
Proteome and Peptidome Changes and Zn Concentration in Chicken after In Ovo Stimulation with a Multi-Strain Probiotic and Zn-Gly Chelate: Preliminary Research

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Incubation Period
2.3. In Ovo Inoculations
2.4. Birds and Housing
2.5. Blood and Tissue Sample Collection
2.6. Determination of the Zinc Concentration in Serum and Tissues by Flame Atomic Absorption Spectrometry (FAAS)
2.7. MALDI—TOF MS
2.8. SDS-PAGE Analysis
2.9. Statistical Analysis
3. Results
3.1. Zn Concentration in the Serum, Liver, Small Intestine, and Yolk Sac
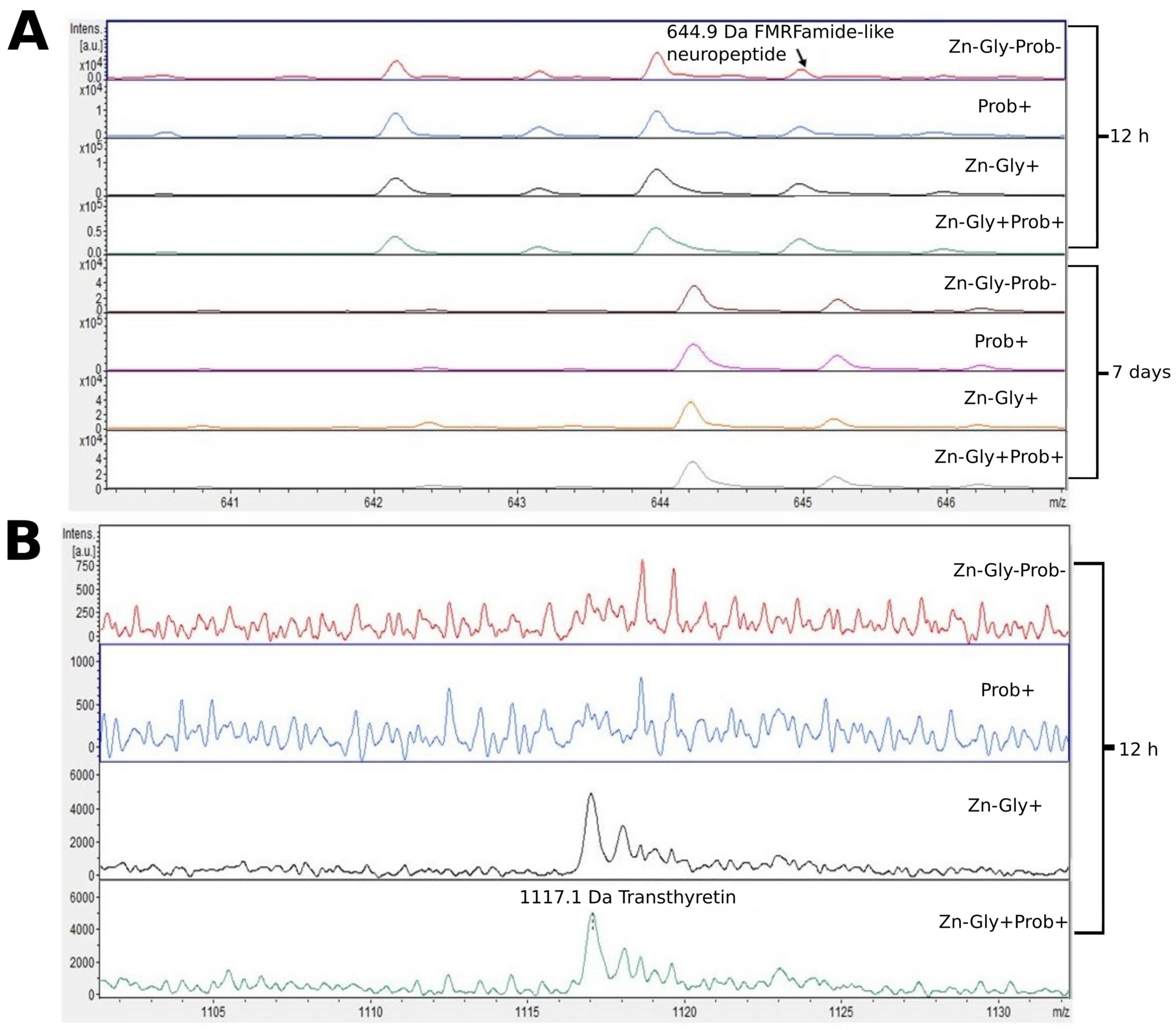
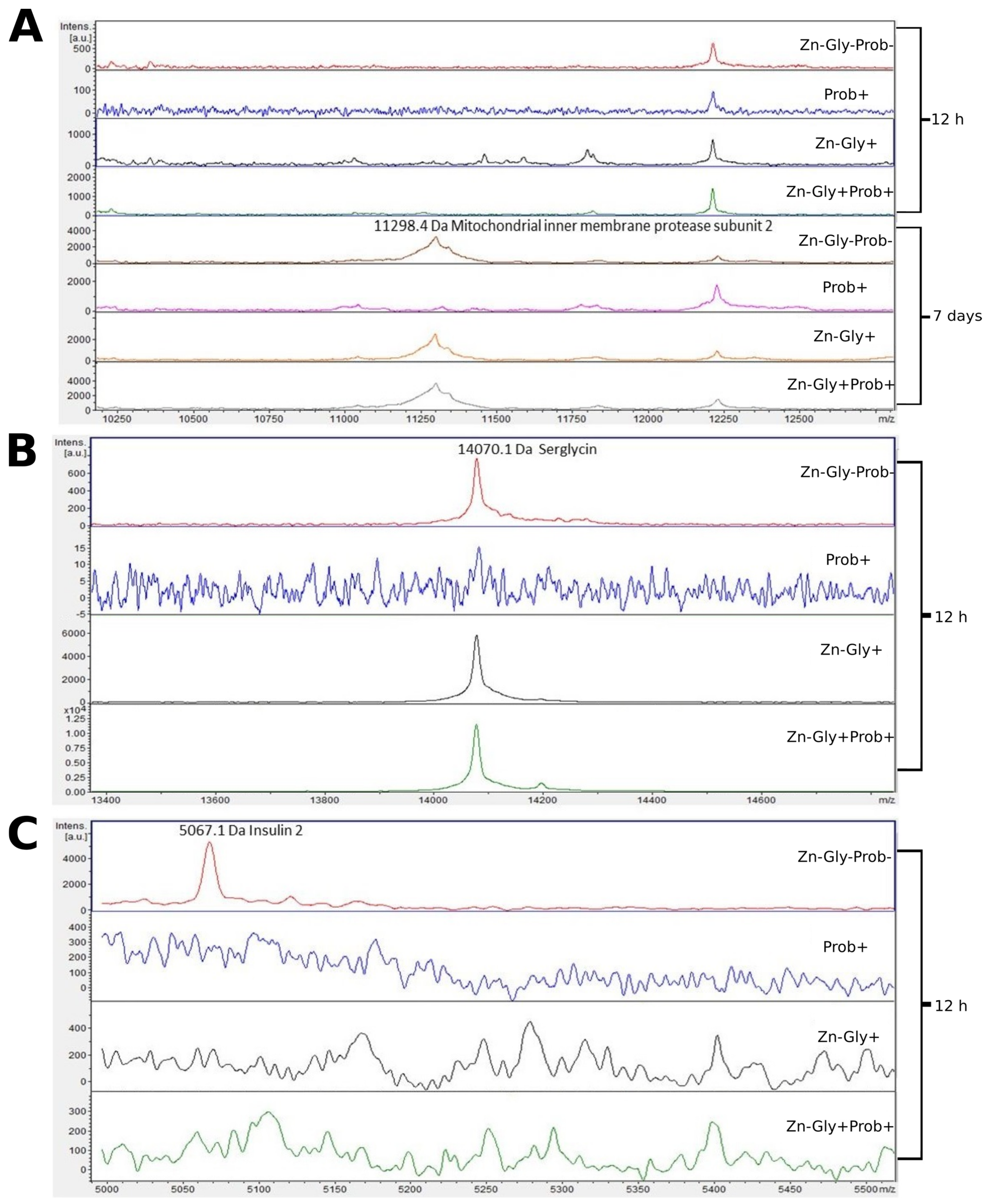
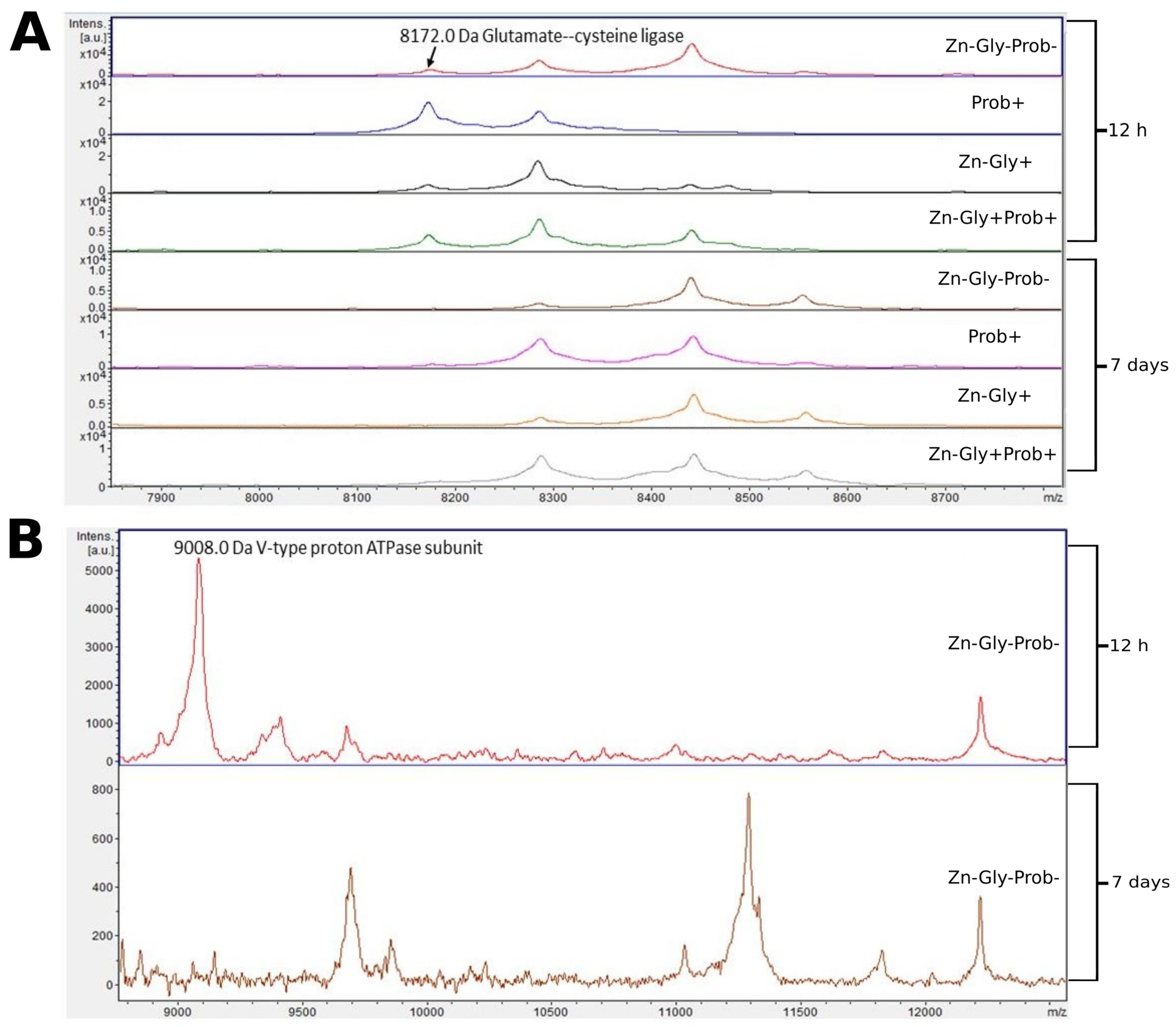
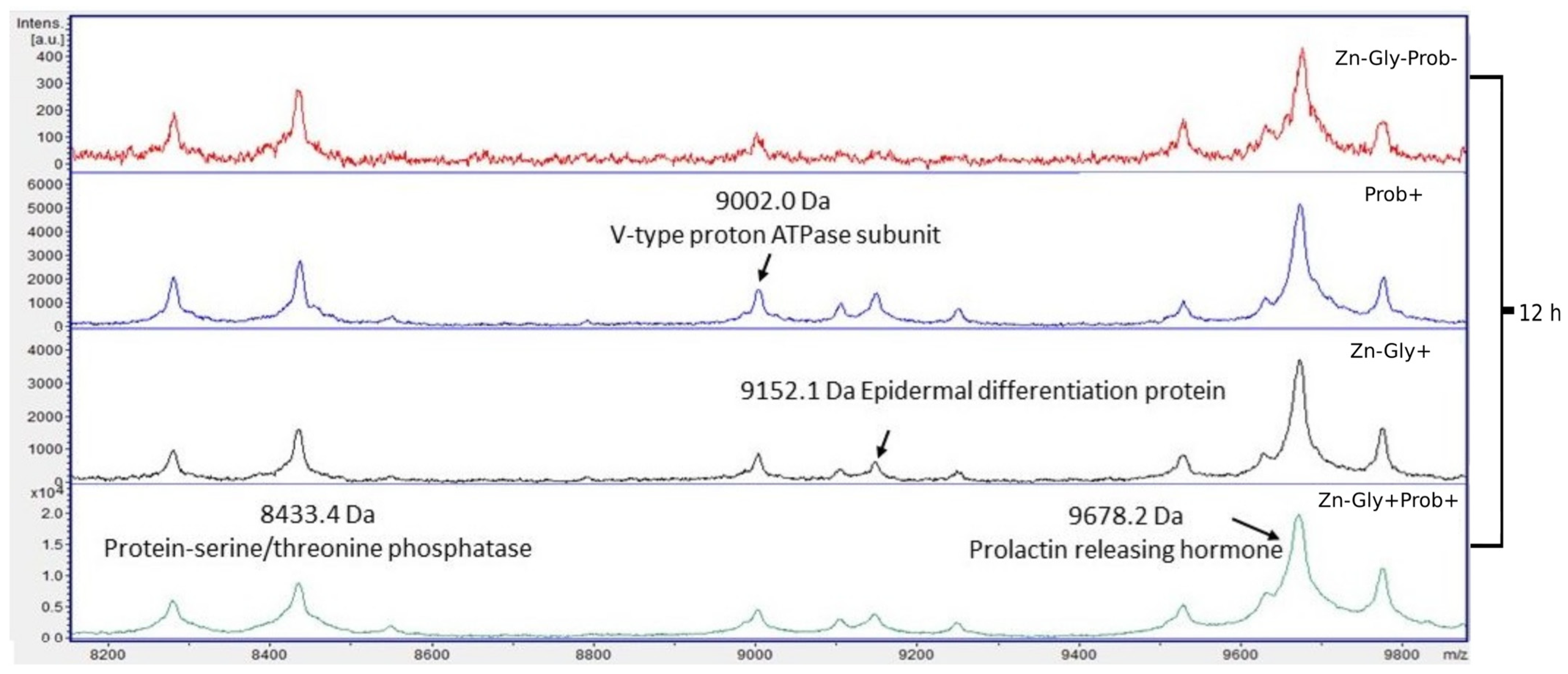
3.2. Peptide Profile Analysis
3.3. Protein Mass Distribution Visualized by SDS-PAGE
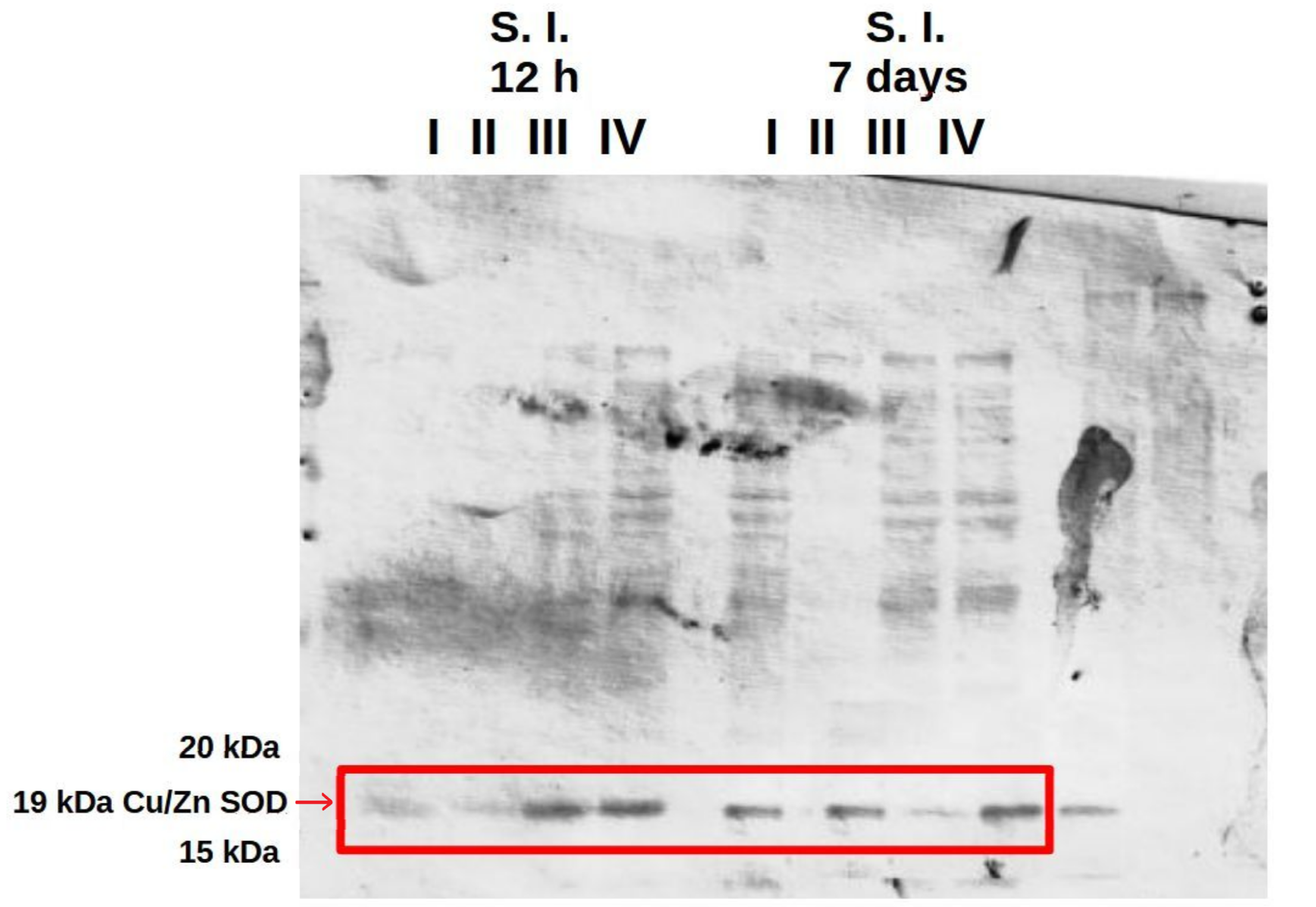
3.4. Analysis of the Presence of Superoxide Dismutase (SOD) in the Liver and in the Small and Large Intestine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Olejnik, K.; Popiela, E.; Opaliński, S. Emerging precision management methods in poultry sector. Agriculture 2022, 12, 718. [Google Scholar] [CrossRef]
- Gržinić, G.; Piotrowicz-Cieślak, A.; Klimkowicz-Pawlas, A.; Górny, R.L.; Ławniczek-Wałczyk, A.; Piechowicz, L.; Olkowska, E.; Potrykus, M.; Tankiewicz, M.; Krupka, M.; et al. Intensive poultry farming: A review of the impact on the environment and human health. Sci. Total Environ. 2023, 858, 160014. [Google Scholar] [CrossRef]
- Mesquita, M.A.; Araújo, I.C.S.; Café, M.B.; Arnhold, E.; Mascarenhas, A.G.; Carvalho, F.B.; Stringhini, J.H.; Leandro, N.S.M.; Gonzales, E. Results of hatching and rearing broiler chickens in different incubation systems. Poult. Sci. 2021, 100, 94–102. [Google Scholar] [CrossRef]
- Yalcin, S.; Özkan, S.; Shah, T. Incubation temperature and lighting: Effect on embryonic development, post-hatch growth, and adaptive response. Front. Physiol. 2022, 13, 899977. [Google Scholar] [CrossRef]
- Adedokun, S.A.; Olojede, O.C. Optimizing gastrointestinal integrity in poultry: The role of nutrients and feed additives. Front. Vet. Sci. 2019, 5, 348. [Google Scholar] [CrossRef]
- Jha, R.; Singh, A.K.; Yadav, S.; Berrocoso, J.F.D.; Mishra, B. Early nutrition programming (in ovo and post-hatch feeding) as a strategy to modulate gut health of poultry. Front. Vet. Sci. 2019, 6, 82. [Google Scholar] [CrossRef]
- Peebles, E.D. In ovo applications in poultry: A review. Poult. Sci. 2018, 97, 2322–2338. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.F.B.; Bertechini, A.G.; Bricka, R.M.; Kim, E.J.; Gerard, P.D.; Peebles, E.D. Effects of in ovo injection of organic zinc, manganese, and copper on the hatchability and bone parameters of broiler hatchlings. Poult. Sci. 2015, 94, 2488–2494. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (direct-fed microbials) in poultry nutrition and their effects on nutrient utilization, growth and laying performance, and gut health: A systematic review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef] [PubMed]
- Oladokun, S.; Adewole, D.I. In ovo delivery of bioactive substances: An alternative to the use of antibiotic growth promoters in poultry production—A review. J. Appl. Poult. Res. 2020, 29, 744–763. [Google Scholar] [CrossRef]
- Ciszewski, A.; Jarosz, Ł.; Marek, A.; Michalak, K.; Grądzki, Z.; Kaczmarek, B.; Rysiak, A. Effect of combined in ovo administration of zinc glycine chelate (Zn-Gly) and a multistrain probiotic on the modulation of cellular and humoral immune responses in broiler chickens. Poult. Sci. 2023, 102, 102823. [Google Scholar] [CrossRef]
- Ezzulddin, T.A.; Jwher, D.M. Beneficial microorganisms in animal production and environment. J. Appl. Vet. Sci. 2022, 7, 64–71. [Google Scholar] [CrossRef]
- Jwher, D.M.T.; Abd, S.K.; Mohammad, A.G. The study of using effective microorganisms (EM) on health and performance of broiler chicks. Iraqi J. Vet. Sci. 2013, 27, 73–78. [Google Scholar] [CrossRef]
- Esatu, W.; Melesse, A.; Dessie, T. Effect of effective microorganisms on growth parameters and serum cholesterol levels in broilers. Afr. J. Agric. Res. 2011, 6, 3841–3846. [Google Scholar] [CrossRef]
- Reszka, P.; Dunislawska, A.; Slawinska, A.; Siwek, M.; Kapelański, W.; Bogucka, J. Influence of the effective microorganisms (EM) on performance, intestinal morphology and gene expression in the jejunal mucosa of pigs fed different diets. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1444–1453. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F. Programming of host metabolism by the gut microbiota. Ann. Nutr. Metab. 2011, 58, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.L.; Lu, L.; Li, S.F.; Luo, X.G.; Liu, B. Relative bioavailabilities of organic zinc sources with different chelation strengths for broilers fed a conventional corn-soybean meal diet. J. Anim. Sci. 2009, 87, 2038–2046. [Google Scholar] [CrossRef]
- Salim, H.M.; Lee, H.R.; Jo, C.; Lee, S.K.; Lee, B.D. Effect of sources and levels of zinc on the tissue mineral concentration and carcass quality of broilers. Avian Biol. Res. 2010, 3, 23–29. [Google Scholar] [CrossRef]
- Jarosz, Ł.; Marek, A.; Grądzki, Z.; Kwiecień, M.; Kalinowski, M. The effect of feed supplementation with zinc chelate and zinc sulphate on selected humoral and cell-mediated immune parameters and cytokine concentration in broiler chickens. Res. Vet. Sci. 2017, 112, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, Ł.; Marek, A.; Grądzki, Z.; Laskowska, E.; Kwiecień, M. Effect of zinc sulfate and zinc glycine chelate on concentrations of acute phase proteins in chicken serum and liver tissue. Biol. Trace Elem. Res. 2019, 187, 258–272. [Google Scholar] [CrossRef]
- Jarosz, Ł.; Marek, A.; Grądzki, Z.; Kwiecień, M.; Żylinska, B.; Kaczmarek, B. Effect of feed supplementation with zinc glycine chelate and zinc sulfate on cytokine and immunoglobulin gene expression profiles in chicken intestinal tissue. Poult. Sci. 2017, 96, 4224–4235. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J. Physiol. Sci. 2018, 68, 19–31. [Google Scholar] [CrossRef]
- Adisa, A.O.; Odutuga, A.A. Metabolic interactions between zinc and essential fatty acids in the mammalian organism. Nutr. Food Sci. 1999, 99, 99–104. [Google Scholar] [CrossRef]
- Winiarska-Mieczan, A.; Jachimowicz, K.; Kwiecień, M.; Kislova, S.; Baranowska-Wójcik, E.; Zasadna, Z.; Yanovych, D.; Kowalczuk-Vasilev, E. The impact of Zn, Cu and Fe chelates on the fatty-acid profile and dietary value of broiler-chicken thigh meat. Animals 2021, 11, 3115. [Google Scholar] [CrossRef]
- Kim, H.J.; Kang, H.K. Effects of in ovo injection of zinc or diet supplementation of zinc on performance, serum biochemical profiles, and meat quality in broilers. Animals 2022, 12, 630. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, L.; Ding, X.; Wang, Y.; Xu, Y.; Li, D.; Liang, S.; Wang, Y.; Zhang, L.; Fu, A.; et al. Mechanisms underlying the protective effect of maternal zinc (ZnSO4 or Zn-Gly) against heat stress-induced oxidative stress in chicken embryo. Antioxidants 2022, 11, 1699. [Google Scholar] [CrossRef] [PubMed]
- Hatab, M.H.; Rashad, E.; Saleh, H.M.; El-Sayed, E.R.; Taleb, A.M.A. Effects of dietary supplementation of myco-fabricated zinc oxide nanoparticles on performance, histological changes, and tissues Zn concentration in broiler chicks. Sci. Rep. 2022, 12, 18791. [Google Scholar] [CrossRef]
- Rubio, L.A. Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 2019, 98, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Varvara, R.A.; Vodnar, D.C. Probiotic-driven advancement: Exploring the intricacies of mineral absorption in the human body. Food Chem. X 2023, 21, 101067. [Google Scholar] [CrossRef] [PubMed]
- Bielik, V.; Kolisek, M. Bioaccessibility and bioavailability of minerals in relation to a healthy gut microbiome. Int. J. Mol. Sci. 2021, 22, 6803. [Google Scholar] [CrossRef]
- Malyar, R.M.; Li, H.; Liu, D.; Abdulrahim, Y.; Farid, R.A.; Gan, F.; Ali, W.; Enayatullah, H.; Banuree, S.A.H.; Huang, K.; et al. Selenium/Zinc-enriched probiotics improve serum enzyme activity, antioxidant ability, inflammatory factors and related gene expression of Wistar rats inflated under heat stress. Life Sci. 2020, 248, 117464. [Google Scholar] [CrossRef]
- De Palmieri, L.C.; Lima, L.M.; Freire, J.B.; Bleicher, L.; Polikarpov, I.; Almeida, F.C.; Foguel, D. Novel Zn2+-binding sites in human transthyretin: Implications for amyloidogenesis and retinol-binding protein recognition. J. Biol. Chem. 2010, 285, 31731–31741. [Google Scholar] [CrossRef]
- Martinez, M.M.; Hill, G.M.; Link, J.E.; Raney, N.E.; Tempelman, R.J.; Ernst, C.W. Pharmacological zinc and phytase supplementation enhance metallothionein mRNA abundance and protein concentration in newly weaned pigs. J. Nutr. 2004, 134, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Bondzio, A.; Pieper, R.; Gabler, C.; Weise, C.; Schulze, P.; Zentek, J.; Einspanier, R. Feeding low or pharmacological concentrations of zinc oxide changes the hepatic proteome profiles in weaned piglets. PLoS ONE 2013, 8, e81202. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Montemayor, M.M.; Hill, G.M.; Raney, N.E.; Rilington, V.D.; Tempelman, R.J.; Link, J.E.; Wilkinson, C.P.; Ramos, A.M.; Ernst, C.W. Gene expression profiling in hepatic tissue of newly weaned pigs fed pharmacological zinc and phytase supplemented diets. BMC Genom. 2008, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Bavananthasivam, J.; Shojadoost, B.; Astill, J.; Taha-Abdelaziz, K.; Alqazlan, N.; Boodhoo, N.; Doost, J.S.; Sharif, S. In ovo and oral administration of probiotic Lactobacilli modulate cell- and antibody mediated immune responses in newly hatched chicks. Front. Immunol. 2021, 12, 664387. [Google Scholar] [CrossRef] [PubMed]
- AVMA Guidelines for the Euthanasia of Animals: 2020 Edition. Available online: https://www.avma.org/resources-tools/avma-policies/avma-guidelines-euthanasia-animals (accessed on 24 February 2020).
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Radi, A.M.; Abdel Azeem, N.M.; EL-Nahass, E.S. Comparative effects of zinc oxide and zinc oxide nanoparticle as feed additives on growth, feed choice test, tissue residues, and histopathological changes in broiler chickens. Environ. Sci. Pollut. Res. 2021, 28, 5158–5167. [Google Scholar] [CrossRef] [PubMed]
- Ramiah, S.K.; Awad, E.A.; Mookiah, S.; Idrus, Z. Effects of zinc oxide nanoparticles on growth performance and concentrations of malondialdehyde, zinc in tissues, and corticosterone in broiler chickens under heat stress conditions. Poult. Sci. 2019, 98, 3828–3838. [Google Scholar] [CrossRef]
- Mohan, J.; Ali, S.A.; Suvartan, R.; Kapila, S.; Sharma, R.; Tomar, S.K.; Behare, P.; Yadav, H. Bioavailability of biotransformed zinc enriched dahi in wistar rats. Int. J. Probiotics Prebiotics 2018, 13, 45–54. [Google Scholar] [PubMed]
- Mrvcic, J.; Prebeg, T.; Barisic, L.; Stanzer, D.; Bacun-Druzina, V.; Stehlik-Tomas, V. Zinc binding by lactic acid bacteria. Food Sci. Biotechnol. 2009, 47, 381–388. [Google Scholar]
- Świątkiewicz, S.; Koreleski, J.; Zhong, D.Q. The bioavailability of zinc from inorganic and organic sources in broiler chickens as affected by addition of phytase. J. Anim. Feed Sci. 2001, 10, 317–328. [Google Scholar] [CrossRef]
- Yi, Z.; Kornegay, E.T.; Denbow, D.M. Supplemental microbial phytase improves zinc utilization in broilers. Poult. Sci. 1996, 75, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Thiel, U.; Weigand, E. Influence of dietary zinc and microbial phytase supplementation on zinc retention and zinc excretion in broiler chicks. In Proceedings of the XIX World’s Poultry Congress, Amsterdam, The Netherlands, 20–24 September 1992. [Google Scholar]
- Augspurger, N.R.; Spencer, J.D.; Webel, D.M.; Baker, D.H. Pharmacological zinc levels reduce the phosphorus-releasing efficacy of phytase in young pigs and chickens. J. Anim. Sci. 2004, 82, 1732–1739. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Gong, Y.J.; Zhan, H.Q.; Zheng, Y.X.; Zou, X.T. Effects of dietary Zn-methionine supplementation on the laying performance, egg quality, antioxidant capacity, and serum parameters of laying hens. Poult. Sci. 2019, 98, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Lu, L.; Liao, X.; Zhang, L.; Lin, X.; Luo, X.; Ma, Q. Effect of in ovo zinc injection on the embryonic development and epigenetics-related indices of zinc-deprived broiler breeder eggs. Biol. Trace. Elem. Res. 2018, 185, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Lu, L.; Wang, R.L.; Lei, H.L.; Li, S.F.; Zhang, L.Y.; Luo, X.G. Effects of supplemental zinc source and level on antioxidant ability and fat metabolism-related enzymes of broilers. Poult. Sci. 2015, 94, 2686–2694. [Google Scholar] [CrossRef]
- Wang, M.Q.; Tao, W.J.; Ye, S.S.; Du, Y.J.; Wang, C.; Shen, S.X. Effects of dietary pharmacological zinc on growth, liver metallothionein, Cu, Zn-SOD concentration and serum parameters in piglets. J. Anim. Vet. Adv. 2012, 11, 1390–1394. [Google Scholar] [CrossRef]
- Lazar, I., Jr.; Lazar, I., Sr. Gel Analyzer 19.1. Available online: www.gelanalyzer.com (accessed on 1 December 2022).
- Tang, D.; Wu, J.; Jiao, H.; Wang, X.; Zhao, J.; Lin, H. The development of antioxidant system in the intestinal tract of broiler chickens. Poult. Sci. 2019, 98, 664–678. [Google Scholar] [CrossRef]
- Silva, A.P.D.; Gallardo, R.A. The Chicken MHC: Insights into genetic resistance, immunity, and inflammation following infectious bronchitis virus infections. Vaccines 2020, 8, 637. [Google Scholar] [CrossRef]
- Miller, M.M.; Taylor, R.L., Jr. Brief review of the chicken Major Histocompatibility Complex: The genes, their distribution on chromosome 16, and their contributions to disease resistance. Poult. Sci. 2016, 95, 375–392. [Google Scholar] [CrossRef]
- Karsten, S.L.; Sang, T.K.; Gehman, L.T.; Chatterjee, S.; Liu, J.; Lawless, G.M.; Sengupta, S.; Berry, R.W.; Pomakian, J.; Oh, H.S.; et al. A genomic screen for modifiers of tauopathy identifies puromycin-sensitive aminopeptidase as an inhibitor of tau-induced neurodegeneration. Neuron 2006, 51, 549–560. [Google Scholar] [CrossRef]
- Bhutani, N.; Venkatraman, P.; Goldberg, A.L. Puromycin-sensitive aminopeptidase is the major peptidase responsible for digesting polyglutamine sequences released by proteasomes during protein degradation. EMBO J. 2007, 26, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Stoltze, L.; Schirle, M.; Schwarz, G.; Schröter, C.; Thompson, M.W.; Hersh, L.B.; Kalbacher, H.; Stevanovic, S.; Rammensee, H.G.; Schild, H. Two new proteases in the MHC class I processing pathway. Nat. Immunol. 2000, 1, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Hui, K.S.; Saito, M.; Hui, M.; Saito, M.; Lajtha, A.; Yamamoto, K.; Osawa, T. Two cytosolic puromycin-sensitive aminopeptidase isozymes in chicken brain: Molecular homology to brain-specific 14-3-3 protein. Neurochem. Int. 1993, 22, 445–453. [Google Scholar] [CrossRef]
- Li, X.; Buxbaum, J.N. Transthyretin and the brain re-visited: Is neuronal synthesis of transthyretin protective in Alzheimer’s disease? Mol. Neurodegener. 2011, 6, 79. [Google Scholar] [CrossRef]
- Rapezzi, C.; Quarta, C.C.; Riva, L.; Longhi, S.; Gallelli, I.; Lorenzini, M.; Ciliberti, P.; Biagini, E.; Salvi, F.; Branzi, A. Transthyretin-related amyloidoses and the heart: A clinical overview. Nat. Rev. Cardiol. 2010, 7, 398–408. [Google Scholar] [CrossRef]
- Dockray, G.J.; Sault, C.; Holmes, S. Antibodies to FMRF amide, and the related pentapeptide LPLRF amide, reveal two groups of immunoreactive peptides in chicken brain. Regul. Pept. 1986, 16, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Schick, B.P.; Gradowski, J.F.; San Antonio, J.D. Synthesis, secretion, and subcellular localization of serglycin proteoglycan in human endothelial cells. Blood 2001, 97, 449–458. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Franklin, C.C.; Backos, D.S.; Mohar, I.; White, C.C.; Forman, H.J.; Kavanagh, T.J. Structure, function, and post-translational regulation of the catalytic and modifier subunits of glutamate cysteine ligase. Mol. Aspects. Med. 2009, 30, 86–98. [Google Scholar] [CrossRef]
- Dalton, T.P.; Chen, Y.; Schneider, S.N.; Nebert, D.W.; Shertzer, H.G. Genetically altered mice to evaluate glutathione homeostasis in health and disease. Free Radic. Biol. Med. 2004, 37, 1511–1526. [Google Scholar] [CrossRef]
- Forgac, M. Vacuolar ATPases: Rotary proton pumps in physiology and pathophysiology. Nat. Rev. Mol. Cell. Biol. 2007, 8, 917–929. [Google Scholar] [CrossRef]
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell. Biol. 2010, 11, 50–61. [Google Scholar] [CrossRef]
- Xu, H.; Ren, D. Lysosomal physiology. Annu. Rev. Physiol. 2015, 77, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Mo, G.; Hu, B.; Wei, P.; Luo, Q.; Zhang, X. The role of chicken prolactin, growth hormone and their receptors in the immune system. Front. Microbiol. 2022, 13, 900041. [Google Scholar] [CrossRef] [PubMed]
- Bastan, R.; Eskandari, N.; Sabzghabaee, A.M.; Manian, M. Serine/threonine phosphatases: Classification, roles and pharmacological regulation. Int. J. Immunopathol. Pharmacol. 2014, 27, 473–484. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element/Method | Zn/FAAS 1 |
|---|---|
| Flame/assay technique | acetylene/air/absorption |
| Wavelength [nm] | 213.9 |
| Slit width [nm] | 1.0 |
| Lamp current [mA] | 5.0 |
| LOD—limit of detection [mg/kg] | 0.4 |
| LOQ—limit of quantitation [mg/kg] | 0.9 |
| Precision (coefficient of variation) (%) | 5.1 |
| Recovery of the CRM (%) | 106 |
| Expanded uncertainty [%] | 14 |
| Effect | Serum | Liver | Small Intestine | Yolk Sac | |||
|---|---|---|---|---|---|---|---|
| 12 h | 7 Days | 12 h | 7 Days | 12 h | 7 Days | 12 h | |
| Prob+ (group II) | 1.57 ± 0.20 | 2.99 ± 0.31 | 18.40 ± 2.35 | 22.8 ± 0.53 | 23.63 ± 2.83 | 27.35 ± 0.33 | 14.98 ± 1.59 |
| F = 2.75 | F = 25.74 | F = 9.42 | F = 0.0 | F = 0.10 | F = 8.40 | F = 0.50 | |
| p = 0.11 | p < 0.001 | p < 0.001 | p = 0.77 | p = 0.76 | p < 0.001 | p = 0.49 | |
| η2 = 0.12% | η2 = 56.27% | η2 =29.4% | η2 = 0.44% | η2 =0.47% | η2 = 8.32% | η2 = 0.31% | |
| Zn-Gly+ (group III) | 2.20 ± 0.30 | 2.93 ± 0.22 | 21.00 ± 3.43 | 34.73 ± 7.14 | 24.67 ± 2.33 | 24.85 ± 1.56 | 32.82 ± 4.76 |
| F = 1375.24 | F = 0.25 | F = 1.78 | F = 131 | F = 0.29 | F = 58.64 | F = 94.43 | |
| p < 0.001 | p = 0.63 | p = 0.2 | p < 0.001 | p = 0.59 | p < 0.001 | p < 0.001 | |
| η2 = 60.11% | η2 = 1.22% | η2 = 5.50% | η2 = 86.77% | η2 = 1.39% | η2 = 61.62% | η2 = 60.34% | |
| Zn-Gly+Prob+ (group IV) | 13.88 ± 1.69 | 2.58 ± 0.26 | 23.30 ± 2.28 | 30.85 ± 0.56 | 24.43 ± 2.16 | 24.78 ± 1.09 | 25.12 ± 2.82 |
| F = 889.93 | F = 1.23 | F = 1.03 | F = 3.0 | F = 0.28 | F = 8.13 | F = 41.56 | |
| p < 0.001 | p = 0.29 | p = 0.32 | p = 0.12 | p = 0.60 | p < 0.001 | p < 0.001 | |
| η2 = 38.90% | η2 = 5.78% | η2 =3.2% | η2 = 11.77% | η2 = 96.77% | η2 = 8.54% | η2 = 12.78% | |
| Protein Name | Weight (Da) | UniProt Entry |
|---|---|---|
| Liver | ||
| FMRFamide-like neuropeptide | 645 | P83308 |
| Transthyretin | 1117 | Q6LAP1 |
| Puromycin-sensitive aminopeptidase isozyme II | 2162 | Q9PS17 |
| MHC class I antigen | (7052.50)/7053 | Q4ZGL8 |
| Small intestine | ||
| Insulin 2 | 5067 | A0A3G3C4S4 |
| Mitochondrial inner membrane protease subunit 2 | 11,298 | A0A8V0X568 |
| Serglycin | 14,070 | A0A8V0ZRY6 |
| Large intestine | ||
| Glutamate—cysteine ligase | 8172 | A0A8V0X4D6 |
| V-type proton ATPase subunit | 9008 | Q5ZJC4 |
| Yolk sac | ||
| Protein-serine/threonine phosphatase | 8433 | A0A3Q2U207 |
| V-type proton ATPase subunit | 9002 | Q5ZJC4 |
| Epidermal differentiation protein | 9152 | A0A088BHA8 |
| Prolactin-releasing hormone | 9678 | A0A1D5PYU4 |
| Time | Tissue | SOD Band Volume | SD (±) | Time | Tissue | SOD Band Volume | SD (±) |
|---|---|---|---|---|---|---|---|
| Liver | Liver | ||||||
| 12 h | I | 901 | 4.32 | 7 days | I | 859 | 3.83 |
| II | 940 | 4.37 | II | 751 | 1.87 | ||
| III | 994 | 3.85 | III | 686 | 4.08 | ||
| IV | 1034 | 6.06 | IV | 765 | 4.08 | ||
| Small intestine | Small intestine | ||||||
| 12 h | I | 977 | 8.89 | 7 days | I | 926 | 2.34 |
| II | 844 | 4.04 | II | 445 | 6.28 | ||
| III | 1574 | 3.83 | III | 683 | 5.89 | ||
| IV | 1788 | 2.16 | IV | 1537 | 5.01 | ||
| Large intestine | Large intestine | ||||||
| 12 h | I | 406 | 5.09 | 7 days | I | 1672 | 6.92 |
| II | 297 | 2.88 | II | 421 | 8.01 | ||
| III | 817 | 3.50 | III | 1819 | 6.10 | ||
| IV | 911 | 3.08 | IV | 976 | 5.16 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciszewski, A.; Jarosz, Ł.S.; Michalak, K.; Marek, A.; Grądzki, Z.; Wawrzykowski, J.; Szymczak, B.; Rysiak, A. Proteome and Peptidome Changes and Zn Concentration in Chicken after In Ovo Stimulation with a Multi-Strain Probiotic and Zn-Gly Chelate: Preliminary Research. Curr. Issues Mol. Biol. 2024, 46, 1259-1280. https://doi.org/10.3390/cimb46020080
Ciszewski A, Jarosz ŁS, Michalak K, Marek A, Grądzki Z, Wawrzykowski J, Szymczak B, Rysiak A. Proteome and Peptidome Changes and Zn Concentration in Chicken after In Ovo Stimulation with a Multi-Strain Probiotic and Zn-Gly Chelate: Preliminary Research. Current Issues in Molecular Biology. 2024; 46(2):1259-1280. https://doi.org/10.3390/cimb46020080
Chicago/Turabian StyleCiszewski, Artur, Łukasz S. Jarosz, Katarzyna Michalak, Agnieszka Marek, Zbigniew Grądzki, Jacek Wawrzykowski, Bartłomiej Szymczak, and Anna Rysiak. 2024. "Proteome and Peptidome Changes and Zn Concentration in Chicken after In Ovo Stimulation with a Multi-Strain Probiotic and Zn-Gly Chelate: Preliminary Research" Current Issues in Molecular Biology 46, no. 2: 1259-1280. https://doi.org/10.3390/cimb46020080
APA StyleCiszewski, A., Jarosz, Ł. S., Michalak, K., Marek, A., Grądzki, Z., Wawrzykowski, J., Szymczak, B., & Rysiak, A. (2024). Proteome and Peptidome Changes and Zn Concentration in Chicken after In Ovo Stimulation with a Multi-Strain Probiotic and Zn-Gly Chelate: Preliminary Research. Current Issues in Molecular Biology, 46(2), 1259-1280. https://doi.org/10.3390/cimb46020080