
Zinc Finger E-Box Binding Homeobox 2 as a Prognostic Biomarker in Various Cancers and Its Correlation with Infiltrating Immune Cells in Ovarian Cancer


Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Expression Profiling Interactive Analysis (GEPIA)
2.2. Kaplan–Meier Plotter Database Analysis
2.3. PrognoScan Database Analysis
2.4. Tumor Immune Estimation Resource (TIMER) Analysis
2.5. STRING Analysis
2.6. Statistical Analysis
3. Results
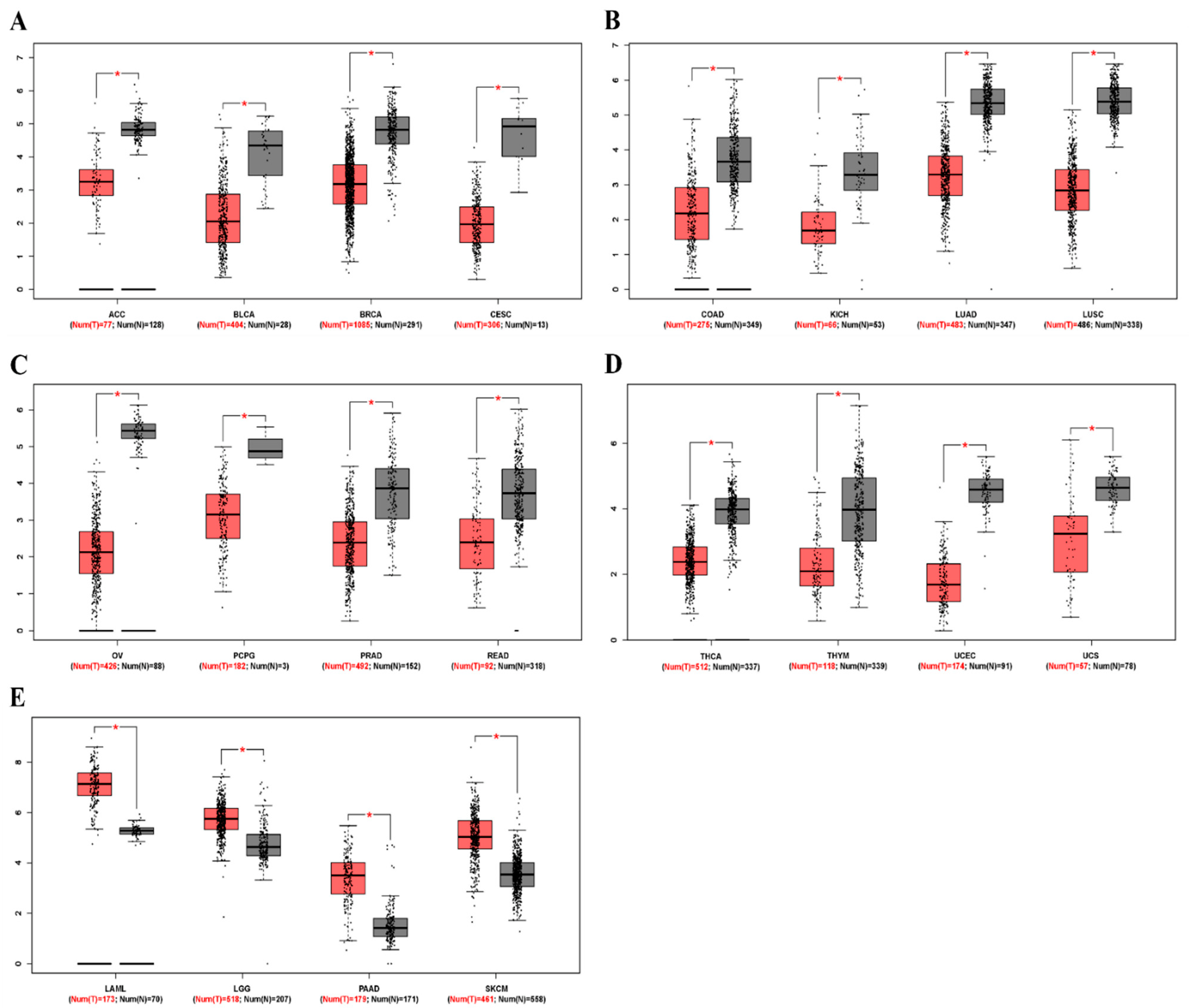
3.1. mRNA Levels of ZEB2 in Various Types of Cancer
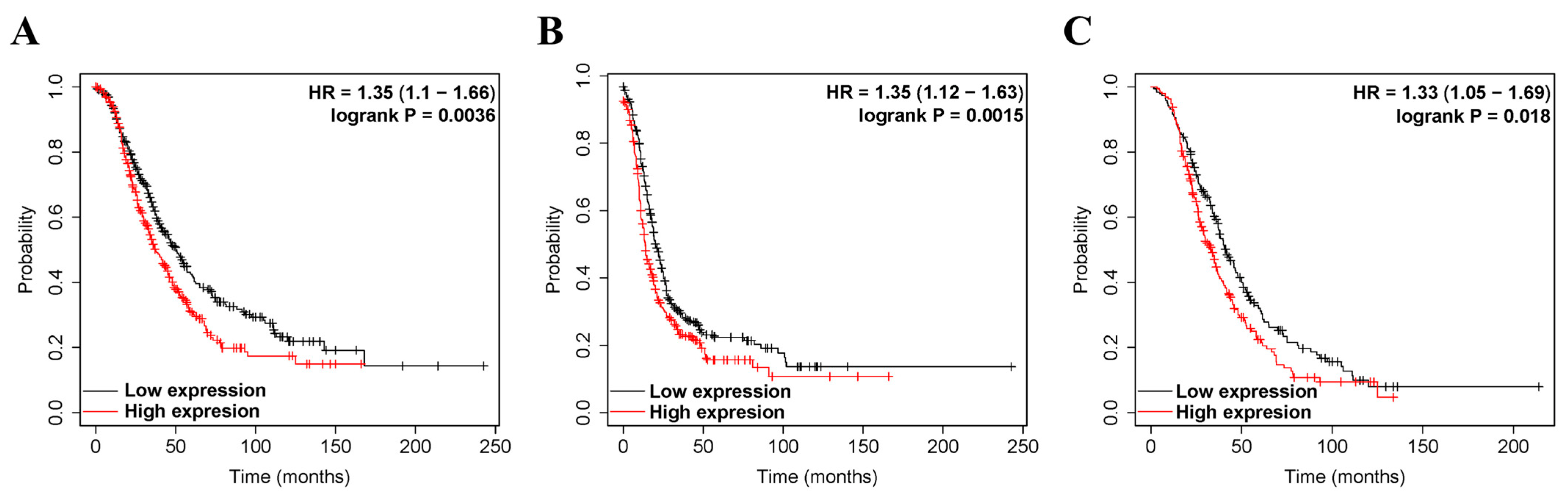
3.2. The Prognostic Value of ZEB2 Expression in Various Types of Cancer
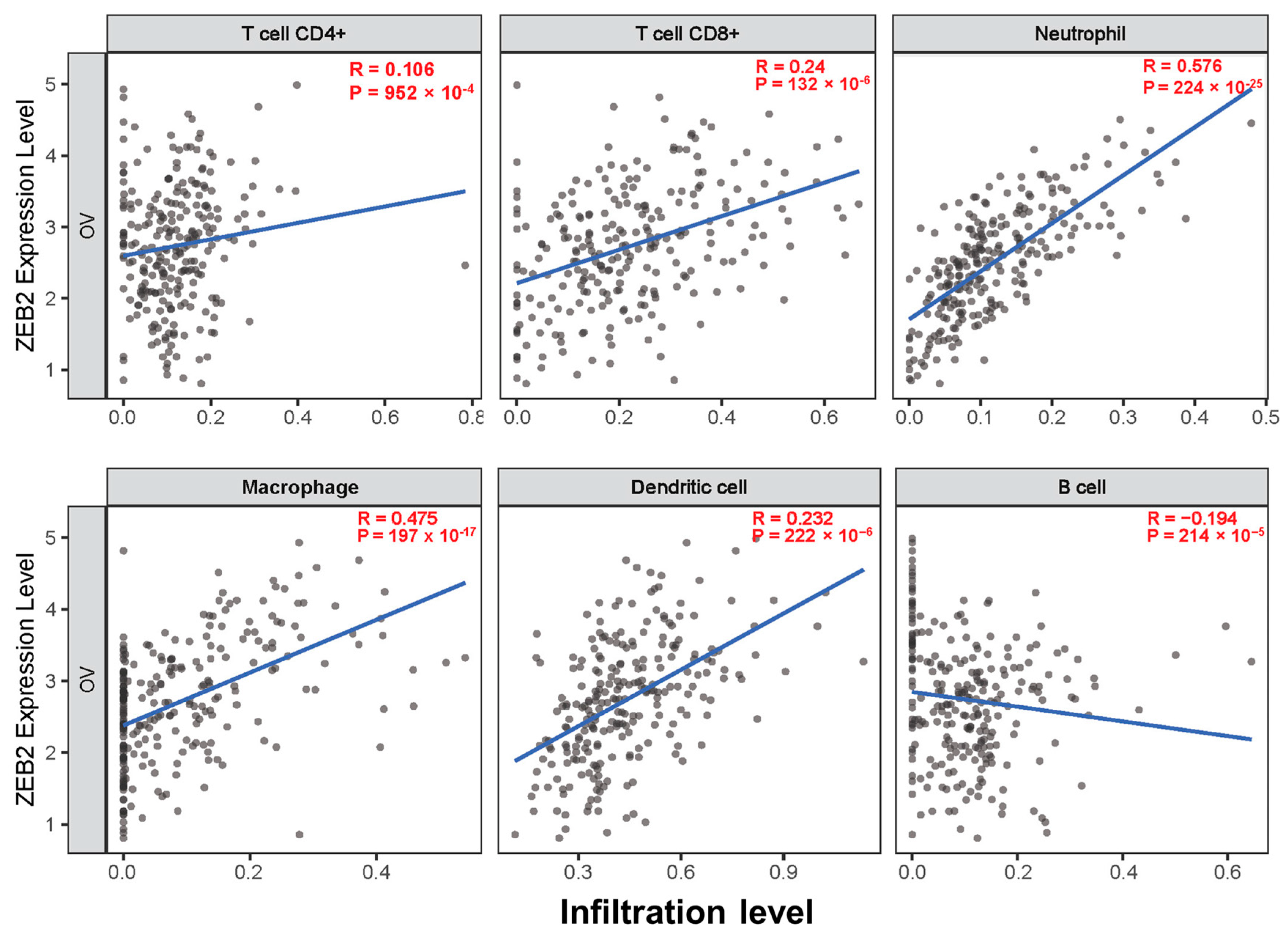
3.3. Correlation between ZEB2 and Infiltrating Immune Cells in Various Types of Cancer
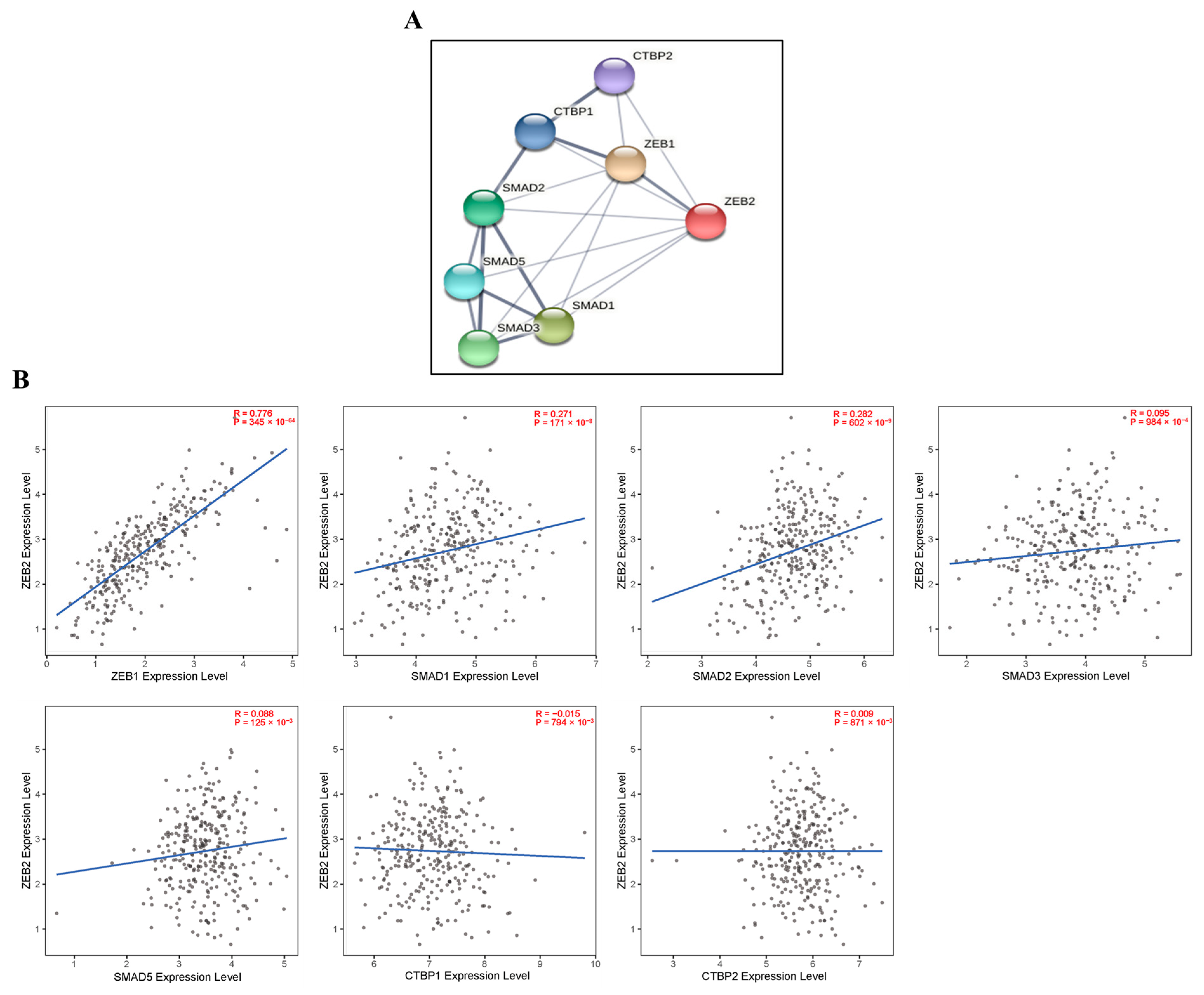
3.4. Correlation between ZEB1 and ZEB2 Expression in Various Types of Cancer
3.5. Correlations with ZEB2-Related Proteins in Various Types of Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roett, M.A.; Evans, P. Ovarian cancer: An overview. Am. Fam. Physician 2009, 80, 609–616. [Google Scholar] [PubMed]
- Lisio, M.A.; Fu, L.; Goyeneche, A.; Gao, Z.H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [Green Version]
- Odunsi, K. Immunotherapy in ovarian cancer. Ann. Oncol. 2017, 28, viii1–viii7. [Google Scholar] [CrossRef] [PubMed]
- Berek, J.S.; Kehoe, S.T.; Kumar, L.; Friedlander, M. Cancer of the ovary, fallopian tube, and peritoneum. Int. J. Gynaecol. Obstet. 2018, 143, 59–78. [Google Scholar] [CrossRef]
- Elias, K.M.; Guo, J.; Bast, R.C., Jr. Early Detection of Ovarian Cancer. Hematol. Oncol. Clin. N. Am. 2018, 32, 903–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.P.; Lin, Q.; Mu, X.L. MicroRNA-155 and FOXP3 jointly inhibit the migration and invasion of colorectal cancer cells by regulating ZEB2 expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6131–6138. [Google Scholar]
- Ma, F.; Li, W.; Liu, C.; Li, W.; Yu, H.; Lei, B.; Ren, Y.; Li, Z.; Pang, D.; Qian, C. MiR-23a promotes TGF-beta1-induced EMT and tumor metastasis in breast cancer cells by directly targeting CDH1 and activating Wnt/beta-catenin signaling. Oncotarget 2017, 8, 69538–69550. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhou, W.; Han, Y.; Peng, F.; Wang, R.; Yu, R.; Wang, C.; Liang, H.; Guo, Z.; Gu, Y. EMT-Regulome: A database for EMT-related regulatory interactions, motifs and network. Cell Death Dis. 2017, 8, e2872. [Google Scholar] [CrossRef] [Green Version]
- Chaffer, C.L.; San Juan, B.P.; Lim, E.; Weinberg, R.A. EMT, cell plasticity and metastasis. Cancer Metastasis Rev. 2016, 35, 645–654. [Google Scholar] [CrossRef]
- Liang, H.; Yu, T.; Han, Y.; Jiang, H.; Wang, C.; You, T.; Zhao, X.; Shan, H.; Yang, R.; Yang, L.; et al. LncRNA PTAR promotes EMT and invasion-metastasis in serous ovarian cancer by competitively binding miR-101-3p to regulate ZEB1 expression. Mol. Cancer 2018, 17, 119. [Google Scholar] [CrossRef]
- Wang, P.; Liu, X.; Han, G.; Dai, S.; Ni, Q.; Xiao, S.; Huang, J. Downregulated lncRNA UCA1 acts as ceRNA to adsorb microRNA-498 to repress proliferation, invasion and epithelial mesenchymal transition of esophageal cancer cells by decreasing ZEB2 expression. Cell Cycle 2019, 8, 2359–2376. [Google Scholar] [CrossRef] [PubMed]
- Fardi, M.; Alivand, M.; Baradaran, B.; Farshdousti Hagh, M.; Solali, S. The crucial role of ZEB2: From development to epithelial-to-mesenchymal transition and cancer complexity. J. Cell Physiol. 2019. Online ahead of print. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, X.; Ge, G.; Zang, X.; Shao, M.; Zou, S.; Zhang, Y.; Mao, Z.; Zhang, J.; Mao, F.; et al. miR-498 inhibits the growth and metastasis of liver cancer by targeting ZEB2. Oncol. Rep. 2019, 41, 1638–1648. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wang, W.; Lin, X.; Zheng, X.; Yang, A. Roles of ZEB2 and RBM38 in liver cancer stem cell proliferation. J. BUON 2020, 25, 1390–1394. [Google Scholar]
- Feng, S.; Liu, W.; Bai, X.; Pan, W.; Jia, Z.; Zhang, S.; Zhu, Y.; Tan, W. LncRNA-CTS promotes metastasis and epithelial-to-mesenchymal transition through regulating miR-505/ZEB2 axis in cervical cancer. Cancer Lett. 2019, 465, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhu, W.; Yang, R.; Xie, W.; Wang, D. LncRNA ZEB2-AS1 contributes to the tumorigenesis of gastric cancer via activating the Wnt/beta-catenin pathway. Mol. Cell. Biochem. 2019, 456, 73–83. [Google Scholar] [CrossRef]
- Meng, L.; Yue, X.; Zhou, D.; Li, H. Long non coding RNA OIP5-AS1 promotes metastasis of breast cancer via miR-340-5p/ZEB2 axis. Oncol. Rep. 2020, 44, 1662–1670. [Google Scholar] [CrossRef]
- Li, Y.; Fei, H.; Lin, Q.; Liang, F.; You, Y.; Li, M.; Wu, M.; Qu, Y.; Li, P.; Yuan, Y.; et al. ZEB2 facilitates peritoneal metastasis by regulating the invasiveness and tumorigenesis of cancer stem-like cells in high-grade serous ovarian cancers. Oncogene 2021, 40, 5131–5141. [Google Scholar] [CrossRef]
- Prislei, S.; Martinelli, E.; Zannoni, G.F.; Petrillo, M.; Filippetti, F.; Mariani, M.; Mozzetti, S.; Raspaglio, G.; Scambia, G.; Ferlini, C. Role and prognostic significance of the epithelial-mesenchymal transition factor ZEB2 in ovarian cancer. Oncotarget 2015, 6, 18966–18979. [Google Scholar] [CrossRef] [Green Version]
- Scott, C.L.; Omilusik, K.D. ZEBs: Novel Players in Immune Cell Development and Function. Trends Immunol. 2019, 40, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Lai, J.; Dai, D.; Chen, R.; Li, X.; Liao, N. JAK1 as a prognostic marker and its correlation with immune infiltrates in breast cancer. Aging 2019, 11, 11124–11135. [Google Scholar] [CrossRef]
- Gu, Y.; Li, X.; Bi, Y.; Zheng, Y.; Wang, J.; Li, X.; Huang, Z.; Chen, L.; Huang, Y.; Huang, Y. CCL14 is a prognostic biomarker and correlates with immune infiltrates in hepatocellular carcinoma. Aging 2020, 12, 784–807. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, H.; Kitada, K.; Nakai, K.; Sarai, A. PrognoScan: A new database for meta-analysis of the prognostic value of genes. BMC Med. Genom. 2009, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Reddel, R.R. Telomere maintenance mechanisms in cancer: Clinical implications. Curr. Pharm. Des. 2014, 20, 6361–6374. [Google Scholar] [CrossRef] [Green Version]
- Japanese Gastric Cancer Association. Japanese gastric cancer treatment guidelines 2014 (ver. 4). Gastric Cancer 2017, 20, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, H. Focus on TILs: Prognostic significance of tumor infiltrating lymphocytes in human colorectal cancer. Cancer Immun. 2007, 7, 4. [Google Scholar]
- Soen, B.; Vandamme, N.; Berx, G.; Schwaller, J.; Van Vlierberghe, P.; Goossens, S. ZEB Proteins in Leukemia: Friends, Foes, or Friendly Foes? Hemasphere 2018, 2, e43. [Google Scholar] [CrossRef]
- Katsura, A.; Tamura, Y.; Hokari, S.; Harada, M.; Morikawa, M.; Sakurai, T.; Takahashi, K.; Mizutani, A.; Nishida, J.; Yokoyama, Y.; et al. ZEB1-regulated inflammatory phenotype in breast cancer cells. Mol. Oncol. 2017, 11, 1241–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Ma, Q.; Zhang, M.; Wang, X.; Zhang, D.; Li, W.; Wang, F.; Wu, E. Alterations of TP53 are associated with a poor outcome for patients with hepatocellular carcinoma: Evidence from a systematic review and meta-analysis. Eur. J. Cancer 2012, 48, 2328–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, F.; Tian, W.Y.; Wang, Y.M.; Zhang, Y.F.; Guo, F.; Zhao, J.; Gao, C.; Xue, F.X. Cancer-associated fibroblasts promote the progression of endometrial cancer via the SDF-1/CXCR4 axis. J. Hematol. Oncol. 2016, 9, 8–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, P.L.P.; Cordeiro, Y.G.; Rochetti, A.L.; Sangalli, J.R.; Zuccari, D.A.P.C.; Silveira, J.C.; Bressan, F.F.; Fukumasu, H. ZEB1 and ZEB2 transcription factors are potential therapeutic targets of canine mammary cancer cells. Vet. Comp. Oncol. 2018, 16, 596–605. [Google Scholar] [CrossRef] [PubMed]
- van Helden, M.J.; Goossens, S.; Daussy, C.; Mathieu, A.L.; Faure, F.; Marçais, A.; Vandamme, N.; Farla, N.; Mayol, K.; Viel, S.; et al. Terminal NK cell maturation is controlled by concerted actions of T-bet and Zeb2 and is essential for melanoma rejection. J. Exp. Med. 2015, 212, 2015–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez, C.X.; Amezquita, R.A.; Guan, T.; Marshall, H.D.; Joshi, N.S.; Kleinstein, S.H.; Kaech, S.M. The transcription factors ZEB2 and T-bet cooperate to program cytotoxic T cell terminal differentiation in response to LCMV viral infection. J. Exp. Med. 2015, 212, 2041–2056. [Google Scholar] [CrossRef] [Green Version]
- Omilusik, K.D.; Best, J.A.; Yu, B.; Goossens, S.; Weidemann, A.; Nguyen, J.V.; Seuntjens, E.; Stryjewska, A.; Zweier, C.; Roychoudhuri, R.; et al. The transcriptional repressor ZEB2 promotes the terminal differentiation of CD8+ effector and memory T cell populations during infection. J. Exp. Med. 2015, 212, 2027–2039. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Sun, Y.; Ma, L. ZEB1: At the crossroads of epithelial-mesenchymal transition, metastasis and therapy resistance. Cell Cycle 2015, 14, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Vandewalle, C.; Comijn, J.; De Craene, B.; Vermassen, P.; Bruyneel, E.; Andersen, H.; Tulchinsky, E.; Van Roy, F.; Berx, G. SIP1/ZEB2 induces EMT by repressing genes of different epithelial cell-cell junctions. Nucleic Acids Res. 2005, 33, 6566–6578. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [Green Version]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heldin, C.H.; Vanlandewijck, M.; Moustakas, A. Regulation of EMT by TGFbeta in cancer. FEBS Lett. 2012, 586, 1959–1970. [Google Scholar] [CrossRef] [Green Version]
- Miyazono, K.; Ehata, S.; Koinuma, D. Tumor-promoting functions of transforming growth factor-beta in progression of cancer. J. Med. Sci. 2012, 117, 143–152. [Google Scholar]
- Diaferia, G.R.; Balestrieri, C.; Prosperini, E.; Nicoli, P.; Spaggiari, P.; Zerbi, A.; Natoli, G. Dissection of transcriptional and cis-regulatory control of differentiation in human pancreatic cancer. EMBO J. 2016, 35, 595–617. [Google Scholar] [CrossRef] [Green Version]
- Krebs, A.M.; Mitschke, J.; Lasierra Losada, M.; Schmalhofer, O.; Boerries, M.; Busch, H.; Boettcher, M.; Mougiakakos, D.; Reichardt, W.; Bronsert, P.; et al. The EMT-activator Zeb1 is a key factor for cell plasticity and promotes metastasis in pancreatic cancer. Nat. Cell Biol. 2017, 19, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verschueren, K.; Remacle, J.E.; Collart, C.; Kraft, H.; Baker, B.S.; Tylzanowski, P.; Nelles, L.; Wuytens, G.; Su, M.T.; Bodmer, R.; et al. SIP1, a novel zinc finger/homeodomain repressor, interacts with Smad proteins and binds to 5′-CACCT sequences in candidate target genes. Biol. Chem. 1999, 274, 20489–20498. [Google Scholar] [CrossRef] [Green Version]
- Wen, H.; Qian, M.; He, J.; Li, M.; Yu, Q.; Leng, Z. Inhibiting of self-renewal, migration and invasion of ovarian cancer stem cells by blocking TGF-beta pathway. PLoS ONE 2020, 15, e0230230. [Google Scholar]
- Postigo, A.A.; Dean, D.C. ZEB represses transcription through interaction with the corepressor CtBP. Proc. Natl. Acad. Sci. USA 1999, 96, 6683–6688. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Characteristics | Progression-Free Survival
(n = 614) | Overall Survival (n = 1144) | Post-progression Survival
(n = 138) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | Hazard Ratio | p-Value | n | Hazard Ratio | p-Value | n | Hazard Ratio | p-Value | |
| STAGE | |||||||||
| I | 74 | 1.3 (0.36–4.7) | 0.68 | 51 | 1.57 (0.39–6.32) | 0.52 | 7 | - | - |
| I+ II | 115 | 2.13 (0.94–4.8) | 0.064 | 83 | 1.2 (0.43–3.4) | 0.73 | 20 | 1.85 (0.52–6.7) | 0.33 |
| II | 14 | 2.5 (1.0–6.4) | 0.043 | 32 | 2.7 (0.52–14) | 0.22 | 13 | 5.26 (0.59–48) | 0.093 |
| II + III | 465 | 1.34 (1.09–1.64) | 0.0056 | 458 | 1.21 (0.95–1.54) | 0.13 | 325 | 1.31 (1.01–1.7) | 0.041 |
| II + III + IV | 535 | 1.33 (1.1–1.61) | 0.0034 | 519 | 1.29 (1.03–1.61) | 0.025 | 374 | 1.29 (1.02–1.64) | 0.034 |
| III | 424 | 1.29 (1.05–1.6) | 0.017 | 426 | 1.26 (0.99–1.62) | 0.063 | 312 | 1.25 (0.96–1.62) | 0.097 |
| III + IV | 494 | 1.37 (1.12–1.66) | 0.0017 | 487 | 1.26 (1.0–1.58) | 0.048 | 361 | 1.22 (0.96–1.56) | 0.099 |
| IV | 70 | 1.65 (0.99–2.75) | 0.05 | 61 | 1.13 (0.63–2.01) | 0.69 | 49 | 1.1 (0.59–2.1) | 0.76 |
| GRADE | |||||||||
| I | 54 | 4.16 (0.86–20) | 0.054 | 41 | 1.12 (0.39–3.22) | 0.84 | 9 | - | - |
| I+ II | 189 | 1.38 (0.97–1.96) | 0.074 | 203 | 1.17 (0.78–1.75) | 0.44 | 118 | 1.12 (0.72–1.74) | 0.62 |
| II | 161 | 1.27 (0.88–1.83) | 0.19 | 162 | 1.18 (0.76–1.83) | 0.46 | 109 | 1.09 (0.68–1.74) | 0.71 |
| II + III | 476 | 1.33 (1.08–1.64) | 0.0061 | 554 | 1.31 (1.05–1.62) | 0.016 | 349 | 1.31 (1.02–1.68) | 0.033 |
| III | 315 | 1.38 (1.07–1.77) | 0.012 | 392 | 1.29 (1.01–1.66) | 0.044 | 240 | 1.37 (1.02–1.84) | 0.035 |
| IV | 18 | - | - | 18 | 1.06 (0.39–2.9) | 0.91 | 18 | - | - |
| TP53 mutation | |||||||||
| Mutated | 124 | 1.26 (0.87–1.84) | 0.22 | 124 | 1.18 (0.81–1.73) | 0.38 | 116 | 1.02 (0.7–1.5) | 0.92 |
| Wild type | 19 | 2.2 (0.77–6.33) | 0.13 | 19 | 2.83 (0.98–8.16) | 0.046 | 17 | 2.02 (0.7–5.8) | 0.18 |
| Cancer Type | R | p |
|---|---|---|
| Adrenocortical carcinoma | 0.30 | 0.007 |
| Bladder urothelial carcinoma | 0.82 | 0.000 |
| Breast invasive carcinoma | 0.80 | 0.000 |
| Cervical squamous cell carcinoma andendocervical adenocarcinoma | 0.71 | 0.000 |
| Cholangiocarcinoma | 0.70 | 0.000 |
| Colon adenocarcinoma | 0.85 | 0.000 |
| Lymphoid neoplasm diffuse large B-cellLymphoma | 0.79 | 0.000 |
| Esophageal carcinoma | 0.87 | 0.000 |
| Glioblastoma multiforme | 0.49 | 0.000 |
| Head and neck squamous cell carcinoma | 0.87 | 0.000 |
| Kidney chromophobe | 0.71 | 0.000 |
| Kidney renal clear cell carcinoma | 0.75 | 0.000 |
| Kidney renal papillary cell carcinoma | 0.47 | 0.000 |
| Brain lower grade glioma | 0.54 | 0.000 |
| Liver hepatocellular carcinoma | 0.57 | 0.000 |
| Lung adenocarcinoma | 0.78 | 0.000 |
| Lung squamous cell carcinoma | 0.76 | 0.000 |
| Mesothelioma | 0.51 | 0.000 |
| Ovarian serous cystadenocarcinoma | 0.78 | 0.000 |
| Pancreatic adenocarcinoma | 0.88 | 0.000 |
| Pheochromocytoma and paraganglioma | 0.12 | 0.097 |
| Prostate adenocarcinoma | 0.86 | 0.000 |
| Rectum adenocarcinoma | 0.89 | 0.000 |
| Sarcoma | 0.14 | 0.029 |
| Skin cutaneous melanoma | 0.20 | 0.000 |
| Stomach adenocarcinoma | 0.82 | 0.000 |
| Testicular germ cell tumors | 0.76 | 0.000 |
| Thyroid carcinoma | 0.69 | 0.000 |
| Thymoma | 0.22 | 0.017 |
| Uterine corpus endometrial carcinoma | 0.71 | 0.000 |
| Uterine carcinosarcoma | 0.52 | 0.000 |
| Uveal melanoma | 0.61 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-R.; Seo, C.W.; Han, S.J.; Lee, J.-H.; Kim, J. Zinc Finger E-Box Binding Homeobox 2 as a Prognostic Biomarker in Various Cancers and Its Correlation with Infiltrating Immune Cells in Ovarian Cancer. Curr. Issues Mol. Biol. 2022, 44, 1203-1214. https://doi.org/10.3390/cimb44030079
Kim H-R, Seo CW, Han SJ, Lee J-H, Kim J. Zinc Finger E-Box Binding Homeobox 2 as a Prognostic Biomarker in Various Cancers and Its Correlation with Infiltrating Immune Cells in Ovarian Cancer. Current Issues in Molecular Biology. 2022; 44(3):1203-1214. https://doi.org/10.3390/cimb44030079
Chicago/Turabian StyleKim, Hye-Ran, Choong Won Seo, Sang Jun Han, Jae-Ho Lee, and Jongwan Kim. 2022. "Zinc Finger E-Box Binding Homeobox 2 as a Prognostic Biomarker in Various Cancers and Its Correlation with Infiltrating Immune Cells in Ovarian Cancer" Current Issues in Molecular Biology 44, no. 3: 1203-1214. https://doi.org/10.3390/cimb44030079
APA StyleKim, H.-R., Seo, C. W., Han, S. J., Lee, J.-H., & Kim, J. (2022). Zinc Finger E-Box Binding Homeobox 2 as a Prognostic Biomarker in Various Cancers and Its Correlation with Infiltrating Immune Cells in Ovarian Cancer. Current Issues in Molecular Biology, 44(3), 1203-1214. https://doi.org/10.3390/cimb44030079