_Kim.png)
Transcriptomic Analysis of the Negative Effect of Epigallocatechin-3-Gallate from Tea Plant (Camellia sinensis) on Agrobacterium-Mediated Transformation Efficiency

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sample Collection
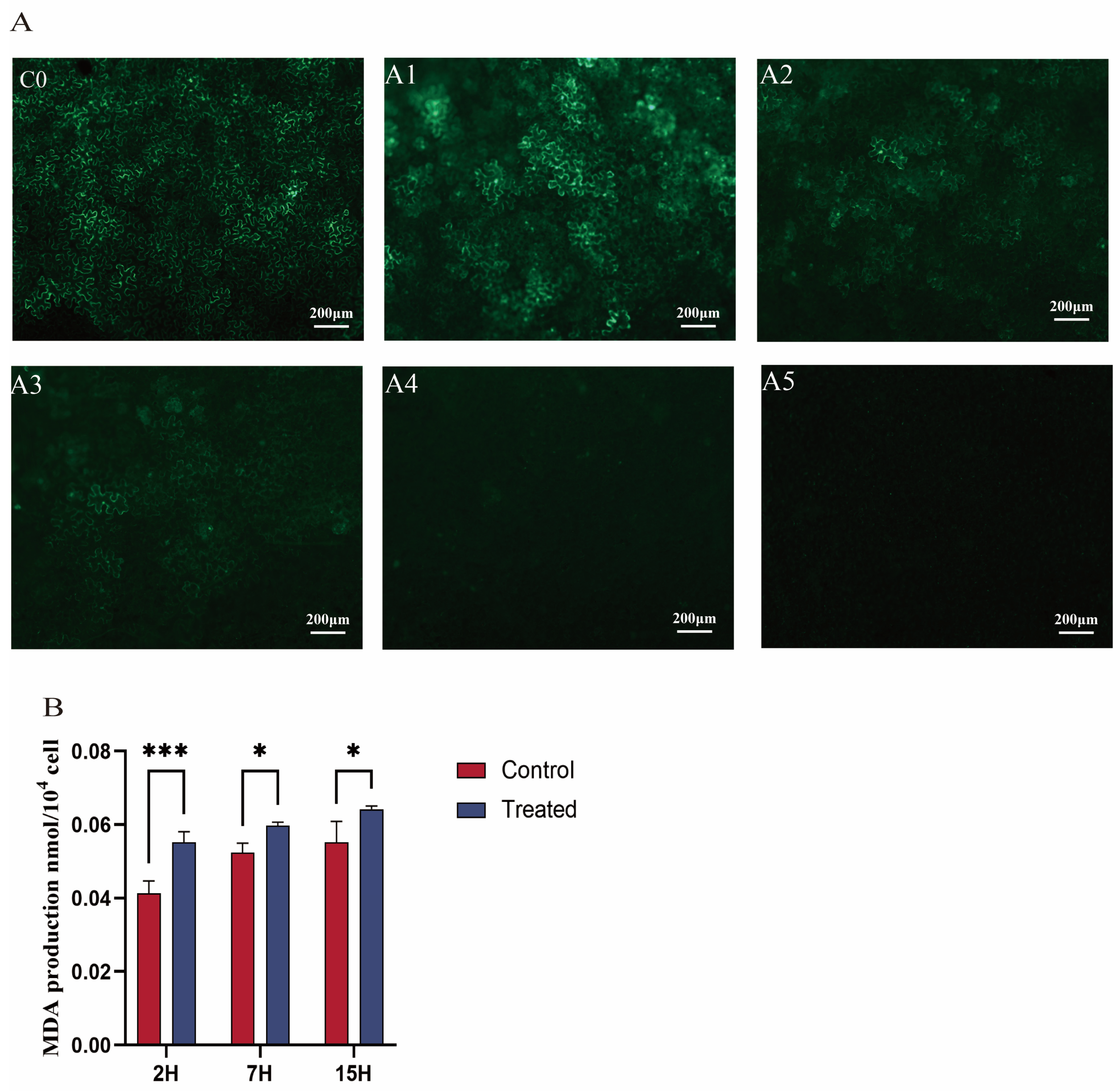
2.2. Green Fluorescence GFP Observation
2.3. Measurement of MDA
2.4. RNA Extraction and Transcriptome Sequencing
2.5. Gene Annotation, Enrichment, and Differential Expression Analysis
2.6. Co-Expression Network Analysis
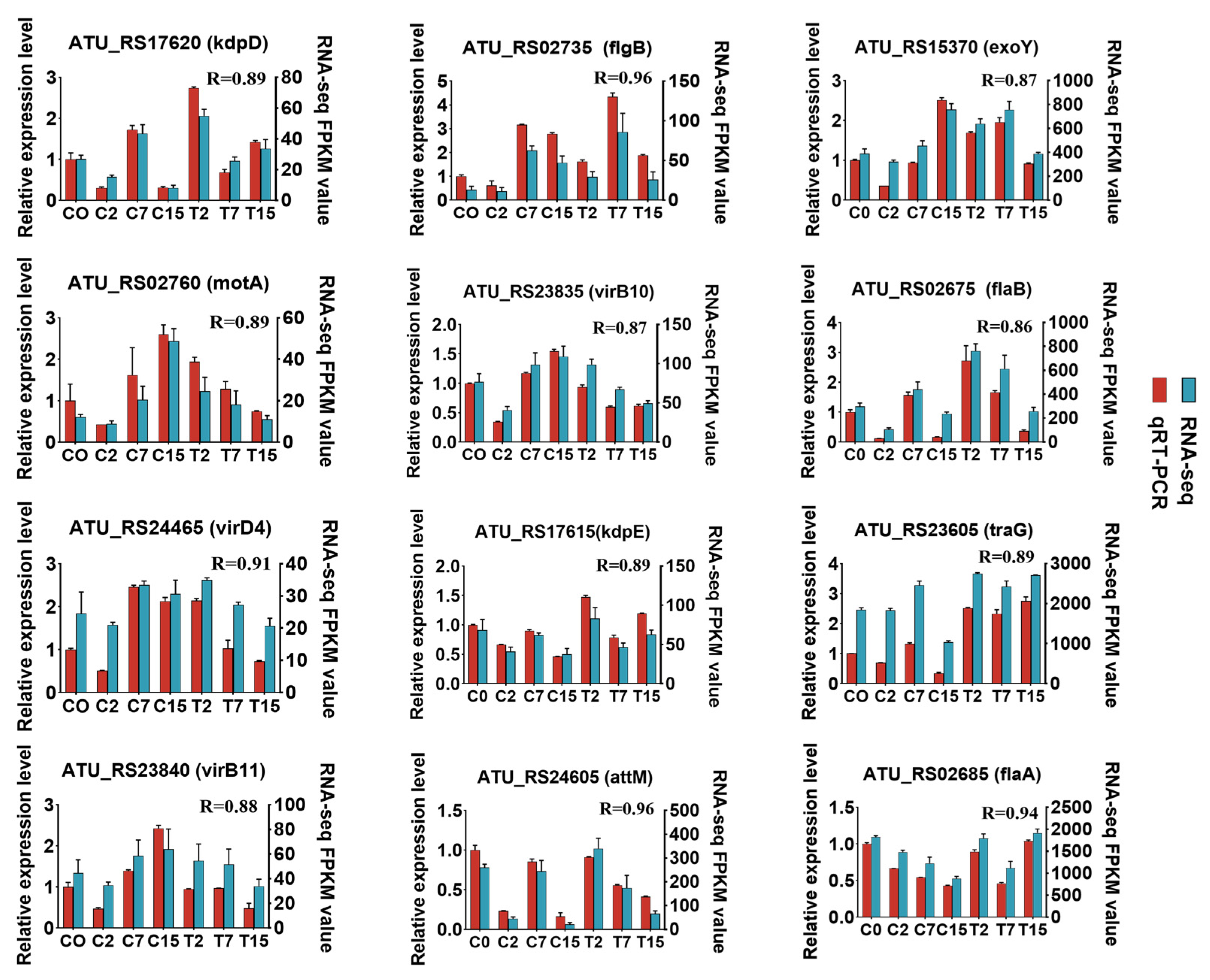
2.7. Quantitative RT-PCR (qRT-PCR) Validation
3. Results
3.1. Effect of EGCG on Agrobacterium-Mediated Genetic Transformation
3.2. Effect of EGCG on the Agrobacterium Cell Membrane
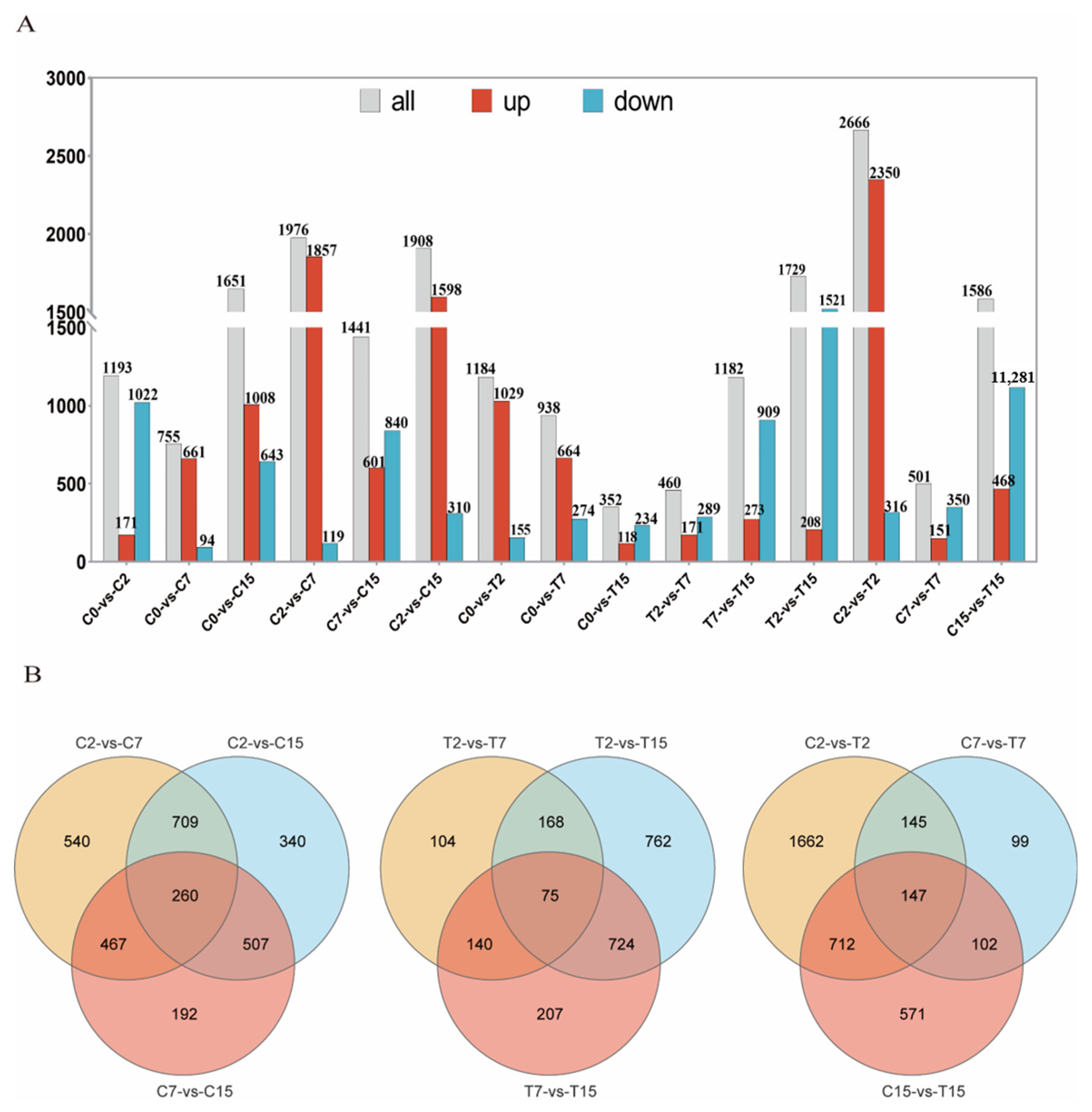
3.3. Differential Gene Expression Analysis in Agrobacterium Caused by EGCG
3.4. Venn Analysis of the DEGs
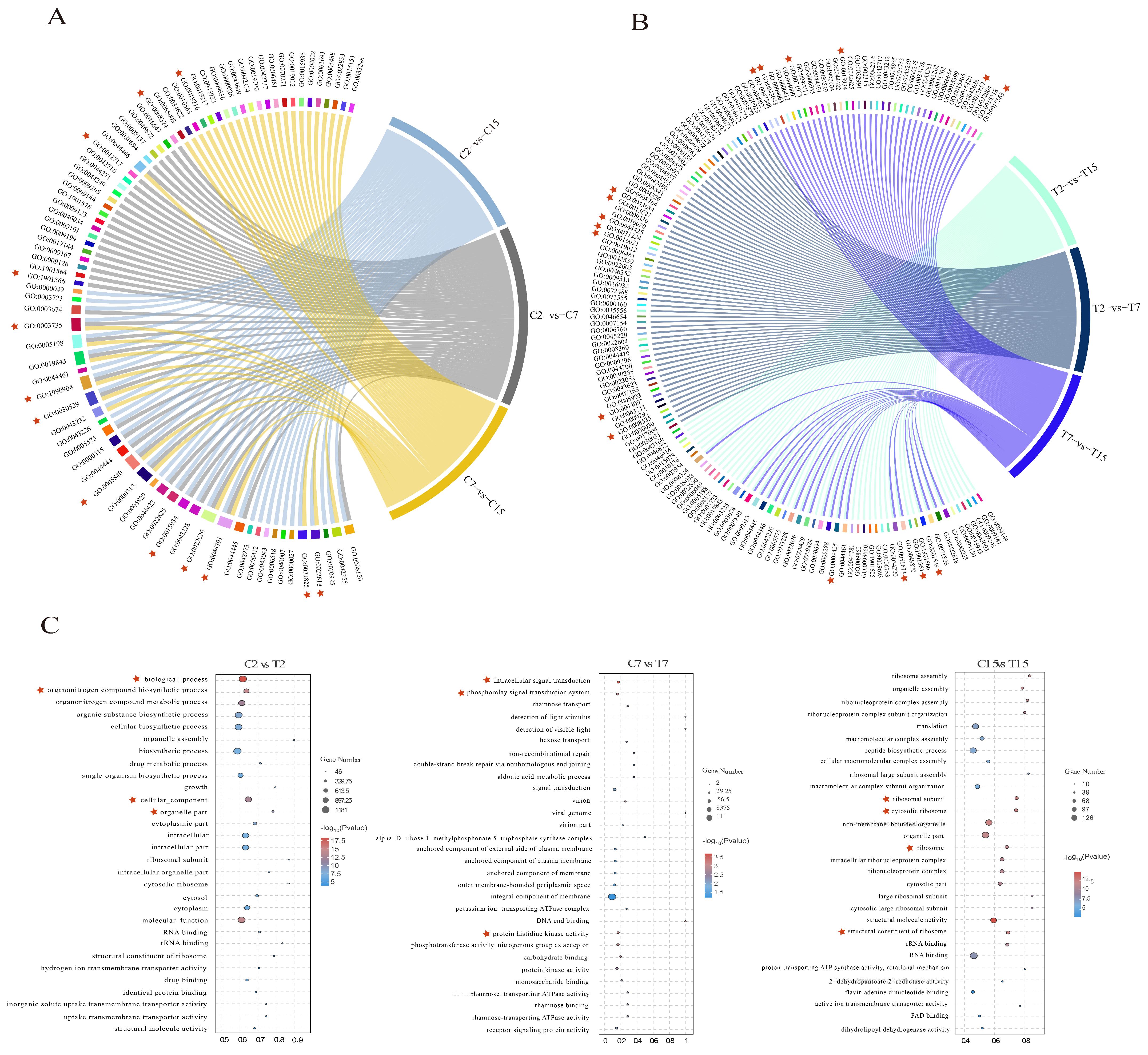
3.5. GO Enrichment Analysis of the DEGs
3.6. KEGG Enrichment Analysis
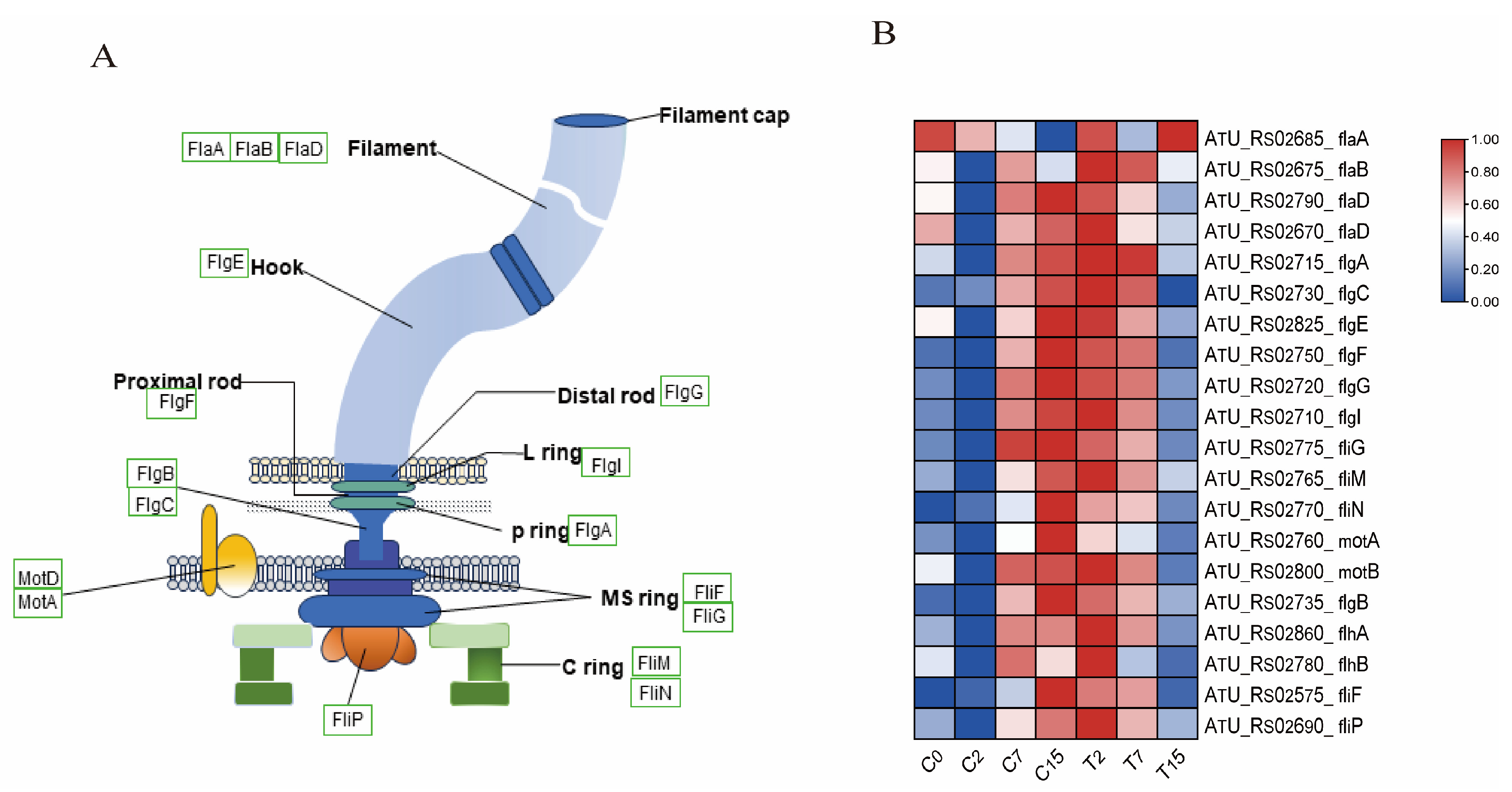
3.7. The DEGs Involved in the Flagellar Synthesis Pathway
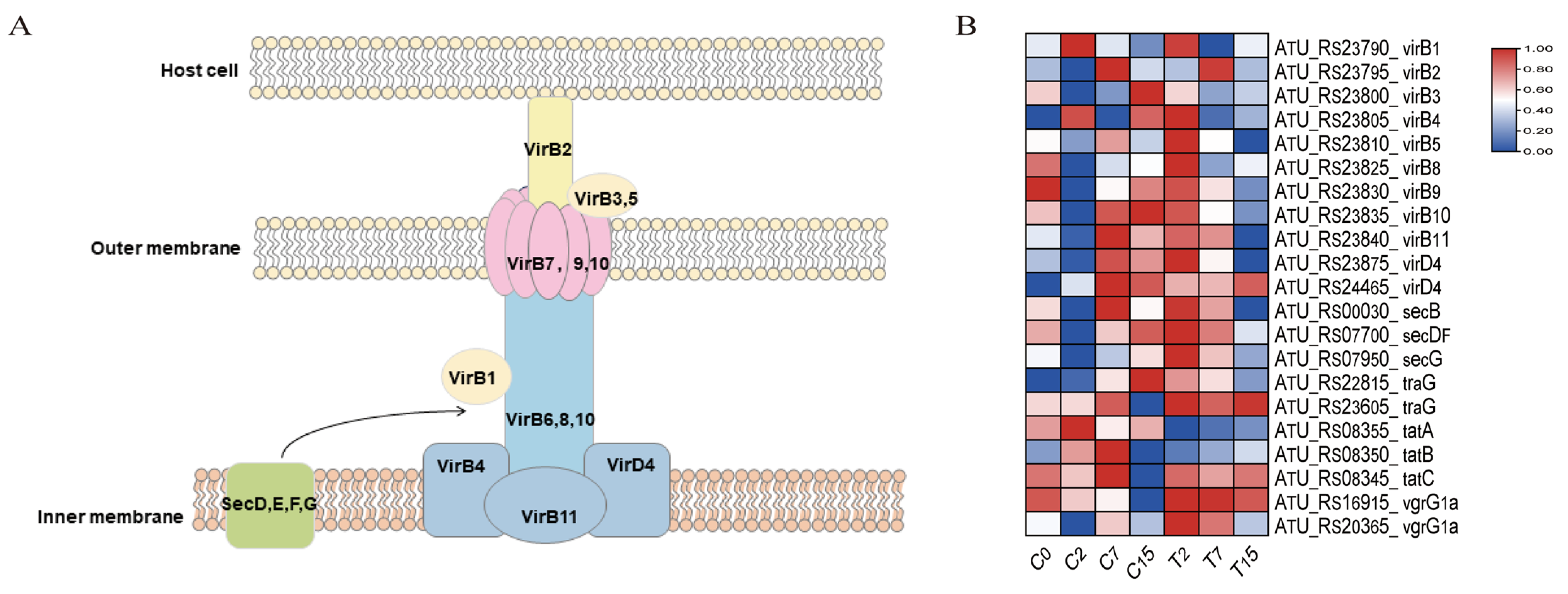
3.8. The DEGs Involved in the Bacterial Secretion System Pathway
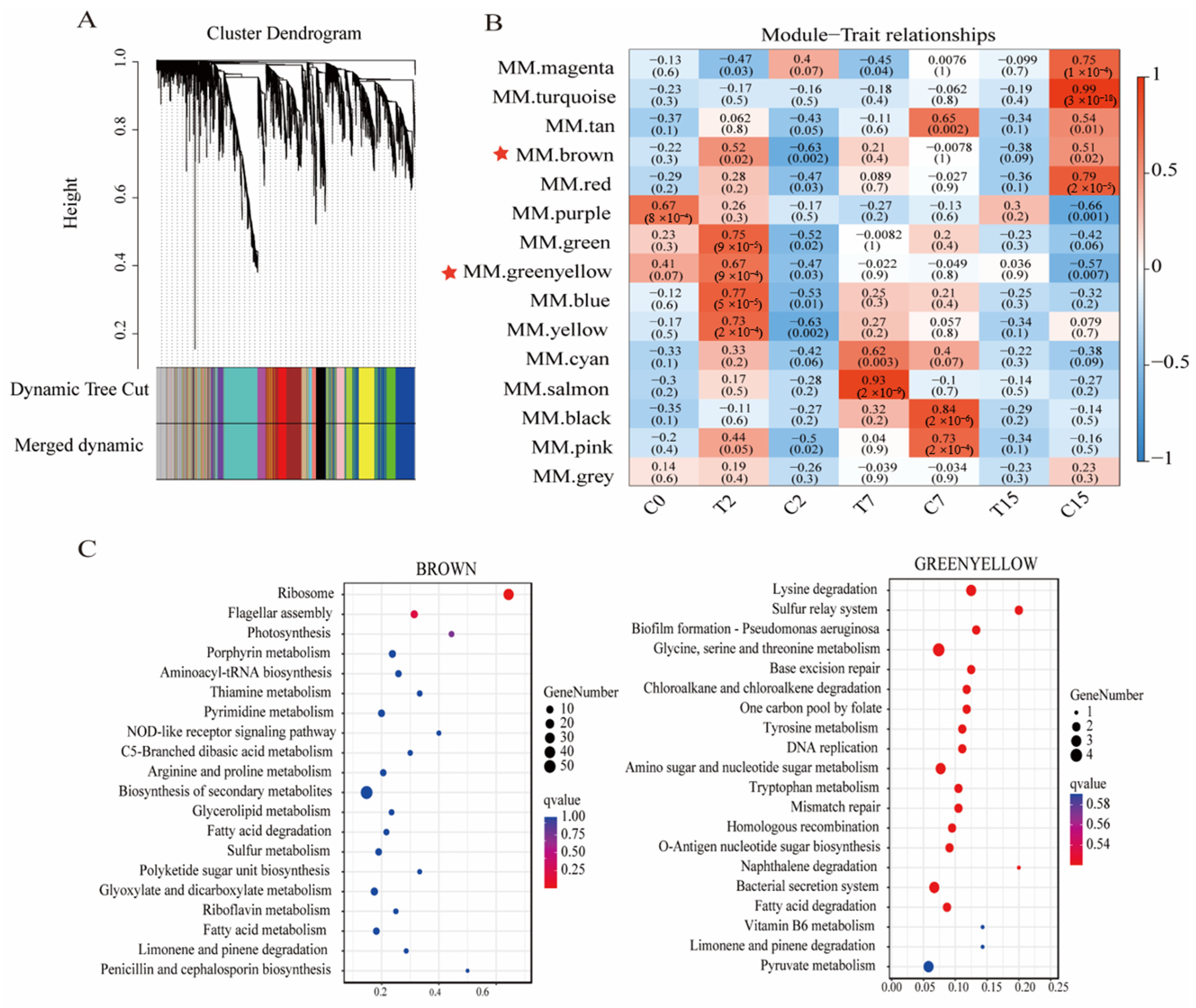
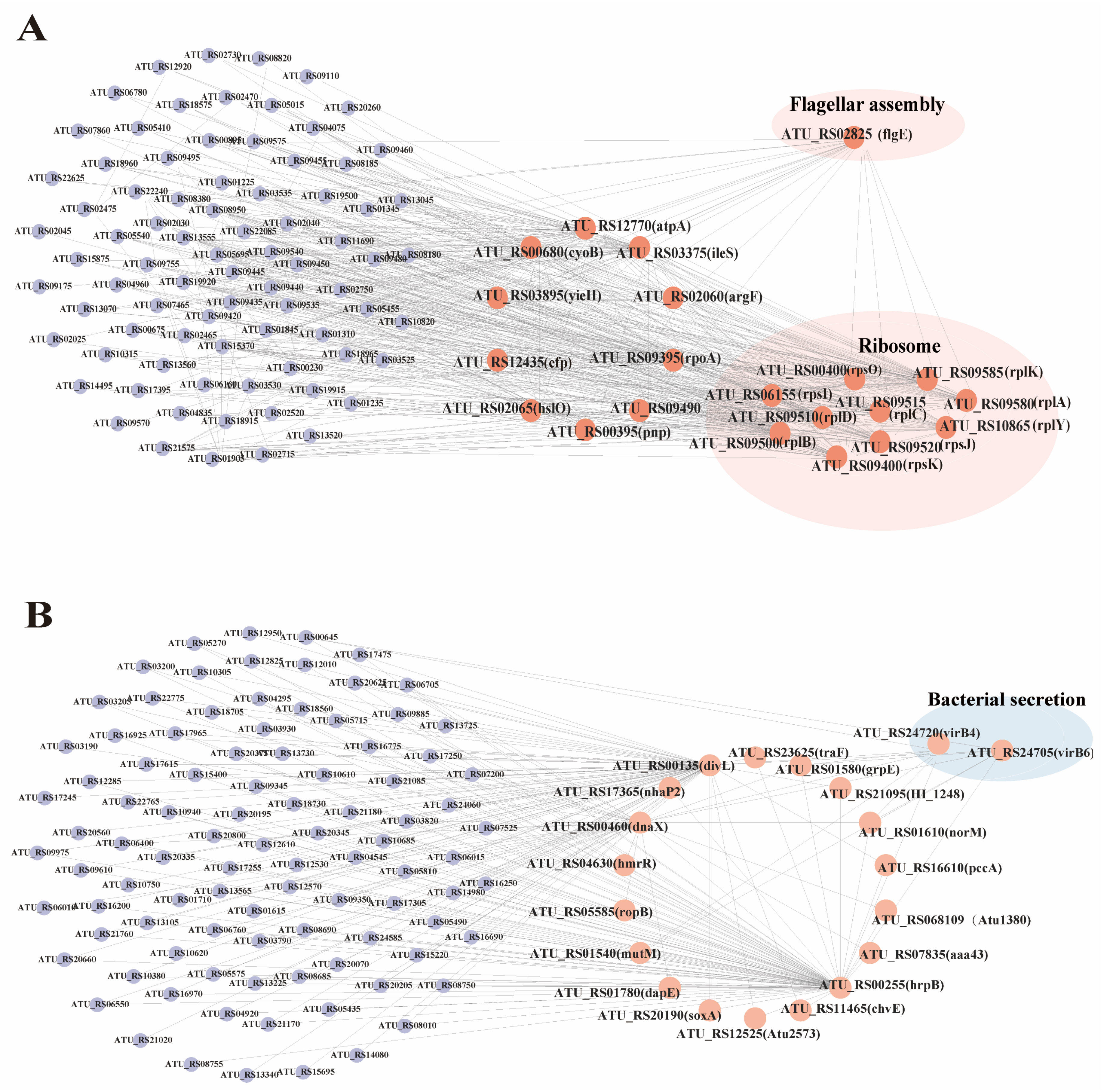
3.9. WGCNA Analysis
4. Discussion
4.1. EGCG Is an Important Factor in Tea Plant Recalcitrance to Agrobacterium-Mediated Transformation
4.2. EGCG Hinders Agrobacterium Biofilm Formation
4.3. EGCG Disrupted the T-DNA Transfer of Agrobacterium
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, H.J.; Liu, N.G.; Sun, X.P.; Zhu, M.L.; Mao, T.F.; Huang, S.Y.; Meng, X.Y.; Li, H.F.; Wang, M.; Liang, H.L. Establishment of an efficient transformation system and its application in regulatory mechanism analysis of biological macromolecules in tea plants. Int. J. Biol. Macromol. 2023, 244, 125372. [Google Scholar] [CrossRef]
- Azizi-Dargahlou, S.; Pouresmaeil, M. Agrobacterium tumefaciens-mediated plant Transformation: A review. Mol. Biotechnol. 2024, 66, 1563–1580. [Google Scholar] [CrossRef]
- Schröpfer, S.; Lempe, J.; Emeriewen, O.F.; Emeriewen, O.F.; Flachowsky, H. Recent Developments and Strategies for the Application of Agrobacterium-Mediated Transformation of Apple Malus × domestica Borkh. Front. Plant Sci. 2022, 13, 928292. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.H.; Wu, Z.Y.; Wang, X.Q.; Mei, Y.L.; Chen, W.R.; Liao, F.L.; Guo, W.D. Research on the Establishment of TRV-VIGS System of Fruit Shape- related Genes CRC in Fingered Citron and Hongkong Kumquat. Acta Hortic. Sin. 2024, 51, 715–726. [Google Scholar]
- Han, P.A.; Cheng, Y.; Tang, K.G.; Li, X.D.; Wang, L.W.; Liang, Y.H.; Jing, Y.; Shi, H.B.; Wu, X.R. Study on Agrobacterium mediated transfer of CP4-EPSPS gene into maize inbred line B73. J. North. Agric. 2023, 51, 31–37. [Google Scholar]
- Jiao, X.Y.; Wang, L.G.; Liu, D.D.; Ruan, X.; Xu, Y.D.; Wu, Q.; Sun, M.H.; Wang, W.J. Research progress on genetic transformation of Camellia sinensis (L.) O. Kuntze. Plant Sci. J. 2021, 39, 446–456. [Google Scholar]
- Zhang, Z.L.; Ren, L.L.; Yan, X.M.; Nie, X.R.; Yang, T.Y. Research progress on regeneration in vitro and genetic transformation system of Camellia sinensis. J. Anhui Agric. Univ. 2021, 48, 738–743. [Google Scholar]
- Matthysse, A.G. Initial interactions of Agrobacterium tumefaciens with plant host cells. Crit. Rev. Microbiol. 1986, 13, 281–307. [Google Scholar] [CrossRef]
- Hwang, H.H.; Yu, M.; Lai, E.M. Agrobacterium-mediated plant transformation: Biology and applications. Arab. Book 2017, 15, e0186. [Google Scholar] [CrossRef]
- Heindl, J.E.; Wang, Y.; Heckel, B.C.; Mohari, B.; Feirer, N.; Fuqua, C. Mechanisms and regulation of surface interactions and biofilm formation in Agrobacterium. Front. Plant Sci. 2014, 5, 176. [Google Scholar] [CrossRef]
- Laventie, B.J.; Jenal, U. Surface Sensing and Adaptation in Bacteria. Annu. Rev. Microbiol. 2020, 74, 735–760. [Google Scholar] [CrossRef]
- Wood, T.K.; González, B.A.F.; Herzberg, M.; Lee, J. Motility influences biofilm architecture in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 72, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Bruzaud, J.; Tarrade, J.; Coudreuse, A.; Canette, A.; Herry, J.M.; Taffin de Givenchy, E.; Darmanin, T.; Guittard, F.; Guilbaud, M.; Bellon-Fontaine, M.N. Flagella but not type IV pili are involved in the initial adhesion of Pseudomonas aeruginosa PAO1 to hydrophobic or superhydrophobic surfaces. Colloids Surf. B Biointerfaces 2015, 131, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Chesnokova, O.; Coutinho, J.B.; Khan, I.H.; Mikhail, M.S.; Kado, C.I. Characterization of flagella genes of Agrobacterium tumefaciens, and the effect of a bald strain on virulence. Mol. Microbiol. 1997, 23, 579–590. [Google Scholar] [CrossRef]
- Merritt, P.M.; Danhorn, T.; Fuqua, C. Motility and chemotaxis in Agrobacterium tumefaciens surface attachment and biofilm formation. J. Bacteriol. 2007, 189, 8005–8014. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zou, H.; Wang, Y.; Jin, J.; Wang, H.; Zhou, M. Inhibition effect of epigallocatechin gallate on the growth and biofilm formation of Vibrio parahaemolyticuss. Lett. Appl. Microbiol. 2022, 75, 81–88. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Ma, R.; Sun, W.; Ji, Z. Antibacterial Activity of Epigallocatechin Gallate (EGCG) against Shigella flexneri. Int. J. Environ. Res. Public Health 2023, 20, 4676. [Google Scholar] [CrossRef]
- Schneider-Rayman, M.; Steinberg, D.; Sionov, R.V.; Friedman, M.; Shalish, M. Effect of epigallocatechin gallate on dental biofilm of Streptococcus mutans: An in vitro study. BMC Oral Health 2021, 21, 447. [Google Scholar] [CrossRef]
- Wang, G.P.; Zhang, T.; Shi, Z.G.; Chen, R.H. Research Progress on T-DNA Transfer Related Genes Mediated of Agrobacterium. Mol. Plant Breed. 2017, 15, 1752–1761. [Google Scholar]
- Thompson, M.G.; Moore, W.M.; Hummel, N.F.C.; Pearson, A.N.; Barnum, C.R.; Scheller, H.V.; Shih, P.M. Agrobacterium tumefaciens: A bacterium primed for synthetic biology. Biodesign Res. 2020, 2020, 8189219. [Google Scholar] [CrossRef]
- Wangthaisong, P.; Piromyou, P.; Songwattana, P.; Wongdee, J.; Teamtaisong, K.; Tittabutr, P.; Boonkerd, N.; Teaumroong, N. The Type IV Secretion System (T4SS) Mediates Symbiosis between Bradyrhizobium sp. SUTN9-2 and Legumes. Appl. Environ. Microbiol. 2023, 89, e0004023. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.M.; Yue, Y.B.; Luo, M.C.; Lu, X.L. Roles of the type IV secretion systems in bacterial resistance to heavy metals. Microbiol. China 2024, 51, 1460–1470. [Google Scholar]
- Christie, P.J.; Whitaker, N.; González-Rivera, C. Mechanism and structure of the bacterial type IV secretion systems. Biochim. Biophys. Acta 2014, 1843, 1578–1591. [Google Scholar] [CrossRef] [PubMed]
- De Saeger, J.; Park, J.; Chung, H.S. Agrobacterium strains and strain improvement: Present and outlook. Biotechnol. Adv. 2021, 53, 107677. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ling, C.R.; Huang, Y.; Deng, X.M.; Wang, Y.Q.; Sun, W.J. Effects of Tea Polyphenols on Agrobacterium-mediated Plant Genetic Transformation System. J. Tea Sci. 2022, 42, 477–490. [Google Scholar]
- Alagarsamy, K.; Shamala, L.F.; Wei, S. Protocol: High-efficiency in-planta Agrobacterium-mediated transgenic hairy root induction of Camellia sinensis var. sinensis. Plant Methods. 2018, 14, 17. [Google Scholar] [CrossRef]
- Sandal, I.; Saini, U.; Lacroix, B.; Bhattacharya, A.; Ahuja, P.S. Agrobacterium-mediated genetic transformation of tea leaf explants: Effects of counteracting bactericidity of leaf polyphenols without loss of bacterial virulence. Plant Cell Rep. 2007, 26, 169–176. [Google Scholar] [CrossRef]
- Kumar, N.; Gulati, A.; Bhattacharya, A. L-glutamine and L-glutamic Acid Facilitate Successful Agrobacterium Infection of Recalcitrant Tea Cultivars. Appl. Biochem. Biotechnol. 2013, 170, 1649–1664. [Google Scholar] [CrossRef]
- Bhattacharyya, N.; Singh, H.R.; Bhagawati, P. Agrobacterium mediated transfer of nptII and gus genes in Camellia assamica. Agric. Food Sci. 2014, 6, 22–28. [Google Scholar]
- Han, X.Y.; Chen, Y.; Yang, Y.; Xu, W.; Wei, L. Advances in the Anti-infective Effects of Epigallocatechin Gallate. Chin. J. Mod. Appl. Pharm. 2021, 38, 2616–2620. [Google Scholar]
- Hao, S.; Yang, D.; Zhao, L.; Shi, F.; Ye, G.; Fu, H.; Lin, J.; Guo, H.; He, R.; Li, J.; et al. EGCG-Mediated Potential Inhibition of Biofilm Development and Quorum Sensing in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 4946. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Tian, N.; da Silva Ferreira, J.F.; Sandhu, D.; Xiao, L.; Gu, M.; Luo, Y.; Zhang, X.; Liu, G.; Liu, Z.; et al. Comparative Transcriptome Analysis of Agrobacterium tumefaciens Reveals the Molecular Basis for the Recalcitrant Genetic Transformation of Camellia sinensis L. Biomolecules 2022, 12, 688. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, Y.; Wu, H.Z.; Min, X.Y.; Zhang, B.; Kim, D.; Yan, X.B.; Chuan-Jie Zhang, C.J. Optimization and establishment of Agrobacterium-mediated transformation in alfalfa (Medicago sativa L.) using eGFP as a visual reporter. Plant Cell Tissue Organ Cult. 2024, 156, 7. [Google Scholar] [CrossRef]
- Goodner, B.; Hinkle, G.; Gattung, S.; Miller, N.; Blanchard, M.; Qurollo, B.; Goldman, B.S.; Cao, Y.; Askenazi, M.; Halling, C.; et al. Genome sequence of the plant pathogen and biotechnology agent Agrobacterium tumefaciens C58. Science 2001, 294, 2323–2328. [Google Scholar] [CrossRef] [PubMed]
- Peter, L.; Steve, H. WGCNA: An R package for weighted gene co-expression network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar]
- Shannon, P. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Hsu, K.C.; Hsu, P.F.; Chen, Y.C.; Lin, H.C.; Hung, C.C.; Chen, P.C.; Huang, Y.L. Oxidative stress during bacterial growth characterized through microdialysis sampling coupled with HPLC/fluorescence detection of malondialdehyde. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1019, 112–116. [Google Scholar] [CrossRef]
- Xie, Y.; Li, J.; Ding, Y.; Shao, X.; Sun, Y.; Xie, F.; Liu, S.; Tang, S.; Deng, X. An atlas of bacterial two-component systems reveals function and plasticity in signal transduction. Cell Rep. 2022, 41, 111502. [Google Scholar] [CrossRef]
- Wang, M.M.; Wu, S.B.; Wu, H.; Zhang, P.; Zhang, Y.M.; Qiao, J.J.; Cai, Y.Q.G.L. Research Progress on Polyphenol-based Quorum Sensing Interfering. China Biotechnol. 2022, 42, 93–104. [Google Scholar]
- Hengge, R. Targeting Bacterial Biofilms by the Green Tea Polyphenol EGCG. Molecules 2019, 24, 2403. [Google Scholar] [CrossRef]
- Song, D.P.; Lin, F.; Rana, M.M.; Ming, J.G.; Wei, S. Effects of catechins on Agrobacterium-mediated genetic transformation of Camellia sinensis. Plant Cell Tissue Organ Cult. (Pctoc) 2014, 119, 27–37. [Google Scholar] [CrossRef]
- Gong, G.L.; Hu, L.M.; Jia, X.Y.; Qi, Q. Inhibitory effect of catechin on Enterobacter sakazakii in vitro. J. Shaanxi Univ. Sci. Technol. 2023, 41, 62–67. [Google Scholar]
- Chen, X.Q.; Zhao, Y.Y.; Zhen, Z.; Wu, P.J.; Li, W.; Du, J.M.; Zhang, S.X.; Wang, P.; Wang, L. Analysis of cinnamon oil composition and its mechanism of cell membrane damage in Salmonella enterica. Food Ferment. Ind. 2022, 14, 24–32. [Google Scholar]
- Petrarca, P.; Ammendola, S.; Pasquali, P.; Battistoni, A. The Zur-Regulated ZinT Protein Is an Auxili ary Component of the High-Affinity ZnuABC Zinc Transporter That Facilitates Metal R ecruitment during Severe Zinc Shortage. J. Bacteriol. 2010, 192, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Chinnam, N.; Dadi, P.K.; Sabri, S.A.; Ahmad, M.; Kabir, M.A.; Ahmad, Z. Dietary bioflavonoids inhibit Escherichia coli A TP synthase in a differential manner. Int. J. Biol. Macromol. 2010, 46, 478–486. [Google Scholar] [CrossRef]
- Dan, Y.; Zhang, S.; Zhong, H.; Hochul Yi, H.C.; Sainz, M.B. Novel compounds that enhance Agrobacterium-mediated plant transformation by mitigating oxidative stress. Plant Cell Rep. 2015, 34, 291–309. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Jailani, A.; Lee, J.H.; Lee, J. Effect of halogenated indoles on biofilm formation, virulence, and root surface colonization by Agrobacterium tumefaciens. Chemosphere 2022, 293, 133603. [Google Scholar] [CrossRef]
- Jailani, A.; Ahmed, B.; Lee, j.h.; Lee, J.T. Inhibition of Agrobacterium tumefaciens growth and biofilm formation by tannic acid. Biomedicines 2022, 10, 1619. [Google Scholar] [CrossRef]
- Feirer, N.; Kim, D.; Xu, J.; Fernandez, N.; Waters, C.M.; Fuqua, C. The Agrobacterium tumefaciens CheY-like protein ClaR regulates biofilm formation. Microbiology 2017, 163, 1680–1691. [Google Scholar] [CrossRef]
- Inoue, Y.; Morimoto, Y.V.; Namba, K.; Minamino, T. Novel insights into the mechanism of well-ordered assembly of bacterial flagellar proteins in Salmonella. Sci. Rep. 2018, 8, 1787. [Google Scholar] [CrossRef]
- Hu, H.; Santiveri, M.; Wadhwa, N.; Berg, H.C.; Erhardt, M.; Taylor, N.M.I. Structural basis of torque generation in the bi-directional bacterial flagellar motor. Trends Biochem. Sci. 2022, 47, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Mohari, B.; Thompson, M.A.; Trinidad, J.C.; Setayeshgar, S.; Fuqua, C. Multiple Flagellin Proteins Have Distinct and Synergistic Roles in Agrobacterium tumefaciens Motility. J. Bacteriol. Am. Soc. Microbiol. 2018, 200, e00327-18. [Google Scholar] [CrossRef]
- Xu, S.; Li, C.; Jian, J.L.; Xie, X.C.; Wang, B.; Huo, Z.M.; Fang, L. Digging the chemokine receptors-encoding genes and analyzing the relative protein sequences of a Halomonas strain. J. Dalian Ocean. Univ. 2024, 39, 203–214. [Google Scholar]
- Alakavuklar, M.A.; Heckel, B.C.; Stoner, A.M.; Stembel, J.A.; Fuqua, C. Motility control through an anti-activation mechanism in Agrobacterium tumefaciens. Mol. Microbiol. 2021, 116, 1281–1297. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Sinha, R.; Mukherjee, P.; Howlader, D.R.; Nag, D.; Sarkar, S.; Koley, H.; Withey, J.H.; Gachhui, R. Anti-virulence activity of polyphenolic frac Agrobacterium strains and strain improvement: Present and out tion isolated from Kombucha against Vibrio cholerae. Microb. Pathog. 2020, 140, 103927. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Lu, J.Y.; Duan, H.B.; Yang, J. Biofilm inhibition and mode of action of epigallocatechin gallate against Vibrio mimicus. Food Control 2020, 113, 107148. [Google Scholar] [CrossRef]
- You, Y.; Ye, F.; Mao, W.; Yang, H.; Lai, J.; Deng, S. An overview of the structure and function of the flagellar hook FlgE protein. World J. Microbiol. Biotechnol. 2023, 39, 126. [Google Scholar] [CrossRef]
- Valentin, J.D.P.; Straub, H.; Pietsch, F.; Lemare, M.; Ahrens, C.H.; Schreiber, F.; Webb, J.S.; Mei, H.; Ren, Q. Role of the flagellar hook in the structural development and antibiotic tolerance of Pseudomonas aeruginosa motility. ISME J. 2022, 16, 1176–1186. [Google Scholar] [CrossRef]
- Zheng, H.; Mao, Y.; Teng, J.; Zhu, Q.; Ling, J.; Zhong, Z. Flagellar-Dependent Motility in Mesorhizobium tianshanense is Involved in the Early Stage of Plant Host Interaction: Study of an flgE Mutant. Curr. Microbiol. 2015, 70, 219–227. [Google Scholar] [CrossRef]
- Thompson, M.A.; Onyeziri, M.C.; Fuqua, C. Function and regulation of Agrobacterium tumefaciens cell surface structures that promote attachment. In Agrobacterium Biology: From Basic Science to Biotechnology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 143–184. [Google Scholar]
- Hwang, H.H.; Gelvin, S.B. Plant Proteins That Interact with VirB2, the Agrobacterium tumefaciens Pilin Protein, Mediate Plant Transformation. Plant Cell 2004, 16, 3148–3167. [Google Scholar] [CrossRef]
- Li, Y.G.; Christie, P.J. The Agrobacterium VirB/VirD4 T4SS: Mechanism and Architecture Defined Through In Vivo Mutagenesis and Chimeric Systems. Curr. Top. Microbiol. Immunol. 2018, 418, 233–260. [Google Scholar]
- Yu, M.; Wang, Y.C.; Huang, C.J.; Ma, L.S.; Lai, E.M. Agrobacterium tumefaciens Deploys a Versatile Antibacterial Strategy To Increase Its Competitiveness. J. Bacteriol. 2021, 203, e00490-20. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, B.; Citovsky, V. Genetic factors governing bacterial virulence and host plant susceptibility during Agrobacterium infection. Adv. Genet. 2022, 110, 1–29. [Google Scholar] [PubMed]
- Grohmann, E.; Christie, P.J.; Waksman, G.; Backert, S. Type IV secretion in Gram-negative and Gram-positive bacteria. Mol. Microbiol. 2018, 107, 455–471. [Google Scholar] [CrossRef] [PubMed]
- Mary, C.; Fouillen, A.; Bessette, B.; Nanci, A.; Baron, C. Interaction via the N terminus of the type IV secretion system (T4SS) protein VirB6 with VirB10 is required for VirB2 and VirB5 incorporation into T-pili and for T4SS function. J. Biol. Chem. 2018, 293, 13415–13426. [Google Scholar] [CrossRef]
- Gupta, R.; Gupta, N. Fundamentals of Bacterial Physiology and Metabolism; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Zhang, J.; Feng, T.; Wang, J.; Wang, Y.; Zhang, X.H. The Mechanisms and Applications of Quorum Sensing (QS) and Quorum Quenching (QQ). J. Ocean Univ. China 2019, 18, 1427–1442. [Google Scholar] [CrossRef]
- Dessaux, Y.; Faure, D. Quorum Sensing and Quorum Quenching in Agrobacterium: A “Go/No Go System”? Genes 2018, 9, 210. [Google Scholar] [CrossRef]
- Lin, W.; Li, D. Zinc and Zinc Transporters: Novel Regulators of Ventricular Myocardial Development. Pediatr. Cardiol. 2018, 39, 1042–1051. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Tian, N.; Chen, L.; Xie, S.; Hu, J.; Jin, Q.; Shao, C.; Huang, M.; Su, Q.; Huang, J.; et al. Transcriptomic Analysis of the Negative Effect of Epigallocatechin-3-Gallate from Tea Plant (Camellia sinensis) on Agrobacterium-Mediated Transformation Efficiency. Curr. Issues Mol. Biol. 2025, 47, 178. https://doi.org/10.3390/cimb47030178
Liu G, Tian N, Chen L, Xie S, Hu J, Jin Q, Shao C, Huang M, Su Q, Huang J, et al. Transcriptomic Analysis of the Negative Effect of Epigallocatechin-3-Gallate from Tea Plant (Camellia sinensis) on Agrobacterium-Mediated Transformation Efficiency. Current Issues in Molecular Biology. 2025; 47(3):178. https://doi.org/10.3390/cimb47030178
Chicago/Turabian StyleLiu, Guizhi, Na Tian, Lan Chen, Siyi Xie, Jinyu Hu, Qifang Jin, Chenyu Shao, Mengdi Huang, Qin Su, Jianan Huang, and et al. 2025. "Transcriptomic Analysis of the Negative Effect of Epigallocatechin-3-Gallate from Tea Plant (Camellia sinensis) on Agrobacterium-Mediated Transformation Efficiency" Current Issues in Molecular Biology 47, no. 3: 178. https://doi.org/10.3390/cimb47030178
APA StyleLiu, G., Tian, N., Chen, L., Xie, S., Hu, J., Jin, Q., Shao, C., Huang, M., Su, Q., Huang, J., Liu, Z., & Liu, S. (2025). Transcriptomic Analysis of the Negative Effect of Epigallocatechin-3-Gallate from Tea Plant (Camellia sinensis) on Agrobacterium-Mediated Transformation Efficiency. Current Issues in Molecular Biology, 47(3), 178. https://doi.org/10.3390/cimb47030178