
Can the Biological Activity of Abandoned Soils Be Changed by the Growth of Paulownia elongata × Paulownia fortunei?—Preliminary Study on a Young Tree Plantation





Abstract
1. Introduction
2. Materials and Methods
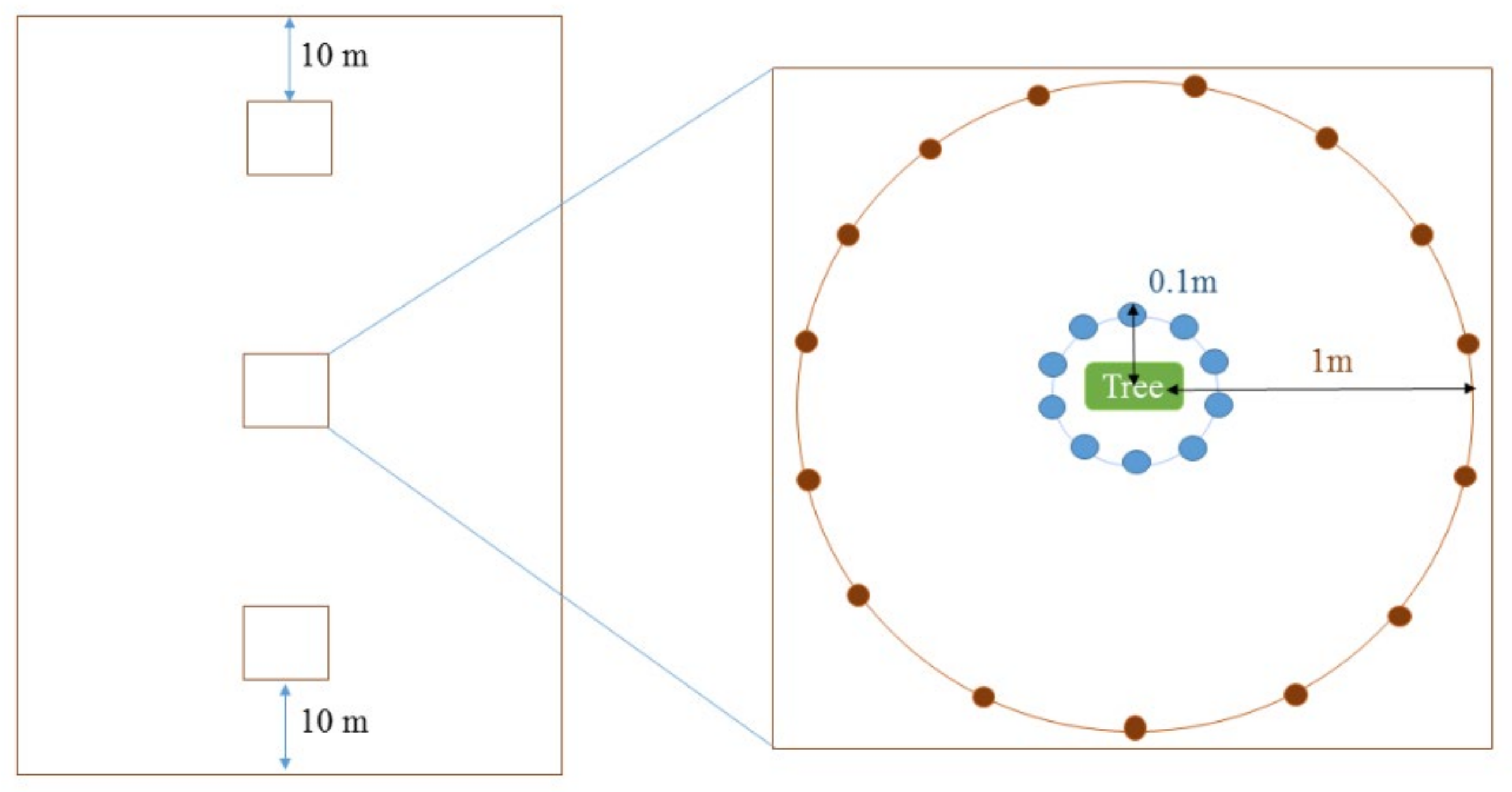
2.1. Study Sites and Soil Sampling
2.2. Determination of Soil Microbial Biomass and Enzyme Activity
2.3. Bacterial Community-Level Physiological Profiling (CLPP)
2.4. Statistical Analysis
3. Results
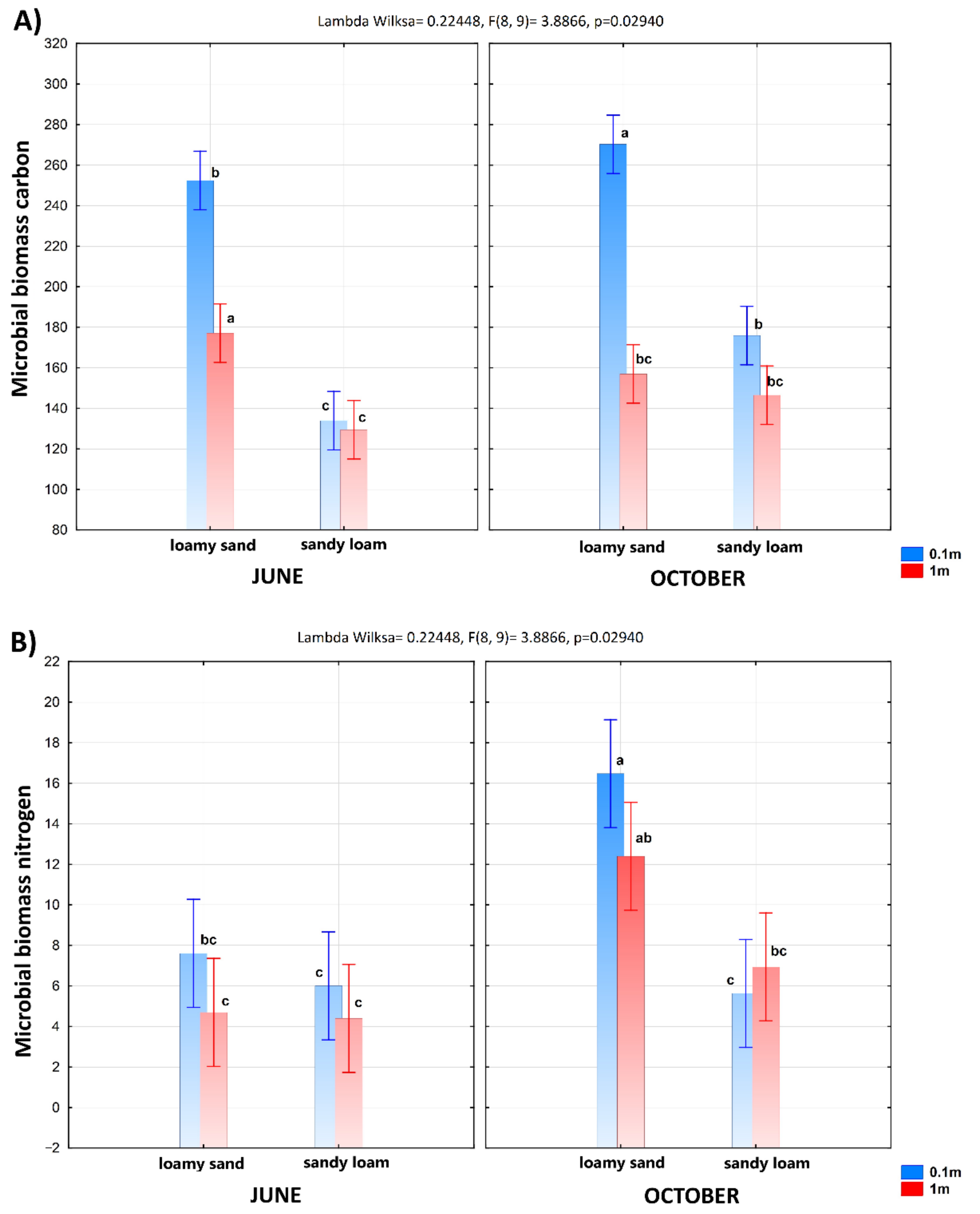
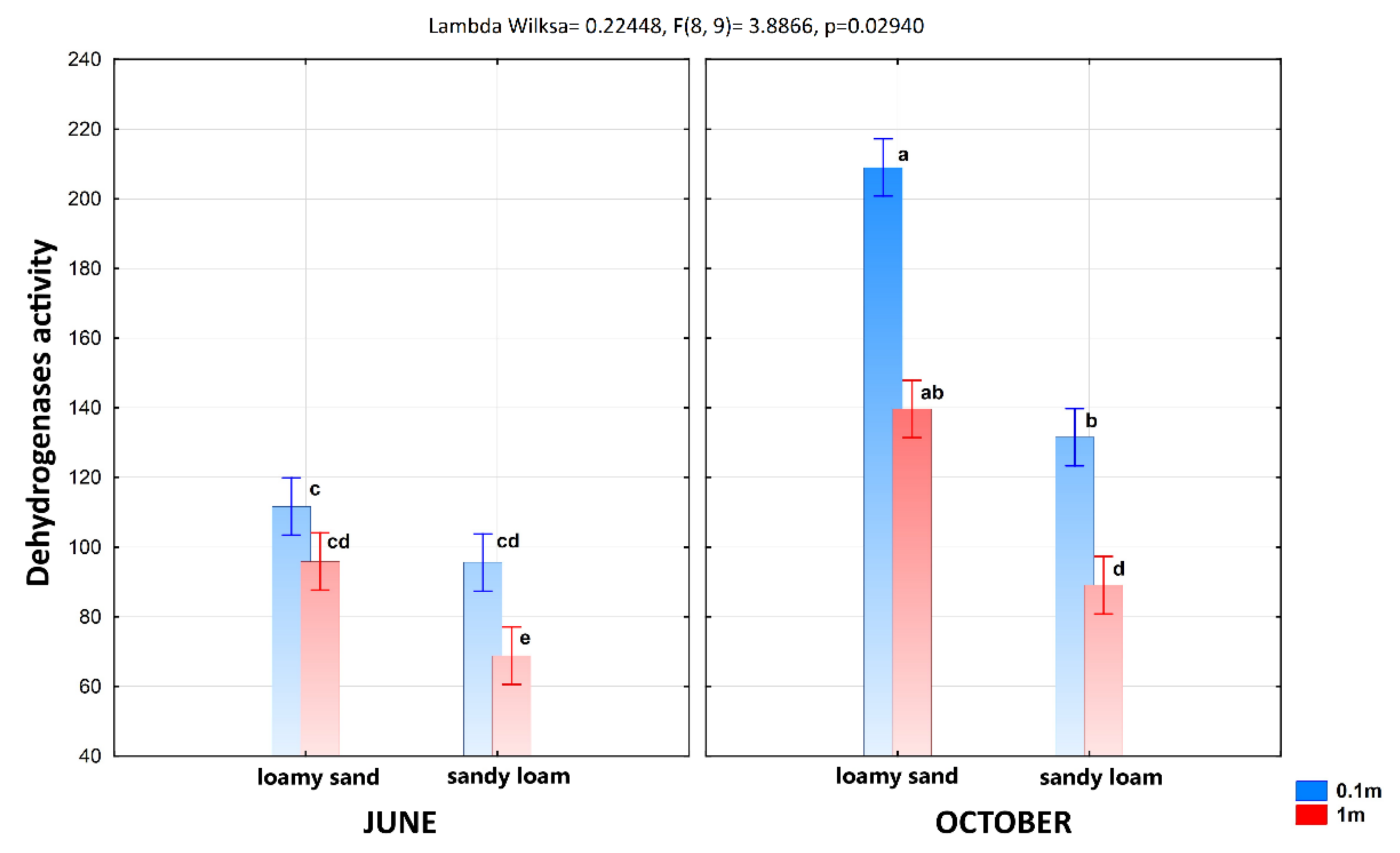
3.1. The Soil Microbial Biomass and Enzyme Activity
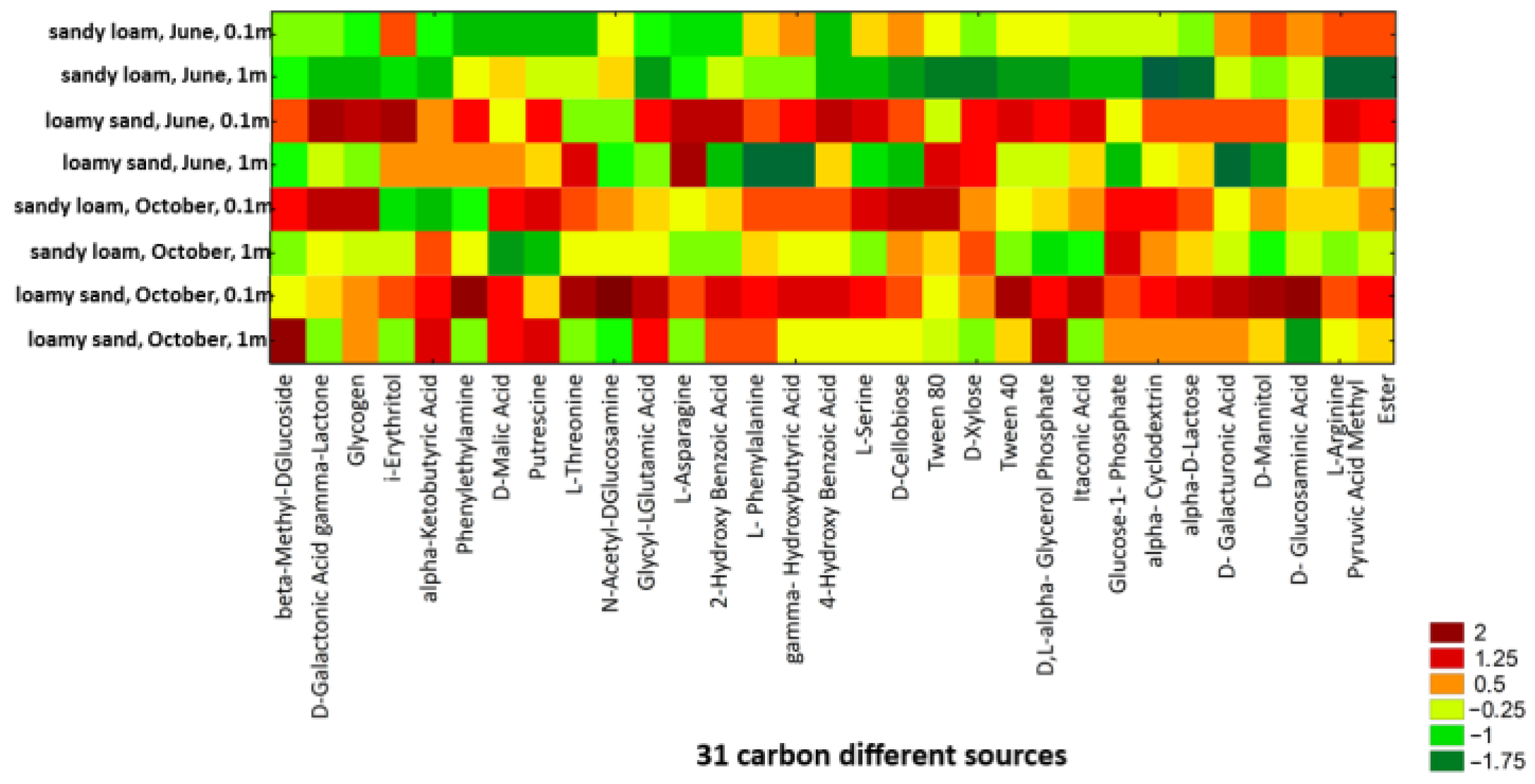
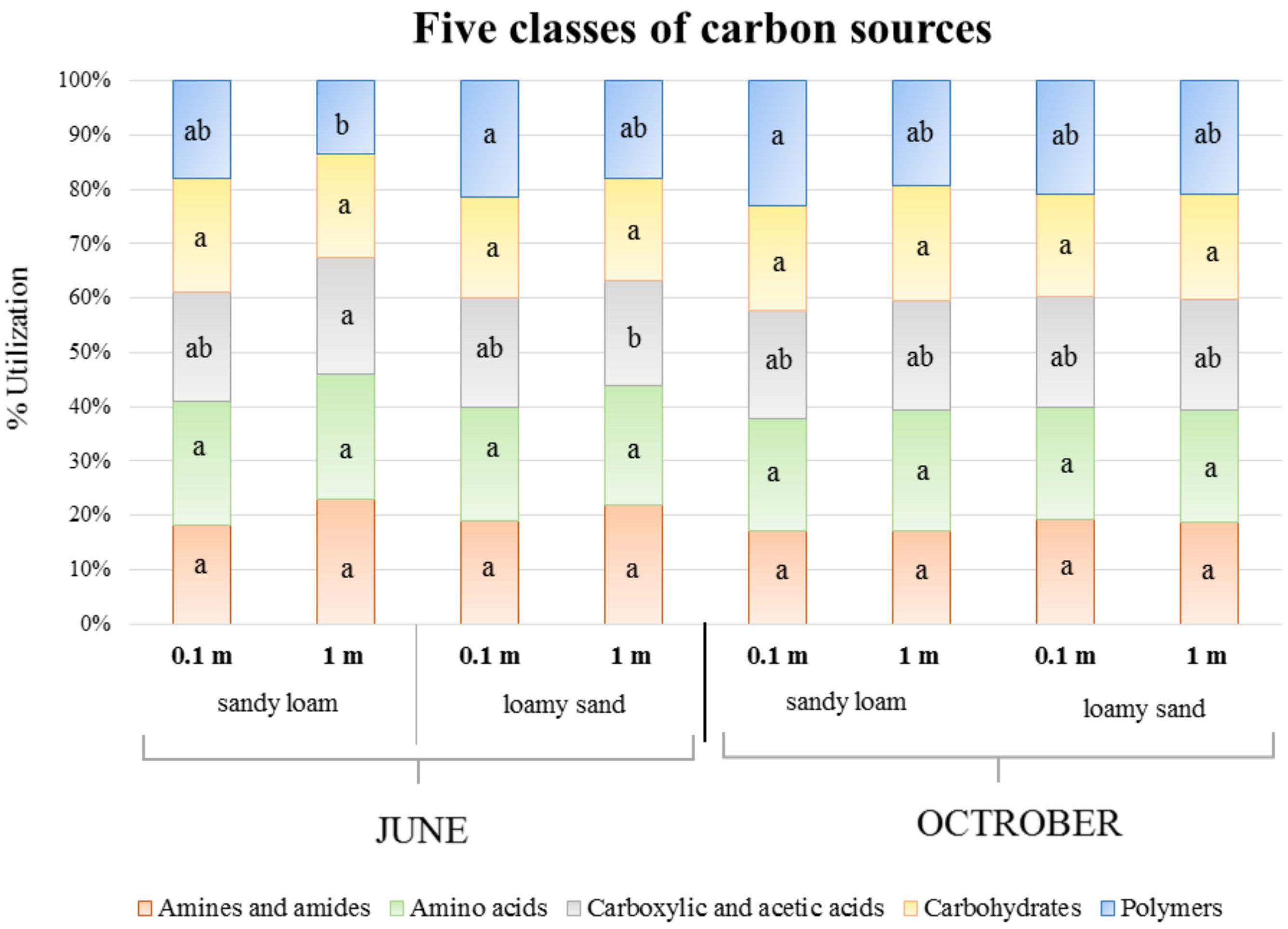
3.2. Bacterial Community-Level Physiological Profiling (CLPP)
3.3. Statistical Analysis
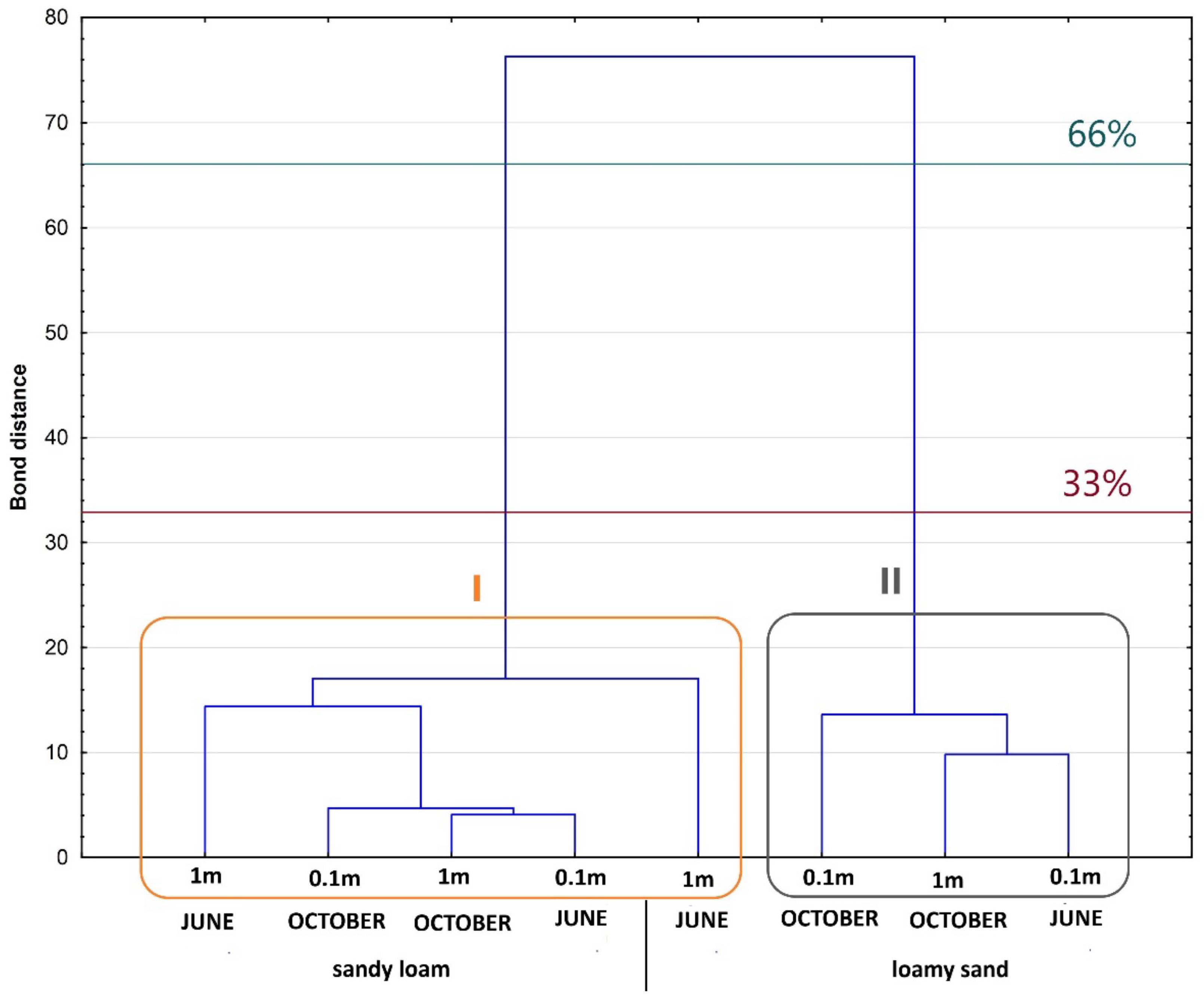
3.3.1. Hierarchical Cluster Analysis
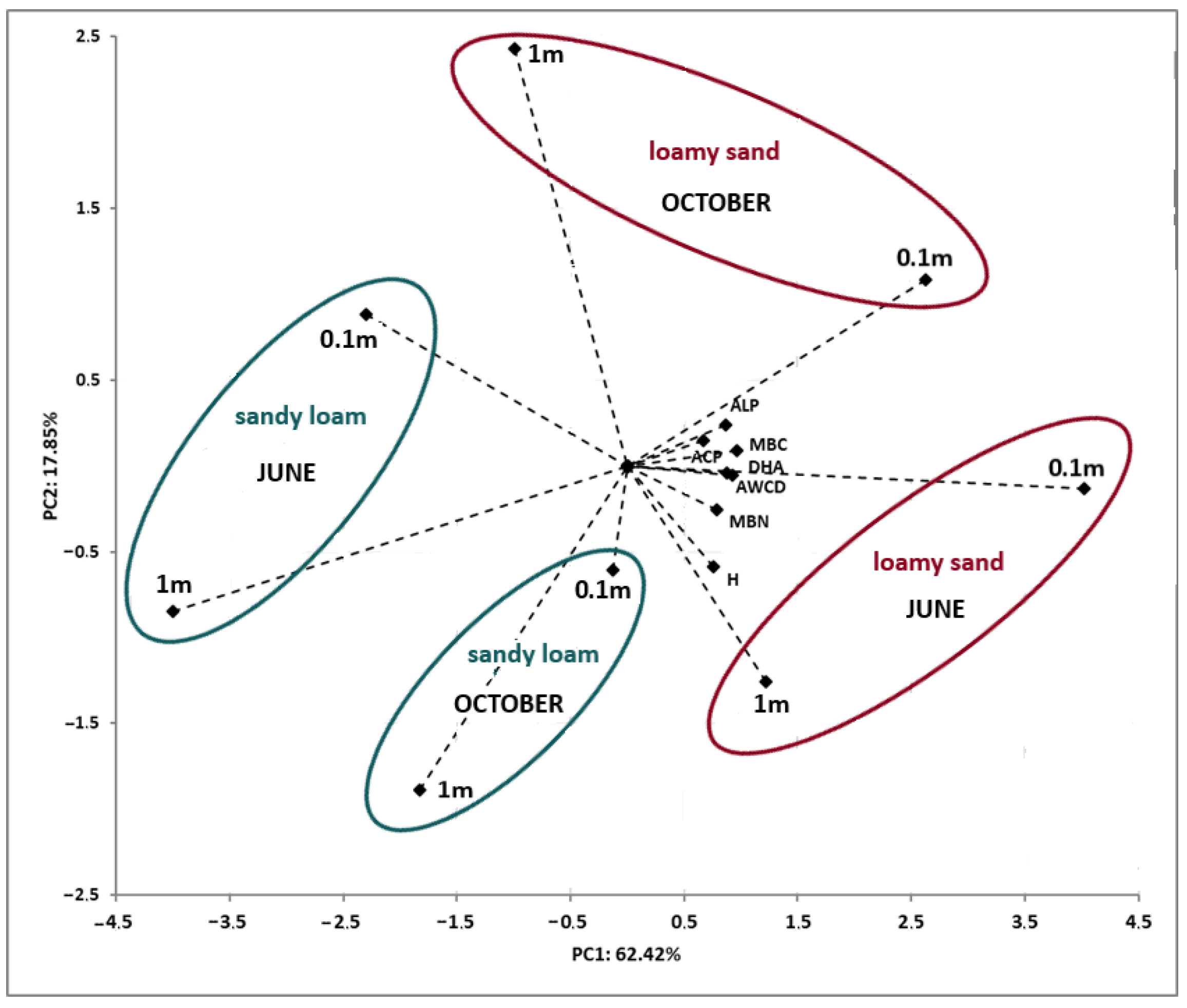
3.3.2. Principal Component Analysis
3.3.3. Pearson’s Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pazúr, R.; Lieskovský, J.; Bürgi, M.; Müller, D.; Lieskovský, T.; Zhang, Z.; Prishchepov, A.V. Abandonment and recultivation of agricultural lands in Slovakia—Patterns and determinants from the past to the future. Land 2020, 9, 316. [Google Scholar] [CrossRef]
- Helming, K.; Daedlow, K.; Hansjürgens, B.; Koellner, T. Assessment and governance of sustainable soil management. Sustainability 2018, 10, 4432. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Snyder, P.K. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Hatna, E.; Bakker, M.M. Abandonment and expansion of arable land in Europe. Ecosystems 2011, 14, 720–731. [Google Scholar] [CrossRef]
- Ustaoglu, E.; Collier, M.J. Farmland abandonment in Europe: An overview of drivers, consequences, and assessment of the sustainability implications. Environ. Rev. 2018, 26, 396–416. [Google Scholar] [CrossRef]
- FAO. FAOSTAT, Methods & standards [WWW Document]. URL. 2016. Available online: http://www.fao.org/ag/agn/nutrition/Indicatorsfiles/Agriculture.pdf (accessed on 20 November 2021).
- Pointereau, P.; Coulon, F.; Girard, P.; Lambotte, M.; Stuczynski, T.; Sánchez Ortega, V.; Del Rio, A. Analysis of Farmland Abandonment and the Extent and Location of Agricultural Areas that are Actually Abandoned or are in Risk to be Abandoned; Publications Office of the European Union: Luxembourg, 2008. [Google Scholar]
- Alcantara, C.; Kuemmerle, T.; Baumann, M.; Bragina, E.V.; Griffiths, P.; Hostert, P.; Radeloff, V.C. Mapping the extent of abandoned farmland in Central and Eastern Europe using MODIS time series satellite data. Environ. Res. Lett. 2013, 8, 035035. [Google Scholar] [CrossRef]
- Van der Zanden, E.H.; Verburg, P.H.; Schulp, C.J.; Verkerk, P.J. Trade-offs of European agricultural abandonment. Land Use Policy 2017, 62, 290–301. [Google Scholar] [CrossRef]
- Castillo, C.P.; Kavalov, B.; Diogo, V.; Jacobs-Crisioni, C.; e Silva, F.B.; Lavalle, C. Agricultural Land Abandonment in the EU within 2015–2023. 2018. Available online: https://ec.europa.eu/jrc/sites/default/files/jrc113718.pdf (accessed on 20 November 2021).
- Bell, S.; Barriocanal, C.; Terrer, C.; Rosell-Melé, A. Management Opportunities for Soil Carbon Sequestration Following Agricultural Land Abandonment. Environ. Sci. Policy 2020, 108, 104–111. [Google Scholar] [CrossRef]
- Schou, J.S.; Olsen, J.V. Multifunctional land consolidation evaluation from an agricultural economics perspective. In International Farm Management Congress; Grand Chancellor Hotel: Launceston, Australia, 2019. [Google Scholar]
- Renting, H.; Rossing, W.A.H.; Groot, J.C.J.; Van der Ploeg, J.D.; Laurent, C.; Perraudd, D.; Stobbelaar, D.J.; Van Ittersum, M.K. Exploring multifunctional agriculture. A review of conceptual approches and prospects for an integrative transitional framework. J. Environ. Manag. 2009, 90, 112–123. [Google Scholar] [CrossRef]
- Di Bene, C.; Pellegrino, E.; Tozzini, C.; Bonari, E. Changes in soil quality following poplar short-rotation forestry under different cutting cycles. Ital. J. Agron. 2011, 6, e6. [Google Scholar] [CrossRef]
- Hirschmugl, M.; Sobe, C.; Khawaja, C.; Janssen, R.; Traverso, L. Pan-European Mapping of Underutilized Land for Bioenergy Production. Land 2021, 10, 102. [Google Scholar] [CrossRef]
- Wise, M.; Calvin, K.; Thomson, A.; Clarke, L.; Bond-Lamberty, B.; Sands, R.; Smith, S.J.; Janetos, A.; Edmonds, J. Implications of limiting CO2 concentrations for land use and energy. Science 2009, 324, 1183–1186. [Google Scholar] [CrossRef]
- Popp, A.; Lotze-Campen, H.; Leimbach, M.; Knof, B.; Beringer, T.; Bauer, N.; Bodirsky, B. On sustainability of bionenergy prduction: Integrating co-emissions from agricultural intensification. Biomass Bionerg. 2010, 35, 4770–4780. [Google Scholar] [CrossRef]
- Kumar, A.; Jyske, T.; Möttönen, V. Properties of Injection Molded Biocomposites Reinforced with Wood Particles of Short-Rotation Aspen and Willow. Polymers 2020, 12, 257. [Google Scholar] [CrossRef] [PubMed]
- Schulze, J.; Gawel, E.; Nolzen, H.; Weise, H.; Frank, K. The expansion of short rotation forestry: Characterization of determinants with an agent-based land use model. GCB Bioenergy 2017, 9, 1042–1056. [Google Scholar] [CrossRef]
- Rahman, M.A.; Rahman, F.; Rahmatullah, M. In vitro regeneration of Paulownia tomentosa Steud. plants through the induction of adventitious shoots in explants derived from selected mature trees, by studying the effect of different plant growth regulators. Am.-Eurasian J. Sustain. Agric. 2013, 7, 259–268. [Google Scholar]
- Popović, J.; Suzana, M.; Milorad, V.; Dragica, V. Impact of soil to dimensions of mechanical fibres of a juvenile wood of Paulownia elongata S.Y.HU. In Proceedings of the International Conference Reforestation Challenges, Belgrade, Serbia, 3–6 June 2015; Ivetić, V., Stanković, D., Eds.; Reforesta: Belgrade, Serbia, 2015; pp. 175–184. [Google Scholar]
- El-Showk, S.; El-Showk, N. The Paulownia Tree: An Alternative for Sustainable Forestry. The Farm. 2003. Available online: http://www.cropdevelopment.org/docs/PaulowniaBooklet_print.pdf (accessed on 20 November 2021).
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The Role of Soil Microorganisms in Plant Mineral Nutrition—Current Knowledge and Future Directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Attiwill, P.M.; Adams, M.A. Nutrient cycling in forests. New Phytol. 1993, 124, 561–582. [Google Scholar] [CrossRef]
- Yang, C.H.; Crowley, D.E. Rhizosphere microbial community structure in relation to root location and plant iron nutritional status. Appl. Environ. Microbiol. 2000, 66, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Menyailo, O.V.; Hungate, B.A.; Zech, W. Tree species mediated soil chemical changes in a Siberian artificial afforestation experiment. Plant Soil 2002, 242, 171–182. [Google Scholar] [CrossRef]
- Margesin, R.; Minerbi, S.; Schinner, F. Long-term monitoring of soil microbiological activities in two forest sites in South Tyrol in the Italian Alps. Microbes Environ. 2014, 29, 277–285. [Google Scholar] [CrossRef]
- Frąc, M.; Oszust, K.; Lipiec, J. Community level physiological profiles (CLPP), characterization and microbial activity of soil amended with dairy sewage sludge. Sensors 2012, 12, 3253–3268. [Google Scholar] [CrossRef]
- Houben, D.; Daoulas, G.; Faucon, M.P.; Dulaurent, A.M. Potential use of mealworm frass as a fertilizer: Impact on crop growth and soil properties. Sci. Rep. 2020, 10, 4659. [Google Scholar] [CrossRef]
- Zuccarini, P.; Asensio, D.; Ogaya, R.; Sardans, J.; Peñuelas, J. Effects of seasonal and decadal warming on soil enzymatic activity in a P-deficient Mediterranean shrubland. Glob. Chang. Biol. 2020, 26, 3698–3714. [Google Scholar] [CrossRef] [PubMed]
- Rajper, A.M.; Udawatta, R.P.; Kremer, R.J.; Lin, C.H.; Jose, S. Effects of probiotics on soil microbial activity, biomass and enzymatic activity under cover crops in field and greenhouse studies. Agrofor. Syst. 2016, 90, 811–827. [Google Scholar] [CrossRef]
- Van Leeuwen, J.P.; Djukic, I.; Bloem, J.; Lehtinen, T.; Hemerik, L.; de Ruiter, P.C.; Lair, G.J. Effects of land use on soil microbial biomass, activity and community structure at different soil depths in the Danube floodplain. Eur. J. Soil Biol. 2017, 79, 14–20. [Google Scholar] [CrossRef]
- Insam, H. A new set of substrates proposed for community characterization in environmental samples. In Microbial Communities; Insam, H., Rangger, A., Eds.; Springer: Berlin, Germany, 1997; pp. 259–260. [Google Scholar] [CrossRef]
- Garland, J.L.; Millis, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Lehman, R.M.; Colwell, F.S.; Ringelbergb, D.B.; Whiteb, D.C. Combined microbial community-level analyses for quality assurance of terrestrial subsurface cores. J. Microbiol. Methods 1995, 22, 263–281. [Google Scholar] [CrossRef]
- Ros, M.; Goberna, M.; Pascual, J.A.; Klammer, S.; Insam, H.J. 16S rDNA analysis reveals low microbial diversity in community level physiological profile assays. Microbiol. Methods 2008, 72, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Varma, A. Role of enzymes in maintaining soil health. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer-Verlag: Berlin, Germany, 2010; pp. 25–42. [Google Scholar] [CrossRef]
- Calfapietra, C.; Gielen, B.; Karnosky, D.; Ceulemans, R.; Scarascia-Mugnozza, G. Response and potential of agroforestry crops under global change. Environ. Pollut. 2010, 158, 1095–1104. [Google Scholar] [CrossRef]
- Stauffer, M.; Leyval, C.; Brun, J.J.; Leportier, P.; Berthelin, J. Effect of willow short rotation coppice on soil properties after three years of growth as compared to forest, grassland and arable land uses. Plant Soil 2014, 377, 423–438. [Google Scholar] [CrossRef][Green Version]
- Porta, C.J.; López-Acevedo, R.M.; Roquero, L.C. Edafología Para la Agricultura y el Medio Ambiente; Mundi-Prensa: Madrid, Spain, 2003. [Google Scholar]
- Lucas-Borja, M.E.; Wic-Baena, C.; Moreno, J.L.; Dadi, T.; García, C.; Andrés-Abellán, M. Microbial activity in soils under fast-growing Paulownia (Paulownia elongata x fortunei) plantations in Mediterranean areas. Appl. Soil Ecol. 2011, 51, 42–51. [Google Scholar] [CrossRef]
- Woźniak, M.; Grządziel, J.; Gałązka, A.; Frąc, M. Metagenomic analysis of bacterial and fungal community composition associated with Paulownia elongata × Paulownia fortunei. BioResources 2019, 14, 8511–8529. [Google Scholar] [CrossRef]
- Moreno, J.L.; Bastida, F.; Ondoño, S.; García, C.; Andrés-Abellán, M.; López-Serrano, F.R. Agro-forestry management of Paulownia plantations and their impact on soil biological quality: The effects of fertilization and irrigation treatments. Appl. Soil Ecol. 2017, 117, 46–56. [Google Scholar] [CrossRef]
- Graham, W.R.; Leff, J.W.; Weintraub, S.R.; Townsend, A.R.; Cleveland, C.C.; Philippot, L.; Nemergut, D.R. Do we need to understand microbial communities to predict ecosystem function? A comparison of statistical models of nitrogen cycling processes. Soil Biol. Biochem. 2014, 68, 279–282. [Google Scholar] [CrossRef]
- PN-R-04023; Chemical and Agricultural Analysis. Determination of the Content Available Phosphorus in Mineral Soils. Polish Standards Committee: Warszawa, Poland, 1996.
- PN-R-04022; Chemical and Agricultural Analysis. Determination of the Content Available Potassium in Mineral Soils. Polish Standards Committee: Warszawa, Poland, 1996.
- PN-R-04020; Chemical and Agricultural Analysis. Determination of the Content Available Magnesium. Polish Standards Committee: Warszawa, Poland, 1994.
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction—an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Jenkinson, D.S. Determination of microbial biomass carbon and nitrogen in soil. In Advances in Nitrogen Cycling in Agricultural Ecosystems; Wilson, J.R., Ed.; CAB International: Wallingford, UK, 1988; pp. 368–386. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenol phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Casida, L.E.; Klein, D.A.; Santoro, T. Soil dehydrogenase activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Pohland, B.; Owen, B. Biolog EcoPlates standard methods. TAS Tech. Bull. Biol. 2009, 1, 1–3. [Google Scholar]
- Wolbert-Haverkamp, M.; Musshoff, O. Are short rotation coppices an economically interesting form of land use? A real options analysis. Land Use Policy 2014, 38, 163–174. [Google Scholar] [CrossRef]
- Abolina, E.; Luzadis, V.A. Abandoned agricultural land and its potential for short rotation woody crops in Latvia. Land Use Policy 2015, 49, 435–445. [Google Scholar] [CrossRef]
- Rowe, R.L.; Hanley, M.E.; Goulson, D.; Clarke, D.J.; Doncaster, C.P.; Taylor, G. Potential benefits of commercial willow Short Rotation Coppice (SRC) for farm-scale plant and invertebrate communities in the agri-environment. Biomass Bioenerg. 2011, 35, 325–336. [Google Scholar] [CrossRef]
- Holland, R.A.; Eigenbrod, F.; Muggeridge, A.; Brown, G.; Clarke, D.; Taylor, G. A synthesis of the ecosystem services impact of second generation bioenergy crop production. Renew. Sust. Energ. Rev. 2015, 46, 30–40. [Google Scholar] [CrossRef]
- Schmidt-Walter, P.; Lamersdorf, N.P. Biomass production with willow and poplar short rotation coppices on sensitive areas-the impact on nitrate leaching and groundwater recharge in a drinking water catchment near Hanover, Germany. Bioenergy Res. 2012, 5, 546–562. [Google Scholar] [CrossRef]
- Doran, J.W.; Parkin, T.B. Defining and assessing soil quality. In Defining Soil Quality for a Sustainable Environment; Doran, J.W., Coleman, D.C., Bezdicek, D.F., Stewart, B.A., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 1–20. [Google Scholar]
- Acosta-Martínez, V.; Zobeck, T.M.; Allen, V. Soil microbial, chemical and physical properties in continuous cotton and integrated crop–livestock systems. Soil Sci. Soc. Am. J. 2004, 68, 1875–1884. [Google Scholar] [CrossRef]
- Das, P.K.; Nath, S.; Banerjee, S.K. Distribution of microorganisms in soils under different forest cover at different altitudes. Indian Agric. 1991, 35, 217–223. [Google Scholar]
- Liu, G.; Jin, M.; Cai, C.; Ma, C.; Chen, Z.; Gao, L. Soil microbial community structure and physicochemical properties in amomum tsaoko-based agroforestry systems in the Gaoligong Mountains, Southwest China. Sustainability 2019, 11, 546. [Google Scholar] [CrossRef]
- Pang, D.; Wang, G.; Liu, Y.; Cao, J.; Wan, L.; Wu, X.; Zhou, J. The Impacts of Vegetation Types and Soil Properties on Soil Microbial Activity and Metabolic Diversity in Subtropical Forests. Forests 2019, 10, 497. [Google Scholar] [CrossRef]
- Neina, D. The role of soil pH in plant nutrition and soil remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef]
- Hinsinger, P.; Plassard, C.; Jaillard, B. Rhizosphere: A new frontier for soil biogeochemistry. J. Geochem. Explor. 2006, 88, 210–213. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Callesen, I.; Armolaitis, K.; Nihlgård, B. The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For. Ecol. Manag. 2004, 195, 373–384. [Google Scholar] [CrossRef]
- Khan, B.; Ablimit, A.; Mahmood, R.; Qasim, M. Robinia pseudoacacia leaves improve soil physical and chemical properties. J. Arid Land 2010, 2, 266–271. [Google Scholar] [CrossRef]
- Xu, Y.; Du, A.; Wang, Z.; Zhu, W.; Li, C.; Wu, L. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity. For. Ecol. Manag. 2020, 456, 117683. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Baath, E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef]
- Staelens, J.; Rütting, T.; Huygens, D.; Schrijver, A.D.; Müller, C.; Verheyen, K.; Boeckx, P. In situ gross nitrogen transformations differ between temperate deciduous and coniferous forest soils. Biogeochemistry 2012, 108, 259–277. [Google Scholar] [CrossRef]
- Hackl, E.; Pfeffer, M.; Donat, C.; Bachmann, G.; Zechmeister-Boltenstern, S. Composition of the microbial communities in the mineral soil under different types of natural forest. Soil Biol. Biochem. 2005, 37, 661–671. [Google Scholar] [CrossRef]
- Banerjee, S.; Baah-Acheamfour, M.; Carlyle, C.N.; Bissett, A.; Richardson, A.E.; Siddique, T.; Bork, E.W.; Chang, S.X. Determinants of bacterial communities in Canadian agroforestry systems: Co-occurrence patterns of soil bacterial communities. Environ. Microbiol. 2016, 18, 1805–1816. [Google Scholar] [CrossRef]
- Mungai, N.W.; Motavalli, P.P.; Kremer, R.J.; Nelson, K.A. Spatial variation of soil enzyme activities and microbial functional diversity in temperate alley cropping systems. Biol. Fertil. Soils 2005, 42, 129–136. [Google Scholar] [CrossRef]
- Prasad, R.; Mertia, R.S. Dehydrogenase activity and VAM fungi in tree-rhizosphere of agroforestry systems in Indian arid zone. Agrofor. Syst. 2005, 63, 219–223. [Google Scholar] [CrossRef]
- Yadav, R.S.; Yadav, B.L.; Chhipa, B.R.; Dhyani, S.K.; Munna, R. Soil biological properties under different tree based traditional agroforestry systems in a semi-arid region of Rajasthan, India. Agrofor. Syst. 2011, 81, 195–202. [Google Scholar] [CrossRef]
- Wan, F.; Chen, P. Soil enzyme activities under agroforestry systems in Northern Jiangsu Province. For. Stud. China 2004, 6, 21–26. [Google Scholar] [CrossRef]
- Yadav, N.K.; Vaidya, B.N.; Henderson, K.; Lee, J.F.; Stewart, W.M.; Dhekney, S.A.; Joshee, N. A review of Paulownia biotechnology: A short rotation, fast growing multipurpose bioenergy tree. Am. J. Plant Sci. 2013, 4, 2070–2082. [Google Scholar] [CrossRef]
- Cederlund, H.; Wessen, E.; Enwall, K.; Jones, C.M.; Juhanson, J.; Pell, M.; Philippot, L.; Hallin, S. Soil carbon quality and nitrogen fertilization structure bacterial communities with predictable responses of major bacterial phyla. Appl. Soil Ecol. 2014, 84, 62–68. [Google Scholar] [CrossRef]
- Li, F.; Chen, L.; Zhang, J.; Yin, J.; Huang, S. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed]
- Uthappa, A.R.; Bana, O.P.S.; Kumar, M.; Kanwal, M. Soil physico-bio-chemical properties as influenced by varying tree densities in poplar (Populus deltoides Bartr. ex Marsh.) based agroforestry system. Indian J. Agrofor. 2015, 17, 81–90. [Google Scholar]
- Wang, Q.; Shogren, J.F. Characteristics of the crop-Paulownia system in China. Agr. Ecosyst. Environ. 1992, 39, 145–152. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Bowman, W.D.; Kaufmann, R.; Schmidt, S.K. A temporal approach to linking aboveground and belowground ecology. Trends Ecol. Evol. 2005, 20, 634–641. [Google Scholar] [CrossRef]
- Grogan, P.; Illeris, L.; Michelsen, A.; Jonasson, S. Respiration of recently-fixed plant carbon dominates mid-winter ecosystem CO2 production in sub-arctic heath tundra. Clim. Chang. 2001, 50, 129–142. [Google Scholar] [CrossRef]
- Lipson, D.A.; Schmidt, S.K. Seasonal changes in an alpine soil bacterial community in the Colorado Rocky Mountains. Appl. Environ. Microbiol. 2004, 70, 2867–2879. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.K.; Costello, E.K.; Nemergut, D.R.; Cleveland, C.C.; Reed, S.C.; Weintraub, M.N.; Martin, A.M. Biogeochemical consequences of rapid microbial turnover and seasonal succession in soil. Ecology 2007, 88, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.P.; Mikan, C. Changing microbial substrate use in Arctic tundra soils through a freeze-thaw cycle. Soil Biol. Biochem. 2005, 37, 1411–1418. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Costello, E.K.; Meyer, A.F.; Pescador, M.Y.; Weintraub, M.N.; Schmidt, S.K. Structure and function of alpine and arctic soil microbial communities. Res. Microbiol. 2005, 156, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Koranda, M.; Kaiser, C.; Fuchslueger, L.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S.; Richter, A. Seasonal variation in functional properties of microbial communities in beech forest soil. Soil Biol. Biochem. 2013, 60, 95–104. [Google Scholar] [CrossRef]
- Bini, D.; Figueiredo, A.F.; Silva, M.C.P.D.; Vasconcellos, R.L.D.F.; Cardoso, E.J.B.N. Microbial biomass and activity in litter during the initial development of pure and mixed plantations of Eucalyptus grandis and Acacia mangium. Rev. Bras. Cienc. Solo 2013, 37, 76–85. [Google Scholar] [CrossRef]
- Shigyo, N.; Umeki, K.; Hirao, T. Seasonal Dynamics of Soil Fungal and Bacterial Communities in Cool-Temperate Montane Forests. Front. Microbiol. 2019, 10, 1944. [Google Scholar] [CrossRef] [PubMed]
- Gou, X.; Tan, B.; Wu, F.; Yang, W.; Xu, Z.; Li, Z.; Zhang, X. Seasonal dynamics of soil microbial biomass C and N along an elevational gradient on the eastern Tibetan Plateau, China. PLoS ONE 2015, 10, e0132443. [Google Scholar] [CrossRef]
- Ge, G.; Li, Z.; Fan, F.; Chu, G.; Hou, Z.; Liang, Y. Soil biological activity and their seasonal variations in response to long-term application of organic and inorganic fertilizers. Plant Soil 2010, 326, 31. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.K.; Sharma, R.; Chander, J. Short rotation forestry: It’s application for biomass, energy, soil health and carbon sequestration. In Agroforestry Strategies for Climate Change: Mitigation and Adaptation; Parthiban, K.T., Sudhagar, R.J., Fernandaz, C.C., Suresh, K.K., Eds.; Jaya Publishing House: Delhi, India, 2017; pp. 139–168. [Google Scholar]
- Swamy, S.L.; Mishra, A.; Puri, S. Comparison of growth, biomass and nutrient distribution in five promising clones of Populus deltoides under an agrisilviculture system. Bioresour. Technol. 2006, 97, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sharma, K.N. Tree growth and nutrient status of soil in a poplar (Populus deltoides Bartr.)-based agroforestry system in Punjab, India. Agrofor. Syst. 2007, 70, 125–134. [Google Scholar] [CrossRef]
- Gupta, N.; Kukal, S.S.; Bawa, S.S.; Dhaliwal, G.S. Soil organic carbon and aggregation under poplar based agroforestry system in relation to tree age and soil type. Agrofor. Syst. 2009, 76, 27–35. [Google Scholar] [CrossRef]
- Turley, N.E.; Bell-Dereske, L.; Evans, S.E.; Brudvig, L.A. Agricultural land-use history and restoration impact soil microbial biodiversity. J. Appl. Ecol. 2020, 57, 852–863. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Chen, Y.; Wang, M.; Liu, G.; Xue, S.; Yang, X. Disentangling the direct and indirect effects of cropland abandonment on soil microbial activity in grassland soil at different depths. Catena 2020, 194, 104774. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Soil Texture | June | October | ||
|---|---|---|---|---|---|
| Podkampinos | sandy loam | 0.1 m | 1 m | 0.1 m | 1 m |
| Otrębusy | loamy sand | 0.1 m | 1 m | 0.1 m | 1 m |
| Soil Properties | Site | |
|---|---|---|
| Podkampinos | Otrębusy | |
| Sand % | 62.35 | 80.60 |
| Silt % | 34.66 | 17.73 |
| Clay % | 2.99 | 1.66 |
| Texture 1* | sandy loam | loamy sand |
| N-NH4 (mg kg−1) 2* | 0.75 | 0.28 |
| N-NO2 (mg kg−1) 2* | 0.03 | 0.09 |
| N-NO3 (mg kg−1) 2* | 66.70 | 108.45 |
| Total nitrogen (TN %) 3* | 0.10 | 0.18 |
| Soil organic carbon % 3* | 0.81 | 2.07 |
| pH in H2O 4* | 5.82 | 6.39 |
| Available phosphorus (mg P2O5 100 g−1) 5* | 17.90 | 18.00 |
| Available potasium (mg K2O 100 g−1) 6* | 26.10 | 27.90 |
| Exhangeable Ca (cmol kg−1) 7* | 2.02 | 5.49 |
| Exhangeable Mg (cmol kg−1) 7* | 0.44 | 1.14 |
| Exhangeable K (cmol kg−1) 7* | 0.49 | 0.44 |
| Exhangeable Na (cmol kg−1) 7* | 0.13 | 0.13 |
| Soil | Date | Distance | Biodiversity Indices | |||
|---|---|---|---|---|---|---|
| AWCD | H’ | E | 1-D | |||
| sandy loam | June | 0.1 m | 1.75 cd | 3.29 bc | 0.97 ab | 0.98 bc |
| 1 m | 1.48 e | 3.27 c | 0.98 ab | 0.98 a | ||
| sandy loam | October | 0.1 m | 2.01 b | 3.33 abc | 0.98 ab | 0.98 bc |
| 1 m | 1.79 cd | 3.33 abc | 0.98 a | 0.98 b | ||
| loamy sand | June | 0.1 m | 2.14 ab | 3.35 ab | 0.98 a | 0.98 c |
| 1 m | 1.66 de | 3.27 c | 0.96 b | 0.98 bc | ||
| loamy sand | October | 0.1 m | 2.23 a | 3.36 a | 0.98 a | 0.98 c |
| 1 m | 1.94 bc | 3.36 a | 0.99 a | 0.98 ab | ||
| Variables | AWCD | H’ | E | 1-D | |||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | ||
| Main effects | Soil | 62.2 | 0.00 | 8 | 0.01 | 0.2 | 0.69 | 15 | 0.00 |
| Date | 62.4 | 0.00 | 25.1 | 0.00 | 13.7 | 0.00 | 1 | 0.25 | |
| Distance | 110.9 | 0.00 | 6.4 | 0.02 | 0.6 | 0.46 | 56 | 0.00 | |
| Interaction effects | Soil * Date | 3.4 | 0.08 | 0.30 | 0.59 | 2.1 | 0.16 | 10 | 0.01 |
| Soil * Distance | 5.3 | 0.03 | 3.9 | 0.07 | 11.7 | 0.00 | 4 | 0.08 | |
| Date * Distance | 4.1 | 0.06 | 6.9 | 0.02 | 4.5 | 0.05 | 0 | 0.98 | |
| Soil * Date * Distance | 1.3 | 0.27 | 4.1 | 0.06 | 4.8 | 0.04 | 8 | 0.01 | |
| Parameters | DHA | AlP | AcP | MBC | MBN | pH | AWCD | H’ | E | 1-D | AandA | A | CandAA | C |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DHA | ||||||||||||||
| AlP | −0.36 | |||||||||||||
| AcP | 0.79 * | −0.38 | ||||||||||||
| MBC | 0.64 * | −0.01 | 0.49 | |||||||||||
| MBN | 0.06 | −0.39 | 0.17 | −0.07 | ||||||||||
| pH | 0.88 * | −0.35 | 0.68 * | 0.65 * | 0.14 | |||||||||
| AWCD | 0.87 * | −0.44 | 0.75 * | 0.78 * | 0.22 | 0.86 * | ||||||||
| H’ | 0.64 * | −0.53 | 0.64 * | 0.71 * | 0.18 | 0.49 | 0.80 * | |||||||
| E | −0.19 | −0.20 | −0.04 | 0.17 | −0.33 | −0.35 | −0.01 | 0.46 | ||||||
| 1-D | −0.74 * | 0.34 | −0.67 * | −0.42 | −0.43 | −0.87 * | −0.71 * | −0.25 | 0.622 * | |||||
| AandA | −0.36 | 0.66 * | −0.28 | −0.28 | 0.04 | −0.45 | −0.30 | −0.36 | −0.03 | 0.30 | ||||
| A | −0.37 | 0.03 | −0.02 | −0.50 | −0.24 | −0.45 | −0.54 | −0.41 | 0.09 | 0.24 | 0.00 | |||
| CandAA | −0.22 | 0.01 | −0.30 | −0.62 * | −0.16 | −0.27 | −0.49 | −0.38 | −0.04 | 0.32 | −0.10 | 0.14 | ||
| C | −0.01 | −0.51 | −0.11 | −0.19 | 0.13 | 0.02 | −0.20 | −0.15 | −0.22 | −0.11 | −0.73 * | 0.25 | 0.22 | |
| P | 0.76 * | −0.31 | 0.64 * | 0.64 * | −0.15 | 0.81 * | 0.70 * | 0.54 | −0.06 | −0.57 | −0.68 * | −0.24 | 0.00 | 0.10 |
| Parameters | DHA | AlP | AcP | MBC | MBN | pH | AWCD | H’ | E | 1− D | AandA | A | CandAA | C |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DHA | ||||||||||||||
| AlP | 0.09 | |||||||||||||
| AcP | 0.87 * | 0.04 | ||||||||||||
| MBC | 0.36 | −0.25 | 0.61 * | |||||||||||
| MBN | 0.87 * | 0.07 | 0.70 * | 0.21 | ||||||||||
| pH | 0.19 | −0.18 | 0.53 | 0.81 * | 0.06 | |||||||||
| AWCD | 0.78 * | 0.10 | 0.87 * | 0.71 * | 0.65 * | 0.69 * | ||||||||
| H’ | 0.80 * | 0.45 | 0.56 | 0.03 | 0.73 * | −0.10 | 0.55 | |||||||
| E | 0.80 * | 0.45 | 0.56 | 0.03 | 0.73 * | −0.10 | 0.55 | 1.00 | ||||||
| 1−D | −0.17 | 0.39 | −0.35 | −0.68 * | 0.04 | −0.71 * | −0.57 | 0.22 | 0.22 | |||||
| AandA | −0.53 | −0.45 | −0.57 | −0.32 | −0.54 | −0.23 | −0.52 | −0.57 | −0.57 | −0.08 | ||||
| A | −0.39 | −0.38 | −0.27 | 0.00 | −0.33 | −0.19 | −0.46 | −0.36 | −0.36 | 0.11 | 0.11 | |||
| CandAA | 0.30 | −0.44 | 0.25 | −0.05 | 0.43 | 0.10 | 0.22 | 0.08 | 0.08 | −0.29 | −0.13 | 0.15 | ||
| C | −0.12 | 0.01 | −0.20 | −0.13 | 0.06 | −0.48 | −0.23 | −0.06 | −0.06 | 0.09 | −0.11 | 0.52 | 0.15 | |
| P | 0.12 | 0.57 | 0.15 | 0.17 | 0.11 | 0.15 | 0.25 | 0.25 | 0.25 | 0.20 | −0.53 | −0.59 * | −0.56 | −0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woźniak, M.; Gałązka, A.; Siebielec, G.; Frąc, M. Can the Biological Activity of Abandoned Soils Be Changed by the Growth of Paulownia elongata × Paulownia fortunei?—Preliminary Study on a Young Tree Plantation. Agriculture 2022, 12, 128. https://doi.org/10.3390/agriculture12020128
Woźniak M, Gałązka A, Siebielec G, Frąc M. Can the Biological Activity of Abandoned Soils Be Changed by the Growth of Paulownia elongata × Paulownia fortunei?—Preliminary Study on a Young Tree Plantation. Agriculture. 2022; 12(2):128. https://doi.org/10.3390/agriculture12020128
Chicago/Turabian StyleWoźniak, Małgorzata, Anna Gałązka, Grzegorz Siebielec, and Magdalena Frąc. 2022. "Can the Biological Activity of Abandoned Soils Be Changed by the Growth of Paulownia elongata × Paulownia fortunei?—Preliminary Study on a Young Tree Plantation" Agriculture 12, no. 2: 128. https://doi.org/10.3390/agriculture12020128
APA StyleWoźniak, M., Gałązka, A., Siebielec, G., & Frąc, M. (2022). Can the Biological Activity of Abandoned Soils Be Changed by the Growth of Paulownia elongata × Paulownia fortunei?—Preliminary Study on a Young Tree Plantation. Agriculture, 12(2), 128. https://doi.org/10.3390/agriculture12020128