improves Mitochondrial Dynamics and Hyperglycemia in Aging and Obesity. Cell Chem. Biol. 2023, 30, 1585–1600.e6. [Google Scholar] [CrossRef]
- Kim, M.S.; Baek, J.-H.; Lee, J.; Sivaraman, A.; Lee, K.; Chun, K.-H. Deubiquitinase USP1 Enhances CCAAT/Enhancer-Binding Protein Beta (C/EBPβ) Stability and Accelerates Adipogenesis and Lipid Accumulation. Cell Death Dis. 2023, 14, 776. [Google Scholar] [CrossRef]
- Liu, B.; Wang, T.; Mei, W.; Li, D.; Cai, R.; Zuo, Y.; Cheng, J. Small Ubiquitin-like Modifier (SUMO) Protein-Specific Protease 1 de-SUMOylates Sharp-1 Protein and Controls Adipocyte Differentiation. J. Biol. Chem. 2014, 289, 22358–22364. [Google Scholar] [CrossRef]
- Wu, Y.; Yu, B.; Wang, M. SENP1 Is Required for the Growth, Migration, and Survival of Human Adipose-Derived Stem Cells. Adipocyte 2021, 10, 38–47. [Google Scholar] [CrossRef]
- Lee, J.S.; Chae, S.; Nan, J.; Koo, Y.D.; Lee, S.-A.; Park, Y.J.; Hwang, D.; Han, W.; Lee, D.-S.; Kim, Y.-B.; et al. SENP2 Suppresses Browning of White Adipose Tissues by De-Conjugating SUMO from C/EBPβ. Cell Rep. 2022, 38, 110408. [Google Scholar] [CrossRef]
- Bjune, J.-I.; Laber, S.; Lawrence-Archer, L.; Zhao, X.; Yamada, S.; Al-Sharabi, N.; Mustafa, K.; Njølstad, P.R.; Claussnitzer, M.; Cox, R.D.; et al. Mechanisms of the FTO Locus Association with Obesity: Irx3 Controls a Sumoylation-Dependent Switch between Adipogenesis and Osteogenesis. bioRxiv 2023. [Google Scholar] [CrossRef]
- Cossec, J.-C.; Theurillat, I.; Chica, C.; Búa Aguín, S.; Gaume, X.; Andrieux, A.; Iturbide, A.; Jouvion, G.; Li, H.; Bossis, G.; et al. SUMO Safeguards Somatic and Pluripotent Cell Identities by Enforcing Distinct Chromatin States. Cell Stem Cell 2018, 23, 742–757.e8. [Google Scholar] [CrossRef]
- Theurillat, I.; Hendriks, I.A.; Cossec, J.-C.; Andrieux, A.; Nielsen, M.L.; Dejean, A. Extensive SUMO Modification of Repressive Chromatin Factors Distinguishes Pluripotent from Somatic Cells. Cell Rep. 2020, 33, 108251. [Google Scholar] [CrossRef]
- Zhao, X.; Hendriks, I.A.; Le Gras, S.; Ye, T.; Ramos-Alonso, L.; Nguéa P, A.; Lien, G.F.; Ghasemi, F.; Klungland, A.; Jost, B.; et al. Waves of Sumoylation Support Transcription Dynamics during Adipocyte Differentiation. Nucleic Acids Res. 2022, 50, 1351–1369. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Wen, R.; Yang, N.; Zhang, T.-N.; Liu, C.-F. Roles of Protein Post-Translational Modifications in Glucose and Lipid Metabolism: Mechanisms and Perspectives. Mol. Med. Camb. Mass 2023, 29, 93. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, F.; Li, M.; Zhang, Y.; Li, N.; Shao, L. MiR-34a-5p Promotes Hepatic Gluconeogenesis by Suppressing SIRT1 Expression. Exp. Cell Res. 2022, 420, 113336. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, S.; Xiao, M.; Lin, Y.; Zhou, L.; Lei, Q.; Xiong, Y.; Guan, K.-L.; Zhao, S. Acetylation Regulates Gluconeogenesis by Promoting PEPCK1 Degradation via Recruiting the UBR5 Ubiquitin Ligase. Mol. Cell 2011, 43, 33–44. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Sarma, P.; Barge, S.R.; Swargiary, D.; Devi, G.S.; Borah, J.C. Xanthosine, a Purine Glycoside Mediates Hepatic Glucose Homeostasis through Inhibition of Gluconeogenesis and Activation of Glycogenesis via Regulating the AMPK/ FoxO1/AKT/GSK3β Signaling Cascade. Chem. Biol. Interact. 2023, 371, 110347. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Li, E.; Chen, L.; Zhang, Y.; Wei, F.; Liu, J.; Deng, H.; Wang, Y. The CREB Coactivator CRTC2 Controls Hepatic Lipid Metabolism by Regulating SREBP1. Nature 2015, 524, 243–246. [Google Scholar] [CrossRef]
- Li, N.; Luo, X.; Yu, Q.; Yang, P.; Chen, Z.; Wang, X.; Jiang, J.; Xu, J.; Gong, Q.; Eizirik, D.L.; et al. SUMOylation of Pdia3 Exacerbates Proinsulin Misfolding and ER Stress in Pancreatic Beta Cells. J. Mol. Med. Berl. Ger. 2020, 98, 1795–1807. [Google Scholar] [CrossRef]
- Xie, H.; Wang, Y.-H.; Liu, X.; Gao, J.; Yang, C.; Huang, T.; Zhang, L.; Luo, X.; Gao, Z.; Wang, T.; et al. SUMOylation of ERp44 Enhances Ero1α ER Retention Contributing to the Pathogenesis of Obesity and Insulin Resistance. Metabolism 2023, 139, 155351. [Google Scholar] [CrossRef]
- Liu, Y.; Dou, X.; Zhou, W.-Y.; Ding, M.; Liu, L.; Du, R.-Q.; Guo, L.; Qian, S.-W.; Tang, Y.; Yang, Q.-Q.; et al. Hepatic Small Ubiquitin-Related Modifier (SUMO)-Specific Protease 2 Controls Systemic Metabolism through SUMOylation-Dependent Regulation of Liver-Adipose Tissue Crosstalk. Hepatology 2021, 74, 1864–1883. [Google Scholar] [CrossRef]
- Maschari, D.; Saxena, G.; Law, T.D.; Walsh, E.; Campbell, M.C.; Consitt, L.A. Lactate-Induced Lactylation in Skeletal Muscle Is Associated with Insulin Resistance in Humans. Front. Physiol. 2022, 13, 951390. [Google Scholar] [CrossRef]
- Gao, R.; Li, Y.; Xu, Z.; Zhang, F.; Xu, J.; Hu, Y.; Yin, J.; Yang, K.; Sun, L.; Wang, Q.; et al. Mitochondrial Pyruvate Carrier 1 Regulates Fatty Acid Synthase Lactylation and Mediates Treatment of Nonalcoholic Fatty Liver Disease. Hepatology 2023, 78, 1800–1815. [Google Scholar] [CrossRef]
- Li, F.; Jing, J.; Movahed, M.; Cui, X.; Cao, Q.; Wu, R.; Chen, Z.; Yu, L.; Pan, Y.; Shi, H.; et al. Epigenetic Interaction between UTX and DNMT1 Regulates Diet-Induced Myogenic Remodeling in Brown Fat. Nat. Commun. 2021, 12, 6838. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Meng, X.; Wang, W.; Duan, F.; Chen, S.; Zhang, Y.; Sheng, Z.; Gao, Y.; Zhou, L. METTL16 Inhibits Papillary Thyroid Cancer Tumorigenicity through m6A/YTHDC2/SCD1-Regulated Lipid Metabolism. Cell. Mol. Life Sci. CMLS 2024, 81, 81. [Google Scholar] [CrossRef]
- Zhang, T.; Ren, Z.; Mao, R.; Yi, W.; Wang, B.; Yang, H.; Wang, H.; Liu, Y. LINC00278 and BRG1: A Key Regulatory Axis in Male Obesity and Preadipocyte Adipogenesis. Metabolism 2025, 168, 156194. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, G.-L.; Zhu, X.; Peng, T.-H.; Lv, Y.-C. Critical Roles of FTO-Mediated mRNA m6A Demethylation in Regulating Adipogenesis and Lipid Metabolism: Implications in Lipid Metabolic Disorders. Genes Dis. 2022, 9, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Song, M.; Hong, Z.; Chen, W.; Zhang, Q.; Zhou, J.; Yang, C.; He, Z.; Yu, J.; Peng, X.; et al. The N6-Methyladenosine METTL3 Regulates Tumorigenesis and Glycolysis by Mediating m6A Methylation of the Tumor Suppressor LATS1 in Breast Cancer. J. Exp. Clin. Cancer Res. CR 2023, 42, 10. [Google Scholar] [CrossRef] [PubMed]
- Hinte, L.C.; Castellano-Castillo, D.; Ghosh, A.; Melrose, K.; Gasser, E.; Noé, F.; Massier, L.; Dong, H.; Sun, W.; Hoffmann, A.; et al. Adipose Tissue Retains an Epigenetic Memory of Obesity after Weight Loss. Nature 2024, 636, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Chao, S.; Lu, J.; Li, L.-J.; Guo, H.-Y.; Xu, K.; Wang, N.; Zhao, S.-X.; Jin, X.-W.; Wang, S.-G.; Yin, S.; et al. Maternal Obesity May Disrupt Offspring Metabolism by Inducing Oocyte Genome Hyper-Methylation via Increased DNMTs. eLife 2024, 13, RP97507. [Google Scholar] [CrossRef]
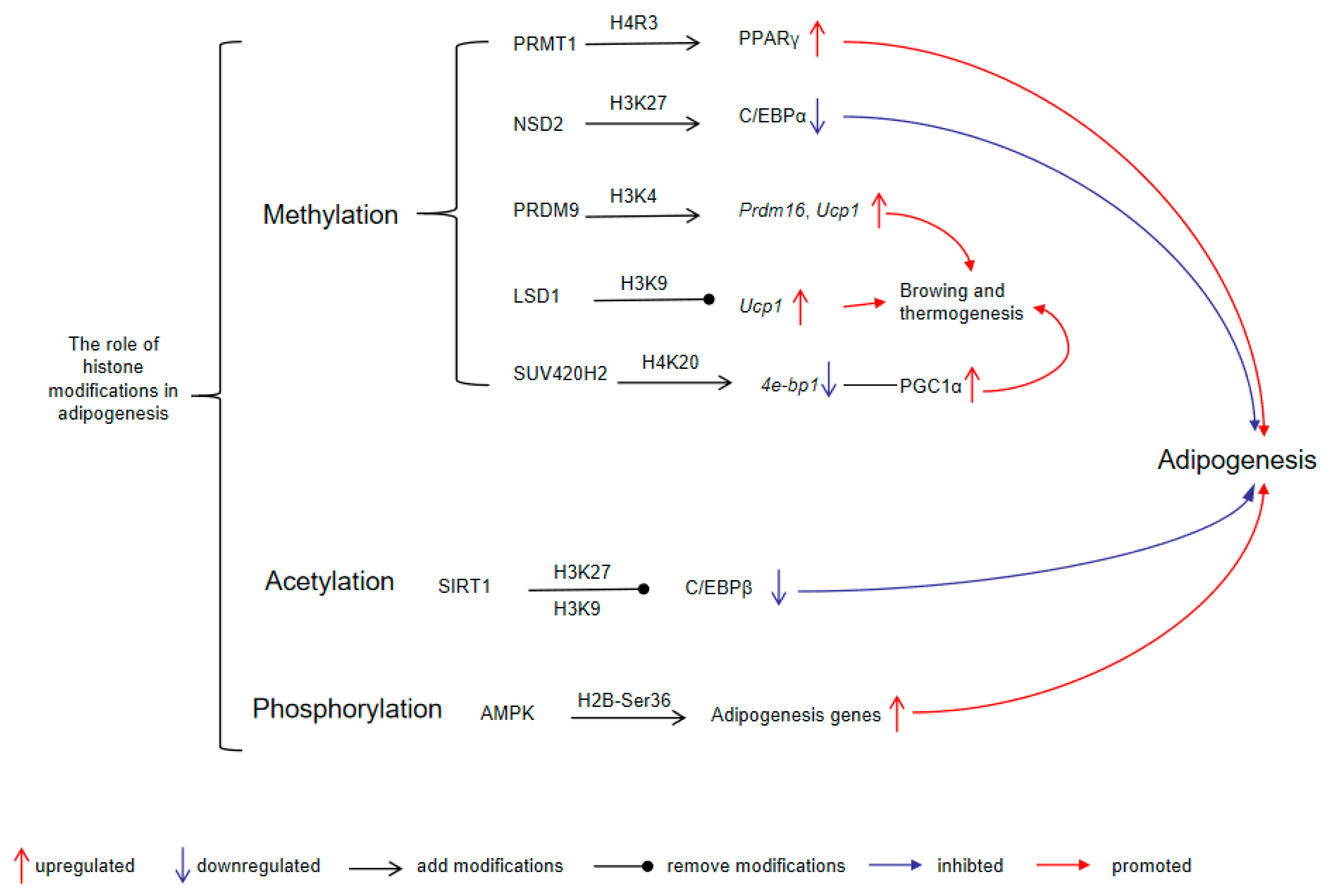
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).