Abstract
Ficus simplicissima Lour. is an Asian species of fig tree in the family Moraceae. The chloroplast (cp) genome of F. simplicissima m3 was sequenced using the Pacbio sequel platform. The F. simplicissima cpDNA has a size of 160,321 bp in length, of which GC content accounts for 36.13%. The cp genome of F. simplicissima consists of a single large copy (LSC) with a size of 91,346 bp, a single small copy (SSC) with a size of 20,131 bp, and a pair of inverted repeats with a size of 24,421 to 24,423 bp. The cp genome of F. simplicissima has 127 genes, including 85 protein-coding genes, eight rRNA genes, and 34 tRNA genes; 92 simple sequence repeats and 39 long repeats were detected in the cpDNA of F. simplicissim. A comparative cp genome analysis among six species in the Ficus genus indicated that the genome structure and gene content were highly conserved. The non-coding regions show more differentiation than the coding regions, and the LSC and SSC regions show more differences than the inverted repeat regions. Phylogenetic analysis supported that F. simplicissima m3 had a close relationship with F. hirta. The complete cp genome of F. simplicissima was proposed as a chloroplast DNA barcoding for genus-level in the Moraceae family and the psbA-trnH gene region for species-level identification.
1. Introduction
The chloroplast (cp) genome is circular and relatively conserved among plants in terms of size, structure, and gene content [1]. The cp genome generally comprises two copies of inverted regions that divide the genome into a large single-copy region (LSC) and a small single-copy region (SSC). The cp genome provides genes participating in photosynthesis, transcription, and translation. In addition, its non-coding intergenic spacer regions are highly conserved and can be used for phylogenetics, population genetics, and species identification [2]. Earlier research used partial cp sequences for plant barcoding [3], but they are only universal for some plant taxa and have limitations at lower taxonomic levels [4,5]. Therefore, the whole cp genome is informative and extensive for plant barcoding [6,7]. With the advance of next-generation sequencing, whole chloroplast genome acquisition is now simpler and faster than ever. Coverage of four cp junctions between the inverted repeat (IR) and single-copy regions is performed by a third–generation sequencer, the PacBio system with Single Molecule Real-Time (SMRT) technology [8].
Ficus simplicissima Lour. is a species of fig tree in the Ficus genus. This genus contains about 1000 species of trees, shrubs, and vines and is distinct with a unique fruit structure known as syconium [9]. The species is found predominantly native to East Asian tropical regions. In folk medicine, F. simplicissima was used to treat pneumonia, vitiligo, diarrhea, tonsillitis, cough, and rheumatic pain and promote lactation [10]. Moreover, combining modified Radix Fici Simplicissimae with Western medicines was considered a potential treatment for SARS-CoV-2 patients [11].
Although the Ficus genus has different applications, its phylogenetic relationship is controversial. The Ficus classification system was based on morphological characteristics, and analysis of ribosomal DNA was reported by Corner (1965) and Huang et al. (2022). The information on the cp genomes of Ficus can be used in species identification and phylogenetic analysis of Ficus species [12,13]. However, the cp genome of F. simplicissima Lour. needed more because there were only raw reads of the cp genome of F. simplicissima Lour. at the China National GenBank [13].
The purpose of this study was to sequence and annotate the cp genome of F. simplicissima Lour m3; cp genome analysis, evaluate diversity and molecular evolutionary analysis of Ficus genus in the Moraceae family, search for cpDNA markers as potential DNA barcodes for Ficus species identification.
2. Materials and Methods
2.1. Plant Material, DNA Extraction, and cp Genome Sequencing
Ficus simplicissima Lour. m3 seeds were collected in the Thai Nguyen province (Hung Son township, Dai Tu district), then cultured on Murashige and Skoog medium. Regeneration of plants in vitro and plants grown in pots and in experimental gardens. Ficus simplicissima samples were identified by comparative morphology by the Department of Botany, Thai Nguyen University of Education, Vietnam (Figure 1). Fresh leaves from plantlets were harvested for genomic DNA extraction using the DNeasy® Plant Mini Kit. Absorption spectroscopy analysis on a Shimadzu Biospec Nano instrument at wavelengths A260 and A230 assessed DNA sample purity.

Figure 1.
Sample image of Ficus simplicissima Lour. m3 regenerated from seeds collected in the Thai Nguyen province (Hung Son township, Dai Tu district). (a) Plants regenerated in vitro from seeds grown in the experimental garden and (b) grown in pots, they are kept at the Department of Biology, Thai Nguyen University of Education; (c,d) fruit of F. simplicissima.
The concentration of DNA was determined using a Qubit 3 Fluorometer and Qubit HS DNA reagents. The integrity of the gDNA is assessed by 0.8% agarose gel electrophoresis. Besides, the total genomic DNA was used for library preparation using SMRTbell Express Template Prep Kit 2.0 (Pacific Biosciences, Menlo Park, CA, USA) following the manufacturer’s protocol (Pacific Biosciences). SMRTbell libraries were loaded on one chip and sequenced on a Pacbio SEQUEL system at the Key Laboratory for Gene Technology, Institute of Biotechnology (Hanoi, Vietnam).
2.2. Genome Assembly and Annotation
The cp genome sequences were determined via pbmm2 (https://github.com/PacificBiosciences/pbmm2, accessed on 20 August 2022) by mapping to Ficus hirta cp genomes (NC_051532.1) as the reference. Afterward, the CANU program [14] was used to assemble the cp genome. The assembled genome was annotated and analyzed using the GeSeq tool [15]. The tRNAscan-SE ve has confirmed the tRNA genes with default parameters. 1.21 software [16]. A circular genome map was created by the OrganellarGenomeDRAW tool (OGDRAW) ver. 1.3.1 [17]. Two methods searched Repeat sequences. MISA-web [18] was used to detect microsatellites with the following setting: 10 repeat units for mono-, five repeat units for di, four repeat units for tri-, and three repeat units for tetra-, penta-, and hexanucleotide SSRs. Dispersed repeats (including forward, reverse, complement, and palindromic repeats) were identified by REPuter [19] with a minimum repetition size of 20 times, hamming distance = 3, and sequence identities ≥90%.
2.3. Genome Comparison
For comparative analysis, five Ficus cp genomes were obtained from NCBI, and their accession numbers are as follows: Ficus concinna (MZ128521), Ficus formosana (NC_059898), Ficus pandurata var. angustifolia (NC_063593), Ficus sarmentosa voucher ZZ063 (NC_061976), Ficus hirta (NC_051532). The cp DNA nucleotide sequences of the six Ficus species, including the genome of F. simplicissima m3, have been linked with the MAFFT server and visualized by mVISTA software (Shuffle-LAGAN mode) [20] and used the F. simplicissima m3 genome to compare with genome five remaining species. Large single-copy (LSC), small single-copy (SSC), and inverted repeat (IR) regions among the Ficus species were visualized as the junction sites of chloroplast genomes and compared using the IRscope online program. Codon usage trends, Pi values, and nucleotide sequence polymorphisms among six Ficus species were determined by calculating the pi sliding window analysis between cp DNAs in DnaSP ver. 6.12.03 [21]. We chose a window size of 600 bp with a step size of 200 bp for sequence divergence analysis.
2.4. Phylogenetic Identification
The sequences of psbA-trnH and complete cp genome were downloaded from the GenBank of NCBI to illustrate phylogenetic relationship and position. The MAFFT server [22] was applied to align these sequences and maximum likelihood trees with 1000 bootstrap replicates were constructed by FastTree version 2.1.11 [23]. Subsequently, FigTree (version 1.4.4) [24] was employed to visualize phylogenetic trees.
3. Results
3.1. Chloroplast Genome Features of Ficus Simplicissima m3
In total, 51,578 reads and 3.7 Gb raw data sequences of the whole genome were generated from F. simplicissima m3. After trimming and selecting reads, the F. simplicissima m3 cp genome with a size 160,321 bp was assembled. The Ficus plastome possessed the classic quadripartite structure (Figure 2), containing one LSC region (68,977 bp), one SSC region (20,131 bp) and two inverted repeat (IR) regions (24,421 and 24,423 bp). The overall GC content was 35.9 (%) (Table 1). The cp genome of F. simplicissima m3 contains 127 genes, including 85 protein-coding genes, 8 rRNA genes, 34 tRNA genes. Of these, 18 genes were duplicated in the IR region and 21 genes contain introns. Additionally, 11 protein-coding genes (rps16, petB, petD, atpF, ndhA, ndhB (× 2), rpoC1, rpl16 and rpl2 (× 2)) and six RNA genes (rrn23 (×2), trnI-GAU (× 2) and trnA-UGC (× 2)) contained only one intron, while two protein-coding genes (clpP and ycf3) had two introns (Supplementary File S1 Table S1). The complete chloroplast genome of the F. simplicissima was submitted to GenBank in November 2022 and was granted the code BankIt2647431 Ficus_simplicissima_m3 OP928145 on 5 December 2022, and is waiting for the accession number (Supplementary File S2).
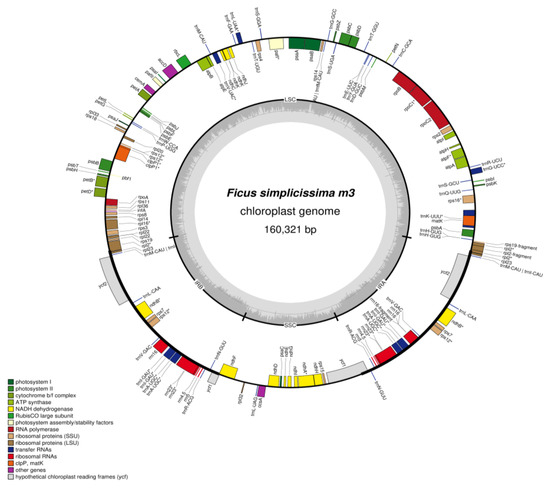
Figure 2.
Circular map of the chloroplast genome of F. simplicissima m3. Genes located outside of the circle are transcribed counter-clockwise, while genes shown inside are transcribed clockwise. The darker gray in the inner circle indicates GC content, and the lighter gray corresponds to AT content. Genes marked with the sign ‘*’ are the gene with intron.

Table 1.
Summary of the chloroplast genome of Ficus simplicissima m3 species.
3.2. Codon Usage
The chloroplast genome of F. simplicissima was analyzed for its codon usage frequency based on the nucleotide sequence of protein-coding genes and on relative synonymous codon usage (RSCU). The relative frequency of synonymous codons of the F. simplicissima m3 cp coding sequence was estimated. The results indicate that protein-coding genes were encoded by 54,960 codons and the four most common codons were UUU (phenylalanine), AAA (lysine), AAU (asparagine), and AUU (Isoleucine), corresponding to 2393 (4.35%), 2287 (4.16%), 1967 (3.57%), and 1830 (3.32%) codons, respectively. In terms of the prevalence of translated amino acids, leucine (5719) and isoleucine (4569) were the two most frequently used amino acids, while the least abundant was tryptophan (692 codons, approximately 1.25%). Thirty codons were used more frequently than other synonymous codons (RSCU > 1) and thirty-two codons were considered as relatively less used codons (RSCU < 1). Furthermore, AUG and UGG (methionine and tryptophan) showed no bias (RSCU = 1).
3.3. Repeat Sequence Analysis
Simple sequence repeats (SSRs) are tandemly repeated DNA sequences consisting of short, tandemly repeated di-, tri-, tetra-or penta-nucleotide motifs [25]. A total of 92 SSRs were identified in the F. simplicissima m3 cp genome. Among them, there were 49 mononucleotide repeats, 22 dinucleotide repeats, 5 trinucleotide repeats, 10 tetranucleotide repeats, 4 pentanucleotide, and 2 hexanucleotides. In addition, all the mononucleotide repeats belonged to A/T and were the highest (Figure 3).
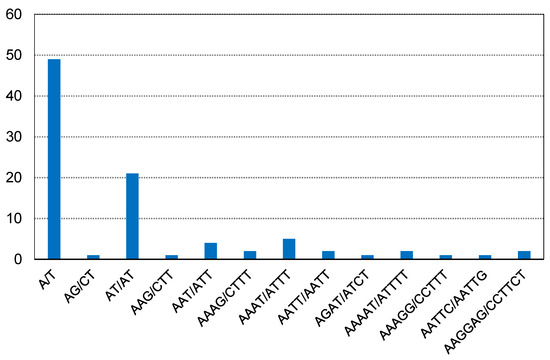
Figure 3.
Number of different repeat units of SSRs.
Additionally, the long complex repetitive sequences were explored, containing forward repeats, reverse repeats, palindromic repeats and complement repeats in the F. simplicissima m3 chloroplast genome. We identified 11 forward, 1 reverse, 1 complement, and 26 palindromic repeats (Supplementary File S1 Figure S1).
3.4. Phylogenetic Analysis
To examine the phylogenetic relationships within the Ficus genus and Moraceae family, ML analysis was constructed based on the similarity of chloroplast sequences and the psbA-trnH intergenic region. As illustrated in the complete cp genome (Figure 4a), species are divided into groups on the phylogenetic tree with high bootstrap values (86.20–100%). The studied F. simplicissima m3 and F. hirta (Accession number in GenBank: NC_051532.1) were located in one group with 100% support. The branch support value (86.2%) for F. pumila (NC_058617.1) and this group was lower than the other branch support values. F. benjamina (NC_053836.1) and F. lyrata (NC_053838.1) formed a well-supported monophyletic group.

Figure 4.
Phylogenetic relationships among Moraceae family based on complete cp genome (a) and psbA-trnH intergenic region (b).
The phylogenetic tree inferred from the psbA-trnH data displayed six distinct groups with strong support, with the support values ranging from 89.3 to 100% (Figure 4b). Group I contained two species of outgroup Morus alba (voucher A. chaveerach 976.1, (Accession number in GenBank: MF405185.1 and voucher A. chaveerach 977.1, MF405186.1). The selected species belonging to the Ficus genus generated the five remaining groups. Almost all individuals in the same species fell into the same clade, except five F. hirta voucher. Group II included two vouchers of F. variolosa species (JQ774218.1 and JQ774174.1) with 100% bootstrap value. Three F. simplicissima species constituted group III, in which F. simplicissima voucher HSNU2014113 (KX055795.1) and F. simplicisima voucher HSNU2014119 (KX055795.1) formed one clade (bootstrap value = 92.2%) resolved as sister to the studied F. simplicissima m3 (bootstrap value = 89.3%). Five F. hirta species split into three separate groups. Group IV comprised two clades, one clade encompassed F. hirta voucher HSNU2013079 (KX055770.1) and F. hirta voucher HSNU2013080 (KX055773.1) with bootstrap value = 100, the rest were sister to F. hirta voucher HSNU2013229 (KX055778.1) with 92.5% branch support value. Three selected F. simplicissima species were the most closely related to three F. hirta species (KX0055774.1; KX055773.1; KX055778) in group V with a 94% bootstrap value.
3.5. Comparative Genomic Analysis
The junction sites in these cp genomes were relatively conserved. The length of IR ranged from 24,421 to 25,898 bp. Most of these species had a length of the LSC region of approximately 88,500 bp. By contrast, F. simplicissima m3 exhibited a larger LSC region of 91,346 bp (Figure 5) and the large single-copy (LSC) region of chloroplast DNA is highly efficient in species identification [26].
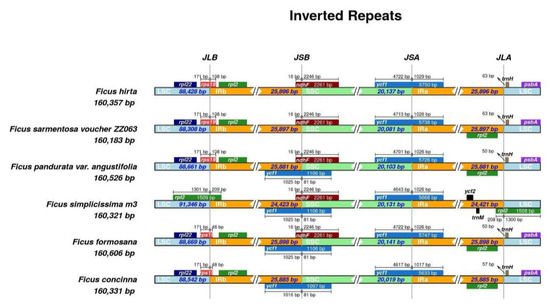
Figure 5.
Contiguous positions between LSC, SSC, and IR regions of six Ficus cp genomes.
The rps19 gene is located in the junction region between LSC and IRb (JLB), while the rpl2 gene covered this location in F. simplicissima m3. The trnH gene was shifted from JLA from 50 to 63 bp, except for F. simplicissima m3. The ycf1 gene was found to have crossed the junction located in JSA and JSB, while it was absent in the JSB of F. hirta and F. sarmentosa. The ndhF gene covered the IRB-SSC region with a similarity size of 2261 bp. However, the ndhF gene was not observed in this region of F. concinna (Figure 5).
The F. simplicissima m3 cp genome was used as a reference to analyze the cp genome identity of the six Ficus species (Figure 6). The non-coding regions were supposed to be more divergent than the coding regions. A considerable number of variations were found including ycf1, rpoC2, rpoC1, ycf2, ndhF, rps16—trnQ-UUG, trnS-UGA—trnG-GCC, trnT-UGU—trnF-GAA, petN—psbM, trnT-GGU—psbD, rpl32—trnL-UAG in the intergenic spacer regions.
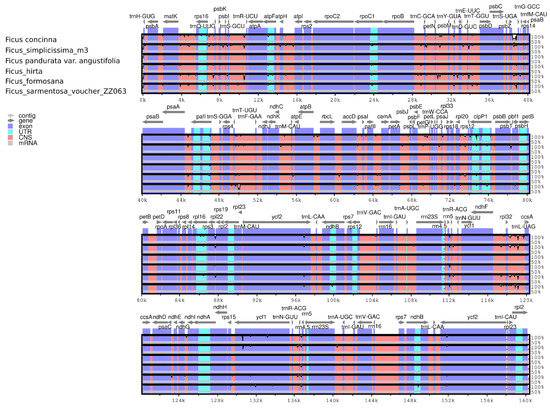
Figure 6.
Sequence alignment of chloroplast genomes among six Ficus species. The x-axis shows the coordinates in the cp genome. The y-axis shows the recognition percentage (50% to 100%).
Nucleotide diversity in LSC and SSC regions was significantly higher than that in the IR regions (Figure 6). The Pi value among six Ficus species ranged from 0 to 0.01701, with an average of 0.00306. The results showed that six highly variable regions were detected consisting of rps16—trnQ-UUG, trnC-GCA, rps14—psaB, clpB—psbB, trnL—ccsA—ndhD, rrn23S (Figure 6). Among divergent regions, five belonged to the LSC and SSC regions, only one was located in the IRa region.
4. Discussion
The gene content and genomic organization of the Ficus cp genomes were highly conserved, and no rearrangement had been found. The F. simplicissima m3 cp genome was 160,321 bp in length and had a typical quadripartite structure including an SSC, and an LSC as well as a pair of IRs. The studied cp genome sequence showed a bias toward a higher A/T ratio in composition. The GC content of the IR regions was higher than that of the non-coding intergenic regions because of the presence of rRNA genes [27]. In addition, the number of predicted genes was smaller (127) than that (131) which were previously reported in the F. simplicissima [13], F. concinna [28], and Broussonetia species [29] genomes with 15 intron-containing genes. Several genes are known to possess structural intron variation, such as atpF, rpoC2, rpl12, rps12, and rps16 [2,30]. The cp genome loses and gains introns during evolution, which plays a significant role in regulating gene expression via the alternative splicing or the stabilization of the transcript [31]. In the F. simplicissima m3 plastome, both clpP and ycf3 genes contained double introns, while 11 other protein-coding genes, six rrRNA genes and five tRNA genes contain one intron.
In this study, the use of codons in the nucleotide sequence of protein-coding genes of the chloroplast genome of F. simplicissima tends to be specific codons used more frequently than other synonymous codons. This result was consistent with previous related reports [32,33,34,35,36]. The level of use of codons varies between individuals within species and between species of the genus. RSCU is often used to reflect codon bias. In the chloroplast genome of Z. officinale, most of the preferred synonymous codons (RSCU > 1) possess A- or U-ending codons, except for trnL-CAA, which UUG encodes. Codons ending with A and/or U accounted for 71.2%, resulting in the bias for A/T bases [33]. However, most codons with an RSCU > 1 of the chloroplast genome of Litsea contained either an A- or G-terminal. By contrast, RSCU values for codons that ended with a C-terminal, such as CGC (Arg), UGC (Cys), CAC (His), and AGC (Ser), were relatively low [32,34].
Repeated sequences are involved in stabilizing and rearranging sequences in the cp genome. Repeated sequences can be used to construct molecular markers for plant identification and molecular evolutionary genetic analysis [37,38,39]. In the F. simplicissima m3 plastid, the majority of SSRs were found in the intergenic spacer regions rather than in the coding regions. They primarily consisted of AT subunits, which were similar to those in the cp genomes of angiosperms has been reported in previous works [40,41]. Besides, mono- and tri- nucleotide SSRs were more prevalent than any other type of SSRs in the studied F. simplicissima m3 species. It was reported that large and complex repeats also involved the sequence rearrangement and the evolution of cp genomes [42,43]. The REPuter analysis resulted in 39 dispersed repeats distributed mainly in the intergenic spacer and intron sequences of the F. simplicissima m3 plastome. This number was much higher than that of the recently published F. simplicissima cp genome [13]; however, the repeat type content performed similarly to the prevalent palindromic and forward repeats.
The IR regions’ size of the F. simplicissima Lour m3 was 24.4 kb, which was consistent with the data observed in most angiosperm cp genomes (20–28 kb) [44]. In most land plants, the cp genome commonly displays some significant variations, such as gene loss, sequence inversion, and expansion/contraction of the IR regions, which lead to length differences among cp genomes [45,46,47]. The sizes of the Ficus cp genomes differed with some remarkable variations in the junction regions. According to the Irscope result, there was a significant IR contraction in the F. simplicissima m3 plastome, which was a decrease in size from about 25.8 kb to 24.4 kb. The boundaries between the SSC and the IRs were similar among the Ficus species. Besides, the junctions between LSC and IRs of Ficus plastomes were commonly located within the rps19 gene (Figure 5). However, there were several cp genomes reported that the rps19 gene does not extend into the IR region [48,49]. The studied cp genome witnessed an absence of this gene in the JLA region, which might be the cause of IR contraction. On the other hand, a difference of two bp in length between IRa (24,421 bp) and IRb (24,423 bp) was predicted. The cp genome annotation tools differed notably in the number of sequences identified, in which the IR regions differed in length [50]. The ndhF gene extended over the JSB and overlapped with the ycf1 gene in half of the compared genomes, which had also been observed in some published cp genomes [13,51].
This study demonstrates that the variability of the ycf1, rpoC2, rpoC1, ycf2, and ndhF gene regions of F. simplicissima species is higher than that of the five compared species. Therefore, these gene regions can be used to elucidate the phylogenetic relationships within the Ficus genus. The Ycf1, ycf2, rpoC2, and ndhF gene regions have been confirmed to be the most disparate regions in the Ficus cp genome of the Moraceae family [52]. For the Asteraceae family, the two gene regions rpoC1 and ycf1 were also found to be the most distinct of the cp genomes of this family [53].
In the Ficus cp genomes, the sequence regions located on the genes trnL-ccsA-ndhD, trnC-GCA, and rrn23S, along with three intergenic spacer regions including rps16-trnQ, rps14-psaB and clpP-psbB, were highly variable regions and trnL-ccsA-ndhD sequence regions had the highest nucleotide diversity value (>0.015). Thus, these regions could be potential DNA barcodes for species identification.
Chloroplast genome data play an important role in species definitions due to the application of organelle-based “barcodes” to reveal the phylogenetic relationships among species [40]. The Moraceae family is known to have about 1100 species of 40 genera and they are distributed mainly in tropical and subtropical regions [54]. Currently, the study of Moraceae’s molecular evolutionary and phylogenetic analysis is limited, especially the species of the Ficus genus. Recently, Huang et al. (2022) evaluated the cp genomes of ten species in the Ficus genus. The results showed that Morus and Ficus had a close relationship compared to other genera of the Moraceae family with high bootstrap values [13,55]. In this study, the phylogenetic relationships by ML analyses were constructed based on two approaches, the complete cp genome sequence and the psbA-trnH intergenic spacer region. The two phylogenetic trees had congruent topologies (Figure 4). The outgroups, Chaetachme, Broussonetia, and Morus, clustered into monophyletic clades and were sisters to Ficus. Our study has made it clear that the complete cp genome of F. simplicissima can support genus-level identification in the Moraceae family.
At the species level, in Figure 4, Ficus were divided into two subgroups and the voucher in the same species clustered together to a certain degree. F. simplicissima and F. hirta diverged, indicating the genetic divergence between these two species and others, followed by F. variolosa. The sister relationships of the subgroups in the Ficus genus are consistent with previous reports [13]. According to Burgess et al. (2011), up to 97% accurate identification of Canadian temperate plant samples was possible based on five gene regions rbcL, matK, rpoC1, psbA-trnH, and atpF-atpH [56]. Research by Newmaster et al. (2008) showed that using matK and psbA-trnH data could identify more than 94% of species in the Myristicaceae family [57]. The study’s results using the psbA-trnH marker to identify the above plant objects are the basis for us to choose psbA-trnH for phylogenetic analysis of Ficus species. These findings supported the Berg classification system [58]. Thus, the psbA-trnH gene region sequence of the cp genome may be a potential candidate for chloroplast DNA barcoding for species-level identification in the Ficus genus. These recommendations contribute to species and genera identification based on cpDNA molecular markers and morphological support methods and help to further illustrate a monophyletic group within the Moraceae family.
5. Conclusions
In this study, the complete cp genomes of F. simplicissima were sequenced using the Pacbio sequel platform and compared with five other species of the Ficus genus in the Moraceae family. The cp genome of F. simplicissima m3 with a size of 160,321 bp contains 127 genes, including 85 protein-coding genes, eight rRNA genes, and 34 tRNA genes. Of these, 18 genes were duplicated in the IR region, and 21 contained introns. The molecular evolutionary genetics analysis results based on the complete cp genome and psbA-trnH intergenic region established the phylogenetic tree. The genetic relationship of F. simplicissima to other species in the Moraceae family was determined. The complete cp genome of F. simplicissima can support genus-level identification in the Moraceae family and the psbA-trnH gene region of the cp genome may be a potential candidate for chloroplast DNA barcoding for species-level identification in the Ficus genus.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cimb45020067/s1, Figure S1: The long repeat sequences in the cp genome of Ficus simplicissima m3; Table S1: Gene contents in the complete chloroplast genome of Ficus simplicissima m3.
Author Contributions
Conceived and designed the experiments: T.T.T.V. and M.H.C. Performed the experiments: T.T.T.V., L.T.K.V., L.T.L. and T.T.M.L. Performed analyses and wrote the article: T.T.T.V., L.T.L., L.T.K.V., T.T.M.L. and M.H.C. Made the Proof-reading: M.H.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Instances and data are available under request.
Acknowledgments
This research is funded by Thai Nguyen University of Education under grant number CS.2021.19.
Conflicts of Interest
All authors declare that the research was conducted in the absence of any commercial or financial relationships which could be construed as potential conflicts of interest.
References
- Downie, S.; Palmer, J. Use of Chloroplast DNA Rearrangements in Reconstructing Plant Phylogeny. In Molecular Systematics of Plants; Springer: Boston, MA, USA, 1992; pp. 14–35. [Google Scholar]
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- CBOL Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef]
- Yang, Z.; Zhao, T.; Ma, Q.; Liang, L.; Wang, G. Comparative genomics and phylogenetic analysis revealed the chloroplast genome variation and interspecific relationships of Corylus (Betulaceae) species. Front. Plant Sci. 2018, 9, 927. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xie, X.; Chen, A.; Li, F.; Tian, E.; Chao, Z. The chloroplast genomes of four Bupleurum (Apiaceae) species endemic to Southwestern China, a diversity center of the genus, as well as their evolutionary implications and phylogenetic inferences. BMC Genom. 2021, 22, 714. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Yan, Y.H.; Harris, A.J.; Kang, J.S.; Shen, H.; Xiang, Q.P.; Zhang, X.C. Plastid phylogenomics resolve deep relationships among eupolypod II ferns with rapid radiation and rate heterogeneity. Genome Biol. Evol. 2017, 9, 1646–1657. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Huang, W.; Sun, H.; Yer, H.; Li, X.; Li, Y.; Yan, B.; Wang, Q.; Wen, Y.; Huang, M.; et al. Comparative chloroplast genome analysis of Impatiens species (Balsaminaceae) in the karst area of China: Insights into genome evolution and phylogenomic implications. BMC Genom. 2021, 22, 571. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, D.S.; Zou, L.; Yao, C.Y. Comparison of chloroplast genomes and phylogenomics in the Ficus sarmentosa complex (Moraceae). PLoS ONE 2022, 17, e0279849. [Google Scholar] [CrossRef]
- eFloras. Missouri Botanical Garden: St. Louis, MO, USA; Harvard University Herbaria: Cambridge, MA, USA. 2008. Available online: http://www.efloras.org (accessed on 20 October 2022).
- Au, D.T.; Chen, H.; Jiang, Z.; Zhao, Z. A novel method to identify the Chinese herbal medicine Wuzhimaotao by quantification of laticifers. Microsc. Res. Tech. 2009, 72, 293–298. [Google Scholar] [CrossRef]
- Chen, J.; Tan, Y.; Wu, Z.; Liu, Z.; Zhao, W.; Wang, B.; Zhang, H.; Lin, S.; Li, J.; Lin, C.; et al. The efficacy and safety of traditional Chinese medicines, modified Radix Fici Simplicissimae, combined with Western medicines amongst patients infected with the 2019 novel coronavirus (SARS-CoV-2) in tropical tourist area, China. Integr. Med. Res. 2021, 10, 100665. [Google Scholar] [CrossRef]
- Corner, E.J.H. Check-list of Ficus in Asia and Australasia with Key; to Identification. Gard. Bull. Singap. 1965, 21, 1–186. Available online: https://biostor.org/reference/248325 (accessed on 20 October 2022).
- Huang, Y.; Li, J.; Yang, Z.; An, W.; Xie, C.; Liu, S.; Zheng, X. Comprehensive analysis of complete chloroplast genome and phylogenetic aspects of ten Ficus species. BMC Plant Biol. 2022, 22, 253. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.; Berlin, K.; Miller, J.; Bergman, N.; Phillippy, A. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A Program for Improved Detection of Transfer RNA Genes in Genomic Sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Kurtz, S.; Schleiermacher, C. REPuter: Fast computation of maximal repeats in complete genomes. Bioinformatics 1999, 15, 426–427. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32 (Suppl. S2), W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Figtree v1.4.4. Available online: https://github.com/rambaut/figtree/releases (accessed on 20 December 2022).
- Gebhardt, C. Chapter 5—Molecular Markers, Maps and Population Genetics; Vreugdenhil, D., Bradshaw, J., Gebhardt, C., Govers, F., Mackerron, D.K.L., Taylor, M.A., Ross, B., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2007; pp. 77–89. [Google Scholar] [CrossRef]
- Li, L.; Jiang, Y.; Liu, Y.; Niu, Z.; Xue, Q.; Liu, W.; Ding, X. The large single-copy (LSC) region functions as a highly effective and efficient molecular marker for accurate authentication of medicinal Dendrobium species. Acta Pharm. Sin. B 2020, 10, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Bock, R. Structure, function, and inheritance of plastid genomes. In Cell and Molecular Biology of Plastids; Springer: Berlin/Heidelberg, Germany, 2007; pp. 29–63. [Google Scholar]
- Zhang, P.-H.; Hou, Z. The complete chloroplast genome sequence of Ficus concinna (Moraceae) from Sichuan province. Mitochondrial DNA Part B 2022, 7, 390–392. [Google Scholar] [CrossRef]
- Yang, J.; Chu, Q.; Meng, G.; Kong, W. The complete chloroplast genome sequences of three Broussonetia species and comparative analysis within the Moraceae. PeerJ 2022, 10, e14293. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Qian, J.; Li, X.; Sun, Z.; Xu, X.; Chen, S. Complete Chloroplast Genome of Medicinal Plant Lonicera japonica: Genome Rearrangement, Intron Gain and Loss, and Implications for Phylogenetic Studies. Molecules 2017, 22, 249. [Google Scholar] [CrossRef]
- Daniell, H.; Wurdack, K.J.; Kanagaraj, A.; Lee, S.-B.; Saski, C.; Jansen, R.K. The complete nucleotide sequence of the cassava (Manihot esculenta) chloroplast genome and the evolution of atpF in Malpighiales: RNA editing and multiple losses of a group II intron. Theor. Appl. Genet. 2008, 116, 723–737. [Google Scholar] [CrossRef]
- Zuo, L.H.; Shang, A.Q.; Zhang, S.; Yu, X.Y.; Ren, Y.C.; Yang, M.S.; Wang, J.M. The first complete chloroplast genome sequences of Ulmus species by de Novo sequencing: Genome comparative and taxonomic position analysis. PLoS ONE 2017, 12, e0171264. [Google Scholar] [CrossRef]
- Cui, Y.; Nie, L.; Sun, W.; Xu, Z.; Wang, Y.; Yu, J.; Song, J.; Yao, H. Comparative and phylogenetic analyses of ginger (Zingiber officinale) in the family Zingiberaceae based on the complete chloroplast genome. Plants 2019, 8, 283. [Google Scholar] [CrossRef]
- Song, W.; Chen, Z.; Shi, W.; Han, W.; Feng, Q.; Shi, C.; Engel, M.S.; Wang, S. Comparative Analysis of Complete Chloroplast Genomes of Nine Species of Litsea (Lauraceae): Hypervariable Regions, Positive Selection, and Phylogenetic Relationships. Genes 2022, 13, 1550. [Google Scholar] [CrossRef]
- Song, W.; Ji, C.; Chen, Z.; Cai, H.; Wu, X.; Shi, C.; Wang, S. Comparative analysis the complete chloroplast genomes of nine Musa Species: Genomic features, comparative analysis, and phylogenetic implications. Front. Plant Sci. 2022, 13, 832884. [Google Scholar] [CrossRef]
- Trofimov, D.; Cadar, D.; Schmidt-Chanasit, J.; Moraes, P.L.R.; Rohwe, J.G. A comparative analysis of complete chloroplast genomes of seven Ocotea species (Lauraceae) confirms low sequence divergence within the Ocotea complex. Sci. Rep. 2022, 12, 1120. [Google Scholar] [CrossRef]
- Vieira, L.D.N.; Faoro, H.; Rogalski, M.; Fraga, H.P.D.F.; Cardoso, R.L.A.; de Souza, E.M.; de Oliveira Pedrosa, F.; Nodari, R.O.; Guerra, M.P. The complete chloroplast genome sequence of Podocarpus lambertii: Genome structure, evolutionary aspects, gene content and SSR detection. PLoS ONE 2014, 9, e90618. [Google Scholar] [CrossRef]
- Cho, K.-S.; Cheon, K.-S.; Hong, S.-Y.; Cho, J.-H.; Im, J.-S.; Mekapogu, M.; Yu, Y.-S.; Park, T.-H. Complete chloroplast genome sequences of Solanum commersonii and its application to chloroplast genotype in somatic hybrids with Solanum tuberosum. Plant Cell Rep. 2016, 35, 2113–2123. [Google Scholar] [CrossRef]
- Williams, A.V.; Miller, J.T.; Small, I.; Nevill, P.G.; Boykin, L.M. Integration of complete chloroplast genome sequences with small amplicon datasets improves phylogenetic resolution in Acacia. Mol. Phylogenet. Evol. 2016, 96, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.L.; Al-Harrasi, A.; Asaf, S.; Park, C.E.; Park, G.-S.; Khan, A.R.; Lee, I.-J.; Al-Rawahi, A.; Shin, J.-H. The First Chloroplast Genome Sequence of Boswellia sacra, a Resin-Producing Plant in Oman. PLoS ONE 2017, 12, e0169794. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.Q.; Nguyen, T.N.L.; Doan, T.N.; Nguyen, T.T.N.; Phạm, M.H.; Le, T.L.; Sy, D.T.; Chu, H.H.; Chu, H.M. Complete chloroplast genome of novel Adinandra megaphylla Hu species: Molecular structure, comparative and phylogenetic analysis. Sci. Rep. 2021, 11, 11731. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Chloroplast evolution: Secondary symbiogenesis and multiple losses. Curr. Biol. 2002, 12, R62–R64. [Google Scholar] [CrossRef] [PubMed]
- Bausher, M.G.; Singh, N.D.; Lee, S.-B.; Jansen, R.K.; Daniell, H. The complete chloroplast genome sequence of Citrus sinensis (L.) Osbeck var “Ridge Pineapple”: Organization and phylogenetic relationships to other angiosperms. BMC Plant Biol. 2006, 6, 21. [Google Scholar] [CrossRef]
- Chumley, T.W.; Palmer, J.D.; Mower, J.P.; Fourcade, H.M.; Calie, P.J.; Boore, J.L.; Jansen, R.K. The Complete Chloroplast Genome Sequence of Pelargonium × hortorum: Organization and Evolution of the Largest and Most Highly Rearranged Chloroplast Genome of Land Plants. Mol. Biol. Evol. 2006, 23, 2175–2190. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Chung, M.G.; Park, S. The Complete Chloroplast Genome Sequences of Three Veroniceae Species (Plantaginaceae): Comparative Analysis and Highly Divergent Regions. Front. Plant Sci. 2016, 7, 355. [Google Scholar] [CrossRef]
- Fu, P.-C.; Zhang, Y.-Z.; Geng, H.-M.; Chen, S.-L. The complete chloroplast genome sequence of Gentiana lawrencei var farreri (Gentianaceae) and comparative analysis with its congeneric species. PeerJ 2016, 4, e2540. [Google Scholar] [CrossRef]
- Fan, R.; Ma, W.; Liu, S.; Huang, Q. Integrated analysis of three newly sequenced fern chloroplast genomes: Genome structure and comparative analysis. Ecol. Evol. 2021, 11, 4550–4563. [Google Scholar] [CrossRef]
- Kazakoff, S.H.; Imelfort, M.; Edwards, D.; Koehorst, J.; Biswas, B.; Batley, J.; Scott, P.T.; Gresshoff, P.M. Capturing the biofuel wellhead and powerhouse: The chloroplast and mitochondrial genomes of the leguminous feedstock tree Pongamia pinnata. PLoS ONE 2012, 7, e51687. [Google Scholar] [CrossRef]
- Martin, G.E.; Rousseau-Gueutin, M.; Cordonnier, S.; Lima, O.; Michon-Coudouel, S.; Naquin, D.; de Carvalho, J.F.; Aïnouche, M.; Salmon, A.; Aïnouche, A. The first complete chloroplast genome of the Genistoid legume Lupinus luteus: Evidence for a novel major lineage-specific rearrangement and new insights regarding plastome evolution in the legume family. Ann. Bot. 2014, 113, 1197–1210. [Google Scholar] [CrossRef]
- Turudić, A.; Liber, Z.; Grdiša, M.; Jakše, J.; Varga, F.; Šatović, Z. Chloroplast Genome Annotation Tools: Prolegomena to the Identification of Inverted Repeats. Int. J. Mol. Sci. 2022, 23, 10804. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Asaf, S.; Khan, A.L.; Khan, A.; Al-Harrasi, A.; Al-Sudairy, O.; AbdulKareem, N.M.; Al-Saady, N.; Al-Rawahi, A. Complete chloroplast genomes of medicinally important Teucrium species and comparative analyses with related species from Lamiaceae. PeerJ 2019, 7, e7260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, X.; Li, W.; Peng, Y.; Gao, J. Comparative chloroplast genome analysis of Ficus (Moraceae): Insight into adaptive evolution and mutational hotspot regions. Front. Plant Sci. 2022, 13, 965335. [Google Scholar] [CrossRef]
- Nie, X.; Lv, S.; Zhang, Y.; Du, X.; Wang, L.; Biradar, S.S.; Tan, X.; Wan, F.; Weining, S. Complete chloroplast genome sequence of a major invasive species, crofton weed (Ageratina adenophora). PLoS ONE 2012, 7, e36869. [Google Scholar] [CrossRef]
- Clement, W.L.; Weiblen, G.D. Morphological Evolution in the Mulberry Family (Moraceae). Syst. Bot. 2009, 34, 530–552. [Google Scholar] [CrossRef]
- Xia, X.; Peng, J.; Yang, L.; Zhao, X.; Anan Duan, A.; Wang, D. Comparative Analysis of the Complete Chloroplast Genomes of Eight Ficus Species and Insights into the Phylogenetic Relationships of Ficus. Life 2022, 12, 848. [Google Scholar] [CrossRef] [PubMed]
- Burgess, K.S.; Fazekas, A.J.; Kesanakurti, P.R.; Graham, S.W.; Husband, B.C.; Newmaster, S.G.; Percy, D.M.; Hajibabaei, M.; Barrett, S.C.H. Discriminating plant species in a local temperate flora using the rbcL+matK DNA barcode. Methods Ecol. Evol. 2011, 2, 333–340. [Google Scholar] [CrossRef]
- Newmaster, S.G.; Fazekas, A.J.; Steeves, R.A.; Janovec, J. Testing candidate plant barcode regions in the Myristicaceae. Mol. Ecol. Resour. 2008, 8, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Berg, C.; Corner, E. Moraceae (Ficus). In Flora Malesiana; Nooteboom, H., Ed.; National Herbarium of the Netherlands: Leiden, The Netherlands, 2005; Volume 17, pp. 1–70. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).