
The Small Metal-Binding Protein SmbP Improves the Expression and Purification of the Recombinant Antitumor-Analgesic Peptide from the Chinese Scorpion Buthus martensii Karsch in Escherichia coli

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Vectors, and Reagents
2.2. Construction of the Expression Vector pET30a(+)_SmbP_BmK-AGAP
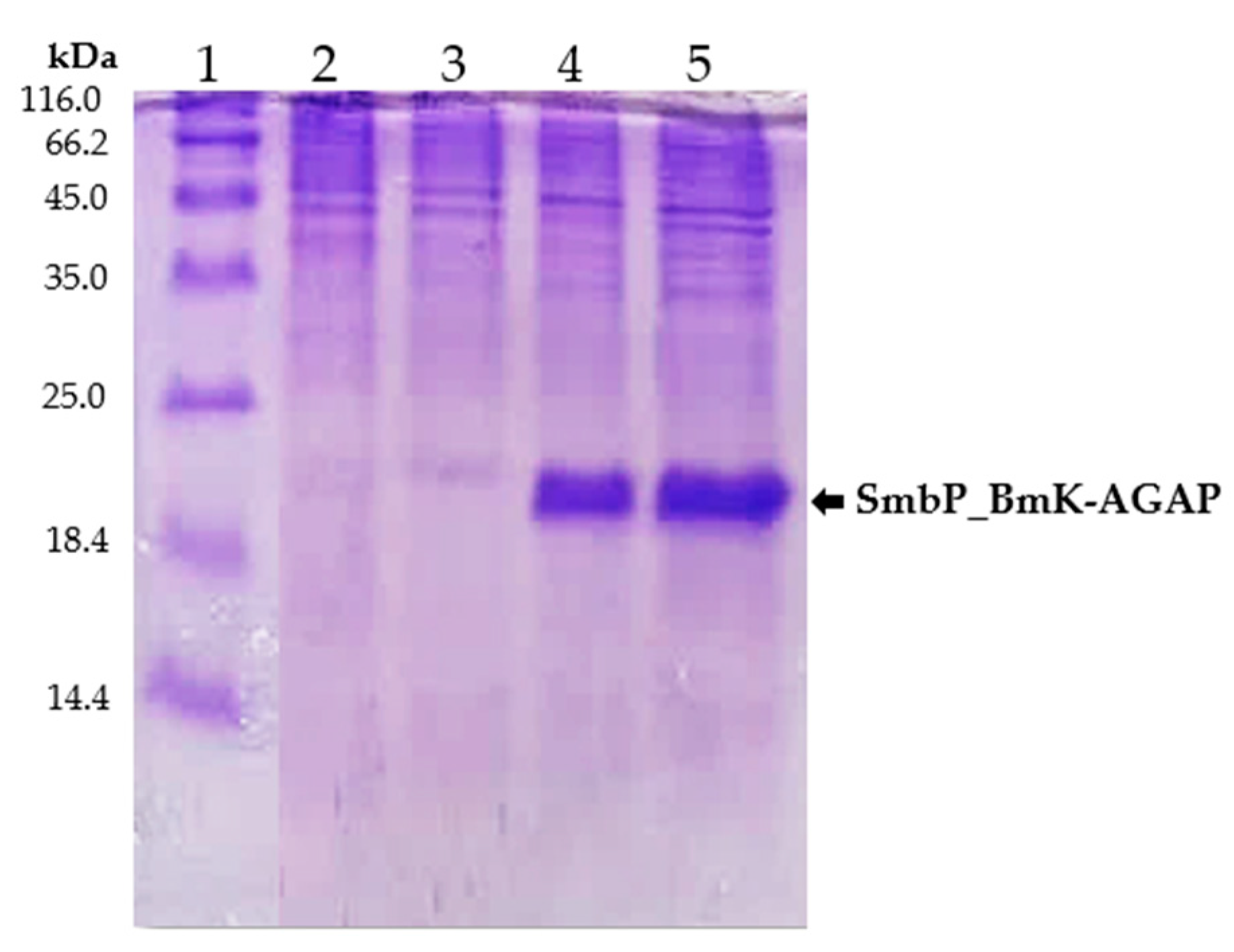
2.3. Expression of SmbP_BmK-AGAP Recombinant Protein
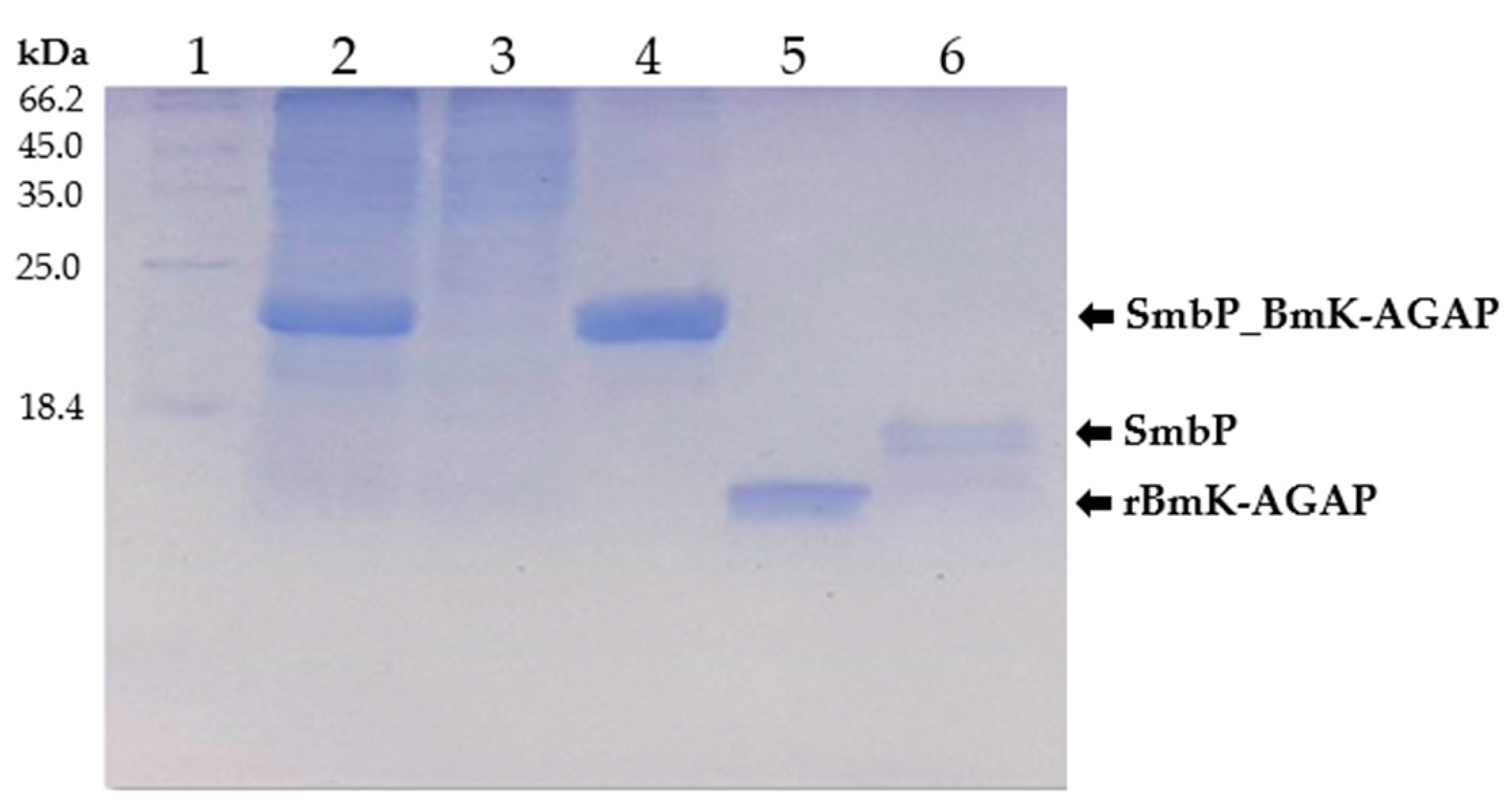
2.4. Purification of SmbP_BmK-AGAP Recombinant Protein
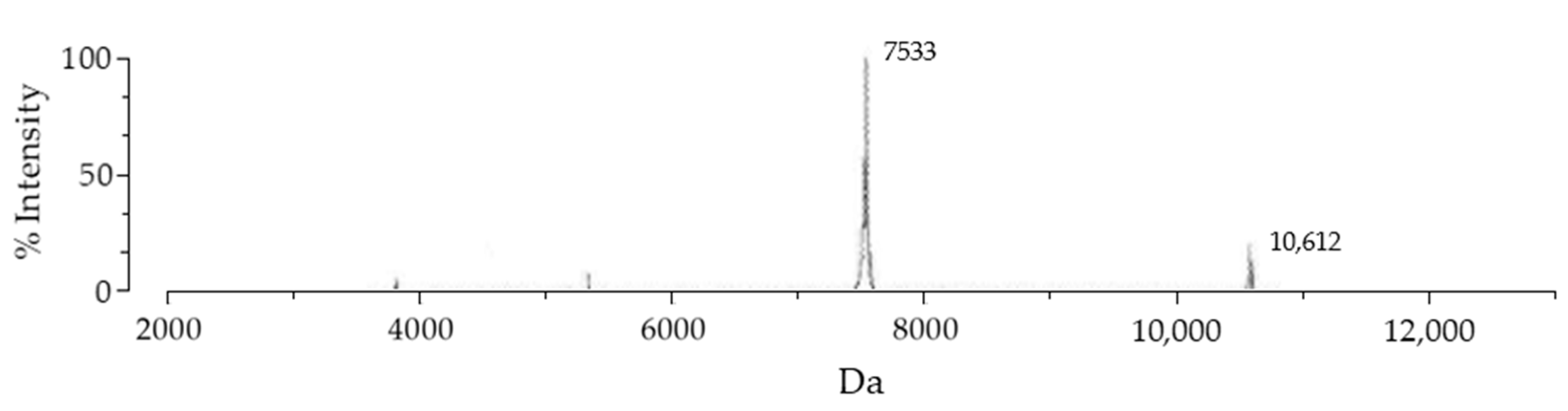
2.5. Enzymatic Removal of Fusion Protein
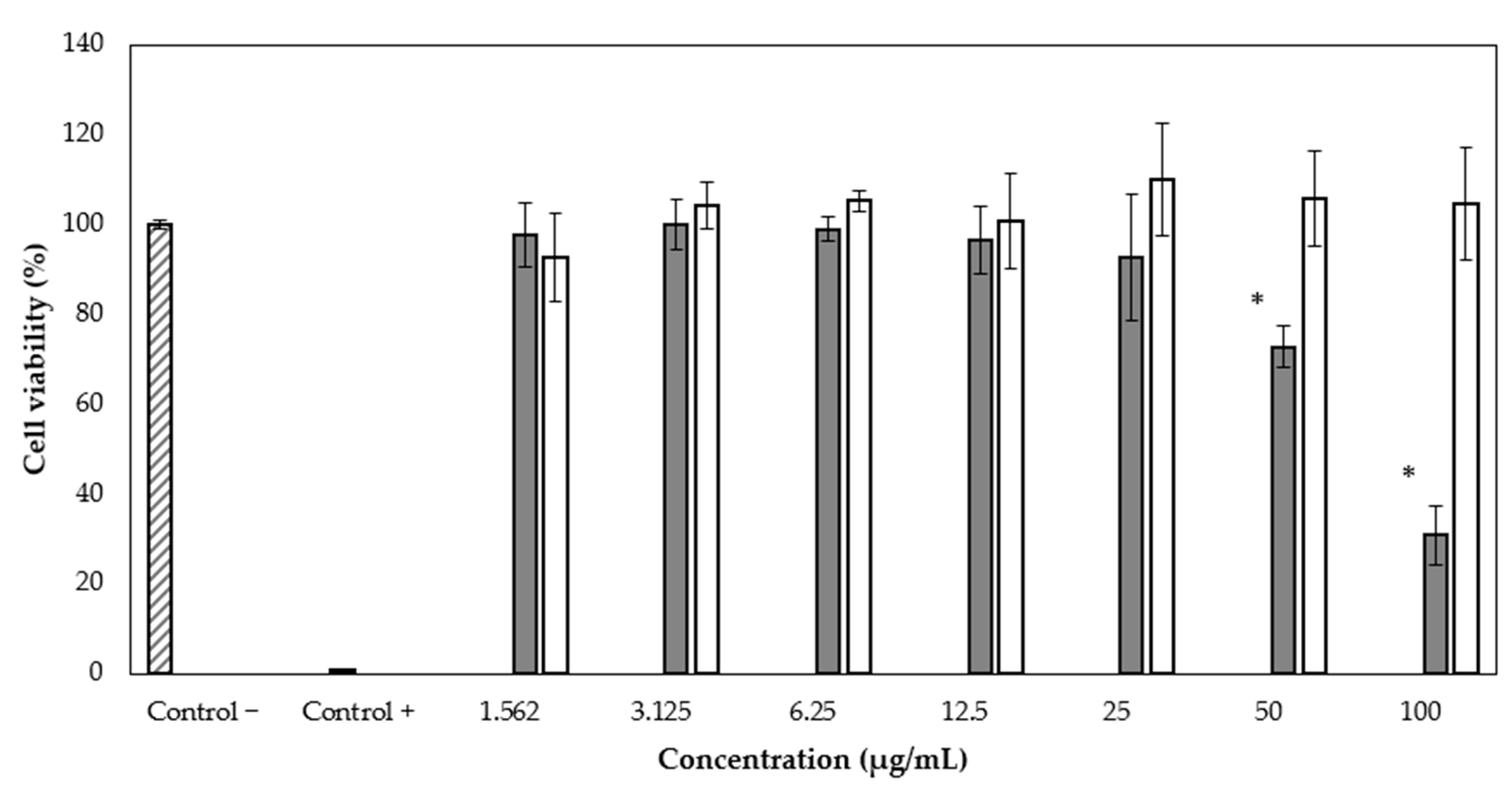
2.6. WST-1 Assay
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Marqus, S.; Pirogova, E.; Piva, T.J. Evaluation of the use of therapeutic peptides for cancer treatment. J. Biomed. Sci. 2017, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.S.; Cheung, R.C.F.; Xia, L.; Wong, J.H.; Ng, T.B.; Chan, W.Y. Snake venom toxins: Toxicity and medicinal applications. Appl. Microbiol. Biotechnol. 2016, 100, 6165–6181. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hu, P.; Wu, W.; Wang, Y. Peptides with therapeutic potential in the venom of the scorpion Buthus martensii Karsch. Peptides 2019, 115, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Richard, S.A.; Kampo, S.; Sackey, M.; Hechavarria, M.E.; Buunaaim, A.D.B. The pivotal potentials of scorpion Buthus martensii Karsch-analgesic-antitumor peptide in pain management and cancer. Evid.-Based Complement. Altern. Med. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Sarfo-Poku, C.; Eshun, O.; Lee, K.H. Medical application of scorpion venom to breast cancer: A mini-review. Toxicon 2016, 122, 109–112. [Google Scholar] [CrossRef]
- Kampo, S.; Ahmmed, B.; Zhou, T.; Owusu, L.; Anabah, T.W.; Doudou, N.R.; Kuugbee, E.D.; Cui, Y.; Lu, Z.; Yan, Q.; et al. Scorpion venom analgesic peptide, BmK AGAP inhibits stemness, and epithelial-mesenchymal transition by down-regulating PTX3 in breast cancer. Front. Oncol. 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; Bordon, K.D.C.F.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor snake venom proteins: Structure, function and potential applications. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 824–838. [Google Scholar] [CrossRef]
- Amorim, F.G.; Cordeiro, F.A.; Pinheiro-Júnior, E.L.; Boldrini-França, J.; Arantes, E.C. Microbial production of toxins from the scorpion venom: Properties and applications. Appl. Microbiol. Biotechnol. 2018, 102, 6319–6331. [Google Scholar] [CrossRef]
- Vargas-Cortez, T.; Morones-Ramirez, J.R.; Balderas-Renteria, I.; Zarate, X. Expression and purification of recombinant proteins in Escherichia coli tagged with a small metal-binding protein from Nitrosomonas europaea. Protein Expr. Purif. 2016, 118, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Barney, B.M.; LoBrutto, R.; Francisco, W.A. Characterization of a small metal binding protein from Nitrosomonas europaea. Biochemistry 2004, 43, 11206–11213. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lugo, J.J.; Santos, B.D.; Perez-Perez, D.A.; Montfort-Gardeazabal, J.M.; McEvoy, M.M.; Zarate, X. Expression and purification of recombinant proteins in Escherichia coli tagged with the metal-binding proteins SmbP and CusF3H+. In Protein Downstream Processing: Design, Development, and Application of High and Low-Resolution Methods, 2nd ed.; Labrou, N.E., Ed.; Humana Press: New York, NY, USA; Springer: Berlin/Heidelberg, Germany, 2021; Volume 1129, pp. 329–344. [Google Scholar] [CrossRef]
- Santos, B.D.; Morones-Ramirez, J.R.; Balderas-Renteria, I.; Casillas-Vega, N.G.; Galbraith, D.W.; Zarate, X. Optimizing periplasmic expression in Escherichia coli for the production of recombinant proteins tagged with the small metal-binding protein SmbP. Mol. Biotechnol. 2019, 61, 451–460. [Google Scholar] [CrossRef]
- Perez-Perez, D.A.; Pioquinto-Avila, E.; Arredondo-Espinoza, E.; Morones-Ramirez, J.R.; Balderas-Renteria, I.; Zarate, X. Engineered small metal-binding protein tag improves the production of recombinant human growth hormone in the periplasm of Escherichia coli. FEBS Open Bio 2020, 10, 546–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montfort-Gardeazabal, J.M.; Balderas-Renteria, I.; Casillas-Vega, N.G.; Zarate, X. Expression and purification of the antimicrobial peptide Bin1b in Escherichia coli tagged with the fusion proteins CusF3H+ and SmbP. Protein Expr. Purif. 2021, 178, 105784. [Google Scholar] [CrossRef] [PubMed]
- Montfort-Gardeazabal, J.M.; Claudio, P.C.M.-S.; Casillas-Vega, N.G.; Zarate, X. Expression and purification of the VpDef defensin in Escherichia coli using the small metal-binding proteins CusF3H+ and SmbP. Protein Pept. Lett. 2020, 28, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Perez-Perez, D.A.; Villanueva-Ramirez, T.D.J.; Hernandez-Pedraza, A.E.; Casillas-Vega, N.G.; Gonzalez-Barranco, P.; Zarate, X. The small metal-binding protein SmbP simplifies the recombinant expression and purification of the antimicrobial peptide LL-37. Antibiotics 2021, 10, 1271. [Google Scholar] [CrossRef]
- Hartig, S.M. Basic image analysis and manipulation in imageJ. Curr. Protoc. Mol. Biol. 2013, 14 (Suppl. 102), 1–12. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Hu, J.; Zhang, J.-H.; Wang, S.-L.; Wu, C.-F. Isolation, purification, and N-terminal partial sequence of an antitumor peptide from the venom of the Chinese scorpion Buthus martensii Karsch. Prep. Biochem. Biotechnol. 2002, 32, 317–327. [Google Scholar] [CrossRef]
- Cui, Y.; Guo, G.-L.; Ma, L.; Hu, N.; Song, Y.-B.; Liu, Y.-F.; Wu, C.-F.; Zhang, J.-H. Structure and function relationship of toxin from Chinese scorpion Buthus martensii Karsch (BmKAGAP): Gaining insight into related sites of analgesic activity. Peptides 2010, 31, 995–1000. [Google Scholar] [CrossRef]
- Lobstein, J.; Emrich, C.A.; Jeans, C.; Faulkner, M.; Riggs, P.; Berkmen, M. SHuffle, a novel Escherichia coli protein expression strain capable of correctly folding disulfide bonded proteins in its cytoplasm. Microb. Cell Fact. 2012, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.F.; Ma, R.L.; Wang, S.L.; Duan, Z.Y.; Zhang, J.H.; Wu, L.J.; Wu, C.F. Expression of an antitumor-analgesic peptide from the venom of Chinese scorpion Buthus martensii Karsch in Escherichia coli. Protein Expr. Purif. 2003, 27, 253–258. [Google Scholar] [CrossRef]
- Cao, P.; Yu, J.; Lu, W.; Cai, X.; Wang, Z.; Gu, Z.; Zhang, J.; Ye, T.; Wang, M. Expression and purification of an antitumor-analgesic peptide from the venom of Mesobuthus martensii Karsch by small ubiquitin-related modifier fusion in Escherichia coli. Biotechnol. Prog. 2010, 26, 1240–1244. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, Y.; Zhao, Y.; Yuan, J.; Mao, W. Inhibition effects of scorpion venom extracts (Buthus martensii Karsch) on the growth of human breast cancer MCF-7 cells. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 105–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Protein (mg) | Peptide (mg) | Yield (%) | Purity (%) |
|---|---|---|---|---|
| Lysate | 60 | 7.2 | 100 | 12 |
| 1° IMAC | 6.5 | 5.6 | 77.8 | 87 |
| Cleavage and 2° IMAC | 1.8 | 1.8 | 25 | 98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Mora, E.; Arredondo-Espinoza, E.; Casillas-Vega, N.G.; Cantu-Cardenas, M.E.; Balderas-Renteria, I.; Zarate, X. The Small Metal-Binding Protein SmbP Improves the Expression and Purification of the Recombinant Antitumor-Analgesic Peptide from the Chinese Scorpion Buthus martensii Karsch in Escherichia coli. Curr. Issues Mol. Biol. 2022, 44, 550-558. https://doi.org/10.3390/cimb44020038
Martinez-Mora E, Arredondo-Espinoza E, Casillas-Vega NG, Cantu-Cardenas ME, Balderas-Renteria I, Zarate X. The Small Metal-Binding Protein SmbP Improves the Expression and Purification of the Recombinant Antitumor-Analgesic Peptide from the Chinese Scorpion Buthus martensii Karsch in Escherichia coli. Current Issues in Molecular Biology. 2022; 44(2):550-558. https://doi.org/10.3390/cimb44020038
Chicago/Turabian StyleMartinez-Mora, Evelyn, Eder Arredondo-Espinoza, Nestor G. Casillas-Vega, Maria Elena Cantu-Cardenas, Isaias Balderas-Renteria, and Xristo Zarate. 2022. "The Small Metal-Binding Protein SmbP Improves the Expression and Purification of the Recombinant Antitumor-Analgesic Peptide from the Chinese Scorpion Buthus martensii Karsch in Escherichia coli" Current Issues in Molecular Biology 44, no. 2: 550-558. https://doi.org/10.3390/cimb44020038
APA StyleMartinez-Mora, E., Arredondo-Espinoza, E., Casillas-Vega, N. G., Cantu-Cardenas, M. E., Balderas-Renteria, I., & Zarate, X. (2022). The Small Metal-Binding Protein SmbP Improves the Expression and Purification of the Recombinant Antitumor-Analgesic Peptide from the Chinese Scorpion Buthus martensii Karsch in Escherichia coli. Current Issues in Molecular Biology, 44(2), 550-558. https://doi.org/10.3390/cimb44020038