
Infliximab Inhibits Colitis Associated Cancer in Model Mice by Downregulating Genes Associated with Mast Cells and Decreasing Their Accumulation
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Cell Lines
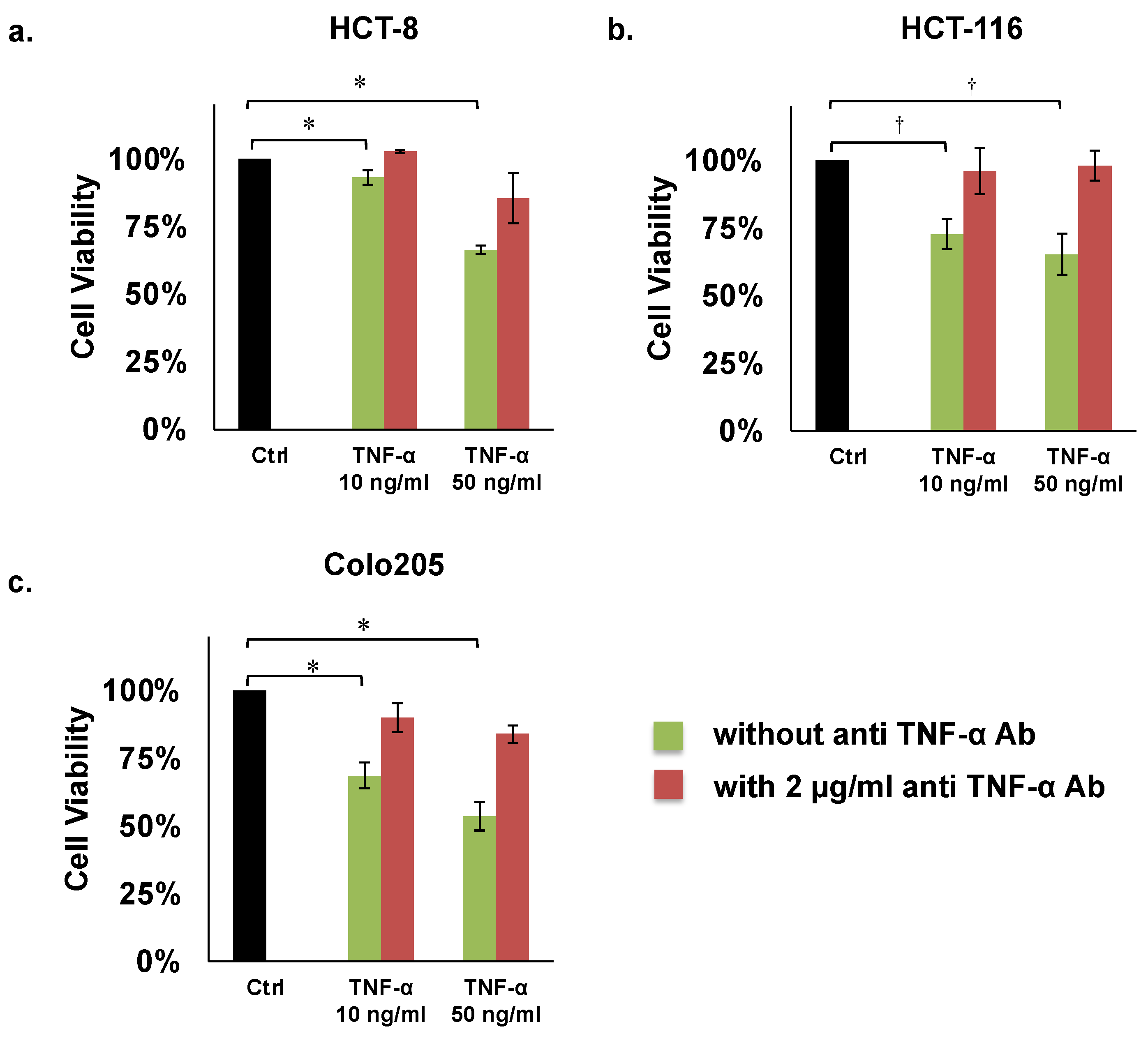
2.2. Proliferation Assays
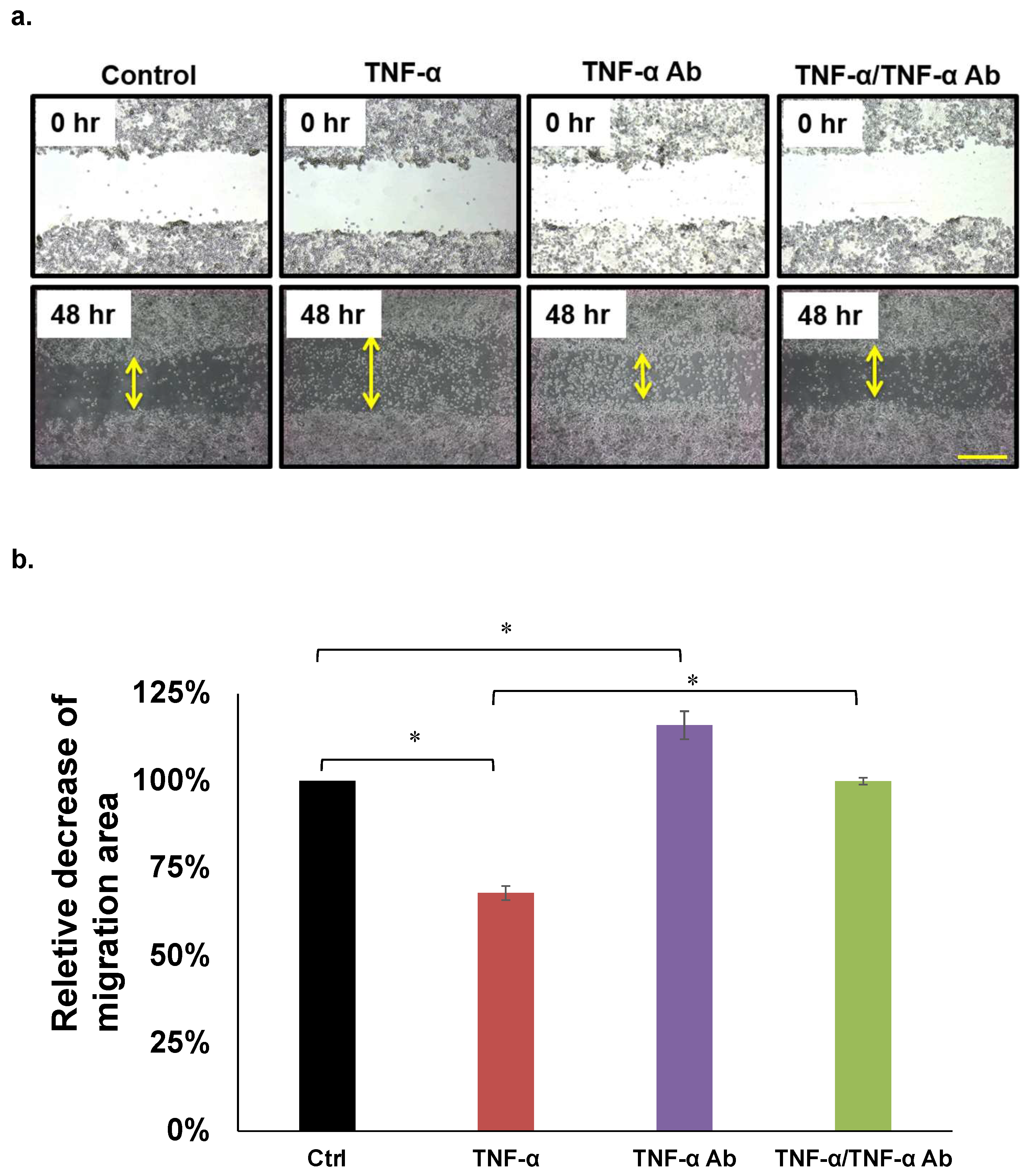
2.3. Migration Assays
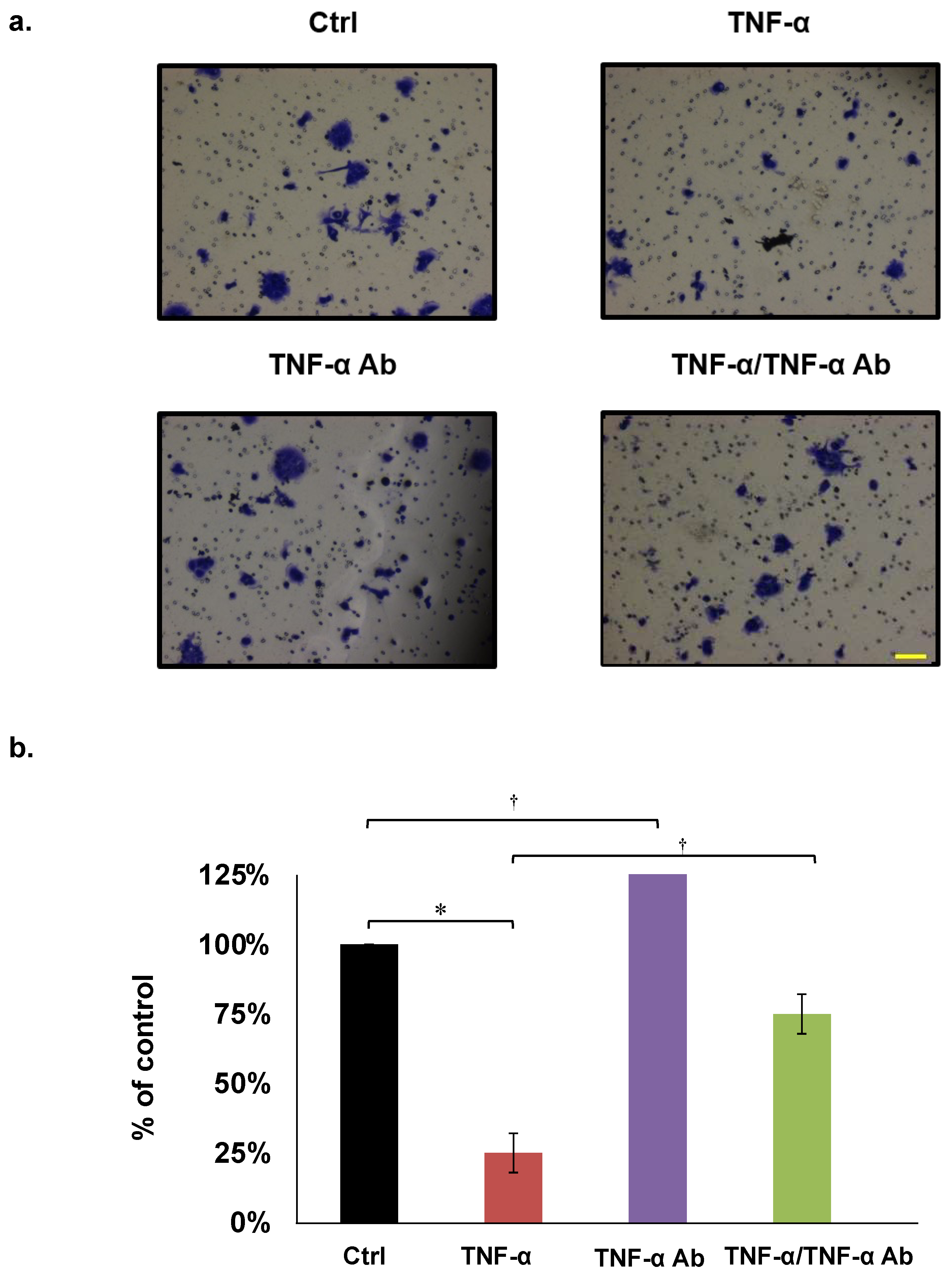
2.4. Invasion Assays
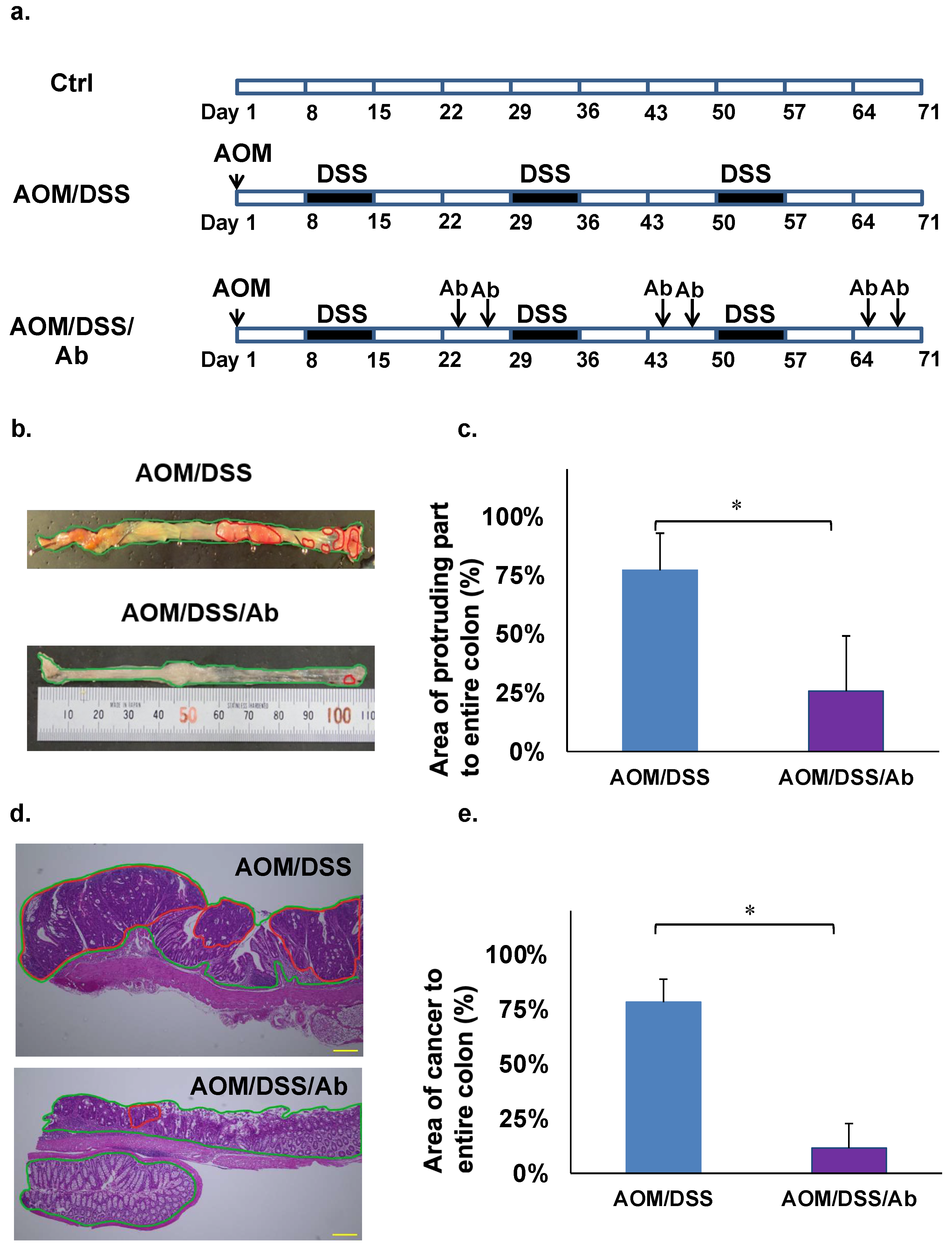
2.5. Animal Experiments
2.6. Calculation of Cancerous Areas in the Colon
2.7. Microarray Analysis
2.8. Staining Mast Cells with Toluidine Blue
2.9. Reverse Transcription Quantitative PCR (RT-qPCR)
2.10. Statistical Analysis
3. Results
3.1. Proliferation Assays
3.2. Migration Assays
3.3. Invasion Assays
3.4. Effects of Infliximab in Azoxymethane/Dextran Sodium Sulfate (AOM/DSS) Model Mice
3.5. Microarray Analysis
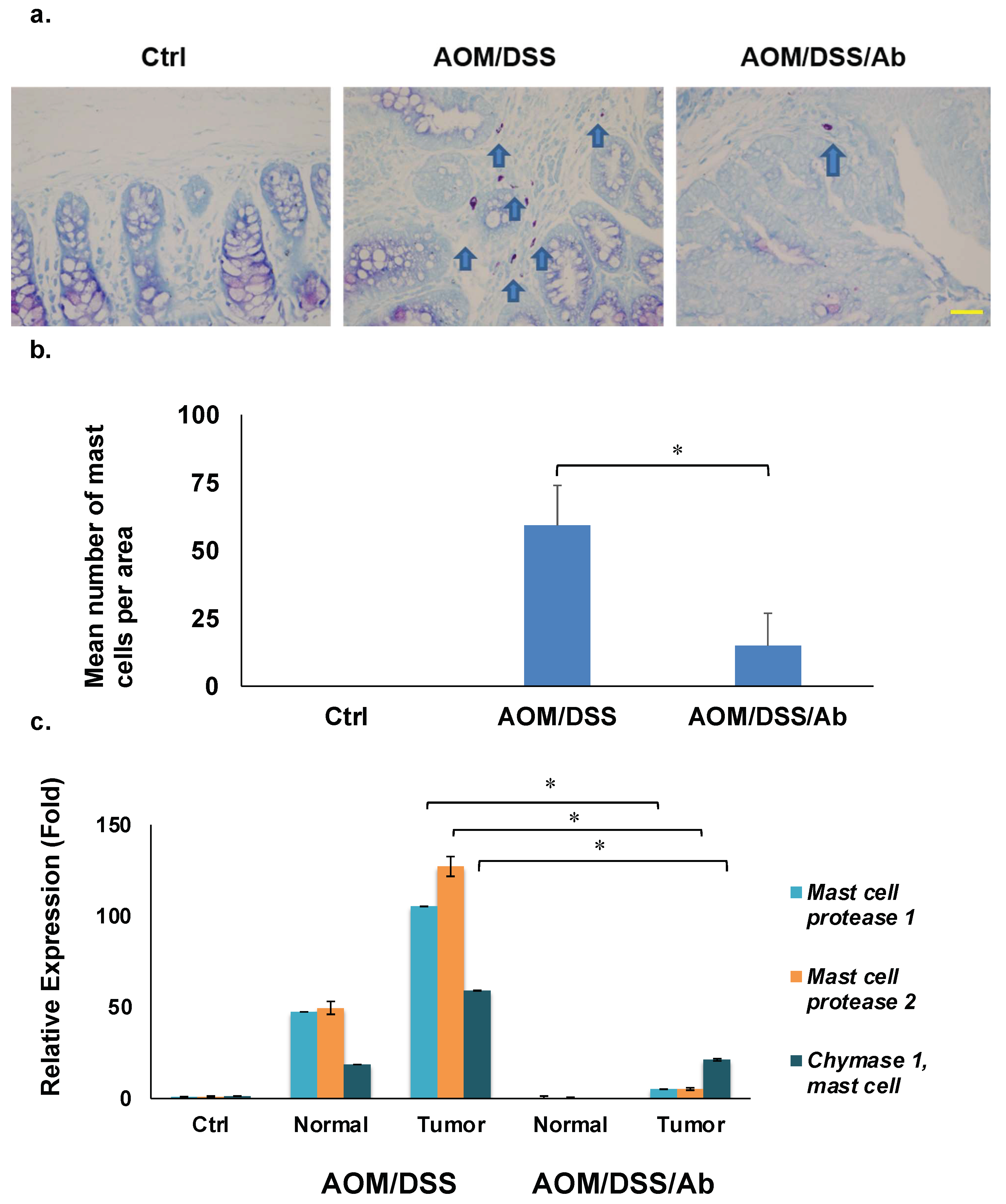
3.6. Toluidine Blue Staining
3.7. Reverse Transcription Quantitative PCR (RT-qPCR)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Chudy-Onwugaje, K.O.; Christian, K.E.; Farraye, F.A.; Cross, R.K. A State-of-the-Art Review of New and Emerging Therapies for the Treatment of IBD. Inflamm. Bowel Dis. 2019, 25, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Higashiyama, M.; Hokaria, R. New and Emerging Treatments for Inflammatory Bowel Disease. Digestion 2023, 104, 74–81. [Google Scholar] [CrossRef]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Hanauer, S.B.; Feagan, B.G.; Lichtenstein, G.R.; Mayer, L.F.; Schreiber, S.; Colombel, J.F.; Rachmilewitz, D.; Wolf, D.C.; Olson, A.; Bao, W.; et al. Maintenance infliximab for Crohn’s disease: The ACCENT I randomised trial. Lancet 2002, 359, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Magro, F.; Caldeira, D.; Alarcao, J.; Sousa, R.; Vaz-Carneiro, A. Infliximab reduces hospitalizations and surgery interventions in patients with inflammatory bowel disease: A systematic review and meta-analysis. Inflamm. Bowel Dis. 2013, 19, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Adegbola, S.O.; Sahnan, K.; Warusavitarne, J.; Hart, A.; Tozer, P. Anti-TNF Therapy in Crohn’s Disease. Int. J. Mol. Sci. 2018, 19, 2244. [Google Scholar] [CrossRef]
- Apostolaki, M.; Armaka, M.; Victoratos, P.; Kollias, G. Cellular mechanisms of TNF function in models of inflammation and autoimmunity. TNF Pathophysiol. 2010, 11, 1–26. [Google Scholar]
- Sethi, G.; Sung, B.; Aggarwal, B.B. TNF: A master switch for inflammation to cancer. Front. Biosci. 2008, 13, 5094–5107. [Google Scholar] [CrossRef]
- Braegger, C.P.; Nicholls, S.; Murch, S.H.; MacDonald, T.T.; Stephens, S. Tumor necrosis factor alpha in stool as a marker of intestinal inflammation. Lancet 1992, 339, 89–91. [Google Scholar] [CrossRef]
- Bazzoni, F.; Beutler, B. The tumor necrosis factor ligand and receptor families. N. Engl. J. Med. 1996, 334, 1717–1725. [Google Scholar] [CrossRef]
- Waters, J.P.; Pober, J.S.; Bradley, J.R. Tumour necrosis factor and cancer. J. Pathol. 2013, 230, 241–248. [Google Scholar] [CrossRef]
- Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 2009, 9, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Mukaida, N.; Baba, T. Chemokines in tumor development and progression. Exp. Cell Res. 2012, 318, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Oguma, K.; Oshima, H.; Aoki, M.; Uchio, R.; Naka, K.; Nakamura, S.; Hirao, A.; Saya, H.; Taketo, M.M.; Oshima, M. Activated macrophages promote Wnt signalling through tumour necrosis factor-alpha in gastric tumour cells. EMBO J. 2008, 27, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Komori, J.; Marusawa, H.; Machimoto, T.; Endo, Y.; Kinoshita, K.; Kou, T.; Haga, H.; Ikai, I.; Uemoto, S.; Chiba, T. Activation-induced cytidine deaminase links bile duct inflammation to human cholangiocarcinoma. Hepatology 2008, 47, 888–896. [Google Scholar] [CrossRef] [PubMed]
- von Roon, A.C.; Reese, G.; Teare, J.; Constantinides, V.; Darzi, A.W.; Tekkis, P.P. The risk of cancer in patients with Crohn’s disease. Dis. Colon Rectum 2007, 50, 839–855. [Google Scholar] [CrossRef]
- Raval, G.; Mehta, P. TNF-alpha inhibitors: Are they carcinogenic? Drug Healthc. Patient Saf. 2010, 2, 241–247. [Google Scholar] [PubMed]
- Ogawa, H.; Haneda, S.; Shibata, C.; Miura, K.; Nagao, M.; Ohnuma, S.; Kohyama, A.; Unno, M. Adenocarcinoma associated with perianal fistulas in Crohn’s disease. Anticancer Res. 2013, 33, 685–689. [Google Scholar] [PubMed]
- Brown, S.L.; Greene, M.H.; Gershon, S.K.; Edwards, E.T.; Braun, M.M. Tumor necrosis factor antagonist therapy and lymphoma development: Twenty-six cases reported to the Food and Drug Administration. Arthritis Rheum. 2002, 46, 3151–3158. [Google Scholar] [CrossRef] [PubMed]
- de Vries, H.S.; van Oijen, M.G.; de Jong, D.J. Serious events with infliximab in patients with inflammatory bowel disease: A 9-year cohort study in the Netherlands. Drug Saf. 2008, 31, 1135–1144. [Google Scholar] [CrossRef]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Yamada, Y.; Sugie, S.; Mori, H. A novel inflammation-related mouse colon carcinogenesis model induced by azoxymethane and dextran sodium sulfate. Cancer Sci. 2003, 94, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Spandidos, A.; Wang, H.; Seed, B. PrimerBank: A PCR primer database for quantitative gene expression analysis, 2012 update. Nucleic Acids Res. 2012, 40, D1144–D1149. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B. Signaling pathways of the TNF superfamily: A double-edged sword. Nat. Rev. Immunol. 2003, 3, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Chantana, C.; Yenjai, C.; Reubroycharoen, P.; Waiwut, P. Combination of Nimbolide and TNF-α-Increases Human Colon Adenocarcinoma Cell Death through JNK-mediated DR5 Up-Regulation. Asian Pac. J. Cancer Prev. 2016, 17, 2637–2641. [Google Scholar]
- Bhat, A.A.; Ahmad, R.; Uppada, S.B.; Singh, A.B.; Dhawan, P. Claudin-1 promotes TNF-α-induced epithelial-mesenchymal transition and migration in colorectal adenocarcinoma cells. Exp. Cell Res. 2016, 349, 119–127. [Google Scholar] [CrossRef]
- Egberts, J.H.; Cloosters, V.; Noack, A.; Schniewind, B.; Thon, L.; Klose, S.; Kettler, B.; von Forstner, C.; Kneitz, C.; Tepel, J.; et al. Anti-tumor necrosis factor therapy inhibits pancreatic tumor growth and metastasis. Cancer Res. 2008, 68, 1443–1450. [Google Scholar] [CrossRef]
- Kim, Y.J.; Hong, K.S.; Chung, J.W.; Kim, J.H.; Hahm, K.B. Prevention of colitis-associated carcinogenesis with infliximab. Cancer Prev. Res. 2010, 3, 1314–1333. [Google Scholar] [CrossRef]
- Popivanova, B.K.; Kitamura, K.; Wu, Y.; Kondo, T.; Kagaya, T.; Kaneko, S.; Oshima, M.; Fujii, C.; Mukaida, N. Blocking TNF-alpha in mice reduces colorectal carcinogenesis associated with chronic colitis. J. Clin. Investig. 2008, 118, 560–570. [Google Scholar]
- Greten, F.R.; Eckmann, L.; Greten, T.F.; Park, J.M.; Li, Z.W.; Egan, L.J.; Kagnoff, M.F.; Karin, M. IKKbeta links inflammation and tumorigenesis in a mouse model of colitis-associated cancer. Cell 2004, 118, 285–296. [Google Scholar] [CrossRef]
- Onizawa, M.; Nagaishi, T.; Kanai, T.; Nagano, K.-I.; Oshima, S.; Nemoto, Y.; Yoshioka, A.; Totsuka, T.; Okamoto, R.; Nakamura, T.; et al. Signaling pathway via TNF-alpha/NF-kappaB in intestinal epithelial cells may be directly involved in colitis-associated carcinogenesis. Am. J. Physiol.-Gastrointest. Liver Physiol. 2009, 296, G850–G859. [Google Scholar] [CrossRef]
- Wilson, J.A. Tumor necrosis factor alpha and colitis-assoiated colon cancer. N. Engl. J. Med. 2008, 358, 2733–2734. [Google Scholar] [CrossRef]
- Andoh, A.; Dequchi, Y.; Inatomi, O.; Yagi, Y.; Bamba, S.; Tsujikawa, T.; Fujiyama, Y. Immunohistochemical study of chymase-positive mast cells in inflammatory bowel disease. Oncol. Rep. 2006, 16, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Raithel, M.; Schneider, H.T.; Hahn, E.G. Effect of substance P on histamine secretion from gut mucosa in inflammatory bowel disease. Scand. J. Gastroenterol. 1999, 34, 496–503. [Google Scholar] [PubMed]
- Raithel, M.; Winterkamp, S.; Pacurar, A.; Ulrich, P.; Hochberger, J.; Hahn, E.G. Release of mast cell tryptase from human colorectal mucosa in inflammatory bowel disease. Scand. J. Gastroenterol. 2001, 36, 174–179. [Google Scholar] [CrossRef]
- Ribatti, D.; Crivellato, E. Mast cells, angiogenesis, and tumor growth. Biochim. Biophys. Acta 2012, 1822, 2–8. [Google Scholar] [CrossRef] [PubMed]
- de Souza Junior, D.A.; Santana, A.C.; da Silva, E.Z.; Oliver, C.; Jamur, M.C. The Role of Mast Cell Specific Chymases and Tryptases in Tumor Angiogenesis. Biomed. Res. Int. 2015, 2015, 142359. [Google Scholar] [CrossRef]
- Ribatti, D.; Vacca, A.; Nico, B.; Crivellato, E.; Roncali, L.; Dammacco, F. The role of mast cells in tumour angiogenesis. Br. J. Haematol. 2001, 115, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Lorentz, A.; Schwengberg, S.; Weier, G.; Raab, R.; Manns, M.P. Mast cells are an important cellular source of tumour necrosis factor alpha in human intestinal tissue. Gut 1999, 44, 643–652. [Google Scholar] [CrossRef]
- Wershil, B.K., IX. Mast cell-deficient mice and intestinal biology. Am. J. Physiol.-Gastrointest. Liver Physiol. 2000, 278, G343:8. [Google Scholar] [CrossRef]
- Misiak-Tloczek, A.; Brzezinska-Blaszczyk, E. IL-6, but not IL-4, stimulates chemokinesis and TNF stimulates chemotaxis of tissue mast cells: Involvement of both mitogen-activated protein kinases and phosphatidylinositol 3-kinase signalling pathways. Apmis 2009, 117, 558–567. [Google Scholar] [CrossRef]
- Yeo, M.; Kim, D.K.; Park, H.J.; Oh, T.Y.; Kim, J.H.; Cho, S.W.; Paik, Y.-K.; Hahm, K.-B. Loss of transgelin in repeated bouts of ulcerative colitis-induced colon carcinogenesis. Proteomics 2006, 6, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Imanishi, M.; Fujikura, D.; Sugiyama, M.; Tanimoto, K.; Mochiji, Y.; Takahashi, Y.; Hiura, K.; Watanabe, M.; Kashimoto, T.; et al. New inducible mast cell-deficient mouse model (Mcpt5/Cma1DTR). Biochem. Biophys. Res. Commun. 2021, 551, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Deveci, F.; Muz, M.H.; Ilhan, N.; Kirkil, G.; Turgut, T.; Akpolat, N. Evaluation of the anti-inflammatory effect of infliximab in a mouse model of acute asthma. Respirology 2008, 13, 488–497. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assays | Cell Lines | Inhibition by TNF-α (50 ng/mL) | Recovery Effect by Anti TNF-α Ab (2 µg/mL) |
|---|---|---|---|
| Proliferation | HCT8 | 66% * | 85% |
| HCT116 | 65% † | 98% | |
| COLO205 | 54% * | 84% | |
| Migration | HCT8 | 74% † | 91% |
| HCT116 | None | None | |
| COLO205 | 61% † | 100% | |
| Invasion | HCT8 | 49% * | 99% |
| HCT116 | 12% * | 20% | |
| COLO205 | 23% * | 75% |
| Gene | Name | Ab(-)/Normal * | Ab(-)/Ab(+) † |
|---|---|---|---|
| Mcpt1 | Mast cell protease 1 | 58.62 | 8.77 |
| Mcpt2 | Mast cell protease 2 | 54.93 | 7.91 |
| Cma1 | Chymase 1, mast cell | 71.01 | 4.84 |
| Mcpt4 | Mast cell protease 4 | 17.21 | 4.76 |
| Fcer1a | Fc receptor, IgE, high affinity I, alpha polypeptide | 27.10 | 4.74 |
| 9530003J23Rik | RIKEN cDNA 9530003J23 | 17.13 | 4.32 |
| Pla2g2e | Phospholipase A2, group IIE | 19.44 | 4.21 |
| Mmp8 | Matrix metallopeptidase 8 | 15.23 | 3.20 |
| Cpa3 | Carboxypeptidase A3, mast cell | 13.30 | 2.79 |
| Krt35 | Keratin 35 | 122.87 | 2.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.-Y.; Ohnuma, S.; Suzuki, H.; Ishida, M.; Ishii, K.; Hirosawa, T.; Hirashima, T.; Murakami, M.; Kobayashi, M.; Kudoh, K.; et al. Infliximab Inhibits Colitis Associated Cancer in Model Mice by Downregulating Genes Associated with Mast Cells and Decreasing Their Accumulation. Curr. Issues Mol. Biol. 2023, 45, 2895-2907. https://doi.org/10.3390/cimb45040189
Wang D-Y, Ohnuma S, Suzuki H, Ishida M, Ishii K, Hirosawa T, Hirashima T, Murakami M, Kobayashi M, Kudoh K, et al. Infliximab Inhibits Colitis Associated Cancer in Model Mice by Downregulating Genes Associated with Mast Cells and Decreasing Their Accumulation. Current Issues in Molecular Biology. 2023; 45(4):2895-2907. https://doi.org/10.3390/cimb45040189
Chicago/Turabian StyleWang, Dan-Yang, Shinobu Ohnuma, Hideyuki Suzuki, Masaharu Ishida, Kentaro Ishii, Takashi Hirosawa, Tomoaki Hirashima, Megumi Murakami, Minoru Kobayashi, Katsuyoshi Kudoh, and et al. 2023. "Infliximab Inhibits Colitis Associated Cancer in Model Mice by Downregulating Genes Associated with Mast Cells and Decreasing Their Accumulation" Current Issues in Molecular Biology 45, no. 4: 2895-2907. https://doi.org/10.3390/cimb45040189
APA StyleWang, D.-Y., Ohnuma, S., Suzuki, H., Ishida, M., Ishii, K., Hirosawa, T., Hirashima, T., Murakami, M., Kobayashi, M., Kudoh, K., Haneda, S., Musha, H., Naitoh, T., & Unno, M. (2023). Infliximab Inhibits Colitis Associated Cancer in Model Mice by Downregulating Genes Associated with Mast Cells and Decreasing Their Accumulation. Current Issues in Molecular Biology, 45(4), 2895-2907. https://doi.org/10.3390/cimb45040189