Antimicrobial Peptides from Fish

Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Piscidins | Defensins | Hepcidins | Cathelicidins | Histone-derived | ||
|---|---|---|---|---|---|---|---|
| Common name | Scientific name | Habitat | |||||
| American plaice | Hippoglossoides platessoides | Marine | 2 [8] | ||||
| Antarctic toothfish | Dissostichus mawsoni | Marine | 3 [9] | ||||
| Atlantic cod | Gadus morhua | Marine and brackish | 2 [10] | 1 [11] | 1 [12] | 1 [13] | 1 [14] |
| Antarctic eelpout | Lycodichthys dearborni | Marine | 2 [9] | ||||
| Atlantic hagfish | Myxine glutinosa | Marine | 3 [15] | ||||
| Atlantic salmon | Salmo salar | Marine, brackish and freshwater | 2 [16] | 2 [17] | 1 [18] | ||
| Ayu | Plecoglossus altivelis | Marine, brackish and freshwater | 1 [19] | 1 [20] | |||
| Barramundi | Lates calcarifer | Marine, brackish and freshwater | 2 [21] | ||||
| Black porgy | Acanthopagrus schlegelii | Marine and brackish | 7 [22,23] | ||||
| Black rockfish | Sebastes schlegelii | Marine | 2 [24] | ||||
| Blotched snakehead | Channa maculata | Freshwater | 1 [25] | ||||
| Blue catfish | Ictalurus furcatus | Freshwater and brackish | 1 [26] | ||||
| Blunt snout bream | Megalobrama amblycephala | Freshwater | 1 [27] | ||||
| Brook trout | Salvelinus fontinalis | Marine, brackish and freshwater | 2 [28] | ||||
| Brown trout | Salmo trutta fario | Marine, brackish and freshwater | 1 [28] | ||||
| Channel catfish | Ictalurus punctatus | Freshwater | 1 [26] | 1 [29] | |||
| Chinese loach | Paramisgurnus dabryanus | Freshwater | 1 [30] | ||||
| Common carp | Cyprinus carpio L. | Freshwater and brackish | 2 [31] | 1 [32] | |||
| European seabass | Dicentrarchus labrax | Marine, brackish and freshwater | 1 [33] | 1 [34] | |||
| Gilthead seabream | Sparus aurata | Marine and brackish | 1 [35] | 1 [36] | |||
| Grayling | Thymallus thymallus | Freshwater and brackish | 1 [28] | ||||
| Half-smooth tongue sole | Cynoglossus semilaevis | Marine, brackish and freshwater | 1 [37] | ||||
| Atlantic halibut | Hippoglossus hippoglossus | Marine | 1 [8] | 1 [38] | |||
| Hybrid striped bass | Morone saxatilis x M. chrysops | Marine, brackish and freshwater | 4 [39,40,41] | 1 [42] | |||
| Icefish | Chionodraco hamatus | Marine | 1 [43] | ||||
| Olive flounder | Paralichthys olivaceus | Marine | 2 [16] | ||||
| Japanese rice fish | Oryzias latipes | Freshwater and brackish | 1 [16] | ||||
| Japanese pufferfish | Takifugu rubripes | Marine, brackish and freshwater | 1 [44] | ||||
| Japanese seabass | Lateolabrax japonicus | Marine, brackish and freshwater | 1 [45] | ||||
| Largemouth bass | Micropterus salmoides | Freshwater | 2 [46] | ||||
| Large yellow croaker | Pseudosciaena crocea | Marine and brackish | 1 [47] | 1 [48,49] | |||
| Mandarin fish | Siniperca chuatsi | Freshwater | 1 [50] | 1 [51] | |||
| Maori chief | Notothenia angustata | Marine | 5 [9] | ||||
| Medaka | Oryzias melastigma | Freshwater and brackish | 1 [52] | 2 [53] | |||
| Miiuy croaker | Miichthys miiuy | Marine and brackish | 1 [54] | ||||
| Mud dab | Limanda limanda | Marine | 1 [55] | ||||
| Mud loach | Misgurnus mizolepis | Freshwater | [56] | ||||
| Olive flounder | Paralichthys olivaceus | Marine | 5 [57] | ||||
| Orange-spotted grouper | Epinephelus coioides | Marine and brackish | 1 [58] | 2 [59,60] | 3 [61,62] | ||
| Pacific mutton hamlet | Alphestes immaculatus | Marine | 1 [63] | ||||
| Rainbow trout | Oncorhynchus mykiss | Marine, brackish and freshwater | 4 [64,65] | 2 [17] | 3 [66,67,68,69] | ||
| Redbanded seabream | Pagrus auriga | Marine | 4 [70] | ||||
| Red sea bream | Chrysophrys major | Marine | 1 [71] | 1 [72] | |||
| Rockbream | Oplegnathus fasciatus | Marine | 4 [73] | ||||
| Sea bass | Dicentrarchus labrax | Marine, brackish and freshwater | 1 [74] | ||||
| Seahorse | Hippocampus kuda | Marine and brackish | 1 [75] | ||||
| Smallmouth bass | Micropterus dolomieu | Freshwater | 2 [46] | ||||
| Snowtrout | Schizothorax richardsonii | Freshwater | 1 [76] | ||||
| Spotted-green pufferfish | Tetraodon nigroviridis | Freshwater and brackish | 2 [44] | ||||
| Sunshine bass | Marine, brackish and freshwater | 1 [69] | |||||
| Thick-lipped lenok | Brachymystax lenok | Freshwater | 1 [77] | ||||
| Tilapia | Oreochromis mossambicus | Freshwater and brackish | 5 [78] | 3 [79] | |||
| Turbot | Scophthalmus maximus | Marine and brackish | 2 [80,81] | ||||
| Winter flounder | Pleuronectesamericanus | Marine | 6 [6,82,83] | 5 [16] | |||
| Witch flounder | Glyptocephalus cynoglossus | Marine | 5 [8] | ||||
| Yellowtail flounder | Pleuronectes ferruginea | Marine | 1 [8] | ||||
| Zebrafish | Danio rerio | Freshwater | 3 [44] | 2 [84] | |||
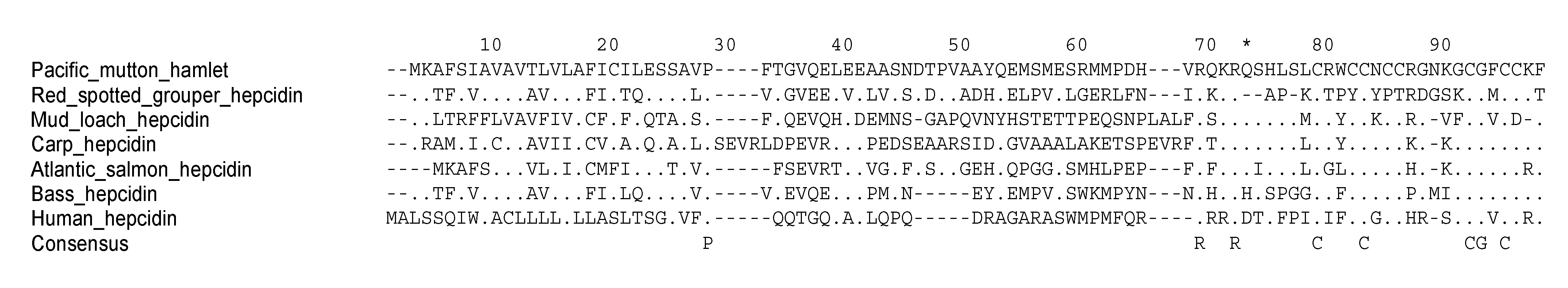
2. Piscidins

3. β-Defensins

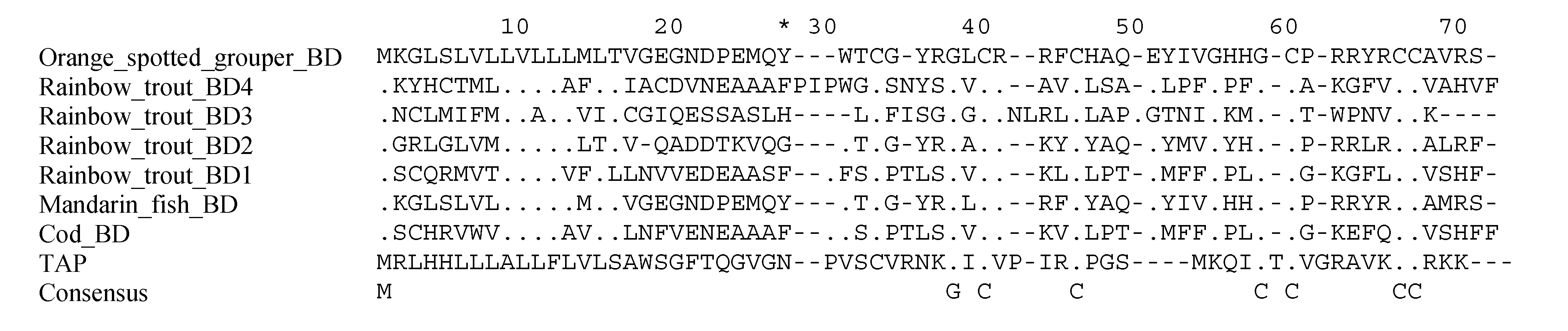
4. Hepcidins
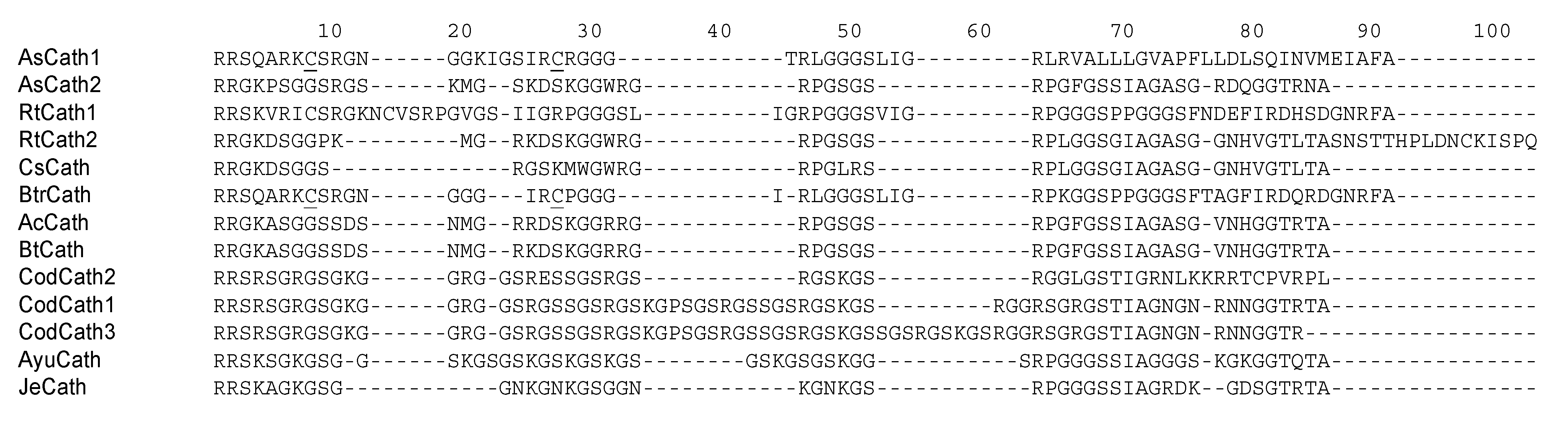
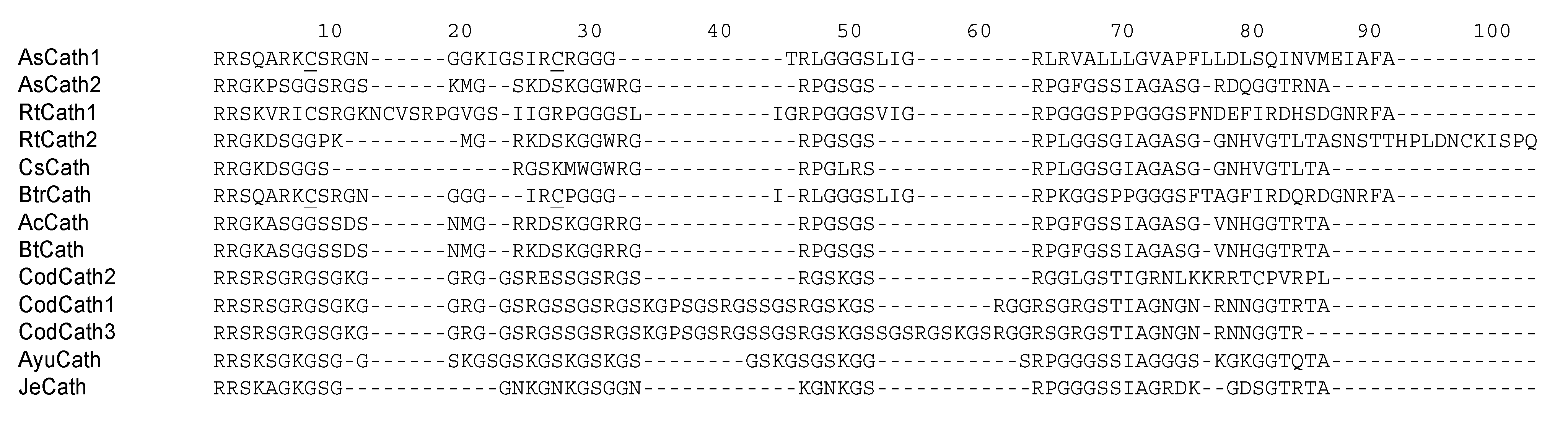
5. Cathelicidins
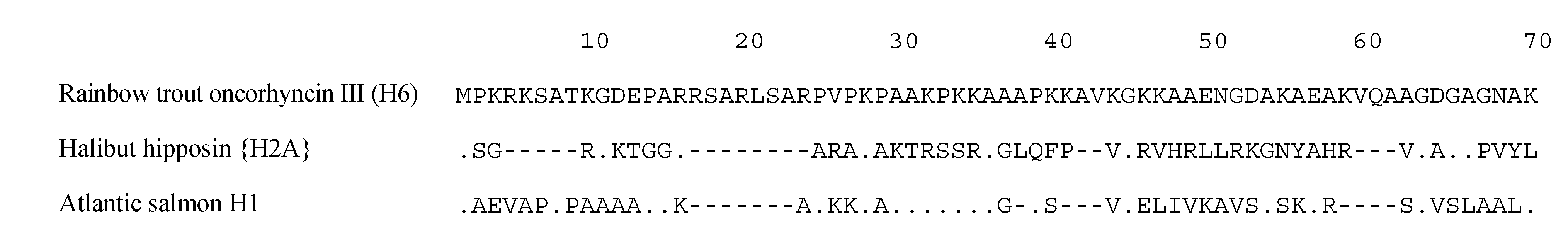
6. Histone-Derived Peptides
7. Therapeutics
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Supplementary Files
| Bacteria | β-Defensin | Piscidin | Hepcdin | Cathelicidin | Histone-derived | Pathogenic | Habitat | |
|---|---|---|---|---|---|---|---|---|
| Fish | Human | |||||||
| Gram negative | ||||||||
| Escherichia coli | 39.0 ± 3.7 (vLD90) [60] 32.6 ± 1.5 (vLD90) [52] IH [10] | 5–10 μM (MIC) [50] 2–10 μM (MIC) [117] 50 μg/mL (MIC) [40] 2.2–3.3 μM (MIC) [6] 5 μM (MIC) [43] 3–6 μM (MIC) [47] 2 μg/mL (MIC) [83] 3 μM (MIC) [75] 1 μg/mL (MIC) [8] | 5–10 μM (MIC) [23] >96 μM (MIC) [62] 6-12 μM (MIC) [171] 18.66 (IC50) [36] 11 μM (MIC) [161] 12–24 μM (MIC) [48] >8.18 μM (MIC) [37] 34.56 mm2 (IH) [49] | <1 μg/mL (MIC) [187] 1 μg/mL (MIC) [225] [20] 2–4 μg/mL (MIC) [77] 5 μg/mL (MIC) [186] | 1 μg/mL(MIC) [29] >10 μg/mL(MIC) [14] 2.5 μg/mL (MIC) [38] | X | X | Freshwater and others |
| Plesiomonas shigelloides | 11 μM (MIC) [161] | X | X | Freshwater | ||||
| Klebsiella pneumoniae | 12.6 μg/mL (MIC) [82] 2.5–5 μM (MIC) [41] | 22 μM (MIC) [161] | 18.8 μM (MIC) [77] | X | Other | |||
| Klebsiella oxytoca | 5–10 μM (MIC) [41] 12.5 μg/mL (MIC) [58] | >44 μM (MIC) [161] | X | Other | ||||
| Shigella sonnei | 5–10 μM (MIC) [41] | >44 μM (MIC) [161] | X | Other | ||||
| Shigella flexneri | 3.1 μg/mL (MIC) [40] 2.5–5 μM (MIC) [41] | >96 μM (MIC) [62] 22 μM (MIC) [161] | X | Freshwater and others | ||||
| Yersinia enterocolitica | 2.5–5 μM (MIC) [41] 100 μM (MIC) [58] | 22 μM (MIC) [161] | X | X | Freshwater and others | |||
| Yersinia ruckeri | 20–40 μM (MIC) [50] | X | Freshwater | |||||
| Aeromonas salmonicida | >50 μM (MIC) [11] | 17.7–35 μM (MIC) [6] 2 μg/mL (MIC) [83] 1 μg/mL (MIC) [8] 5 μM (MLC) [71] | >44 μM (MIC) [161] | 9.4 μg/mL (MIC)[77] 1–5 μg/mL (MIC) [225] 50 μg/mL (MIC) [20] 10 μg/mL (MIC) [186] | 20 μg/mL (MIC) [38] >1.2 μg/mL (MIC) [66] | X | Freshwater and marine | |
| Aeromonas hydrophila | 13.4 ± 0.7 [52] IH [51] | 1.05 μg/mL (MIC) [78] 19.78 μg/mL (MIC) [78] >21.4 μg/mL (MIC) [78] >160 [50] >20 [41] >96 [47] | 10–20 μM (MIC) [23] >96 μM (MIC) [62] 1.5–3 μM (MIC) [171] >44 μM (MIC) [161] 3–6 μM (MIC) [48] 31.66 mm2 (IH) [49] | X | X | Freshwater and others | ||
| Aeromonas sobria | 59.4 ± 8.8 [60] | 10–20 μM (MIC) [50] | 2.5 μM (MIC) [38] | X | X | Freshwater and others | ||
| Aeromonas punctata | 10–20 μM (MIC) [50] | X | Freshwater and others | |||||
| Vibrio anguillarum | >50 μM (MIC) [11] 58.2 ± 23.4 (vLD90) [60] 21.1 ± 1 (vLD90) [52] | 20–40 μM (MIC) [50] 6.3 μg/mL (MIC) [40] 1.25 μM (MLC) [71] | 2.92 μM (MIC) [37] | 5 μM (MIC) [187] 0.5–2.5 μM (MIC) [225] 5 μM (MIC) [186] | X | Marine | ||
| Vibrio parahaemolyticus | >60 μM (MIC) [23] >96 μM (MIC) [62] >48 μM (MIC) [171] 3–6 μM (MIC) [48] 5.84 μM (MIC) [37] 94.25 mm2 (IH) [49] | 25 [20] | X | X | Marine | |||
| Vibrio fluvialis | 43.0 ± 10 (vLD90) [60] 35.1 ± 2.7 (vLD90) [52] | >96 μM (MIC) [62] 123.11 mm2 (IH) [49] | 3.1 μg/mL (MIC) [20] | X | X | Marine | ||
| Vibrio harveyi | 12.5 μg/mL (MIC) [58] 5 μM (MLC) [71] | 20–40 μM (MIC) [23] >96 μM (MIC) [62] 6–12 μM (MIC) [48] 5.84 μM (MIC) [37] 113.10 mm2 (IH) [49] | 6.2 μg/mL (MIC) [20] | X | Marine | |||
| Vibrio alginolyticus | 0.03 μg/mL (MIC) [78] 1.24 μg/mL (MIC) [78] 2.68 μg/mL (MIC) [78] | >60 μM (MIC) [23] >96 μM (MIC) [62] >48 μM (MIC) [171] 12–24 μM (MIC) [48] 118.06 mm2 (IH) [49] | X | X | Marine | |||
| Vibrio vulnificus | 0.03 μg/mL (MIC) [78] 0.62 μg/mL (MIC) [78] 0.67 μg/mL (MIC) [78] 6.25 μg/mL (MIC) [58] 2.5 μM (MLC) [71] | 20 μM (MIC) [61,163] | X | X | Marine | |||
| Vibrio cholera | 2.5–5 μM (MIC) [41] | X | X | Freshwater, marine and others | ||||
| Vibrio damsela | 48 μM (MIC) [75] | X | X | Marine | ||||
| Vibrio penaeicida | 5 μM (MLC) [71] | Marine | ||||||
| Salinivibrio costicola | 3.125 μg/mL (MIC) [58] | Marine and others | ||||||
| Edwardsiella tarda | >44 μM (MIC) [161] 2.92 μM (MIC) [37] | X | X | Marine and freshwater | ||||
| Riemerella anatipestifer | 6.25 μg/mL (MIC) [114] | 25 μg/mL (MIC) [114] | Other | |||||
| Pseudomonas aeruginosa | 44.5 ± 11.8 [60] | 0.52 μg/mL (MIC) [78] >19.78 μg/mL (MIC) [78] 10.70 μg/mL (MIC) [78] 60 μg/mL (MIC) [58] 1 μg/mL (MIC) [83] 28 μg/mL (MIC) [82] >35 μM (MIC) [6] 5–10 μM (MIC) [41] | >44 μM (MIC) [161] >96 μM (MIC) [62] | 12.5 μg/mL (MIC) [20] 1–4 μg/mL (MIC) [77] | X | X | Freshwater, marine and others | |
| Pseudomonas fluorescens | 10–20 μM (MIC) [50] 1.5–3 μM (MIC) [47] | 24–48 μM (MIC) [62] | 20 μM (MIC) [38] | X | X | Freshwater and others | ||
| Pseudomonas stutzeri | <1.5 μM (MIC) [62] 3–6 μM (MIC) [171] | X | Other | |||||
| Enterobacter cloacae | 10–20 μM (MIC) [41] 100 μg/mL (MIC) [58] | >44 μM (MIC) [161] | X | X | Freshwater and others | |||
| Enterobacter aerogenes | 10–20 μM (MIC) [41] 25 μg/mL (MIC) [58] | X | Others | |||||
| Salmonella arizonae | 10–20 μM (MIC) [41] | >44 μM (MIC) [161] | X | Other | ||||
| Salmonella choleraesuis | 10–20 μM (MIC) [41] 6 μM (MIC) [75] | >44 μM (MIC) [161] | X | Other | ||||
| Salmonella typhimurium | 8.8–17.7 μM (MIC) [6] 10–20 μM (MIC) [41] 2 μg/mL (MIC) [83] 1 μg/mL (MIC) [8] | >44 μM (MIC) [161] | 2 μM (MIC) [29] | X | Other | |||
| Serratia marcescens | >35 μM (MIC) [6] >20 μM (MIC) [41] | >44 μM (MIC) [161] | 4 μM (MIC) [29] | X | Other | |||
| Cytophaga columnare | 40–80 μM (MIC) [50] | X | Freshwater | |||||
| Cytophaga aquatilis | 2.2–4.4 μM (MIC) [6] | X | Freshwater | |||||
| Proteus vulgaris | 2–10 μM (MIC) [117]X | X | X | Freshwater and others | ||||
| Photobacterium damsela subsp. piscidida | 1.5 μg/mL (MIC) [40] | X | Marine and freshwater | |||||
| Pasteurella haemolytica | 4.4–8.8 μM (MIC) [6] | X | Other | |||||
| Burkholderia cepacia | >20 μM (MIC) [41] | X | Freshwater and others | |||||
| Moraxella catarrhalis | 2.5–5 μM (MIC) [41] | X | Other | |||||
| Neisseria gonorrhoeae | >20 μM (MIC) [41] | X | Other | |||||
| Psychrobacter sp. | 10 μM (MIC) [43] | Marine and others | ||||||
| Morganella morganii | 12 μM (MIC) [75] | X | Other | |||||
| Enterococcus faecium | 15 μM (MIC) [75] | X | Other | |||||
| Gram positive | ||||||||
| Micrococcus luteus | 25–50 μM (MIC) [11] 296.5 ± 65.5 (vLD90) [60] 311 ± 15.6 (vLD90) [52] | 10–20 μM (MIC) [41] 3.125 μg/mL (MIC) [58] | >96 μM (MIC) [62] 1.5–3 μM (MIC) [48] 2.5–5 μM (MIC) [23] 24–48 μM (MIC) [62] 34.56 mm2 (IH) [49] | X | Freshwater and others | |||
| Staphylococcus aureus | 358.5 ± 46.5 (vLD90) [60] 229.8 ± 12.8 (vLD90) [52] IH [51] | 0–2 μM (MIC) [117] 3.1 μg/mL (MIC) [40] 17.7–35 μM (MIC) [6] 62.5 μg/mL (MIC) [82] 1.25–2.5 μM (MIC) [41] 6–12 μM (MIC) [47] 6.25 μg/mL (MIC) [58] 8 μg/mL (MIC) [83] 1.5 μM (MIC) [75] | 1.25–2.5 μM (MIC) [53] 1.5–3 μM (MIC) [62] 1.5–3 μM (MIC) [171] >44 μM (MIC) [161] 3–6 μM (MIC) [48] 1.25–2.5 μM (MIC) [23] >8.18 μM (MIC) [37] 47.12 mm2 (IH) [49] | 11–45 μM (MIC) [77] | 2μM (MIC) [29] | X | Other | |
| Staphylococcus epidermidis | 5–10 μM (MIC) [41] 12.5 μg/mL (MIC) [58] 8 μg/mL (MIC) [83] 4 μg/mL (MIC) [8] | 20–40 μM (MIC) [23] >96 μM (MIC) [62] 6–12 μM (MIC) [48] | 10 μM (MIC) [38] | X | Other | |||
| Staphylococcus saprophiticus | 5–10 μM (MIC) [41] 7.5 μM (MIC) [75] | X | Other | |||||
| Staphylococcus haemolyticus | 7.5 μM (MIC) [75] | X | Other | |||||
| Staphylococcus xylosus | >20 μM (MIC) [41] 50 μg/mL (MIC) [58] | Other | ||||||
| Staphylococcus warneri | 15 μM (MIC) [75] | Other | ||||||
| Bacillus subtilis | 1.1–2.2 μM (MIC) [6] 0.75–1.5 μM (MIC) [47] 48 μM (MIC) [75] | 5–10 μM (MIC) [23] >96 μM (MIC) [62] 11.41 (IC50) [36] 3–6 μM (MIC) [48] 28.86 mm2 (IH) [49] | 11 μg/mL (MIC) [29] 0.6 μg/mL (MIC) [66] 1.3 μg/mL (MIC) [38] | Other | ||||
| Bacillus cereus | 17.5 ± 4 (vLD90) [60] | 0–2 μM (MIC) [117] 5 μM (MIC) [43] | 40–60 μM (MIC) [23] >96 μM (MIC) [62] 12–24 μM (MIC) [48] | X | Other | |||
| Corynebacterium glutamicum | 2.5–5 μM (MIC) [23] 48–96 μM (MIC) [62] 3–6 μM (MIC) [171] 1.5–3 μM (MIC) [48] | Other | ||||||
| Planococcus citreus | 0.4–0.8 μM (MIC) [11] | 0.08 μg/mL (MIC) [66] | Marine | |||||
| Enterococcus faecalis | 8.39 μg/mL (MIC) [78] | >44 μM (MIC) [161] | X | Other | ||||
| Listeria monocytogenes | 2.5–5 μM (MIC) [41] 25 μg/mL (MIC) [58] | X | Other | |||||
| Streptococcus agalactiae | 0.13 μg/mL (MIC) [78] | X | X | Freshwater, marine and others | ||||
| Streptococcus iniae | 3.1 μg/mL (MIC) [40] 1.25–2.5 μM (MIC) [41] 1.5 μM (MLC) [71] | X | X | Freshwater, marine and others | ||||
| Streptococcus mutans | 2.2 μM (MIC) [207] 8 μg/mL (MIC) [210] 4 μg/mL (MIC) [209] | 1 μg/mL (MIC) [29] | X | Other | ||||
| Streptococcus sobrinus | 8 μg/mL (MIC) [210] 4 μg/mL (MIC) [209] | X | Other | |||||
| Streptococcus sanguinis | 32 μg/mL (MIC) [210] 4–8 μg/mL (MIC) [209] | X | Other | |||||
| Streptococcus gordonii | 8 μg/mL (MIC) [210] 8 μg/mL (MIC) [209] | X | Other | |||||
| Streptococcus bovis | 1.25–2.5 μM (MIC) [41] | X | Other | |||||
| Streptococcus equisimilis | 2.5–5 μM (MIC) [41] | X | Other | |||||
| Streptococcus mitis | 1.25–2.5 μM (MIC) [41] | X | Other | |||||
| Streptococcus pneumoniae | 1.25–2.5 μM (MIC) [41] 12.5 μg/mL (MIC) [58] | X | Other | |||||
| Streptococcus pyogenes | 1.25–2.5 μM (MIC) [41] 25 μg/mL (MIC) [58] | X | Other | |||||
| Lactococcus garviae | 3.1 μg/mL (MIC) [40] 5 μM (MLC) [71] | X | X | Marine and others | ||||
| Leucothrix mucor | >35 μM (MIC) [6] | X | Marine | |||||
| Lactobacillus acidophilus | 128 μg/mL (MIC) [210] 4–8 μg/mL (MIC) [209] | Other | ||||||
| Lactobacillus casei | 32 μg/mL (MIC) [210] 4 μg/mL (MIC) [209] | Other | ||||||
| Lactobacillus fermenti | 2 μg/mL (MIC) [210] 4 μg/mL (MIC) [209] | Other | ||||||
| Actinomyces viscosus | 8 μg/mL (MIC) [209] | X | Other | |||||
| Actinomyces naeslundii | 8 μg/mL (MIC) [209] | X | Other | |||||
| Fungi | ||||||||
| Aspergillus fumigatus | 50–100 μM (MIC) [41] | X | Other | |||||
| Aspergillus niger | 48–96 μM (MIC) [47] | 20–40 μM (MIC) [23] 44 μM (MIC) [161] 12–24 μM (MIC) [48] | X | Other | ||||
| Fusarium graminearum | 20–40 μM (MIC) [23] 12–24 μM (MIC) [48] | Other | ||||||
| Fusarium solani | 20–40 μM (MIC) [23] 12–24 μM (MIC) [48] | X | Other | |||||
| Fusarium oxysporum | 0.78–1.56 μM (MIC) [41] | X | X | Freshwater, marine and others | ||||
| Fusarium culmorum | 0.39–0.78 [41] | Other | ||||||
| Candida albicans | 15.4 μg/mL (MIC) [82] 10–20 μM (MIC) [41] 24–48 μM (MIC) [47] 8 μg/mL (MIC) [83] 96 μM (MIC) [75] 4 μg/mL (MIC) [8] 5 μM (MIC) [115] 6.25 μM (MIC) [226] | >60 μM (MIC) [23] >96 μM (MIC) [62] >44 μM (MIC) [161] >48 μM (MIC) [48] | 2.3 [77] 2.5 [186] | X | Other | |||
| Candida glabrata | 10–20 μM (MIC) [41] | X | Other | |||||
| Candida lusitania | 10–20 μM (MIC) [41] | |||||||
| Candida tropicalis | 10–20 μM (MIC) [41] | X | Other | |||||
| Saccharomyces cerevisiae | 384 μM (MIC) [75] 5 μM (MIC) [115] | X | Other | |||||
| Pichia pastoris | >60 μM (MIC) [23] >96 μM (MIC) [62] >48 μM (MIC) [48] | Other | ||||||
| Saprolegnia sp | 12.5–25 μg/mL (MOC) [98] | X | Freshwater and others | |||||
| Neurospora crassa | 1.56–3.12 μM (MIC) [41] | Other | ||||||
| Trichosporon beigelii | 2.5 μM (MIC) [115] 1.56 μM (MIC) [226] | X | Other | |||||
| Malassezia furfur | 6.25 μM (MIC) [226] | X | Other | |||||
| Parasites | ||||||||
| Trichomonas vaginalis | 12.5 μg/mL (MIC) [115] | X | Other | |||||
| Trichodina | 12.5–100 μg/mL (PCmin) [97] | X | Freshwater and marine | |||||
| Cryptocaryon theront | 12.5 μg/mL (PCmin) [97] | X | Marine | |||||
| Amyloodinium dinospore | 6.3 μg/mL (PCmin) [97] | X | Marine | |||||
| Ichthyophthirius theront | 6.3 μg/mL (PCmin) [97] | X | Freshwater | |||||
| Virus | ||||||||
| VHSV | [64] | [80] | X | Marine | ||||
| NNV | [59] | [227] | [99,169] | X | Marine | |||
| IPNV | [170] | X | Marine | |||||
| RGV | [60] | X | Marine and Freshwater | |||||
| SGIV | [59] | [61] | X | Marine | ||||
| CCV | [100] | X | Freshwater | |||||
| FV3 | [100] | Other | ||||||
References
- Ganz, T.; Selsted, M.E.; Szklarek, D.; Harwig, S.S.; Daher, K.; Bainton, D.F.; Lehrer, R.I. Defensins. Natural peptide antibiotics of human neutrophils. J. Clin. Investig. 1985, 76, 1427–1435. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Selsted, M.E.; Szklarek, D.; Fleischmann, J. Antibacterial activity of microbicidal cationic proteins 1 and 2, natural peptide antibiotics of rabbit lung macrophages. Infect. Immun. 1983, 42, 10–14. [Google Scholar]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef]
- Primor, N.; Tu, A.T. Conformation of pardaxin, the toxin of the flatfish Pardachirus marmoratus. Biochim. Biophys. Acta 1980, 626, 299–306. [Google Scholar] [CrossRef]
- Oren, Z.; Shai, Y. A class of highly potent antibacterial peptides derived from pardaxin, a pore-forming peptide isolated from Moses sole fish Pardachirus marmoratus. Eur. J. Biochem. 1996, 237, 303–310. [Google Scholar]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef]
- Tessera, V.; Guida, F.; Juretic, D.; Tossi, A. Identification of antimicrobial peptides from teleosts and anurans in expressed sequence tag databases using conserved signal sequences. FEBS J. 2012, 279, 724–736. [Google Scholar] [CrossRef]
- Patrzykat, A.; Gallant, J.W.; Seo, J.K.; Pytyck, J.; Douglas, S.E. Novel antimicrobial peptides derived from flatfish genes. Antimicrob. Agents Chemother. 2003, 47, 2464–2470. [Google Scholar] [CrossRef]
- Xu, Q.; Cheng, C.H.; Hu, P.; Ye, H.; Chen, Z.; Cao, L.; Chen, L.; Shen, Y.; Chen, L. Adaptive evolution of hepcidin genes in antarctic notothenioid fishes. Mol. Biol. Evol. 2008, 25, 1099–1112. [Google Scholar] [CrossRef]
- Browne, M.J.; Feng, C.Y.; Booth, V.; Rise, M.L. Characterization and expression studies of Gaduscidin-1 and Gaduscidin-2; paralogous antimicrobial peptide-like transcripts from Atlantic cod (Gadus morhua). Dev. Comp. Immunol. 2011, 35, 399–408. [Google Scholar] [CrossRef]
- Ruangsri, J.; Kitani, Y.; Kiron, V.; Lokesh, J.; Brinchmann, M.F.; Karlsen, B.O.; Fernandes, J.M. A novel beta-defensin antimicrobial peptide in Atlantic cod with stimulatory effect on phagocytic activity. PLoS One 2013, 8, e62302. [Google Scholar]
- Solstad, T.; Larsen, A.N.; Seppola, M.; Jorgensen, T.O. Identification, cloning and expression analysis of a hepcidin cDNA of the Atlantic cod (Gadus morhua L.). Fish Shellfish Immunol. 2008, 25, 298–310. [Google Scholar] [CrossRef]
- Broekman, D.C.; Zenz, A.; Gudmundsdottir, B.K.; Lohner, K.; Maier, V.H.; Gudmundsson, G.H. Functional characterization of codCath, the mature cathelicidin antimicrobial peptide from Atlantic cod (Gadus morhua). Peptides 2011, 32, 2044–2051. [Google Scholar] [CrossRef]
- Bergsson, G.; Agerberth, B.; Jornvall, H.; Gudmundsson, G.H. Isolation and identification of antimicrobial components from the epidermal mucus of Atlantic cod (Gadus morhua). FEBS J. 2005, 272, 4960–4969. [Google Scholar] [CrossRef]
- Uzzell, T.; Stolzenberg, E.D.; Shinnar, A.E.; Zasloff, M. Hagfish intestinal antimicrobial peptides are ancient cathelicidins. Peptides 2003, 24, 1655–1667. [Google Scholar] [CrossRef]
- Douglas, S.E.; Gallant, J.W.; Liebscher, R.S.; Dacanay, A.; Tsoi, S.C. Identification and expression analysis of hepcidin-like antimicrobial peptides in bony fish. Dev. Comp. Immunol. 2003, 27, 589–601. [Google Scholar] [CrossRef]
- Chang, C.I.; Zhang, Y.A.; Zou, J.; Nie, P.; Secombes, C.J. Two cathelicidin genes are present in both rainbow trout (Oncorhynchus mykiss) and atlantic salmon (Salmo salar). Antimicrob. Agents Chemother. 2006, 50, 185–195. [Google Scholar] [CrossRef]
- Richards, R.C.; O’Neil, D.B.; Thibault, P.; Ewart, K.V. Histone H1: An antimicrobial protein of Atlantic salmon (Salmo salar). Biochem. Biophys. Res. Commun. 2001, 284, 549–555. [Google Scholar] [CrossRef]
- Chen, M.Z.; Chen, J.; Lu, X.J.; Shi, Y.H. Molecular cloning, sequence analysis and expression pattern of hepcidin gene in ayu (Plecoglossus altivelis). Dongwuxue Yanjiu 2010, 31, 595–600. [Google Scholar]
- Lu, X.J.; Chen, J.; Huang, Z.A.; Shi, Y.H.; Lv, J.N. Identification and characterization of a novel cathelicidin from ayu, Plecoglossus altivelis. Fish Shellfish Immunol. 2011, 31, 52–57. [Google Scholar] [CrossRef]
- Barnes, A.C.; Trewin, B.; Snape, N.; Kvennefors, E.C.; Baiano, J.C. Two hepcidin-like antimicrobial peptides in Barramundi Lates calcarifer exhibit differing tissue tropism and are induced in response to lipopolysaccharide. Fish Shellfish Immunol. 2011, 31, 350–357. [Google Scholar] [CrossRef]
- Yang, M.; Wang, K.J.; Chen, J.H.; Qu, H.D.; Li, S.J. Genomic organization and tissue-specific expression analysis of hepcidin-like genes from black porgy (Acanthopagrus schlegelii B). Fish Shellfish Immunol. 2007, 23, 1060–1071. [Google Scholar] [CrossRef]
- Yang, M.; Chen, B.; Cai, J.J.; Peng, H.; Ling, C.; Yuan, J.J.; Wang, K.J. Molecular characterization of hepcidin AS-hepc2 and AS-hepc6 in black porgy (Acanthopagrus schlegelii): Expression pattern responded to bacterial challenge and in vitro antimicrobial activity. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 155–163. [Google Scholar] [CrossRef]
- Kim, Y.O.; Park, E.M.; Nam, B.H.; Kong, H.J.; Kim, W.J.; Lee, S.J. Identification and molecular characterization of two hepcidin genes from black rockfish (Sebastes schlegelii). Mol. Cell. Biochem. 2008, 315, 131–136. [Google Scholar] [CrossRef]
- Gong, L.C.; Wang, H.; Deng, L. Molecular characterization, phylogeny and expression of a hepcidin gene in the blotched snakehead Channa maculata. Dev. Comp. Immunol. 2014, 44, 1–11. [Google Scholar] [CrossRef]
- Bao, B.; Peatman, E.; Li, P.; He, C.; Liu, Z. Catfish hepcidin gene is expressed in a wide range of tissues and exhibits tissue-specific upregulation after bacterial infection. Dev. Comp. Immunol. 2005, 29, 939–950. [Google Scholar] [CrossRef]
- Liang, T.; Ji, W.; Zhang, G.R.; Wei, K.J.; Feng, K.; Wang, W.M.; Zou, G.W. Molecular cloning and expression analysis of liver-expressed antimicrobial peptide 1 (LEAP-1) and LEAP-2 genes in the blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2013, 35, 553–563. [Google Scholar] [CrossRef]
- Scocchi, M.; Pallavicini, A.; Salgaro, R.; Bociek, K.; Gennaro, R. The salmonid cathelicidins: A gene family with highly varied C-terminal antimicrobial domains. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 376–381. [Google Scholar] [CrossRef]
- Park, I.Y.; Park, C.B.; Kim, M.S.; Kim, S.C. Parasin I, an antimicrobial peptide derived from histone H2A in the catfish, Parasilurus asotus. FEBS Lett. 1998, 437, 258–262. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, H.; Zhang, X.; Luo, H.; Xue, X.; Li, Z.; Yao, B. Identification, expression and bioactivity of Paramisgurnus dabryanus beta-defensin that might be involved in immune defense against bacterial infection. Fish Shellfish Immunol. 2013, 35, 399–406. [Google Scholar] [CrossRef]
- Marel, M.; Adamek, M.; Gonzalez, S.F.; Frost, P.; Rombout, J.H.; Wiegertjes, G.F.; Savelkoul, H.F.; Steinhagen, D. Molecular cloning and expression of two beta-defensin and two mucin genes in common carp (Cyprinus carpio L.) and their up-regulation after beta-glucan feeding. Fish Shellfish Immunol. 2012, 32, 494–501. [Google Scholar] [CrossRef]
- Li, H.; Zhang, F.; Guo, H.; Zhu, Y.; Yuan, J.; Yang, G.; An, L. Molecular characterization of hepcidin gene in common carp (Cyprinus carpio L.) and its expression pattern responding to bacterial challenge. Fish Shellfish Immunol. 2013, 35, 1030–1038. [Google Scholar] [CrossRef]
- Salerno, G.; Parrinello, N.; Roch, P.; Cammarata, M. cDNA sequence and tissue expression of an antimicrobial peptide, dicentracin; a new component of the moronecidin family isolated from head kidney leukocytes of sea bass, Dicentrarchus labrax. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 521–529. [Google Scholar] [CrossRef]
- Rodrigues, P.N.; Vazquez-Dorado, S.; Neves, J.V.; Wilson, J.M. Dual function of fish hepcidin: Response to experimental iron overload and bacterial infection in sea bass (Dicentrarchus labrax). Dev. Comp. Immunol. 2006, 30, 1156–1167. [Google Scholar] [CrossRef]
- Cuesta, A.; Meseguer, J.; Esteban, M.A. Molecular and functional characterization of the gilthead seabream beta-defensin demonstrate its chemotactic and antimicrobial activity. Mol. Immunol. 2011, 48, 1432–1438. [Google Scholar] [CrossRef]
- Cuesta, A.; Meseguer, J.; Esteban, M.A. The antimicrobial peptide hepcidin exerts an important role in the innate immunity against bacteria in the bony fish gilthead seabream. Mol. Immunol. 2008, 45, 2333–2342. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Ma, L.; Yu, Y.; Yu, H.; Mohammed, S.; Chu, G.; Mu, L.; Zhang, Q. Identification and characterization of a hepcidin from half-smooth tongue sole Cynoglossus semilaevis. Fish Shellfish Immunol. 2012, 33, 213–219. [Google Scholar] [CrossRef]
- Birkemo, G.A.; Luders, T.; Andersen, O.; Nes, I.F.; Nissen-Meyer, J. Hipposin, a histone-derived antimicrobial peptide in Atlantic halibut (Hippoglossus hippoglossus L.). Biochim. Biophys. Acta 2003, 1646, 207–215. [Google Scholar] [CrossRef]
- Salger, S.A.; Reading, B.J.; Baltzegar, D.A.; Sullivan, C.V.; Noga, E.J. Molecular characterization of two isoforms of piscidin 4 from the hybrid striped bass (Morone chrysops x Morone saxatilis). Fish Shellfish Immunol. 2011, 30, 420–424. [Google Scholar] [CrossRef]
- Noga, E.J.; Silphaduang, U.; Park, N.G.; Seo, J.K.; Stephenson, J.; Kozlowicz, S. Piscidin 4, a novel member of the piscidin family of antimicrobial peptides. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 299–305. [Google Scholar] [CrossRef]
- Lauth, X.; Shike, H.; Burns, J.C.; Westerman, M.E.; Ostland, V.E.; Carlberg, J.M.; van Olst, J.C.; Nizet, V.; Taylor, S.W.; Shimizu, C.; et al. Discovery and characterization of two isoforms of moronecidin, a novel antimicrobial peptide from hybrid striped bass. J. Biol. Chem. 2002, 277, 5030–5039. [Google Scholar] [CrossRef]
- Shike, H.; Lauth, X.; Westerman, M.E.; Ostland, V.E.; Carlberg, J.M.; van Olst, J.C.; Shimizu, C.; Bulet, P.; Burns, J.C. Bass hepcidin is a novel antimicrobial peptide induced by bacterial challenge. Eur. J. Biochem. 2002, 269, 2232–2237. [Google Scholar] [CrossRef]
- Buonocore, F.; Randelli, E.; Casani, D.; Picchietti, S.; Belardinelli, M.C.; de Pascale, D.; de Santi, C.; Scapigliati, G. A piscidin-like antimicrobial peptide from the icefish Chionodraco hamatus (Perciformes: Channichthyidae): Molecular characterization, localization and bactericidal activity. Fish Shellfish Immunol. 2012, 33, 1183–1191. [Google Scholar] [CrossRef]
- Zou, J.; Mercier, C.; Koussounadis, A.; Secombes, C. Discovery of multiple beta-defensin like homologues in teleost fish. Mol. Immunol. 2007, 44, 638–647. [Google Scholar] [CrossRef]
- Ren, H.L.; Wang, K.J.; Zhou, H.L.; Yang, M. Cloning and organisation analysis of a hepcidin-like gene and cDNA from Japan sea bass, Lateolabrax japonicus. Fish Shellfish Immunol. 2006, 21, 221–227. [Google Scholar] [CrossRef]
- Robertson, L.S.; Iwanowicz, L.R.; Marranca, J.M. Identification of centrarchid hepcidins and evidence that 17beta-estradiol disrupts constitutive expression of hepcidin-1 and inducible expression of hepcidin-2 in largemouth bass (Micropterus salmoides). Fish Shellfish Immunol. 2009, 26, 898–907. [Google Scholar] [CrossRef]
- Niu, S.F.; Jin, Y.; Xu, X.; Qiao, Y.; Wu, Y.; Mao, Y.; Su, Y.Q.; Wang, J. Characterization of a novel piscidin-like antimicrobial peptide from Pseudosciaena crocea and its immune response to Cryptocaryon irritans. Fish Shellfish Immunol. 2013, 35, 513–524. [Google Scholar] [CrossRef]
- Wang, K.J.; Cai, J.J.; Cai, L.; Qu, H.D.; Yang, M.; Zhang, M. Cloning and expression of a hepcidin gene from a marine fish (Pseudosciaena crocea) and the antimicrobial activity of its synthetic peptide. Peptides 2009, 30, 638–646. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, Q.; Ji, R.; Zou, W.; Guo, G. Isolation and characterization of a hepcidin peptide from the head kidney of large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol. 2009, 26, 864–870. [Google Scholar] [CrossRef]
- Sun, B.J.; Xie, H.X.; Song, Y.; Nie, P. Gene structure of an antimicrobial peptide from mandarin fish, Siniperca chuatsi (Basilewsky), suggests that moronecidins and pleurocidins belong in one family: The piscidins. J. Fish Dis. 2007, 30, 335–343. [Google Scholar] [CrossRef]
- Wang, G.; Li, J.; Zou, P.; Xie, H.; Huang, B.; Nie, P.; Chang, M. Expression pattern, promoter activity and bactericidal property of beta-defensin from the mandarin fish Siniperca chuatsi. Fish Shellfish Immunol. 2012, 33, 522–531. [Google Scholar] [CrossRef]
- Zhao, J.G.; Zhou, L.; Jin, J.Y.; Zhao, Z.; Lan, J.; Zhang, Y.B.; Zhang, Q.Y.; Gui, J.F. Antimicrobial activity-specific to Gram-negative bacteria and immune modulation-mediated NF-kappaB and Sp1 of a medaka beta-defensin. Dev. Comp. Immunol. 2009, 33, 624–637. [Google Scholar]
- Bo, J.; Cai, L.; Xu, J.H.; Wang, K.J.; Au, D.W. The marine medaka Oryzias melastigma—A potential marine fish model for innate immune study. Mar. Pollut. Bull. 2011, 63, 267–276. [Google Scholar] [CrossRef]
- Xu, T.; Sun, Y.; Shi, G.; Wang, R. Miiuy croaker hepcidin gene and comparative analyses reveal evidence for positive selection. PLoS One 2012, 7, e35449. [Google Scholar]
- Brocal, I.; Falco, A.; Mas, V.; Rocha, A.; Perez, L.; Coll, J.M.; Estepa, A. Stable expression of bioactive recombinant pleurocidin in a fish cell line. Appl. Microbiol. Biotechnol. 2006, 72, 1217–1228. [Google Scholar] [CrossRef]
- Nam, Y.K.; Cho, Y.S.; Lee, S.Y.; Kim, B.S.; Kim, D.S. Molecular characterization of hepcidin gene from mud loach (Misgurnus mizolepis; Cypriniformes). Fish Shellfish Immunol. 2011, 31, 1251–1258. [Google Scholar] [CrossRef]
- Nam, B.H.; Moon, J.Y.; Kim, Y.O.; Kong, H.J.; Kim, W.J.; Lee, S.J.; Kim, K.K. Multiple beta-defensin isoforms identified in early developmental stages of the teleost Paralichthys olivaceus. Fish Shellfish Immunol. 2010, 28, 267–274. [Google Scholar] [CrossRef]
- Pan, C.Y.; Chen, J.Y.; Cheng, Y.S.; Chen, C.Y.; Ni, I.H.; Sheen, J.F.; Pan, Y.L.; Kuo, C.M. Gene expression and localization of the epinecidin-1 antimicrobial peptide in the grouper (Epinephelus coioides), and its role in protecting fish against pathogenic infection. DNA Cell Biol. 2007, 26, 403–413. [Google Scholar] [CrossRef]
- Guo, M.; Wei, J.; Huang, X.; Huang, Y.; Qin, Q. Antiviral effects of beta-defensin derived from orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2012, 32, 828–838. [Google Scholar] [CrossRef]
- Jin, J.Y.; Zhou, L.; Wang, Y.; Li, Z.; Zhao, J.G.; Zhang, Q.Y.; Gui, J.F. Antibacterial and antiviral roles of a fish beta-defensin expressed both in pituitary and testis. PLoS One 2010, 5, e12883. [Google Scholar]
- Zhou, J.G.; Wei, J.G.; Xu, D.; Cui, H.C.; Yan, Y.; Ou-Yang, Z.L.; Huang, X.H.; Huang, Y.H.; Qin, Q.W. Molecular cloning and characterization of two novel hepcidins from orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2011, 30, 559–568. [Google Scholar] [CrossRef]
- Qu, H.; Chen, B.; Peng, H.; Wang, K. Molecular cloning, recombinant expression, and antimicrobial activity of EC-hepcidin3, a new four-cysteine hepcidin isoform from Epinephelus coioides. Biosci. Biotechnol. Biochem. 2013, 77, 103–110. [Google Scholar] [CrossRef]
- Masso-Silva, J.; Diamond, G.; Macias-Rodriguez, M.; Ascencio, F. Genomic organization and tissue-specific expression of hepcidin in the pacific mutton hamlet, Alphestes immaculatus (Breder, 1936). Fish Shellfish Immunol. 2011, 31, 1297–1302. [Google Scholar] [CrossRef]
- Falco, A.; Chico, V.; Marroqui, L.; Perez, L.; Coll, J.M.; Estepa, A. Expression and antiviral activity of a beta-defensin-like peptide identified in the rainbow trout (Oncorhynchus mykiss) EST sequences. Mol. Immunol. 2008, 45, 757–765. [Google Scholar]
- Casadei, E.; Wang, T.; Zou, J.; Gonzalez Vecino, J.L.; Wadsworth, S.; Secombes, C.J. Characterization of three novel beta-defensin antimicrobial peptides in rainbow trout (Oncorhynchus mykiss). Mol. Immunol. 2009, 46, 3358–3366. [Google Scholar] [CrossRef]
- Fernandes, J.M.; Kemp, G.D.; Molle, M.G.; Smith, V.J. Anti-microbial properties of histone H2A from skin secretions of rainbow trout, Oncorhynchus mykiss. Biochem. J. 2002, 368, 611–620. [Google Scholar] [CrossRef]
- Fernandes, J.M.; Molle, G.; Kemp, G.D.; Smith, V.J. Isolation and characterisation of oncorhyncin II, a histone H1-derived antimicrobial peptide from skin secretions of rainbow trout, Oncorhynchus mykiss. Dev. Comp. Immunol. 2004, 28, 127–138. [Google Scholar] [CrossRef]
- Fernandes, J.M.; Saint, N.; Kemp, G.D.; Smith, V.J. Oncorhyncin III: A potent antimicrobial peptide derived from the non-histone chromosomal protein H6 of rainbow trout, Oncorhynchus mykiss. Biochem. J. 2003, 373, 621–628. [Google Scholar] [CrossRef]
- Noga, E.J.; Fan, Z.; Silphaduang, U. Histone-like proteins from fish are lethal to the parasitic dinoflagellate Amyloodinium ocellatum. Parasitology 2001, 123, 57–65. [Google Scholar]
- Martin-Antonio, B.; Jimenez-Cantizano, R.M.; Salas-Leiton, E.; Infante, C.; Manchado, M. Genomic characterization and gene expression analysis of four hepcidin genes in the redbanded seabream (Pagrus auriga). Fish Shellfish Immunol. 2009, 26, 483–491. [Google Scholar] [CrossRef]
- Iijima, N.; Tanimoto, N.; Emoto, Y.; Morita, Y.; Uematsu, K.; Murakami, T.; Nakai, T. Purification and characterization of three isoforms of chrysophsin, a novel antimicrobial peptide in the gills of the red sea bream, Chrysophrys major. Eur. J. Biochem. 2003, 270, 675–686. [Google Scholar] [CrossRef]
- Chen, S.L.; Xu, M.Y.; Ji, X.S.; Yu, G.C.; Liu, Y. Cloning, characterization, and expression analysis of hepcidin gene from red sea bream (Chrysophrys major). Antimicrob. Agents Chemother. 2005, 49, 1608–1612. [Google Scholar] [CrossRef]
- Cho, Y.S.; Lee, S.Y.; Kim, K.H.; Kim, S.K.; Kim, D.S.; Nam, Y.K. Gene structure and differential modulation of multiple rockbream (Oplegnathus fasciatus) hepcidin isoforms resulting from different biological stimulations. Dev. Comp. Immunol. 2009, 33, 46–58. [Google Scholar] [CrossRef]
- Terova, G.; Cattaneo, A.G.; Preziosa, E.; Bernardini, G.; Saroglia, M. Impact of acute stress on antimicrobial polypeptides mRNA copy number in several tissues of marine sea bass (Dicentrarchus labrax). BMC Immunol. 2011, 12, 69. [Google Scholar] [CrossRef]
- Sun, D.; Wu, S.; Jing, C.; Zhang, N.; Liang, D.; Xu, A. Identification, synthesis and characterization of a novel antimicrobial peptide HKPLP derived from Hippocampus kuda Bleeker. J. Antibiot. (Tokyo) 2012, 65, 117–121. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Dhanik, M.; Pande, A. Characterization and structural analysis of hepcidin like antimicrobial peptide from schizothorax richardsonii (Gray). Protein J. 2014, 33, 1–10. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, S.; Gao, J.; Guang, H.; Tian, Y.; Zhao, Z.; Wang, Y.; Yu, H. Structural and functional characterization of CATH_BRALE, the defense molecule in the ancient salmonoid, Brachymystax lenok. Fish Shellfish Immunol. 2013, 34, 1–7. [Google Scholar] [CrossRef]
- Peng, K.C.; Lee, S.H.; Hour, A.-L.; Pan, C.Y.; Lee, L.H.; Chen, J. Five different piscidins from nile tilapia, oreochromis niloticus: Analysis of their expressions and biological functions. PLoS One 2012, 7, e50263. [Google Scholar]
- Huang, P.H.; Chen, J.Y.; Kuo, C.M. Three different hepcidins from tilapia, Oreochromis mossambicus: Analysis of their expressions and biological functions. Mol. Immunol. 2007, 44, 1922–1934. [Google Scholar] [CrossRef]
- Pereiro, P.; Figueras, A.; Novoa, B. A novel hepcidin-like in turbot (Scophthalmus maximus L.) highly expressed after pathogen challenge but not after iron overload. Fish Shellfish Immunol. 2012, 32, 879–889. [Google Scholar] [CrossRef]
- Chen, S.L.; Li, W.; Meng, L.; Sha, Z.X.; Wang, Z.J.; Ren, G.C. Molecular cloning and expression analysis of a hepcidin antimicrobial peptide gene from turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2007, 22, 172–181. [Google Scholar] [CrossRef]
- Cole, A.M.; Darouiche, R.O.; Legarda, D.; Connell, N.; Diamond, G. Characterization of a fish antimicrobial peptide: Gene expression, subcellular localization, and spectrum of activity. Antimicrob. Agents Chemother. 2000, 44, 2039–2045. [Google Scholar] [CrossRef]
- Douglas, S.E.; Patrzykat, A.; Pytyck, J.; Gallant, J.W. Identification, structure and differential expression of novel pleurocidins clustered on the genome of the winter flounder, Pseudopleuronectes americanus (Walbaum). Eur. J. Biochem. 2003, 270, 3720–3730. [Google Scholar] [CrossRef]
- Shike, H.; Shimizu, C.; Lauth, X.; Burns, J.C. Organization and expression analysis of the zebrafish hepcidin gene, an antimicrobial peptide gene conserved among vertebrates. Dev. Comp. Immunol. 2004, 28, 747–754. [Google Scholar] [CrossRef]
- Tamang, D.G.; Saier, M.H., Jr. The cecropin superfamily of toxic peptides. J. Mol. Microbiol. Biotechnol. 2006, 11, 94–103. [Google Scholar] [CrossRef]
- Park, C.B.; Lee, J.H.; Park, I.Y.; Kim, M.S.; Kim, S.C. A novel antimicrobial peptide from the loach, Misgurnus anguillicaudatus. FEBS Lett. 1997, 411, 173–178. [Google Scholar] [CrossRef]
- Silphaduang, U.; Noga, E.J. Peptide antibiotics in mast cells of fish. Nature 2001, 414, 268–269. [Google Scholar] [CrossRef]
- Silphaduang, U.; Colorni, A.; Noga, E.J. Evidence for widespread distribution of piscidin antimicrobial peptides in teleost fish. Dis. Aquat. Org. 2006, 72, 241–252. [Google Scholar] [CrossRef]
- Pan, C.Y.; Chen, J.Y.; Ni, I.H.; Wu, J.L.; Kuo, C.M. Organization and promoter analysis of the grouper (Epinephelus coioides) epinecidin-1 gene. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 150, 358–367. [Google Scholar] [CrossRef]
- Syvitski, R.T.; Burton, I.; Mattatall, N.R.; Douglas, S.E.; Jakeman, D.L. Structural characterization of the antimicrobial peptide pleurocidin from winter flounder. Biochemistry 2005, 44, 7282–7293. [Google Scholar] [CrossRef]
- Fernandes, J.M.O.; Ruangsri, J.; Kiron, V. Atlantic cod piscidin and its diversification through positive selection. PLoS One 2010, 5, e9501. [Google Scholar] [CrossRef]
- Tennessen, J.A. Enhanced synonymous site divergence in positively selected vertebrate antimicrobial peptide genes. J. Mol. Evol. 2005, 61, 445–455. [Google Scholar] [CrossRef]
- Pinzon-Arango, P.A.; Nagarajan, R.; Camesano, T.A. Interactions of antimicrobial peptide chrysophsin-3 with Bacillus anthracis in sporulated, germinated, and vegetative states. J. Phys. Chem. B 2013, 117, 6364–6372. [Google Scholar] [CrossRef]
- Cho, J.; Lee, D.G. Oxidative stress by antimicrobial peptide pleurocidin triggers apoptosis in Candida albicans. Biochimie 2011, 93, 1873–1879. [Google Scholar] [CrossRef]
- Jung, H.J.; Park, Y.; Sung, W.S.; Suh, B.K.; Lee, J.; Hahm, K.S.; Lee, D.G. Fungicidal effect of pleurocidin by membrane-active mechanism and design of enantiomeric analogue for proteolytic resistance. Biochim. Biophys. Acta 2007, 1768, 1400–1405. [Google Scholar] [CrossRef]
- Pan, C.Y.; Chen, J.Y.; Lin, T.L.; Lin, C.H. In vitro activities of three synthetic peptides derived from epinecidin-1 and an anti-lipopolysaccharide factor against Propionibacterium acnes, Candida albicans, and Trichomonas vaginalis. Peptides 2009, 30, 1058–1068. [Google Scholar] [CrossRef]
- Colorni, A.; Ullal, A.; Heinisch, G.; Noga, E.J. Activity of the antimicrobial polypeptide piscidin 2 against fish ectoparasites. J. Fish Dis. 2008, 31, 423–432. [Google Scholar] [CrossRef]
- Zahran, E.; Noga, E.J. Evidence for synergism of the antimicrobial peptide piscidin 2 with antiparasitic and antioomycete drugs. J. Fish Dis. 2010, 33, 995–1003. [Google Scholar] [CrossRef]
- Wang, Y.D.; Kung, C.W.; Chen, J.Y. Antiviral activity by fish antimicrobial peptides of epinecidin-1 and hepcidin 1–5 against nervous necrosis virus in medaka. Peptides 2010, 31, 1026–1033. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Bryan, L.; Silphadaung, U.; Noga, E.; Wade, D.; Rollins-Smith, L. Inactivation of viruses infecting ectothermic animals by amphibian and piscine antimicrobial peptides. Virology 2004, 323, 268–275. [Google Scholar] [CrossRef]
- Douglas, S.E.; Gallant, J.W.; Gong, Z.; Hew, C. Cloning and developmental expression of a family of pleurocidin-like antimicrobial peptides from winter flounder, Pleuronectes americanus (Walbaum). Dev. Comp. Immunol. 2001, 25, 137–147. [Google Scholar] [CrossRef]
- Mulero, I.; Noga, E.J.; Meseguer, J.; Garcia-Ayala, A.; Mulero, V. The antimicrobial peptides piscidins are stored in the granules of professional phagocytic granulocytes of fish and are delivered to the bacteria-containing phagosome upon phagocytosis. Dev. Comp. Immunol. 2008, 32, 1531–1538. [Google Scholar] [CrossRef]
- Ruangsri, J.; Fernandes, J.M.; Rombout, J.H.; Brinchmann, M.F.; Kiron, V. Ubiquitous presence of piscidin-1 in Atlantic cod as evidenced by immunolocalisation. BMC Vet. Res. 2012, 8, 46. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Pironi, F.; Giari, L.; Noga, E.J. Immunocytochemical localization of piscidin in mast cells of infected seabass gill. Fish Shellfish Immunol. 2010, 28, 476–482. [Google Scholar] [CrossRef]
- Murray, H.M.; Gallant, J.W.; Douglas, S.E. Cellular localization of pleurocidin gene expression and synthesis in winter flounder gill using immunohistochemistry and in situ hybridization. Cell Tissue Res. 2003, 312, 197–202. [Google Scholar]
- Dezfuli, B.S.; Castaldelli, G.; Bo, T.; Lorenzoni, M.; Giari, L. Intestinal immune response of Silurus glanis and Barbus barbus naturally infected with Pomphorhynchus laevis (Acanthocephala). Parasite Immunol. 2011, 33, 116–23. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Lui, A.; Giari, L.; Castaldelli, G.; Mulero, V.; Noga, E.J. Infiltration and activation of acidophilic granulocytes in skin lesions of gilthead seabream, Sparus aurata, naturally infected with lymphocystis disease virus. Dev. Comp. Immunol. 2012, 36, 174–182. [Google Scholar] [CrossRef]
- Lin, W.J.; Chien, Y.L.; Pan, C.Y.; Lin, T.L.; Chen, J.Y.; Chiu, S.J.; Hui, C.F. Epinecidin-1, an antimicrobial peptide from fish (Epinephelus coioides) which has an antitumor effect like lytic peptides in human fibrosarcoma cells. Peptides 2009, 30, 283–290. [Google Scholar] [CrossRef]
- Lin, H.J.; Huang, T.C.; Muthusamy, S.; Lee, J.F.; Duann, Y.F.; Lin, C.H. Piscidin-1, an antimicrobial peptide from fish (hybrid striped bass morone saxatilis x M. chrysops), induces apoptotic and necrotic activity in HT1080 cells. Zool. Sci. 2012, 29, 327–332. [Google Scholar] [CrossRef]
- Hsu, J.C.; Lin, L.C.; Tzen, J.T.; Chen, J.Y. Characteristics of the antitumor activities in tumor cells and modulation of the inflammatory response in RAW264.7 cells of a novel antimicrobial peptide, chrysophsin-1, from the red sea bream (Chrysophrys major). Peptides 2011, 32, 900–910. [Google Scholar] [CrossRef]
- Chen, J.Y.; Lin, W.J.; Wu, J.L.; Her, G.M.; Hui, C.F. Epinecidin-1 peptide induces apoptosis which enhances antitumor effects in human leukemia U937 cells. Peptides 2009, 30, 2365–2373. [Google Scholar] [CrossRef]
- Morash, M.G.; Douglas, S.E.; Robotham, A.; Ridley, C.M.; Gallant, J.W.; Soanes, K.H. The zebrafish embryo as a tool for screening and characterizing pleurocidin host-defense peptides as anti-cancer agents. Dis. Model. Mech. 2011, 4, 622–633. [Google Scholar] [CrossRef]
- Hilchie, A.L.; Doucette, C.D.; Pinto, D.M.; Patrzykat, A.; Douglas, S.; Hoskin, D.W. Pleurocidin-family cationic antimicrobial peptides are cytolytic for breast carcinoma cells and prevent growth of tumor xenografts. Breast Cancer Res. 2011, 13, R102. [Google Scholar] [CrossRef]
- Pan, C.Y.; Chow, T.Y.; Yu, C.Y.; Yu, C.Y.; Chen, J.C.; Chen, J.Y. Antimicrobial peptides of an anti-lipopolysaccharide factor, epinecidin-1, and hepcidin reduce the lethality of Riemerella anatipestifer sepsis in ducks. Peptides 2010, 31, 806–815. [Google Scholar] [CrossRef]
- Sung, W.S.; Lee, D.G. Pleurocidin-derived antifungal peptides with selective membrane-disruption effect. Biochem. Biophys. Res. Commun. 2008, 369, 858–861. [Google Scholar] [CrossRef]
- Rahmanpour, A.; Ghahremanpour, M.M.; Mehrnejad, F.; Moghaddam, M.E. Interaction of Piscidin-1 with zwitterionic versus anionic membranes: A comparative molecular dynamics study. J. Biomol. Struct. Dyn. 2013, 31, 1393–1403. [Google Scholar] [CrossRef]
- Chekmenev, E.Y.; Jones, S.M.; Nikolayeva, Y.N.; Vollmar, B.S.; Wagner, T.J.; Gor’kov, P.L.; Brey, W.W.; Manion, M.N.; Daugherty, K.C.; Cotten, M. High-field NMR studies of molecular recognition and structure-function relationships in antimicrobial piscidins at the water-lipid bilayer interface. J. Am. Chem. Soc. 2006, 128, 5308–5309. [Google Scholar] [CrossRef]
- Mason, A.J.; Bertani, P.; Moulay, G.; Marquette, A.; Perrone, B.; Drake, A.F.; Kichler, A.; Bechinger, B. Membrane interaction of chrysophsin-1, a histidine-rich antimicrobial peptide from red sea bream. Biochemistry 2007, 46, 15175–15187. [Google Scholar] [CrossRef]
- Patrzykat, A.; Friedrich, C.L.; Zhang, L.; Mendoza, V.; Hancock, R.E. Sublethal concentrations of pleurocidin-derived antimicrobial peptides inhibit macromolecular synthesis in Escherichia coli. Antimicrob. Agents Chemother. 2002, 46, 605–614. [Google Scholar] [CrossRef]
- Kim, J.K.; Lee, S.A.; Shin, S.; Lee, J.Y.; Jeong, K.W.; Nan, Y.H.; Park, Y.S.; Shin, S.Y.; Kim, Y. Structural flexibility and the positive charges are the key factors in bacterial cell selectivity and membrane penetration of peptoid-substituted analog of Piscidin 1. Biochim. Biophys. Acta 2010, 1798, 1913–1925. [Google Scholar]
- Lin, S.B.; Fan, T.W.; Wu, J.L.; Hui, C.F.; Chen, J.Y. Immune response and inhibition of bacterial growth by electrotransfer of plasmid DNA containing the antimicrobial peptide, epinecidin-1, into zebrafish muscle. Fish Shellfish Immunol. 2009, 26, 451–458. [Google Scholar] [CrossRef]
- Lee, L.H.; Hui, C.F.; Chuang, C.M.; Chen, J.Y. Electrotransfer of the epinecidin-1 gene into skeletal muscle enhances the antibacterial and immunomodulatory functions of a marine fish, grouper (Epinephelus coioides). Fish Shellfish Immunol. 2013, 35, 1359–1368. [Google Scholar] [CrossRef]
- Pan, C.Y.; Wu, J.L.; Hui, C.F.; Lin, C.H.; Chen, J.Y. Insights into the antibacterial and immunomodulatory functions of the antimicrobial peptide, epinecidin-1, against Vibrio vulnificus infection in zebrafish. Fish Shellfish Immunol. 2011, 31, 1019–1025. [Google Scholar] [CrossRef]
- Pan, C.Y.; Huang, T.C.; Wang, Y.D.; Yeh, Y.C.; Hui, C.F.; Chen, J.Y. Oral administration of recombinant epinecidin-1 protected grouper (Epinephelus coioides) and zebrafish (Danio rerio) from Vibrio vulnificus infection and enhanced immune-related gene expressions. Fish Shellfish Immunol. 2012, 32, 947–957. [Google Scholar] [CrossRef]
- Peng, K.C.; Pan, C.Y.; Chou, H.N.; Chen, J.Y. Using an improved Tol2 transposon system to produce transgenic zebrafish with epinecidin-1 which enhanced resistance to bacterial infection. Fish Shellfish Immunol. 2010, 28, 905–917. [Google Scholar] [CrossRef]
- Huang, H.N.; Pan, C.Y.; Rajanbabu, V.; Chan, Y.L.; Wu, C.J.; Chen, J.Y. Modulation of immune responses by the antimicrobial peptide, epinecidin (Epi)-1, and establishment of an Epi-1-based inactivated vaccine. Biomaterials 2011, 32, 3627–3636. [Google Scholar] [CrossRef]
- Lee, S.C.; Pan, C.Y.; Chen, J.Y. The antimicrobial peptide, epinecidin-1, mediates secretion of cytokines in the immune response to bacterial infection in mice. Peptides 2012, 36, 100–108. [Google Scholar] [CrossRef]
- Huang, H.N.; Rajanbabu, V.; Pan, C.Y.; Chan, Y.L.; Wu, C.J.; Chen, J.Y. Use of the antimicrobial peptide Epinecidin-1 to protect against MRSA infection in mice with skin injuries. Biomaterials 2013, 34, 10319–10327. [Google Scholar] [CrossRef]
- Pundir, P.; Catalli, A.; Leggiadro, C.; Douglas, S.E.; Kulka, M. Pleurocidin, a novel antimicrobial peptide, induces human mast cell activation through the FPRL1 receptor. Mucosal Immunol. 2014, 7, 177–187. [Google Scholar] [CrossRef]
- Bulet, P.; Stocklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef]
- Aerts, A.M.; Francois, I.E.; Cammue, B.P.; Thevissen, K. The mode of antifungal action of plant, insect and human defensins. Cell. Mol. Life Sci. 2008, 65, 2069–2079. [Google Scholar] [CrossRef]
- Zhu, S. Discovery of six families of fungal defensin-like peptides provides insights into origin and evolution of the CSalphabeta defensins. Mol. Immunol. 2008, 45, 828–838. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, B. Evolutionary origin of beta-defensins. Dev. Comp. Immunol. 2013, 39, 79–84. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, C.; Heng, H.H.; Ganz, T. The human beta-defensin-1 and alpha-defensins are encoded by adjacent genes: Two peptide families with differing disulfide topology share a common ancestry. Genomics 1997, 43, 316–320. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Yuan, J.; Osapay, G.; Osapay, K.; Tran, D.; Miller, C.J.; Ouellette, A.J.; Selsted, M.E. A cyclic antimicrobial peptide produced in primate leukocytes by the ligation of two truncated alpha-defensins. Science 1999, 286, 498–502. [Google Scholar] [CrossRef]
- Beckloff, N.; Diamond, G. Computational analysis suggests beta-defensins are processed to mature peptides by signal peptidase. Protein Pept. Lett. 2008, 15, 536–540. [Google Scholar] [CrossRef]
- Falco, A.; Mas, V.; Tafalla, C.; Perez, L.; Coll, J.M.; Estepa, A. Dual antiviral activity of human alpha-defensin-1 against viral haemorrhagic septicaemia rhabdovirus (VHSV): Inactivation of virus particles and induction of a type I interferon-related response. Antivir. Res. 2007, 76, 111–123. [Google Scholar] [CrossRef]
- Madison, M.N.; Kleshchenko, Y.Y.; Nde, P.N.; Simmons, K.J.; Lima, M.F.; Villalta, F. Human defensin alpha-1 causes Trypanosoma cruzi membrane pore formation and induces DNA fragmentation, which leads to trypanosome destruction. Infect. Immun. 2007, 75, 4780–4791. [Google Scholar] [CrossRef]
- Tanaka, T.; Rahman, M.M.; Battur, B.; Boldbaatar, D.; Liao, M.; Umemiya-Shirafuji, R.; Xuan, X.; Fujisaki, K. Parasiticidal activity of human alpha-defensin-5 against Toxoplasma gondii. In Vitro Cell. Dev. Biol. Anim. 2010, 46, 560–565. [Google Scholar] [CrossRef]
- Krishnakumari, V.; Rangaraj, N.; Nagaraj, R. Antifungal activities of human beta-defensins HBD-1 to HBD-3 and their C-terminal analogs Phd1 to Phd3. Antimicrob. Agents Chemother. 2009, 53, 256–260. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, Y.; Wang, B.; Yang, D.; Yu, K.; Yang, X.; Liu, F.; Jiang, Z.; Li, M. Antifungal activity of recombinant mouse beta-defensin 3. Lett. Appl. Microbiol. 2010, 50, 468–473. [Google Scholar] [CrossRef]
- Aerts, A.M.; Thevissen, K.; Bresseleers, S.M.; Sels, J.; Wouters, P.; Cammue, B.P.; Francois, I.E. Arabidopsis thaliana plants expressing human beta-defensin-2 are more resistant to fungal attack: Functional homology between plant and human defensins. Plant Cell Rep. 2007, 26, 1391–1398. [Google Scholar] [CrossRef]
- Semple, F.; Dorin, J.R. Beta-Defensins: Multifunctional modulators of infection, inflammation and more? J. Innate Immun. 2012, 4, 337–348. [Google Scholar] [CrossRef]
- Rohrl, J.; Yang, D.; Oppenheim, J.J.; Hehlgans, T. Specific binding and chemotactic activity of mBD4 and its functional orthologue hBD2 to CCR6-expressing cells. J. Biol. Chem. 2010, 285, 7028–7034. [Google Scholar]
- Liu, Y.; Chang, M.X.; Wu, S.G.; Nie, P. Characterization of C-C chemokine receptor subfamily in teleost fish. Mol. Immunol. 2009, 46, 498–504. [Google Scholar]
- Dixon, B.; Luque, A.; Abos, B.; Castro, R.; Gonzalez-Torres, L.; Tafalla, C. Molecular characterization of three novel chemokine receptors in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2013, 34, 641–651. [Google Scholar]
- Falco, A.; Brocal, I.; Perez, L.; Coll, J.M.; Estepa, A.; Tafalla, C. In vivo modulation of the rainbow trout (Oncorhynchus mykiss) immune response by the human alpha defensin 1, HNP1. Fish Shellfish Immunol. 2008, 24, 102–112. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Ganz, T. Defensins of vertebrate animals. Curr. Opin. Immunol. 2002, 14, 96–102. [Google Scholar] [CrossRef]
- Tollner, T.L.; Venners, S.A.; Hollox, E.J.; Yudin, A.I.; Liu, X.; Tang, G.; Xing, H.; Kays, R.J.; Lau, T.; Overstreet, J.W.; et al. A common mutation in the defensin DEFB126 causes impaired sperm function and subfertility. Sci. Transl. Med. 2011, 3, 92–ra65. [Google Scholar]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef]
- Dorschner, R.A.; Lin, K.H.; Murakami, M.; Gallo, R.L. Neonatal skin in mice and humans expresses increased levels of antimicrobial peptides: Innate immunity during development of the adaptive response. Pediatr. Res. 2003, 53, 566–572. [Google Scholar] [CrossRef]
- Huttner, K.M.; Brezinski-Caliguri, D.J.; Mahoney, M.M.; Diamond, G. Antimicrobial peptide expression is developmentally-regulated in the ovine gastrointestinal tract. J. Nutr. 1998, 128, 297S–299S. [Google Scholar]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Casadei, E.; Bird, S.; Vecino, J.L.; Wadsworth, S.; Secombes, C.J. The effect of peptidoglycan enriched diets on antimicrobial peptide gene expression in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2013, 34, 529–537. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Guardiola, F.; Rojas, M.; Ascencio-Valle, F.; Esteban, M.A. Dietary administration of microalgae Navicula sp. affects immune status and gene expression of gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2013, 35, 883–889. [Google Scholar] [CrossRef]
- Krause, A.; Neitz, S.; Magert, H.J.; Schulz, A.; Forssmann, W.G.; Schulz-Knappe, P.; Adermann, K. LEAP-1, a novel highly disulfide-bonded human peptide, exhibits antimicrobial activity. FEBS Lett. 2000, 480, 147–150. [Google Scholar] [CrossRef]
- Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J. Biol. Chem. 2001, 276, 7806–7810. [Google Scholar] [CrossRef]
- Hilton, K.B.; Lambert, L.A. Molecular evolution and characterization of hepcidin gene products in vertebrates. Gene 2008, 415, 40–48. [Google Scholar] [CrossRef]
- Hunter, H.N.; Fulton, D.B.; Ganz, T.; Vogel, H.J. The solution structure of human hepcidin, a peptide hormone with antimicrobial activity that is involved in iron uptake and hereditary hemochromatosis. J. Biol. Chem. 2002, 277, 37597–37603. [Google Scholar]
- Padhi, A.; Verghese, B. Evidence for positive Darwinian selection on the hepcidin gene of Perciform and Pleuronectiform fishes. Mol. Divers. 2007, 11, 119–130. [Google Scholar] [CrossRef]
- Lauth, X.; Babon, J.J.; Stannard, J.A.; Singh, S.; Nizet, V.; Carlberg, J.M.; Ostland, V.E.; Pennington, M.W.; Norton, R.S.; Westerman, M.E. Bass hepcidin synthesis, solution structure, antimicrobial activities and synergism, and in vivo hepatic response to bacterial infections. J. Biol. Chem. 2005, 280, 9272–9282. [Google Scholar]
- Hu, X.; Camus, A.C.; Aono, S.; Morrison, E.E.; Dennis, J.; Nusbaum, K.E.; Judd, R.L.; Shi, J. Channel catfish hepcidin expression in infection and anemia. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 55–69. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Mu, X.; Klesius, P.H. Expression profiles of seven channel catfish antimicrobial peptides in response to Edwardsiella ictaluri infection. J. Fish Dis. 2012, 35, 227–237. [Google Scholar] [CrossRef]
- Chiou, P.P.; Lin, C.M.; Bols, N.C.; Chen, T.T. Characterization of virus/double-stranded RNA-dependent induction of antimicrobial peptide hepcidin in trout macrophages. Dev. Comp. Immunol. 2007, 31, 1297–1309. [Google Scholar] [CrossRef]
- Yang, C.G.; Liu, S.S.; Sun, B.; Wang, X.L.; Wang, N.; Chen, S.L. Iron-metabolic function and potential antibacterial role of Hepcidin and its correlated genes (Ferroportin 1 and Transferrin Receptor) in turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2013, 34, 744–755. [Google Scholar] [CrossRef]
- Alvarez, C.A.; Santana, P.A.; Guzman, F.; Marshall, S.; Mercado, L. Detection of the hepcidin prepropeptide and mature peptide in liver of rainbow trout. Dev. Comp. Immunol. 2013, 41, 77–81. [Google Scholar] [CrossRef]
- Nemeth, E.; Valore, E.V.; Territo, M.; Schiller, G.; Lichtenstein, A.; Ganz, T. Hepcidin, a putative mediator of anemia of inflammation, is a type II acute-phase protein. Blood 2003, 101, 2461–2463. [Google Scholar] [CrossRef]
- Skov, J.; Kania, P.W.; Holten-Andersen, L.; Fouz, B.; Buchmann, K. Immunomodulatory effects of dietary beta-1,3-glucan from Euglena gracilis in rainbow trout (Oncorhynchus mykiss) immersion vaccinated against Yersinia ruckeri. Fish Shellfish Immunol. 2012, 33, 111–120. [Google Scholar] [CrossRef]
- Chia, T.J.; Wu, Y.C.; Chen, J.Y.; Chi, S.C. Antimicrobial peptides (AMP) with antiviral activity against fish nodavirus. Fish Shellfish Immunol. 2010, 28, 434–439. [Google Scholar] [CrossRef]
- Rajanbabu, V.; Chen, J.Y. Antiviral function of tilapia hepcidin 1–5 and its modulation of immune-related gene expressions against infectious pancreatic necrosis virus (IPNV) in Chinook salmon embryo (CHSE)-214 cells. Fish Shellfish Immunol. 2011, 30, 39–44. [Google Scholar] [CrossRef]
- Cai, L.; Cai, J.J.; Liu, H.P.; Fan, D.Q.; Peng, H.; Wang, K.J. Recombinant medaka (Oryzias melastigmus) pro-hepcidin: Multifunctional characterization. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2012, 161, 140–147. [Google Scholar] [CrossRef]
- Liu, H.; Trinh, T.L.; Dong, H.; Keith, R.; Nelson, D.; Liu, C. Iron regulator hepcidin exhibits antiviral activity against hepatitis C virus. PLoS One 2012, 7, e46631. [Google Scholar]
- Hocquellet, A.; le Senechal, C.; Garbay, B. Importance of the disulfide bridges in the antibacterial activity of human hepcidin. Peptides 2012, 36, 303–307. [Google Scholar] [CrossRef]
- Chen, J.Y.; Lin, W.J.; Lin, T.L. A fish antimicrobial peptide, tilapia hepcidin TH2–3, shows potent antitumor activity against human fibrosarcoma cells. Peptides 2009, 30, 1636–1642. [Google Scholar] [CrossRef]
- Chang, W.T.; Pan, C.Y.; Rajanbabu, V.; Cheng, C.W.; Chen, J.Y. Tilapia (Oreochromis mossambicus) antimicrobial peptide, hepcidin 1–5, shows antitumor activity in cancer cells. Peptides 2011, 32, 342–352. [Google Scholar] [CrossRef]
- Pan, C.Y.; Peng, K.C.; Lin, C.H.; Chen, J.Y. Transgenic expression of tilapia hepcidin 1–5 and shrimp chelonianin in zebrafish and their resistance to bacterial pathogens. Fish Shellfish Immunol. 2011, 31, 275–285. [Google Scholar] [CrossRef]
- Hsieh, J.C.; Pan, C.Y.; Chen, J.Y. Tilapia hepcidin (TH)2-3 as a transgene in transgenic fish enhances resistance to Vibrio vulnificus infection and causes variations in immune-related genes after infection by different bacterial species. Fish Shellfish Immunol. 2010, 29, 430–439. [Google Scholar] [CrossRef]
- Rajanbabu, V.; Pan, C.Y.; Lee, S.C.; Lin, W.J.; Lin, C.C.; Li, C.L.; Chen, J.Y. Tilapia hepcidin 2-3 peptide modulates lipopolysaccharide-induced cytokines and inhibits tumor necrosis factor-alpha through cyclooxygenase-2 and phosphodiesterase 4D. J. Biol. Chem. 2010, 285, 30577–30586. [Google Scholar]
- Rajanbabu, V.; Chen, J.Y. The antimicrobial peptide, tilapia hepcidin 2-3, and PMA differentially regulate the protein kinase C isoforms, TNF-alpha and COX-2, in mouse RAW264.7 macrophages. Peptides 2011, 32, 333–341. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. The hepcidin-ferroportin system as a therapeutic target in anemias and iron overload disorders. Hematol. Am. Soc. Hematol. Educ. Program. 2011, 2011, 538–542. [Google Scholar] [CrossRef]
- Fraenkel, P.G.; Gibert, Y.; Holzheimer, J.L.; Lattanzi, V.J.; Burnett, S.F.; Dooley, K.A.; Wingert, R.A.; Zon, L.I. Transferrin-a modulates hepcidin expression in zebrafish embryos. Blood 2009, 113, 2843–2850. [Google Scholar] [CrossRef]
- Chen, J.; Shi, Y.H.; Li, M.Y. Changes in transferrin and hepcidin genes expression in the liver of the fish Pseudosciaena crocea following exposure to cadmium. Arch. Toxicol. 2008, 82, 525–530. [Google Scholar] [CrossRef]
- Tomasinsig, L.; Zanetti, M. The cathelicidins—Structure, function and evolution. Curr. Protein Pept. Sci. 2005, 6, 23–34. [Google Scholar] [CrossRef]
- Shinnar, A.E.; Butler, K.L.; Park, H.J. Cathelicidin family of antimicrobial peptides: Proteolytic processing and protease resistance. Bioorg. Chem. 2003, 31, 425–436. [Google Scholar] [CrossRef]
- Maier, V.H.; Dorn, K.V.; Gudmundsdottir, B.K.; Gudmundsson, G.H. Characterisation of cathelicidin gene family members in divergent fish species. Mol. Immunol. 2008, 45, 3723–3730. [Google Scholar] [CrossRef]
- Broekman, D.C.; Frei, D.M.; Gylfason, G.A.; Steinarsson, A.; Jornvall, H.; Agerberth, B.; Gudmundsson, G.H.; Maier, V.H. Cod cathelicidin: Isolation of the mature peptide, cleavage site characterisation and developmental expression. Dev. Comp. Immunol. 2011, 35, 296–303. [Google Scholar] [CrossRef]
- Bridle, A.; Nosworthy, E.; Polinski, M.; Nowak, B. Evidence of an antimicrobial-immunomodulatory role of Atlantic salmon cathelicidins during infection with Yersinia ruckeri. PLoS One 2011, 6, e23417. [Google Scholar]
- Maier, V.H.; Schmitt, C.N.; Gudmundsdottir, S.; Gudmundsson, G.H. Bacterial DNA indicated as an important inducer of fish cathelicidins. Mol. Immunol. 2008, 45, 2352–2358. [Google Scholar] [CrossRef]
- Broekman, D.C.; Guethmundsson, G.H.; Maier, V.H. Differential regulation of cathelicidin in salmon and cod. Fish Shellfish Immunol. 2013, 35, 532–538. [Google Scholar] [CrossRef]
- Costa, M.M.; Maehr, T.; Diaz-Rosales, P.; Secombes, C.J.; Wang, T. Bioactivity studies of rainbow trout (Oncorhynchus mykiss) interleukin-6: Effects on macrophage growth and antimicrobial peptide gene expression. Mol. Immunol. 2011, 48, 1903–1916. [Google Scholar] [CrossRef]
- Hong, S.; Li, R.; Xu, Q.; Secombes, C.J.; Wang, T. Two types of TNF-alpha exist in teleost fish: Phylogeny, expression, and bioactivity analysis of type-II TNF-alpha3 in rainbow trout oncorhynchus mykiss. J. Immunol. 2013, 191, 5959–5972. [Google Scholar] [CrossRef]
- Shewring, D.M.; Zou, J.; Corripio-Miyar, Y.; Secombes, C.J. Analysis of the cathelicidin 1 gene locus in Atlantic cod (Gadus morhua). Mol. Immunol. 2011, 48, 782–787. [Google Scholar] [CrossRef]
- De Bruijn, I.; Belmonte, R.; Anderson, V.L.; Saraiva, M.; Wang, T.; van West, P.; Secombes, C.J. Immune gene expression in trout cell lines infected with the fish pathogenic oomycete Saprolegnia parasitica. Dev. Comp. Immunol. 2012, 38, 44–54. [Google Scholar] [CrossRef]
- Chettri, J.K.; Raida, M.K.; Kania, P.W.; Buchmann, K. Differential immune response of rainbow trout (Oncorhynchus mykiss) at early developmental stages (larvae and fry) against the bacterial pathogen Yersinia ruckeri. Dev. Comp. Immunol. 2012, 36, 463–474. [Google Scholar] [CrossRef]
- Choi, K.Y.; Chow, L.N.; Mookherjee, N. Cationic host defence peptides: Multifaceted role in immune modulation and inflammation. J. Innate Immun. 2012, 4, 361–370. [Google Scholar]
- Park, C.B.; Kim, M.S.; Kim, S.C. A novel antimicrobial peptide from Bufo bufo gargarizans. Biochem. Biophys. Res. Commun. 1996, 218, 408–413. [Google Scholar] [CrossRef]
- Parseghian, M.H.; Luhrs, K.A. Beyond the walls of the nucleus: The role of histones in cellular signaling and innate immunity. Biochem. Cell Biol. 2006, 84, 589–604. [Google Scholar] [CrossRef]
- Noga, E.J.; Ullal, A.J.; Corrales, J.; Fernandes, J.M. Application of antimicrobial polypeptide host defenses to aquaculture: Exploitation of downregulation and upregulation responses. Comp. Biochem. Physiol. Part D Genomics Proteomics 2011, 6, 44–54. [Google Scholar] [CrossRef]
- Robinette, D.; Wada, S.; Arroll, T.; Levy, M.G.; Miller, W.L.; Noga, E.J. Antimicrobial activity in the skin of the channel catfish Ictalurus punctatus: Characterization of broad-spectrum histone-like antimicrobial proteins. Cell. Mol. Life Sci. 1998, 54, 467–475. [Google Scholar] [CrossRef]
- Robinette, D.W.; Noga, E.J. Histone-like protein: A novel method for measuring stress in fish. Dis. Aquat. Org. 2001, 44, 97–107. [Google Scholar] [CrossRef]
- Burrowes, O.J.; Hadjicharalambous, C.; Diamond, G.; Lee, T.C. Evaluation of antimicrobial spectrum and cytotoxic activity of pleurocidin for food applicaitons. J. Food Sci. 2004, 69, 66–71. [Google Scholar]
- Bals, R.; Goldman, M.J.; Wilson, J.M. Mouse b-defensin 1 is a salt-sensitive antimicrobial peptide present in epithelia of the lung and urogenital tract. Infect. Immun. 1998, 66, 1225–1232. [Google Scholar]
- Lee, J.Y.; Yang, S.T.; Lee, S.K.; Jung, H.H.; Shin, S.Y.; Hahm, K.S.; Kim, J.I. Salt-resistant homodimeric bactenecin, a cathelicidin-derived antimicrobial peptide. FEBS J. 2008, 275, 3911–3920. [Google Scholar] [CrossRef]
- Tomita, T.; Hitomi, S.; Nagase, T.; Matsui, H.; Matsuse, T.; Kimura, S.; Ouchi, Y. Effect of ions on antibacterial activity of human beta defensin 2. Microbiol. Immunol. 2000, 44, 749–754. [Google Scholar] [CrossRef]
- Subramanian, S.; Ross, N.W.; MacKinnon, S.L. Myxinidin, a novel antimicrobial peptide from the epidermal mucus of hagfish, Myxine glutinosa L. Mar. Biotechnol. (N. Y.) 2009, 11, 748–757. [Google Scholar] [CrossRef]
- Olli, S.; Rangaraj, N.; Nagaraj, R. Effect of selectively introducing arginine and D-amino acids on the antimicrobial activity and salt sensitivity in analogs of human Beta-defensins. PLoS One 2013, 8, e77031. [Google Scholar]
- Mai, J.; Tian, X.L.; Gallant, J.W.; Merkley, N.; Biswas, Z.; Syvitski, R.; Douglas, S.E.; Ling, J.; Li, Y.H. A novel target-specific, salt-resistant antimicrobial peptide against the cariogenic pathogen Streptococcus mutans. Antimicrob. Agents Chemother. 2011, 55, 5205–5213. [Google Scholar] [CrossRef]
- Wu, S.P.; Huang, T.C.; Lin, C.C.; Hui, C.F.; Lin, C.H.; Chen, J.Y. Pardaxin, a fish antimicrobial peptide, exhibits antitumor activity toward murine fibrosarcoma in vitro and in vivo. Mar. Drugs 2012, 10, 1852–1872. [Google Scholar] [CrossRef]
- Wang, W.; Tao, R.; Tong, Z.; Ding, Y.; Kuang, R.; Zhai, S.; Liu, J.; Ni, L. Effect of a novel antimicrobial peptide chrysophsin-1 on oral pathogens and Streptococcus mutans biofilms. Peptides 2012, 33, 212–219. [Google Scholar] [CrossRef]
- Tao, R.; Tong, Z.; Lin, Y.; Xue, Y.; Wang, W.; Kuang, R.; Wang, P.; Tian, Y.; Ni, L. Antimicrobial and antibiofilm activity of pleurocidin against cariogenic microorganisms. Peptides 2011, 32, 1748–1754. [Google Scholar] [CrossRef]
- Pan, C.Y.; Rajanbabu, V.; Chen, J.Y.; Her, G.M.; Nan, F.H. Evaluation of the epinecidin-1 peptide as an active ingredient in cleaning solutions against pathogens. Peptides 2010, 31, 1449–1458. [Google Scholar] [CrossRef]
- Choi, H.; Lee, D.G. The influence of the N-terminal region of antimicrobial peptide pleurocidin on fungal apoptosis. J. Microbiol. Biotechnol. 2013, 23, 1386–1394. [Google Scholar] [CrossRef]
- Luders, T.; Birkemo, G.A.; Fimland, G.; Nissen-Meyer, J.; Nes, I.F. Strong synergy between a eukaryotic antimicrobial peptide and bacteriocins from lactic acid bacteria. Appl. Environ. Microbiol. 2003, 69, 1797–1799. [Google Scholar] [CrossRef]
- Patrzykat, A.; Zhang, L.; Mendoza, V.; Iwama, G.K.; Hancock, R.E. Synergy of histone-derived peptides of coho salmon with lysozyme and flounder pleurocidin. Antimicrob. Agents Chemother. 2001, 45, 1337–1342. [Google Scholar] [CrossRef]
- Ivanov, I.E.; Morrison, A.E.; Cobb, J.E.; Fahey, C.A.; Camesano, T.A. Creating antibacterial surfaces with the peptide chrysophsin-1. ACS Appl. Mater. Interfaces 2012, 4, 5891–5897. [Google Scholar] [CrossRef]
- Pan, C.Y.; Lee, S.C.; Rajanbabu, V.; Lin, C.H.; Chen, J.Y. Insights into the antibacterial and immunomodulatory functions of tilapia hepcidin (TH)2–3 against Vibrio vulnificus infection in mice. Dev. Comp. Immunol. 2012, 36, 166–173. [Google Scholar] [CrossRef]
- Souza, A.L.; Diaz-Dellavalle, P.; Cabrera, A.; Larranaga, P., Dalla-Rizza; de-Simone, S.G. Antimicrobial activity of pleurocidin is retained in Plc-2, a C-terminal 12-amino acid fragment. Peptides 2013, 45, 78–84. [Google Scholar] [CrossRef]
- Choi, H.; Lee, D.G. Antimicrobial peptide pleurocidin synergizes with antibiotics through hydroxyl radical formation and membrane damage, and exerts antibiofilm activity. Biochim. Biophys. Acta 2012, 1820, 1831–1838. [Google Scholar] [CrossRef]
- Som, A.; Vemparala, S.; Ivanov, I.; Tew, G.N. Synthetic mimics of antimicrobial peptides. Biopolymers 2008, 90, 83–93. [Google Scholar] [CrossRef]
- Beckloff, N.; Laube, D.; Castro, T.; Furgang, D.; Park, S.; Perlin, D.; Clements, D.; Tang, H.; Scott, R.W.; Tew, G.N.; et al. Activity of an antimicrobial peptide mimetic against planktonic and biofilm cultures of oral pathogens. Antimicrob. Agents Chemother. 2007, 51, 4125–4132. [Google Scholar] [CrossRef]
- Hua, J.; Yamarthy, R.; Felsenstein, S.; Scott, R.W.; Markowitz, K.; Diamond, G. Activity of antimicrobial peptide mimetics in the oral cavity: I. Activity against biofilms of Candida albicans. Mol. Oral Microbiol. 2010, 25, 418–425. [Google Scholar] [CrossRef]
- Hua, J.; Scott, R.W.; Diamond, G. Activity of antimicrobial peptide mimetics in the oral cavity: II. Activity against periopathogenic biofilms and anti-inflammatory activity. Mol. Oral Microbiol. 2010, 25, 426–432. [Google Scholar] [CrossRef]
- Hancock, R.E.; Nijnik, A.; Philpott, D.J. Modulating immunity as a therapy for bacterial infections. Nat. Rev. Microbiol. 2012, 10, 243–254. [Google Scholar]
- Terova, G.; Forchino, A.; Rimoldi, S.; Brambilla, F.; Antonini, M.; Saroglia, M. Bio-Mos: An effective inducer of dicentracin gene expression in European sea bass (Dicentrarchus labrax). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 153, 372–377. [Google Scholar] [CrossRef]
- Chang, C.I.; Pleguezuelos, O.; Zhang, Y.A.; Zou, J.; Secombes, C.J. Identification of a novel cathelicidin gene in the rainbow trout, Oncorhynchus mykiss. Infect. Immun. 2005, 73, 5053–5064. [Google Scholar] [CrossRef]
- Sung, W.S.; Lee, J.; Lee, D.G. Fungicidal effect and the mode of action of piscidin 2 derived from hybrid striped bass. Biochem. Biophys. Res. Commun. 2008, 371, 551–555. [Google Scholar] [CrossRef]
- Wang, Y.D.; Kung, C.W.; Chi, S.C.; Chen, J.Y. Inactivation of nervous necrosis virus infecting grouper (Epinephelus coioides) by epinecidin-1 and hepcidin 1-5 antimicrobial peptides, and downregulation of Mx2 and Mx3 gene expressions. Fish Shellfish Immunol. 2010, 28, 113–120. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Masso-Silva, J.A.; Diamond, G. Antimicrobial Peptides from Fish. Pharmaceuticals 2014, 7, 265-310. https://doi.org/10.3390/ph7030265
Masso-Silva JA, Diamond G. Antimicrobial Peptides from Fish. Pharmaceuticals. 2014; 7(3):265-310. https://doi.org/10.3390/ph7030265
Chicago/Turabian StyleMasso-Silva, Jorge A., and Gill Diamond. 2014. "Antimicrobial Peptides from Fish" Pharmaceuticals 7, no. 3: 265-310. https://doi.org/10.3390/ph7030265
APA StyleMasso-Silva, J. A., & Diamond, G. (2014). Antimicrobial Peptides from Fish. Pharmaceuticals, 7(3), 265-310. https://doi.org/10.3390/ph7030265