
The Proliferation of Chang Liver Cells After Simulated Microgravity Induction

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and SMG Induction
2.2. Recovery After SMG Induction
2.3. WST-1 Assay
2.4. Cell Cycle Analysis
2.5. Flow Cytometry Analysis
2.6. Western Blot Analysis
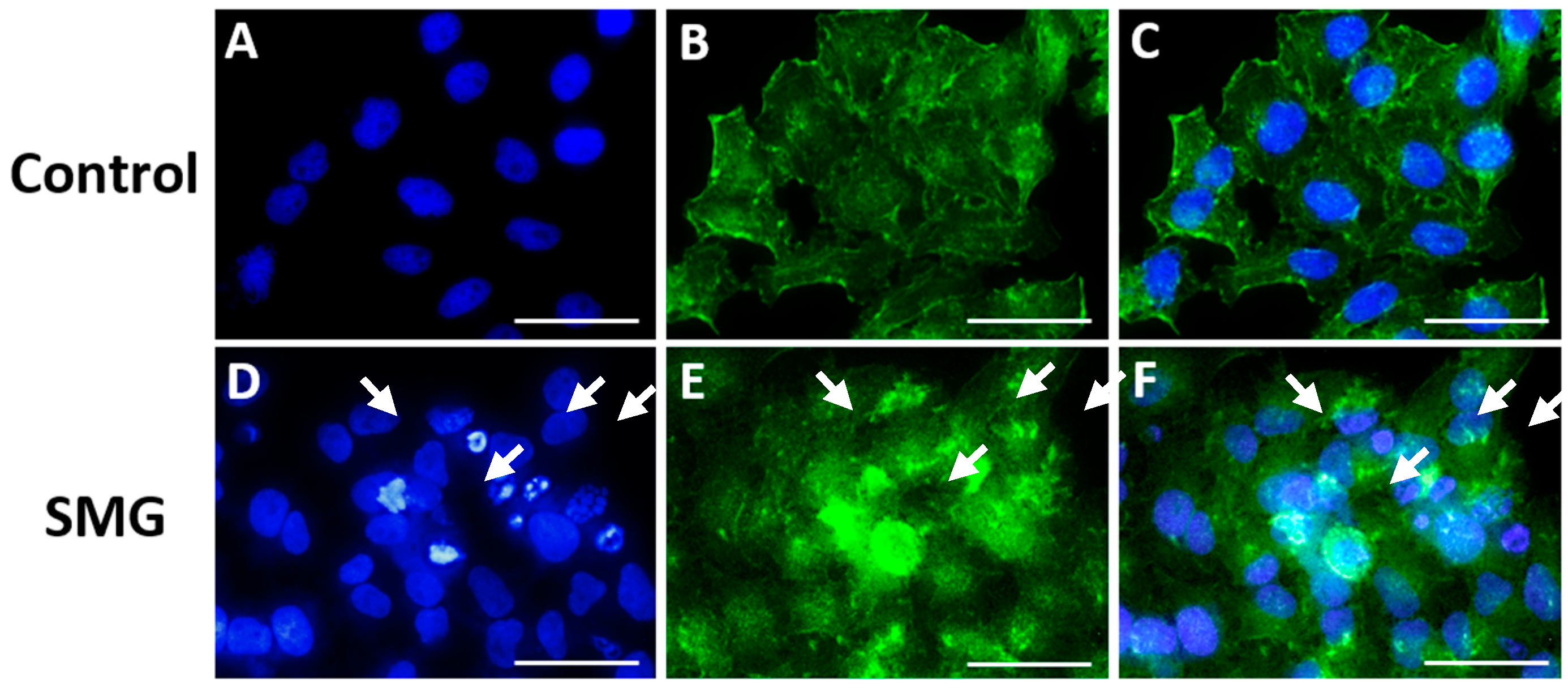
2.7. Microfilament Staining
2.8. Statistical Analysis
3. Results
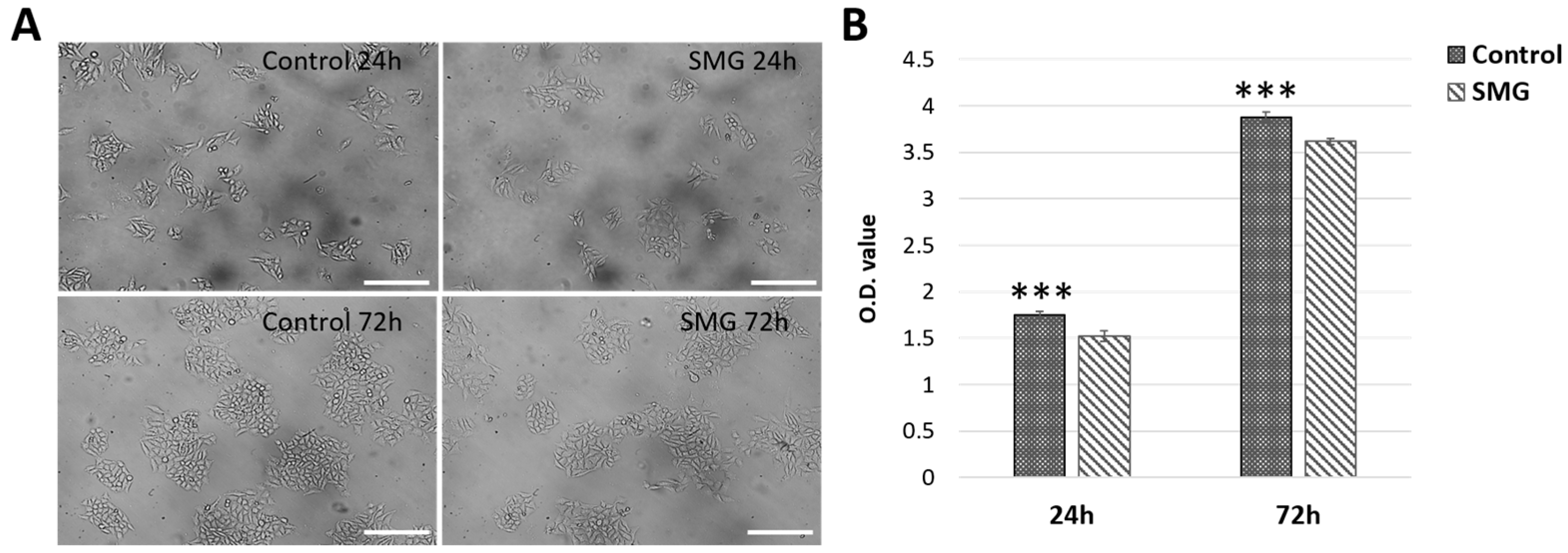
3.1. CCL-13 Cell Proliferation
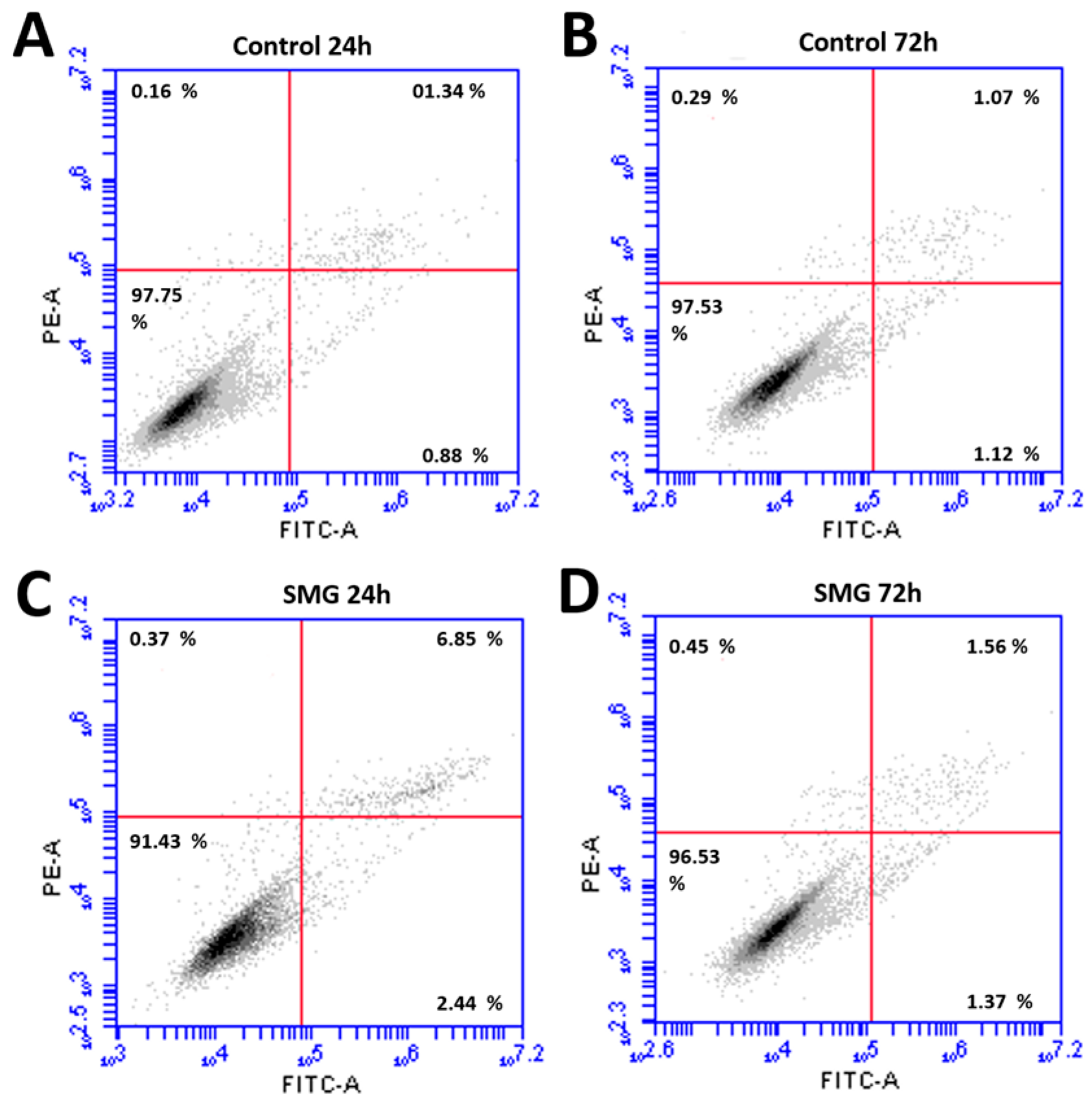
3.2. CCL-13 Cell Viability
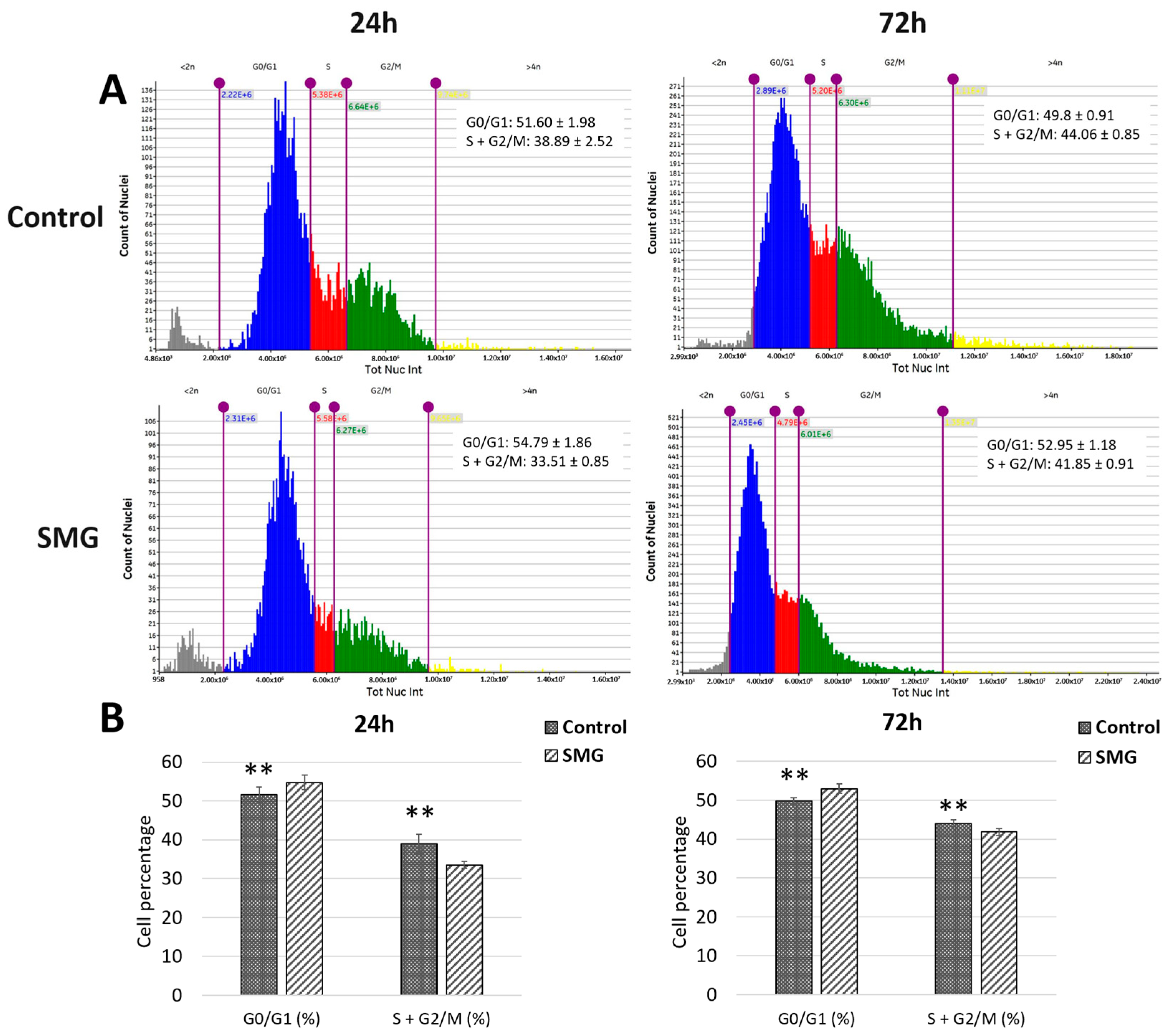
3.3. Cell Cycle Evaluation
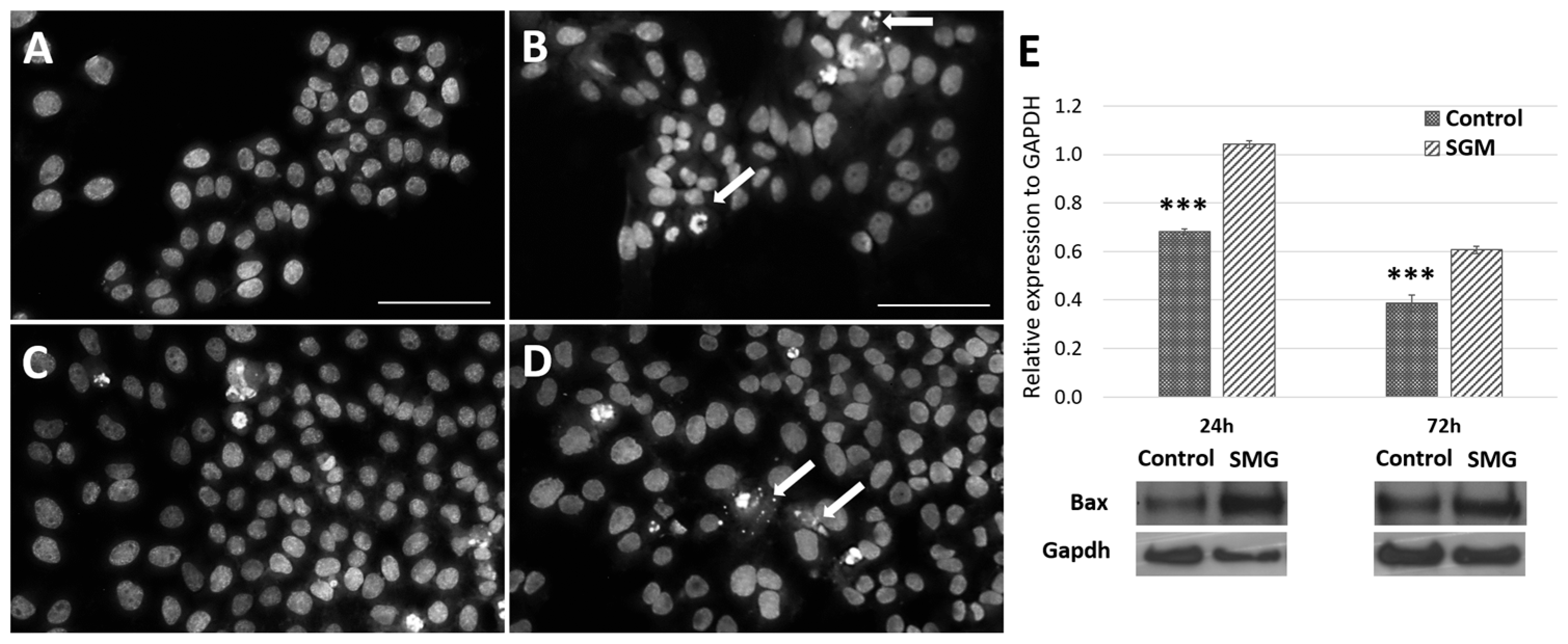
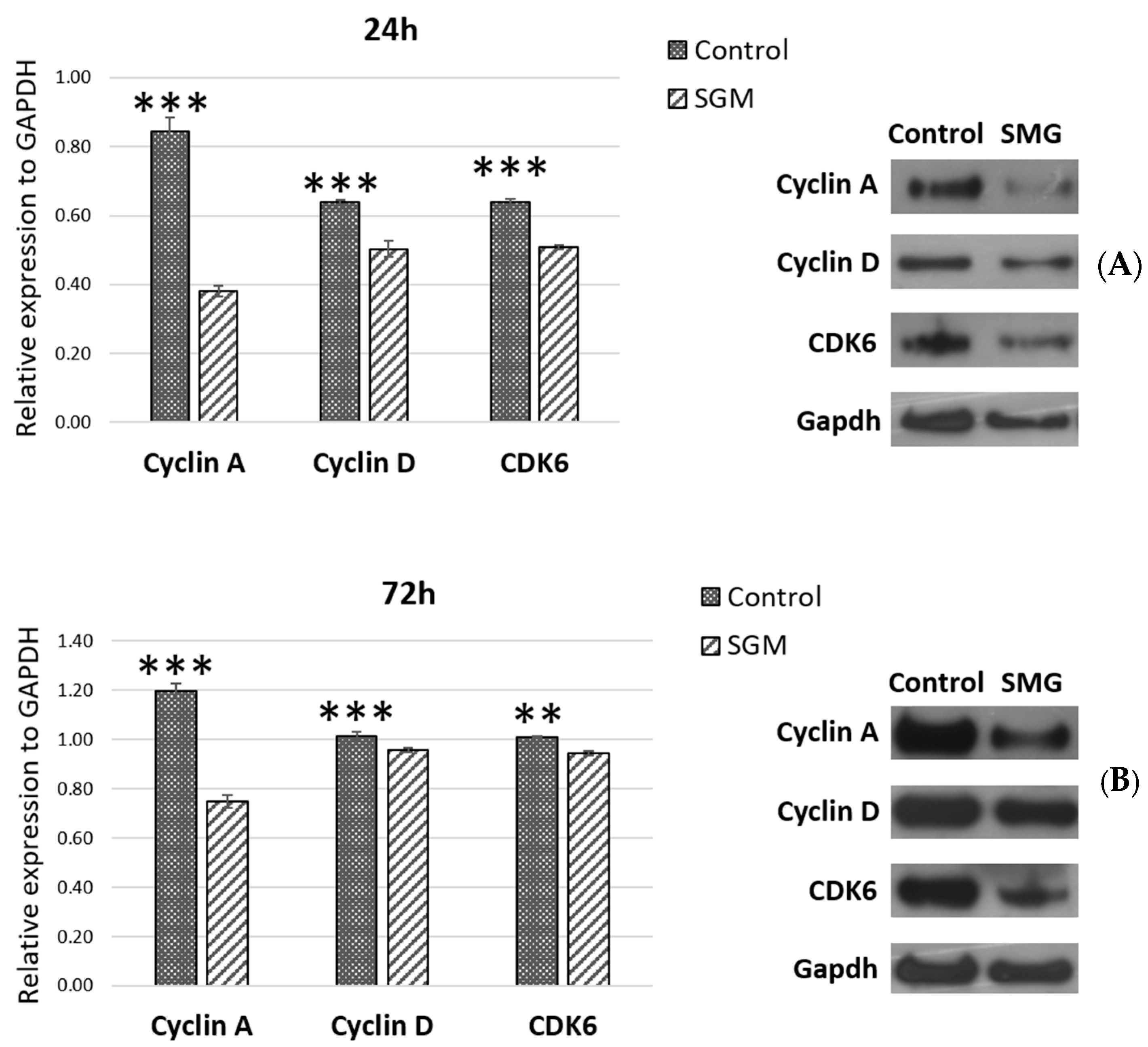
3.4. The Expression of Cell-Cycle-Related Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CDK | Cyclin-dependent kinases |
| ECL kit | Excellent Chemiluminescent Substrate Detection Kit |
| F-actin | Filamentous actin |
| FSC | Forward scatter |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| HRP | Horseradish peroxidase |
| O.D | Optical density |
| LDS | Lithium dodecyl sulfate |
| PBS | Phosphate buffer saline |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| SMG | Simulated microgravity |
| TBST | Tris-buffered saline, 0.1% Tween 20 |
References
- White, O.; Clément, G.; Fortrat, J.-O.; Pavy-LeTraon, A.; Thonnard, J.-L.; Blanc, S.; Wuyts, F.L.; Paloski, W.H. Towards Human Exploration of Space: The THESEUS Review Series on Neurophysiology Research Priorities. Npj Microgravity 2016, 2, 16023. [Google Scholar] [CrossRef]
- Crucian, B.; Simpson, R.J.; Mehta, S.; Stowe, R.; Chouker, A.; Hwang, S.-A.; Actor, J.K.; Salam, A.P.; Pierson, D.; Sams, C. Terrestrial Stress Analogs for Spaceflight Associated Immune System Dysregulation. Brain Behav. Immun. 2014, 39, 23–32. [Google Scholar] [CrossRef]
- Morey-Holton, E.R. The Impact of Gravity on Life. In Evolution on Planet Earth; Elsevier: Amsterdam, The Netherlands, 2003; pp. 143–159. ISBN 978-0-12-598655-7. [Google Scholar]
- Zong, B.; Wang, Y.; Wang, J.; Zhang, P.; Kan, G.; Li, M.; Feng, J.; Wang, Y.; Chen, X.; Jin, R.; et al. Effects of Long-term Simulated Microgravity on Liver Metabolism in Rhesus Macaques. FASEB J. 2022, 36, e22536. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; Van Loon, J.; Ulbrich, C.; Magnusson, N.E.; Infanger, M.; Bauer, J. Growing Tissues in Real and Simulated Microgravity: New Methods for Tissue Engineering. Tissue Eng. Part B Rev. 2014, 20, 555–566. [Google Scholar] [CrossRef]
- Choi, D.H.; Jeon, B.; Lim, M.H.; Lee, D.H.; Ye, S.-K.; Jeong, S.-Y.; Kim, S. 3D Cell Culture Using a Clinostat Reproduces Microgravity-Induced Skin Changes. npj Microgravity 2021, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Wubshet, N.H.; Arreguin-Martinez, E.; Nail, M.; Annamalai, H.; Koerner, R.; Rousseva, M.; Tom, T.; Gillespie, R.B.; Liu, A.P. Simulating Microgravity Using a Random Positioning Machine for Inducing Cellular Responses to Mechanotransduction in Human Osteoblasts. Rev. Sci. Instrum. 2021, 92, 114101. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Guo, J.; Wang, S.; Kang, L.; Deng, Y.; Li, Y. Simulated Microgravity Altered the Metabolism of Loureirin B and the Expression of Major Cytochrome P450 in Liver of Rats. Front. Pharmacol. 2018, 9, 1130. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy Metabolism in the Liver. In Comprehensive Physiology; Terjung, R., Ed.; Wiley: Hoboken, NJ, USA, 2014; pp. 177–197. ISBN 978-0-470-65071-4. [Google Scholar]
- Yang, T.; Li, C.; Zhang, L.; Li, M.; Zhou, P. A Promising Hepatocyte-Like Cell Line, CCL-13, Exhibits Good Liver Function Both In Vitro and in an Acute Liver Failure Model. Transplant. Proc. 2013, 45, 688–694. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Tran, P.H.; Kim, K.-S.; Yang, S.-G. The Effects of Real and Simulated Microgravity on Cellular Mitochondrial Function. npj Microgravity 2021, 7, 44. [Google Scholar] [CrossRef]
- Chung, J.H.; Ahn, C.B.; Son, K.H.; Yi, E.; Son, H.S.; Kim, H.-S.; Lee, S.H. Simulated Microgravity Effects on Nonsmall Cell Lung Cancer Cell Proliferation and Migration. Aerosp. Med. Hum. Perform. 2017, 88, 82–89. [Google Scholar] [CrossRef]
- Martinelli, L.K.; Russomano, T.; Dos Santos, M.A.; Falcao, F.P.; Bauer, M.E.; Machado, A.; Sundaresan, A. Effect of Microgravity on Immune Cell Viability and Proliferation. IEEE Eng. Med. Biol. Mag. 2009, 28, 85–90. [Google Scholar] [CrossRef]
- Blaber, E.A.; Finkelstein, H.; Dvorochkin, N.; Sato, K.Y.; Yousuf, R.; Burns, B.P.; Globus, R.K.; Almeida, E.A.C. Microgravity Reduces the Differentiation and Regenerative Potential of Embryonic Stem Cells. Stem Cells Dev. 2015, 24, 2605–2621. [Google Scholar] [CrossRef]
- Plett, P.A.; Abonour, R.; Frankovitz, S.M.; Orschell, C.M. Impact of Modeled Microgravity on Migration, Differentiation, and Cell Cycle of Primitive Human Hematopoietic Progenitor Cells. Exp. Hematol. 2004, 32, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, C.; Chen, J.; Hong, F.; Chen, P.; Wang, J. Effects of Simulated Microgravity on the Expression Profiles of RNA during Osteogenic Differentiation of Human Bone Marrow Mesenchymal Stem Cells. Cell Prolif. 2019, 52, e12539. [Google Scholar] [CrossRef]
- Dai, T.X.; Son, H.N.; Chi, H.N.Q.; Huy, H.N.Q.; Minh, N.T.; Tram, N.T.T.; Huyen, N.T.T.; Quan, T.M.; Chung, D.C.; Nhung, T.H.; et al. Simulated Microgravity Induces the Proliferative Inhibition and Morphological Changes in Porcine Granulosa Cells. Curr. Issues Mol. Biol. 2021, 43, 2210–2219. [Google Scholar] [CrossRef] [PubMed]
- Tran, M.T.; Ho, C.N.Q.; Hoang, S.N.; Doan, C.C.; Nguyen, M.T.; Van, H.D.; Ly, C.N.; Le, C.P.M.; Hoang, H.N.Q.; Nguyen, H.T.M.; et al. Morphological Changes of 3T3 Cells under Simulated Microgravity. Cells 2024, 13, 344. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.N.Q.; Tran, M.T.; Doan, C.C.; Hoang, S.N.; Tran, D.H.; Le, L.T. Simulated Microgravity Inhibits the Proliferation of Chang Liver Cells by Attenuation of the Major Cell Cycle Regulators and Cytoskeletal Proteins. Int. J. Mol. Sci. 2021, 22, 4550. [Google Scholar] [CrossRef]
- Deng, B.; Liu, R.; Tian, X.; Han, Z.; Chen, J. Simulated Microgravity Inhibits the Viability and Migration of Glioma via FAK/RhoA/Rock and FAK/Nek2 Signaling. In Vitro Cell. Dev. Biol.-Anim. 2019, 55, 260–271. [Google Scholar] [CrossRef]
- Sherr, C.J.; Roberts, J.M. Living with or without Cyclins and Cyclin-Dependent Kinases. Genes Dev. 2004, 18, 2699–2711. [Google Scholar] [CrossRef] [PubMed]
- Nebenfuehr, S.; Kollmann, K.; Sexl, V. The Role of CDK6 in Cancer. Intl J. Cancer 2020, 147, 2988–2995. [Google Scholar] [CrossRef] [PubMed]
- Sherr, C.J. D-Type Cyclins. Trends Biochem. Sci. 1995, 20, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Hitomi, M.; Stacey, D.W. Variations in Cyclin D1 Levels through the Cell Cycle Determine the Proliferative Fate of a Cell. Cell Div. 2006, 1, 32. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.O. CYCLIN-DEPENDENT KINASES: Engines, Clocks, and Microprocessors. Annu. Rev. Cell Dev. Biol. 1997, 13, 261–291. [Google Scholar] [CrossRef] [PubMed]
- Arsic, N.; Bendris, N.; Peter, M.; Begon-Pescia, C.; Rebouissou, C.; Gadéa, G.; Bouquier, N.; Bibeau, F.; Lemmers, B.; Blanchard, J.M. A Novel Function for Cyclin A2: Control of Cell Invasion via RhoA Signaling. J. Cell Biol. 2012, 196, 147–162. [Google Scholar] [CrossRef]
- Loukil, A. Cyclin A2: At the Crossroads of Cell Cycle and Cell Invasion. WJBC 2015, 6, 346. [Google Scholar] [CrossRef]
- Pagano, M.; Pepperkok, R.; Verde, F.; Ansorge, W.; Draetta, G. Cyclin A Is Required at Two Points in the Human Cell Cycle. EMBO J. 1992, 11, 961–971. [Google Scholar] [CrossRef]
- Chibazakura, T.; Kamachi, K.; Ohara, M.; Tane, S.; Yoshikawa, H.; Roberts, J.M. Cyclin A Promotes S-Phase Entry via Interaction with the Replication Licensing Factor Mcm7. Mol. Cell. Biol. 2011, 31, 248–255. [Google Scholar] [CrossRef]
- Dang, B.; Yang, Y.; Zhang, E.; Li, W.; Mi, X.; Meng, Y.; Yan, S.; Wang, Z.; Wei, W.; Shao, C.; et al. Simulated Microgravity Increases Heavy Ion Radiation-Induced Apoptosis in Human B Lymphoblasts. Life Sci. 2014, 97, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Gridley, D.S.; Mao, X.W.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Moldovan, M.; Cunningham, C.E.; Jones, T.A.; Slater, J.M.; Pecaut, M.J. Changes in Mouse Thymus and Spleen after Return from the STS-135 Mission in Space. PLoS ONE 2013, 8, e75097. [Google Scholar] [CrossRef] [PubMed]
- Radugina, E.A.; Almeida, E.A.C.; Blaber, E.; Poplinskaya, V.A.; Markitantova, Y.V.; Grigoryan, E.N. Exposure to Microgravity for 30 Days Onboard Bion M1 Caused Muscle Atrophy and Impaired Regeneration in Murine Femoral Quadriceps. Life Sci. Space Res. 2018, 16, 18–25. [Google Scholar] [CrossRef]
- Ellis, H. Genetic Control of Programmed Cell Death in the Nematode C. Elegans. Cell 1986, 44, 817–829. [Google Scholar] [CrossRef]
- Wang, J.Y. Cellular Responses to DNA Damage. Curr. Opin. Cell Biol. 1998, 10, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Oltval, Z.N.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 Heterodimerizes in Vivo with a Conserved Homolog, Bax, That Accelerates Programed Cell Death. Cell 1993, 74, 609–619. [Google Scholar] [CrossRef]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight Alters Microtubules and Increases Apoptosis in Human Lymphocytes (Jurkat). FASEB J. 1998, 12, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Hughes-Fulford, M. Function of the Cytoskeleton in Gravisensing during Spaceflight. Adv. Space Res. 2003, 32, 1585–1593. [Google Scholar] [CrossRef]
- Sager, P.R.; Syversen, T.L.M.; Clarkson, T.W.; Cavanagh, J.B.; Elgsaeter, A.; Guldberg, H.C.; Lee, S.D.; Lichtman, M.A.; Mottet, N.K.; Olmsted, J.B. Structure and Function of the Cytoskeleton. In The Cytoskeleton; Clarkson, T.W., Sager, P.R., Syversen, T.L.M., Eds.; Springer: Boston, MA, USA, 1986; pp. 3–21. ISBN 978-1-4612-9274-6. [Google Scholar]
- Nakaseko, Y.; Yanagida, M. Cytoskeleton in the Cell Cycle. Nature 2001, 412, 291–292. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Qin, Y.; Kun, L.; Zhou, Y. The Significant Role of the Microfilament System in Tumors. Front. Oncol. 2021, 11, 620390. [Google Scholar] [CrossRef]
- Desouza, M.; Gunning, P.W.; Stehn, J.R. The Actin Cytoskeleton as a Sensor and Mediator of Apoptosis. BioArchitecture 2012, 2, 75–87. [Google Scholar] [CrossRef]
- Gourlay, C.W.; Ayscough, K.R. The Actin Cytoskeleton: A Key Regulator of Apoptosis and Ageing? Nat. Rev. Mol. Cell Biol. 2005, 6, 583–589. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, G.-L.; Vedantam, S.; Li, P.; Field, J. Mitochondrial Shuttling of CAP1 Promotes Actin- and Cofilin-Dependent Apoptosis. J. Cell Sci. 2008, 121, 2913–2920. [Google Scholar] [CrossRef]
- Rehklau, K.; Gurniak, C.B.; Conrad, M.; Friauf, E.; Ott, M.; Rust, M.B. ADF/Cofilin Proteins Translocate to Mitochondria during Apoptosis but Are Not Generally Required for Cell Death Signaling. Cell Death Differ. 2012, 19, 958–967. [Google Scholar] [CrossRef]
- King, V.L.; Campellone, K.G. F-Actin–Rich Territories Coordinate Apoptosome Assembly and Caspase Activation during DNA Damage–Induced Intrinsic Apoptosis. MBoC 2023, 34, ar41. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoang, H.N.Q.; Ho, C.N.Q.; Dang, L.T.T.; Phan, N.L.C.; Doan, C.C.; Nguyen, H.T.M.; Le, C.P.M.; Hoang, S.N.; Le, L.T. The Proliferation of Chang Liver Cells After Simulated Microgravity Induction. Curr. Issues Mol. Biol. 2025, 47, 164. https://doi.org/10.3390/cimb47030164
Hoang HNQ, Ho CNQ, Dang LTT, Phan NLC, Doan CC, Nguyen HTM, Le CPM, Hoang SN, Le LT. The Proliferation of Chang Liver Cells After Simulated Microgravity Induction. Current Issues in Molecular Biology. 2025; 47(3):164. https://doi.org/10.3390/cimb47030164
Chicago/Turabian StyleHoang, Huy Nghia Quang, Chi Nguyen Quynh Ho, Loan Thi Tung Dang, Nhan Lu Chinh Phan, Chung Chinh Doan, Han Thai Minh Nguyen, Cuong Phan Minh Le, Son Nghia Hoang, and Long Thanh Le. 2025. "The Proliferation of Chang Liver Cells After Simulated Microgravity Induction" Current Issues in Molecular Biology 47, no. 3: 164. https://doi.org/10.3390/cimb47030164
APA StyleHoang, H. N. Q., Ho, C. N. Q., Dang, L. T. T., Phan, N. L. C., Doan, C. C., Nguyen, H. T. M., Le, C. P. M., Hoang, S. N., & Le, L. T. (2025). The Proliferation of Chang Liver Cells After Simulated Microgravity Induction. Current Issues in Molecular Biology, 47(3), 164. https://doi.org/10.3390/cimb47030164