
HTRA1 in Placental Cell Models: A Possible Role in Preeclampsia

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Protein Extracts and Western Blotting
2.3. Immunofluorescence Staining
2.4. Hydrogen Peroxide Treatments
2.5. HTRA1 Silencing and Overexpression
2.6. Wound Healing Assay
2.7. Transwell Invasion Assay
2.8. Statistical Analysis
3. Results
3.1. HTRA1 Protein Is Expressed in Placental Cell Lines
3.2. Oxidative Stress Increased HTRA1 Protein Expression in BeWo and HTR8/SVneo Cells
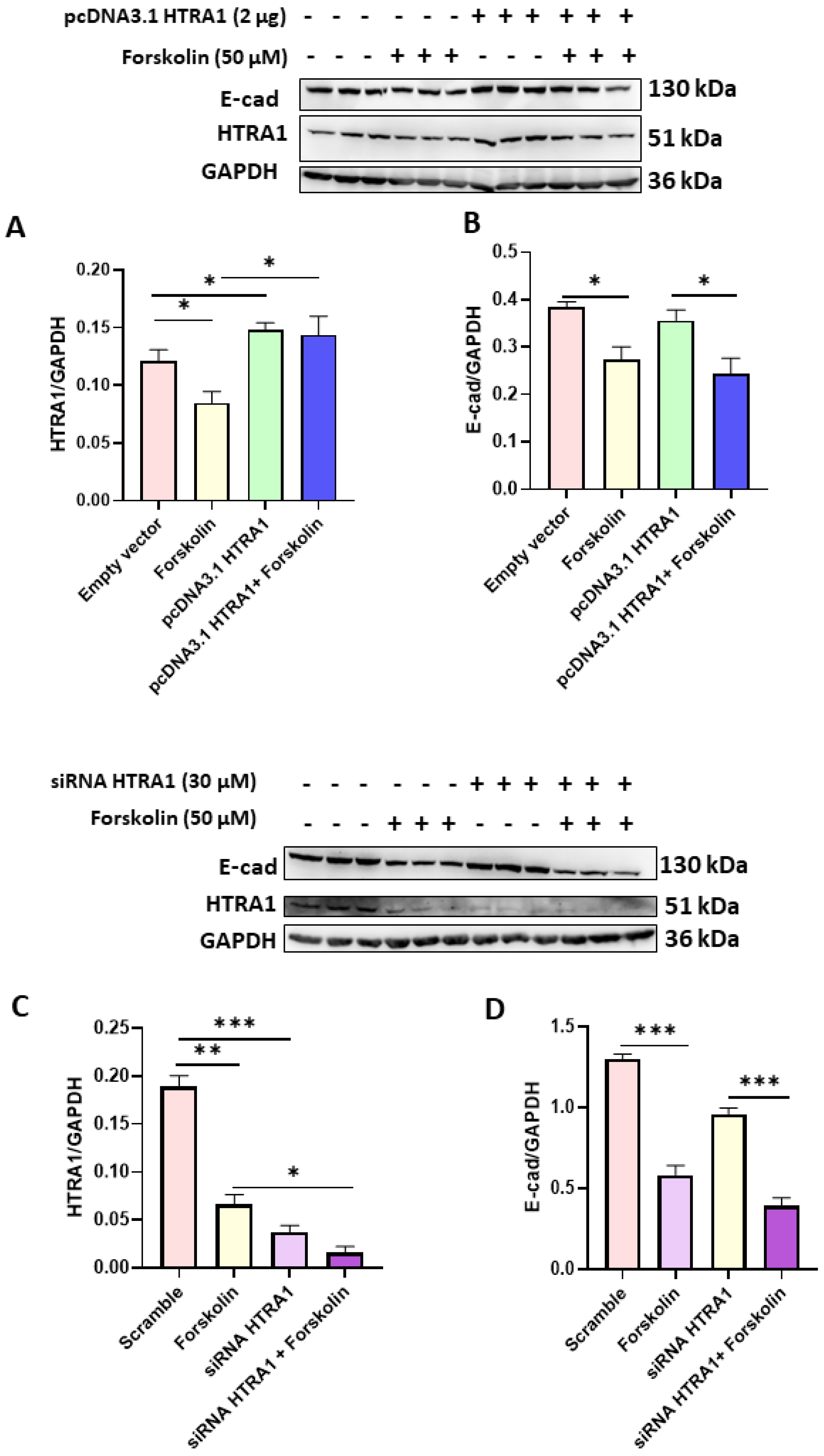
3.3. HTRA1 mRNA and Protein Expression Were Efficiently Modulated under Overexpressing and Silencing Conditions
3.4. HTRA1 Protein Overexpression and Silencing Did Not Alter BeWo Syncytialization
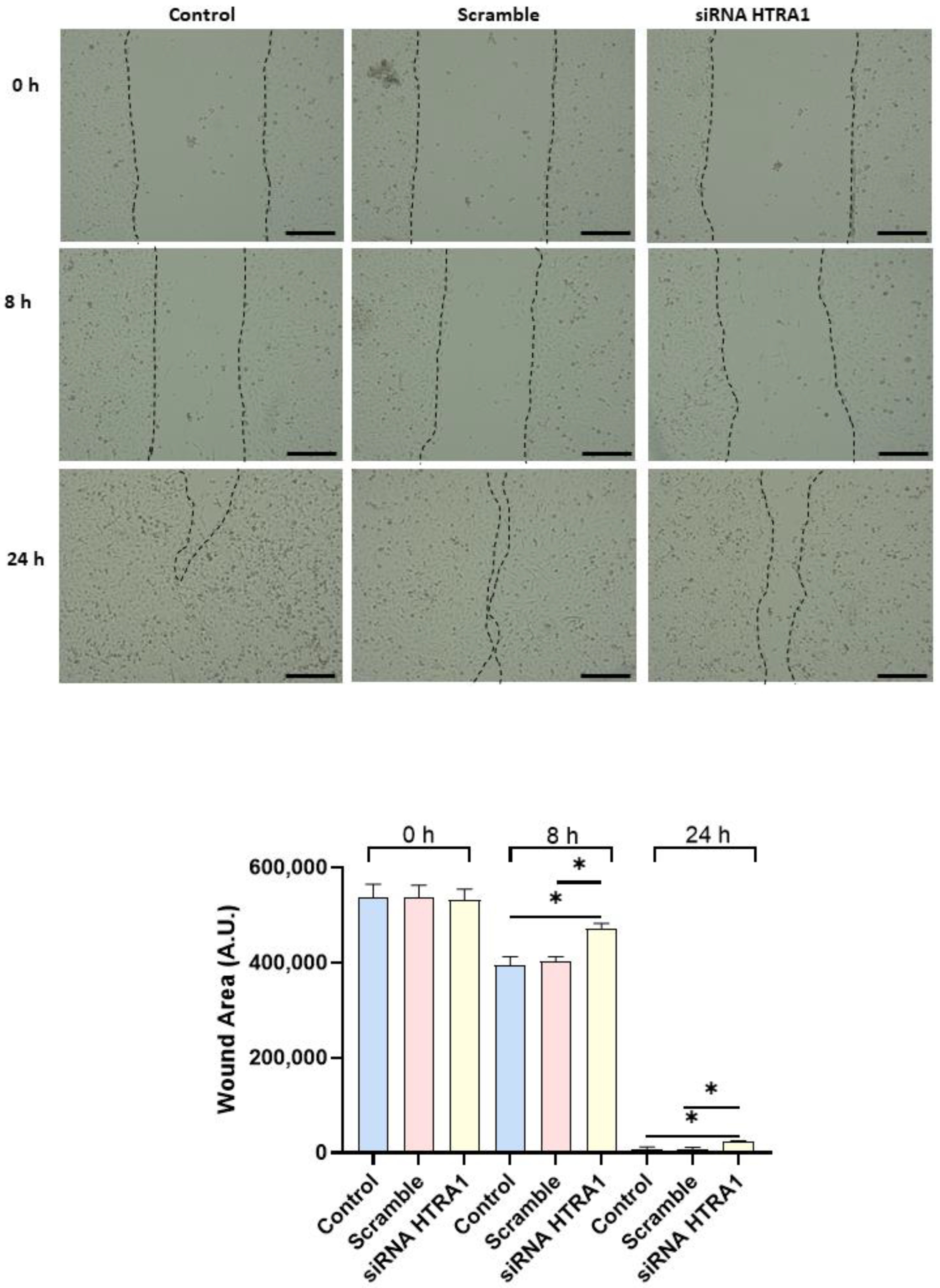
3.5. HTRA1 Protein Regulates HTR8/SVneo Cells Motility
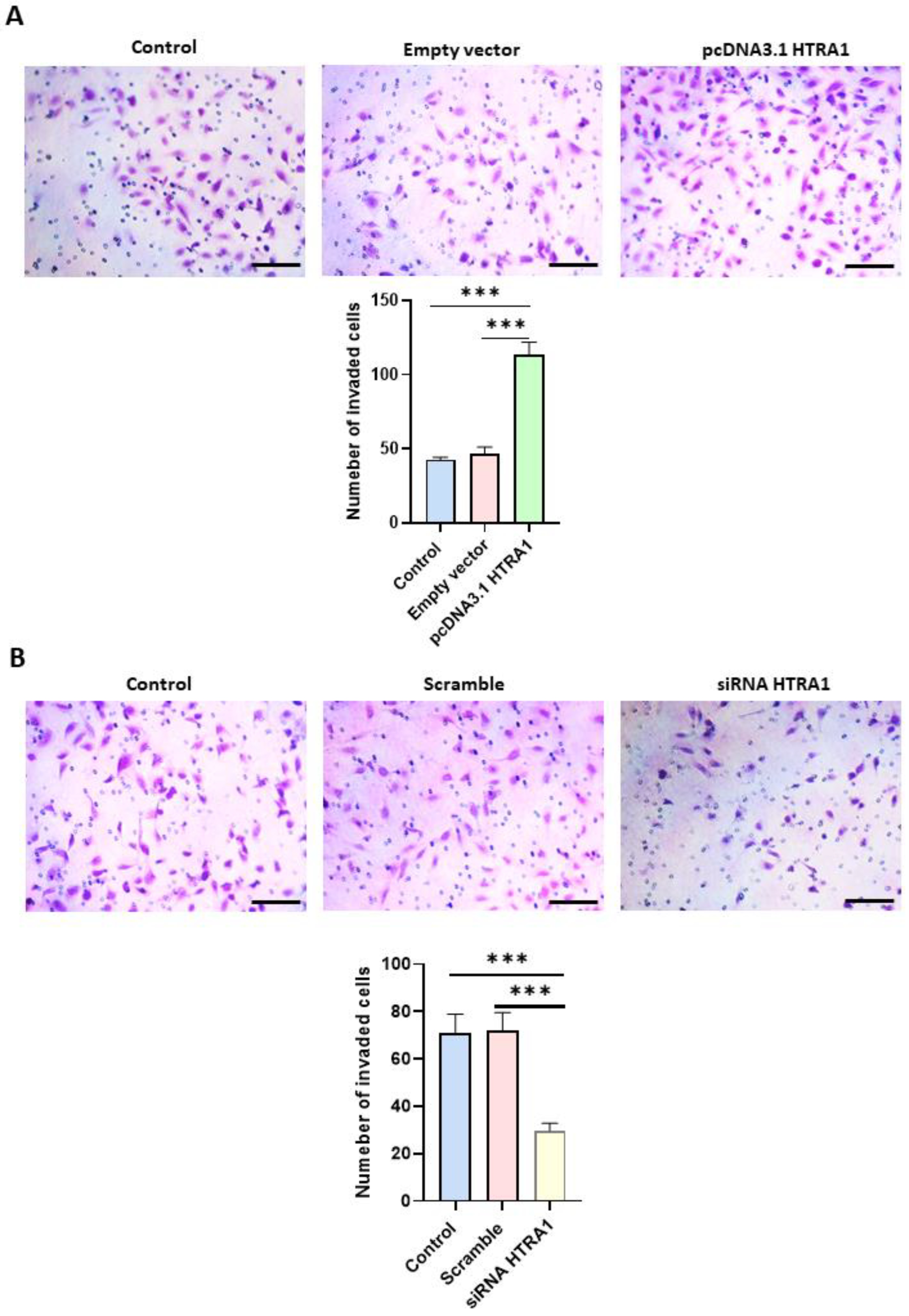
3.6. HTRA1 Protein Regulates HTR8/SVneo Cells Invasion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costa, M.A. The endocrine function of human placenta: An overview. Reprod. Biomed. Online 2016, 32, 14–43. [Google Scholar] [CrossRef] [PubMed]
- Whitley, G.S.; Cartwright, J.E. Cellular and molecular regulation of spiral artery remodelling: Lessons from the cardiovascular field. Placenta 2010, 31, 465–474. [Google Scholar] [CrossRef]
- Whitley, G.S.; Cartwright, J.E. Trophoblast-mediated spiral artery remodelling: A role for apoptosis. J. Anat. 2009, 215, 21–26. [Google Scholar] [CrossRef]
- Ridder, A.; Giorgione, V.; Khalil, A.; Thilaganathan, B. Preeclampsia: The Relationship between Uterine Artery Blood Flow and Trophoblast Function. Int. J. Mol. Sci. 2019, 20, 3263. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef] [PubMed]
- Phipps, E.A.; Thadhani, R.; Benzing, T.; Karumanchi, S.A. Pre-eclampsia: Pathogenesis, novel diagnostics and therapies. Nat. Rev. Nephrol. 2019, 15, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Fantone, S.; Mazzucchelli, R.; Giannubilo, S.R.; Ciavattini, A.; Marzioni, D.; Tossetta, G. AT-rich interactive domain 1A protein expression in normal and pathological pregnancies complicated by preeclampsia. Histochem. Cell Biol. 2020, 154, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Kornacki, J.; Gutaj, P.; Kalantarova, A.; Sibiak, R.; Jankowski, M.; Wender-Ozegowska, E. Endothelial Dysfunction in Pregnancy Complications. Biomedicines 2021, 9, 1756. [Google Scholar] [CrossRef]
- Fantone, S.; Giannubilo, S.R.; Marzioni, D.; Tossetta, G. HTRA family proteins in pregnancy outcome. Tissue Cell 2021, 72, 101549. [Google Scholar] [CrossRef]
- De Luca, A.; De Falco, M.; Fedele, V.; Cobellis, L.; Mastrogiacomo, A.; Laforgia, V.; Tuduce, I.L.; Campioni, M.; Giraldi, D.; Paggi, M.G.; et al. The serine protease HtrA1 is upregulated in the human placenta during pregnancy. J. Histochem. Cytochem. 2004, 52, 885–892. [Google Scholar] [CrossRef]
- Nie, G.Y.; Li, Y.; Minoura, H.; Batten, L.; Ooi, G.T.; Findlay, J.K.; Salamonsen, L.A. A novel serine protease of the mammalian HtrA family is up-regulated in mouse uterus coinciding with placentation. Mol. Hum. Reprod. 2003, 9, 279–290. [Google Scholar] [CrossRef] [PubMed]
- De Luca, A.; De Falco, M.; Severino, A.; Campioni, M.; Santini, D.; Baldi, F.; Paggi, M.G.; Baldi, A. Distribution of the serine protease HtrA1 in normal human tissues. J. Histochem. Cytochem. 2003, 51, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Licini, C.; Marzioni, D.; Mattioli-Belmonte, M. The multifaced role of HtrA1 in the development of joint and skeletal disorders. Bone 2022, 157, 116350. [Google Scholar] [CrossRef] [PubMed]
- Altobelli, E.; Latella, G.; Morroni, M.; Licini, C.; Tossetta, G.; Mazzucchelli, R.; Profeta, V.F.; Coletti, G.; Leocata, P.; Castellucci, M.; et al. Low HtrA1 expression in patients with longstanding ulcerative colitis and colorectal cancer. Oncol. Rep. 2017, 38, 418–426. [Google Scholar] [CrossRef]
- Tossetta, G.; Avellini, C.; Licini, C.; Giannubilo, S.R.; Castellucci, M.; Marzioni, D. High temperature requirement A1 and fibronectin: Two possible players in placental tissue remodelling. Eur. J. Histochem. 2016, 60, 2724. [Google Scholar] [CrossRef]
- Goteri, G.; Altobelli, E.; Tossetta, G.; Zizzi, A.; Avellini, C.; Licini, C.; Lorenzi, T.; Castellucci, M.; Ciavattini, A.; Marzioni, D. High temperature requirement A1, transforming growth factor beta1, phosphoSmad2 and Ki67 in eutopic and ectopic endometrium of women with endometriosis. Eur. J. Histochem. 2015, 59, 2570. [Google Scholar] [CrossRef]
- Marzioni, D.; Quaranta, A.; Lorenzi, T.; Morroni, M.; Crescimanno, C.; De Nictolis, M.; Toti, P.; Muzzonigro, G.; Baldi, A.; De Luca, A.; et al. Expression pattern alterations of the serine protease HtrA1 in normal human placental tissues and in gestational trophoblastic diseases. Histol. Histopathol. 2009, 24, 1213–1222. [Google Scholar] [CrossRef]
- Liu, C.; Xing, F.; He, Y.; Zong, S.; Luo, C.; Li, C.; Duan, T.; Wang, K.; Zhou, Q. Elevated HTRA1 and HTRA4 in severe preeclampsia and their roles in trophoblast functions. Mol. Med. Rep. 2018, 18, 2937–2944. [Google Scholar] [CrossRef]
- Lorenzi, T.; Marzioni, D.; Giannubilo, S.; Quaranta, A.; Crescimanno, C.; De Luca, A.; Baldi, A.; Todros, T.; Tranquilli, A.L.; Castellucci, M. Expression patterns of two serine protease HtrA1 forms in human placentas complicated by preeclampsia with and without intrauterine growth restriction. Placenta 2009, 30, 35–40. [Google Scholar] [CrossRef]
- Gesuita, R.; Licini, C.; Picchiassi, E.; Tarquini, F.; Coata, G.; Fantone, S.; Tossetta, G.; Ciavattini, A.; Castellucci, M.; Di Renzo, G.C.; et al. Association between first trimester plasma htra1 level and subsequent preeclampsia: A possible early marker? Pregnancy Hypertens. 2019, 18, 58–62. [Google Scholar] [CrossRef]
- Giannubilo, S.R.; Licini, C.; Picchiassi, E.; Tarquini, F.; Coata, G.; Fantone, S.; Tossetta, G.; Ciavattini, A.; Castellucci, M.; Giardina, I.; et al. First trimester HtrA1 maternal plasma level and spontaneous preterm birth. J. Matern. Fetal Neonatal Med. 2022, 35, 780–784. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Gesuita, R.; Di Renzo, G.C.; Meyyazhagan, A.; Tersigni, C.; Scambia, G.; Di Simone, N.; Marzioni, D. HtrA1 in Gestational Diabetes Mellitus: A Possible Biomarker? Diagnostics 2022, 12, 2705. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Duan, Y.F.; Bao, W.Y.; Liu, W.S.; Yang, Y.; Cai, H.H. HtrA1 regulates epithelial-mesenchymal transition in hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2015, 467, 589–594. [Google Scholar] [CrossRef]
- Zhang, L.; Lim, S.L.; Du, H.; Zhang, M.; Kozak, I.; Hannum, G.; Wang, X.; Ouyang, H.; Hughes, G.; Zhao, L.; et al. High temperature requirement factor A1 (HTRA1) gene regulates angiogenesis through transforming growth factor-beta family member growth differentiation factor 6. J. Biol. Chem. 2012, 287, 1520–1526. [Google Scholar] [CrossRef] [PubMed]
- Morosin, S.K.; Delforce, S.J.; de Meaultsart, C.C.; Lumbers, E.R.; Pringle, K.G. FURIN and placental syncytialisation: A cautionary tale. Cell Death Dis. 2021, 12, 635. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, G.Y.; Wang, J.; Lu, S.L.; Cao, J.; Sun, L.Z. A novel bridge between oxidative stress and immunity: The interaction between hydrogen peroxide and human leukocyte antigen G in placental trophoblasts during preeclampsia. Am. J. Obstet. Gynecol. 2012, 206, 447.e7–447.e16. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, H.; Silva, C.; Martel, F. The effects of aspartame on the HTR8/SVneo extravillous trophoblast cell line. Reprod. Biol. 2022, 22, 100678. [Google Scholar] [CrossRef]
- Ozmen, A.; Guzeloglu-Kayisli, O.; Tabak, S.; Guo, X.; Semerci, N.; Nwabuobi, C.; Larsen, K.; Wells, A.; Uyar, A.; Arlier, S.; et al. Preeclampsia is Associated with Reduced ISG15 Levels Impairing Extravillous Trophoblast Invasion. Front. Cell Dev. Biol. 2022, 10, 898088. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Lv, X.; Liu, H. Interleukin-17 promotes proliferation, migration, and invasion of trophoblasts via regulating PPAR-gamma/RXR-alpha/Wnt signaling. Bioengineered 2022, 13, 1224–1234. [Google Scholar] [CrossRef]
- Das, M.; Xu, B.; Lin, L.; Chakrabarti, S.; Shivaswamy, V.; Rote, N.S. Phosphatidylserine efflux and intercellular fusion in a BeWo model of human villous cytotrophoblast. Placenta 2004, 25, 396–407. [Google Scholar] [CrossRef]
- Orendi, K.; Gauster, M.; Moser, G.; Meiri, H.; Huppertz, B. The choriocarcinoma cell line BeWo: Syncytial fusion and expression of syncytium-specific proteins. Reproduction 2010, 140, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.M.; Pater, J.A.; MacPhee, D.J. Integrin linked kinase regulates syncytialization of BeWo trophoblast cells. Biol. Reprod. 2017, 96, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Zong, L.; Wang, L.; Huang, P.; Shao, W.; Song, Y.; Gou, W. High temperature requirement A1 in placental tissues and serum from pre-eclamptic pregnancies with or without fetal growth restriction. Arch. Med. Sci. 2013, 9, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Montanari, E.; Marzioni, D.; Goteri, G. Role of NRF2 in Ovarian Cancer. Antioxidants 2022, 11, 663. [Google Scholar] [CrossRef]
- Marzioni, D.; Mazzucchelli, R.; Fantone, S.; Tossetta, G. NRF2 modulation in TRAMP mice: An in vivo model of prostate cancer. Mol. Biol. Rep. 2022, 50, 873–881. [Google Scholar] [CrossRef]
- Sahakyan, G.; Vejux, A.; Sahakyan, N. The Role of Oxidative Stress-Mediated Inflammation in the Development of T2DM-Induced Diabetic Nephropathy: Possible Preventive Action of Tannins and Other Oligomeric Polyphenols. Molecules 2022, 27, 9035. [Google Scholar] [CrossRef]
- Alsawaf, S.; Alnuaimi, F.; Afzal, S.; Thomas, R.M.; Chelakkot, A.L.; Ramadan, W.S.; Hodeify, R.; Matar, R.; Merheb, M.; Siddiqui, S.S.; et al. Plant Flavonoids on Oxidative Stress-Mediated Kidney Inflammation. Biology 2022, 11, 1717. [Google Scholar] [CrossRef]
- Huppertz, B. The Critical Role of Abnormal Trophoblast Development in the Etiology of Preeclampsia. Curr. Pharm. Biotechnol. 2018, 19, 771–780. [Google Scholar] [CrossRef]
- Gao, X.; Qu, H.; Zhang, Y. Circ_0001326 suppresses trophoblast cell proliferation, invasion, migration and epithelial-mesenchymal transition progression in preeclampsia by miR-188-3p/HtrA serine peptidase 1 axis. J. Hypertens. 2023, 10, 1097. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, B.; Chang, J.; Feng, L.; Zhao, X. Melatonin regulates trophoblast pyroptosis, invasion and migration in preeclampsia by inhibiting HtrA1 transcription through the microRNA-520c-3p/SETD7 axis. Am. J. Reprod. Immunol. 2022, 87, e13523. [Google Scholar] [CrossRef]
- Yoshida, K.; Kusama, K.; Azumi, M.; Yoshie, M.; Kato, K.; Tamura, K. Endoplasmic reticulum stress-regulated high temperature requirement A1 (HTRA1) modulates invasion and angiogenesis-related genes in human trophoblasts. J. Pharmacol. Sci. 2022, 150, 267–274. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Western Blot Dilution | Company |
|---|---|---|
| pAb Rabbit anti-human HTRA1 (#PA5-11412) | 1:500 | Thermo Fisher Scientific, Waltham, US |
| mAb Mouse anti-human E-cadherin (#sc-8426) | 1:400 | Santa Cruz Biotechnology, Dallas, US |
| mAb Mouse anti-human Cytokeratin 7 (#sc-23876) | 1:400 | Santa Cruz Biotechnology, Dallas, US |
| mAb Mouse anti-human GAPDH (#sc- 47724) | 1:1000 | Santa Cruz Biotechnology, Dallas, US |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tossetta, G.; Fantone, S.; Giannubilo, S.R.; Ciavattini, A.; Senzacqua, M.; Frontini, A.; Marzioni, D. HTRA1 in Placental Cell Models: A Possible Role in Preeclampsia. Curr. Issues Mol. Biol. 2023, 45, 3815-3828. https://doi.org/10.3390/cimb45050246
Tossetta G, Fantone S, Giannubilo SR, Ciavattini A, Senzacqua M, Frontini A, Marzioni D. HTRA1 in Placental Cell Models: A Possible Role in Preeclampsia. Current Issues in Molecular Biology. 2023; 45(5):3815-3828. https://doi.org/10.3390/cimb45050246
Chicago/Turabian StyleTossetta, Giovanni, Sonia Fantone, Stefano Raffaele Giannubilo, Andrea Ciavattini, Martina Senzacqua, Andrea Frontini, and Daniela Marzioni. 2023. "HTRA1 in Placental Cell Models: A Possible Role in Preeclampsia" Current Issues in Molecular Biology 45, no. 5: 3815-3828. https://doi.org/10.3390/cimb45050246
APA StyleTossetta, G., Fantone, S., Giannubilo, S. R., Ciavattini, A., Senzacqua, M., Frontini, A., & Marzioni, D. (2023). HTRA1 in Placental Cell Models: A Possible Role in Preeclampsia. Current Issues in Molecular Biology, 45(5), 3815-3828. https://doi.org/10.3390/cimb45050246