

Optimization of the Search for Neuroprotectors among Bioflavonoids
 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Docking Analysis
3.2. The Virtual Screening Program
- Linear Regression
- 2.
- Support Vector Machine Regression
- 3.
- Random Forest Regression
- 4.
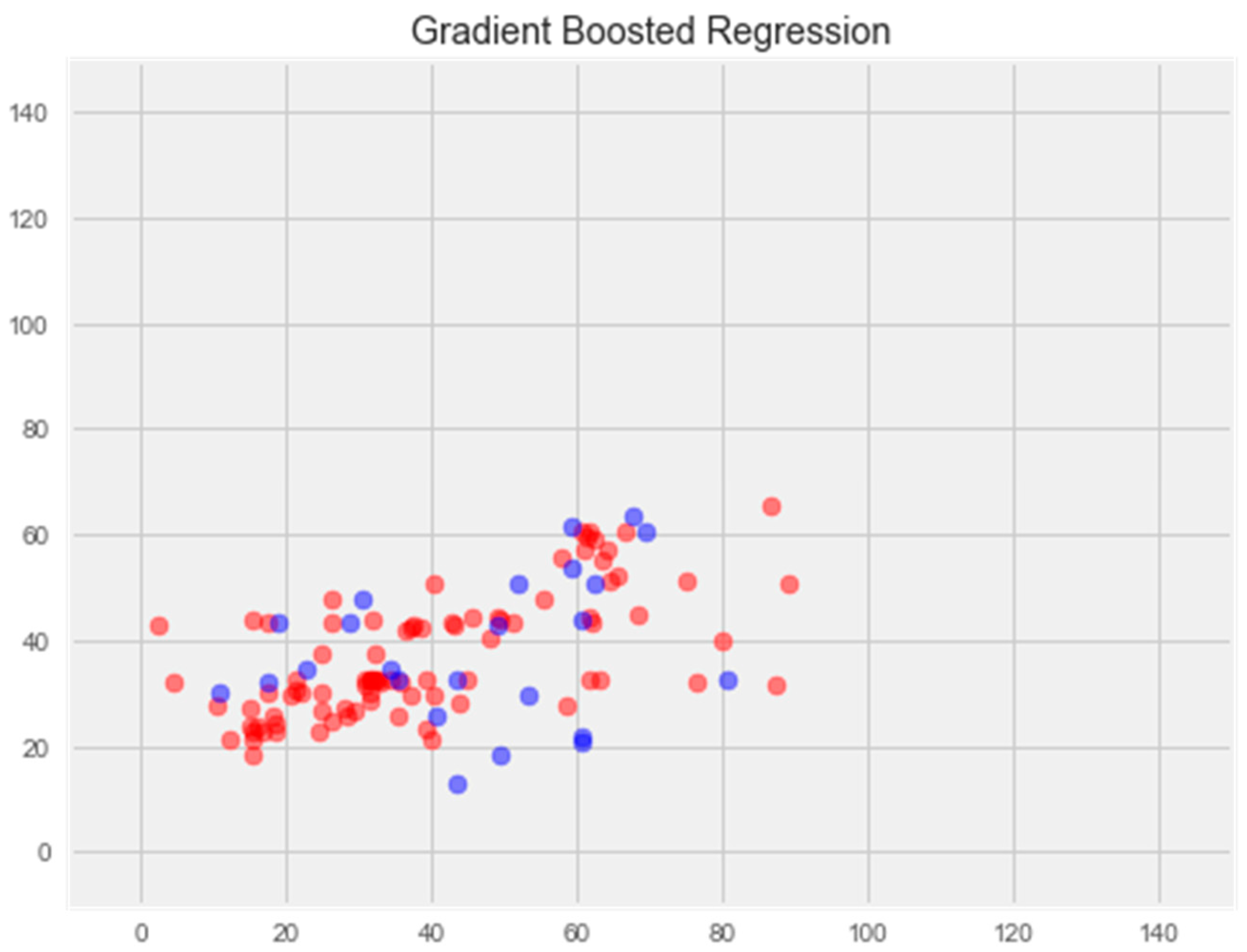
- Gradient Boosting Regression
- 5.
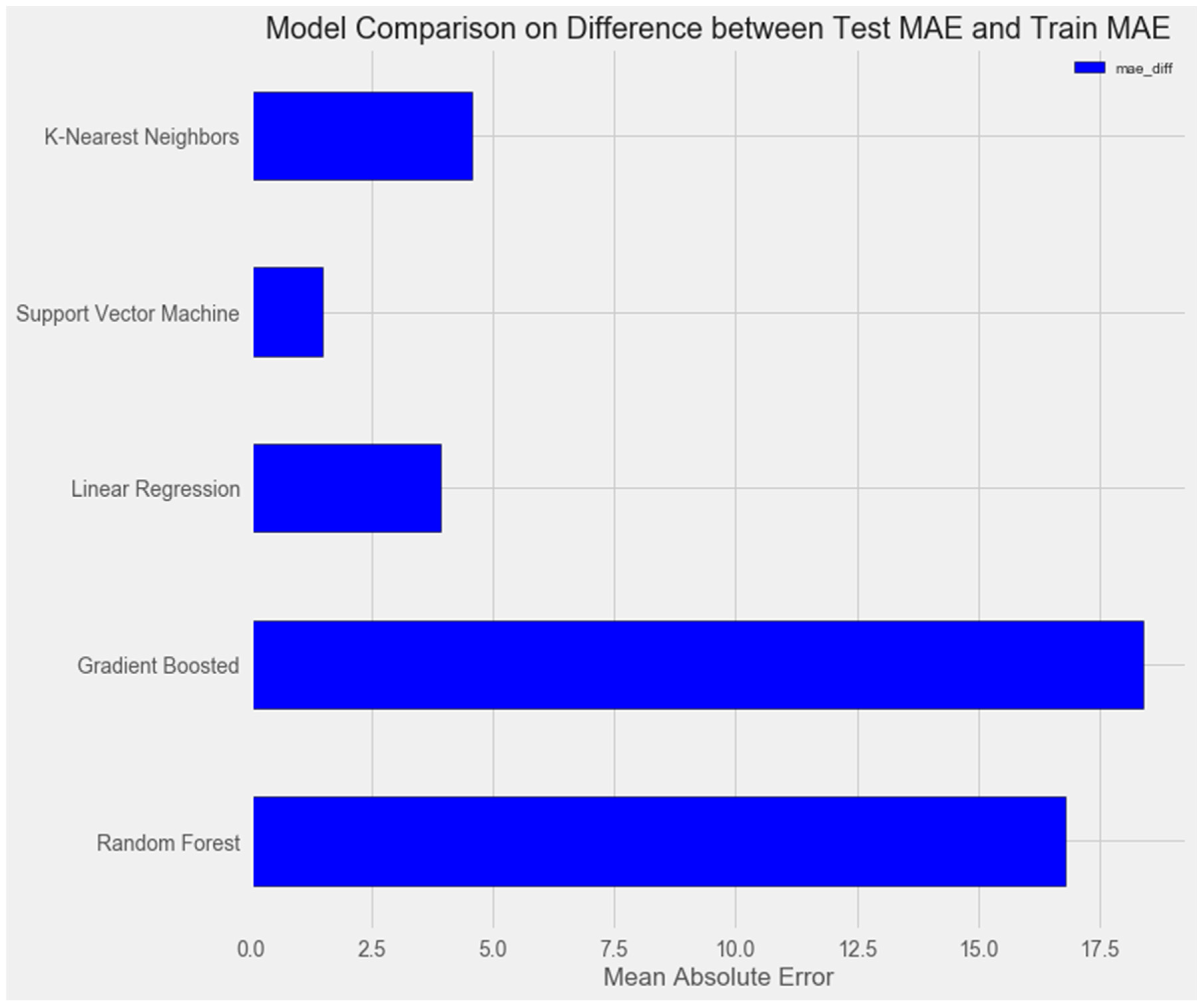
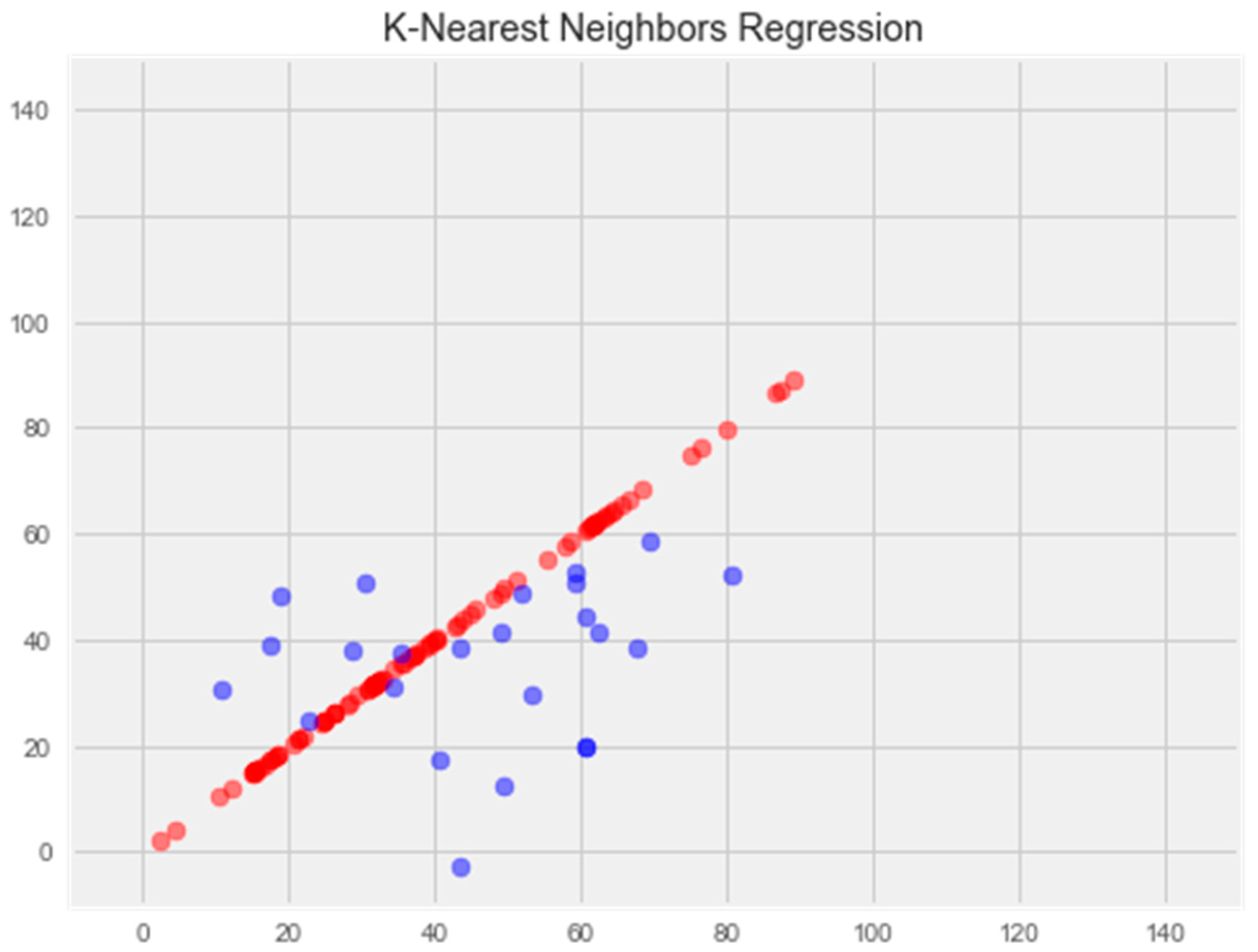
- K-Nearest Neighbors Regression
3.2.1. Linear Regression Model
3.2.2. Regression Model Using the Support Vector Method
3.2.3. Random Forest Model
3.2.4. Gradient Boosting Model
3.2.5. K-Nearest Neighbors Model
3.3. In Vitro Studies
3.4. Experimental Model of Multiple Sclerosis
3.5. Drugs and Doses
- (1)
- Intact (10 rats);
- (2)
- Control—untreated with EAE, received physiological saline (10 rats);
- (3)
- Animals with EAE receiving baseline treatment—methylprednisolone (MP) at 3.4 mg/kg, administered intraperitoneally, slowly, in saline (10 rats);
- (4)
- Animals with EAE receiving MP + catechin at 10 mg/kg, administered intragastrically (10 rats);
- (5)
- Animals with EAE receiving MP + antioxidant mexidol at 250 mg/kg, administered intragastrically (10 rats).
3.6. Preparation of Biological Material
3.7. Enzyme-Linked Immunosorbent Assay
3.8. Statistical Methods of this Study
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Papiri, G.; D’Andreamatteo, G.; Cacchiò, G.; Alia, S.; Silvestrini, M.; Paci, C.; Luzzi, S.; Vignini, A. Multiple Sclerosis: Inflammatory and Neuroglial Aspects. Curr. Issues Mol. Biol. 2023, 45, 1443–1470. [Google Scholar] [CrossRef] [PubMed]
- Balasa, R.; Barcutean, L.; Mosora, O.; Manu, D. Reviewing the Significance of Blood–Brain Barrier Disruption in Multiple Sclerosis Pathology and Treatment. Int. J. Mol. Sci. 2021, 22, 8370. [Google Scholar] [CrossRef] [PubMed]
- Bierhansl, L.; Hartung, H.P.; Aktas, O.; Ruck, T.; Roden, M.; Meuth, S.G. Thinking outside the box: Non-canonical targets in multiple sclerosis. Nat. Rev. Drug Discov. 2022, 21, 578–600. [Google Scholar] [CrossRef] [PubMed]
- Kölliker-Frers, R.; Udovin, L.; Otero-Losada, M.; Kobiec, T.; Herrera, M.I.; Palacios, J.; Razzitte, G.; Capani, F. Neuroinflammation: An Integrating Overview of Reactive-Neuroimmune Cell Interactions in Health and Disease. Mediators Inflamm. 2021, 2021, 9999146. [Google Scholar] [CrossRef] [PubMed]
- Isik, S.; Kiyak, B.Y.; Akbayir, R.; Seyhali, R.; Arpaci, T. Microglia Mediated Neuroinflammation in Parkinson’s Disease. Cells 2023, 12, 1012. [Google Scholar] [CrossRef] [PubMed]
- Alraawi, Z.; Banerjee, N.; Mohanty, S.; Kumar, T.K.S. Amyloidogenesis: What Do We Know So Far? Int. J. Mol. Sci. 2022, 23, 13970. [Google Scholar] [CrossRef] [PubMed]
- Fagla, B.M.; Buhimschi, I.A. Protein Misfolding in Pregnancy: Current Insights, Potential Mechanisms, and Implications for the Pathogenesis of Preeclampsia. Molecules 2024, 29, 610. [Google Scholar] [CrossRef] [PubMed]
- Hadjiagapiou, M.S.; Krashias, G.; Deeba, E.; Christodoulou, C.; Pantzaris, M.; Lambrianides, A. A Preclinical Investigation on the Role of IgG Antibodies against Coagulant Components in Multiple Sclerosis. Biomedicines 2023, 11, 906. [Google Scholar] [CrossRef]
- Singh, M.K.; Shin, Y.; Ju, S.; Han, S.; Choe, W.; Yoon, K.S.; Kim, S.S.; Kang, I. Heat Shock Response and Heat Shock Proteins: Current Understanding and Future Opportunities in Human Diseases. Int. J. Mol. Sci. 2024, 25, 4209. [Google Scholar] [CrossRef]
- Belenichev, I.; Popazova, O.; Bukhtiyarova, N.; Savchenko, D.; Oksenych, V.; Kamyshnyi, O. Modulating Nitric Oxide: Implications for Cytotoxicity and Cytoprotection. Antioxidants 2024, 13, 504. [Google Scholar] [CrossRef]
- Coutinho Costa, V.G.; Araújo, S.E.-S.; Alves-Leon, S.V.; Gomes, F.C.A. Central nervous system demyelinating diseases: Glial cells at the hub of pathology. Front. Immunol. 2023, 14, 1135540. [Google Scholar] [CrossRef] [PubMed]
- Hullsiek, R.; Li, Y.; Snyder, K.M.; Wang, S.; Di, D.; Borgatti, A.; Lee, C.; Moore, P.F.; Zhu, C.; Fattori, C.; et al. Examination of IgG Fc Receptor CD16A and CD64 Expression by Canine Leukocytes and Their ADCC Activity in Engineered NK Cells. Front. Immunol. 2022, 13, 841859. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Mendoza, A.A.; Mendoza-Magaña, M.L.; Ramírez-Herrera, M.A.; Hernández-Nazara, Z.H.; Domínguez-Rosales, J.A. Nitrooxidative Stress and Neuroinflammation Caused by Air Pollutants Are Associated with the Biological Markers of Neurodegenerative Diseases. Antioxidants 2024, 13, 326. [Google Scholar] [CrossRef] [PubMed]
- Park, G.Y.; Jamerlan, A.; Shim, K.H.; An, S.S.A. Diagnostic and Treatment Approaches Involving Transthyretin in Amyloidogenic Diseases. Int. J. Mol. Sci. 2019, 20, 2982. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, C.; Minniti, M.; Susini, V.; Caponi, L.; Panichella, G.; Castiglione, V.; Aimo, A.; Emdin, M.; Vergaro, G.; Franzini, M. The Journey of Human Transthyretin: Synthesis, Structure Stability, and Catabolism. Biomedicines 2022, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, E.; Ożyhar, A. Transthyretin: From Structural Stability to Osteoarticular and Cardiovascular Diseases. Cells 2021, 10, 1768. [Google Scholar] [CrossRef] [PubMed]
- Radwan, A.A.; Alanazi, F.K.; Raish, M. Design and synthesis of multi-functional small-molecule based inhibitors of amyloid-β aggregation: Molecular modeling and in vitro evaluation. PLoS ONE 2023, 18, e0286195. [Google Scholar] [CrossRef] [PubMed]
- Rinauro, D.J.; Chiti, F.; Vendruscolo, M.; Limbocker, R. Misfolded protein oligomers: Mechanisms of formation, cytotoxic effects, and pharmacological approaches against protein misfolding diseases. Mol. Neurodegenerat. 2024, 19, 20. [Google Scholar] [CrossRef] [PubMed]
- Stroo, E.; Koopman, M.; Nollen, E.A.; Mata-Cabana, A. Cellular Regulation of Amyloid Formation in Aging and Disease. Front. Neurosci. 2017, 11, 64. [Google Scholar] [CrossRef]
- Li, J.; Guo, M.; Chen, L.; Chen, Z.; Fu, Y.; Chenet, Y. p53 amyloid aggregation in cancer: Function, mechanism, and therapy. Exp. Hematol. Oncol. 2022, 11, 66. [Google Scholar] [CrossRef]
- Dakterzada, F.; Jové, M.; Cantero, J.L.; Pamplona, R.; Piñoll-Ripoll, G. Plasma and cerebrospinal fluid nonenzymatic protein damage is sustained in Alzheimer’s disease. Redox Biol. 2023, 64, 102772. [Google Scholar] [CrossRef] [PubMed]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Futur. J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Barber, K.; Mendonca, P.; Soliman, K.F.A. The Neuroprotective Effects and Therapeutic Potential of the Chalcone Cardamonin for Alzheimer’s Disease. Brain Sci. 2023, 13, 145. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, A.; Patanè, G.T.; Tellone, E.; Barreca, D.; Ficarra, S.; Misiti, F.; Laganà, G. The Neuroprotective Potentiality of Flavonoids on Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 14835. [Google Scholar] [CrossRef] [PubMed]
- Ciccone, L.; Tonali, N.; Fruchart-Gaillard, C.; Barlettani, L.; Rossello, A.; Braca, A.; Orlandini, E.; Nencetti, S. Antioxidant Quercetin 3-O-Glycosylated Plant Flavonols Contribute to Transthyretin Stabilization. Crystals 2022, 12, 638. [Google Scholar] [CrossRef]
- Mokra, D.; Adamcakova, J.; Mokry, J. Green Tea Polyphenol (-)-Epigallocatechin-3-Gallate (EGCG): A Time for a New Player in the Treatment of Respiratory Diseases? Antioxidants 2022, 11, 1566. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef] [PubMed]
- Ngoc, T.D.; Le, T.N.; Nguyen, T.V.A.; Mechler, A.; Hoa, N.T.; Nam, N.L.; Vo, Q.V. Mechanistic and Kinetic Studies of the Radical Scavenging Activity of 5-O-Methylnorbergenin: Theoretical and Experimental Insights. J. Phys. Chem. B 2022, 126, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Saldaño, T.E.; Zanotti, G.; Parisi, G.; Fernandez-Alberti, S. Evaluating the effect of mutations and ligand binding on transthyretin homotetramer dynamics. PLoS ONE 2017, 12, e0181019. [Google Scholar]
- Cody, V.; Truong, J.Q.; Holdsworth, B.A.; Holien, J.K.; Richardson, S.J.; Chalmers, D.K.; Craik, D.J. Structural Analysis of the Complex of Human Transthyretin with 3′,5′-Dichlorophenylanthranilic Acid at 1.5 Å Resolution. Molecules 2022, 27, 7206. [Google Scholar] [CrossRef]
- Koga, T.; Meydani, M. Effect of plasma metabolites of (+)-catechin and quercetin on monocyte adhesion to human aortic endothelial cells. Am. J. Clin. Nutr. 2001, 73, 941–948. [Google Scholar] [CrossRef]
- Meyer, A.S.; Heinonen, M.; Frankel, E.N. Antioxidant interactions of catechin, cya-nidin, caffeic acid, quercetin, and ellagic acid on human LDL oxidation. Food Chemistry 1998, 61, 71–75. [Google Scholar] [CrossRef]
- Josiah, S.S.; Famusiwa, C.D.; Crown, O.O.; Lawal, A.O.; Olaleye, M.T.; Akindahunsi, A.A.; Akinmoladun, A.C. Neuroprotective effects of catechin and quercetin in experimental Parkinsonism through modulation of dopamine metabolism and expression of IL-1β, TNF-α, NF-κB, IκKB, and p53 genes in male Wistar rats. Neurotoxicology 2022, 90, 158–171. [Google Scholar] [CrossRef]
- Forloni, G. Alpha Synuclein: Neurodegeneration and Inflammation. Int. J. Mol. Sci. 2023, 24, 5914. [Google Scholar] [CrossRef]
- Zhao, J.; Shi, Q.; Zheng, Y.; Liu, Q.; He, Z.; Gao, Z.; Liu, Q. Insights Into the Mechanism of Tyrosine Nitration in Preventing β-Amyloid Aggregation in Alzheimer’s Disease. Front. Mol. Neurosci. 2021, 14, 619836. [Google Scholar] [CrossRef]
- Chen, S.; Saeed, A.F.U.H.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in immunoregulation and therapeutics. Signal Transduct. Target Ther. 2023, 8, 207. [Google Scholar] [CrossRef]
- Wen, L.; Wu, D.; Tan, X.; Zhong, M.; Xing, J.; Li, W.; Li, D.; Cao, F. The Role of Catechins in Regulating Diabetes: An Update Review. Nutrients 2022, 14, 4681. [Google Scholar] [CrossRef]
- Florio, P.; Folli, C.; Cianci, M.; Del Rio, D.; Zanotti, G.; Berni, R. Transthyretin Binding Heterogeneity and Anti-amyloidogenic Activity of Natural Polyphenols and Their Metabolites. J. Biol. Chem. 2015, 290, 29769–29780. [Google Scholar] [CrossRef]
- Nefodov, O.O.; Belenichev, I.F.; Fedchenko, M.P.; Popazova, O.O.; Ryzhenko, V.P.; Morozova, O.V. Evaluation of methods of modeling and formation of experimental allergic encephalomyelitis. Res. Results Pharmacol. 2022, 8, 37–48. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Affinity (kcal/mol) to Human Transthyretin | Docking 2D Visualization | |
|---|---|---|---|
| Thyroxin | −6.4 | 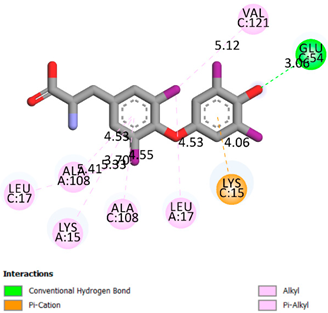 | |
| Quercetin | −6.7 | 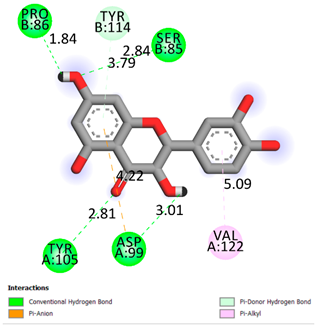 | |
| Catechin | −6.2 | 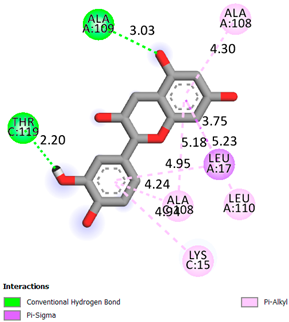 | |
| Epicatechin | −7.2 |  | |
| Catechin-3-gallate | −7.6 |  | |
| Epicatechin-3-gallate | −7.3 | 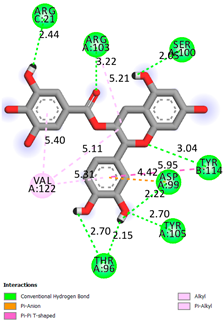 | |
| Epigallocatechin 3-O-Gallate | −7.7 | 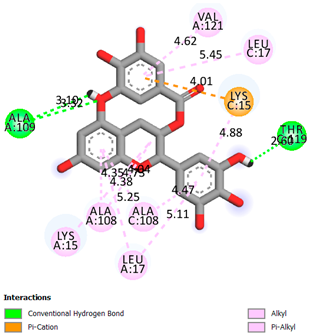 | |
| Gallocatechin-3-gallate | −7.5 |  | |
| Kaempferol | −7.2 | 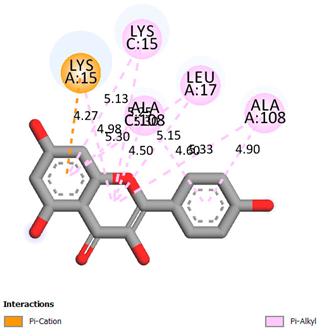 | |
| Luteolin | −8.0 |  | |
| Procyanidin B1 | −7.4 | 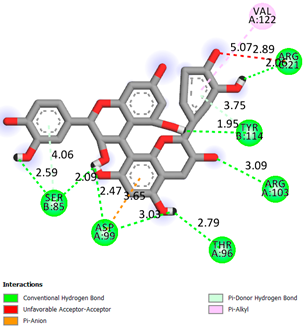 | |
| Procyanidin B2 | −8.6 | 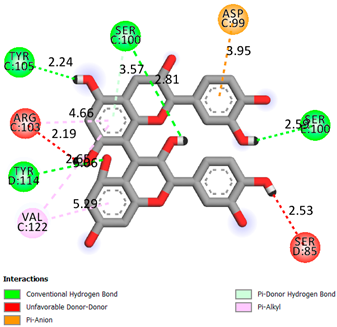 | |
| Procyanidin B3 | −8.7 | 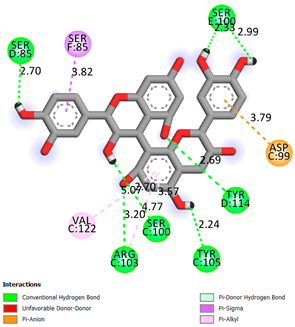 | |
| Experimental Series | Variation in Optical Density at 280 nm (Protein Binding) | Variation in Optical Density at 225 nm (Level of Displaced Thyroxine) |
|---|---|---|
| Indicators | ∆ | ∆ |
| Control (incubation mixture without catechin) | 0 ± 0 | 0 ± 0 |
| Experimental (incubation mixture containing catechin) | 0.063 ± 0.0001 ** | 0.082 ± 0.0002 ** |
| Code | AOA Results at 10−6 M | AOA Prediction, %. | |
|---|---|---|---|
| E, M ± m | % | ||
| Catechin | 1.625 ± 0.001 | 43.67 | 55.15 |
| Control | 1.131 ± 0.002 ** | - | |
| Indicators | Groups of Animals | |||
|---|---|---|---|---|
| Control, EAE (n = 10) | MP (n = 10) | Catechin + MP (n = 10) | Mexidol + MP (n = 10) | |
| % of sick animals (total/severe) | 100/70 | 80/30 | 80/10 *1 | 80/20 * |
| Average clinic index at the peak of EAE, points | 2.6 + 0.5 | 1.80 + 0.5 | 0.9 + 0.5 *1 | 1.65 + 0.152 |
| Average cumulative index, points | 27.2 + 1.5 | 9.4 + 0.4 * | 6.2 + 0.4 *1 | 7.5 + 0.6 * |
| Duration of EAE, days (Student’s test) | 16.0 + 1.2 | 8.4 + 0.7 * | 6.4 + 0.2 *1 | 7.2 + 0.8 * |
| Experimental Groups | NSE, ng/mL | S-100, ng/mL |
|---|---|---|
| Intact (n = 10) | 0.223 ± 0.015 | 0.088 ± 0.002 |
| EAE (control) (n = 10) | 9.11 ± 0.15 1 | 0.97 ± 0.015 1 |
| MP (n = 10) | 9.15 ± 0.14 1 | 0.92 ± 0.033 1 |
| MP+ Mexidol (n = 10) | 7.11 ± 0.21 *1,2 | 0.65 ± 0.042 *1,2 |
| MP+ catechin (n = 10) | 5.74 ± 0.11 *1,2,3 | 0.438 ± 0.014 *1,2,3 |
| Experimental Groups | Nitrotyrosine, ng/mL | IL-1b, ng/mL |
|---|---|---|
| Intact (n = 10) | 0.88 ± 0.042 | 0.31 ± 0.018 |
| EAE (control) (n = 10) | 9.89 ± 0.33 *1 | 3.88 ± 0.055 1 |
| MP (n = 10) | 8.11 ± 0.40 1 | 1.44 ± 0.022 *1 |
| MP+ Mexidol (n = 10) | 5.32 ± 0.32 *1,2, | 1.39 ± 0.033 *1, |
| MP+ catechin (n = 10) | 4.11 ± 0.07 *1,2,3 | 1.00 ± 0.02 *2,3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belenichev, I.; Ryzhenko, V.; Popazova, O.; Bukhtiyarova, N.; Gorchakova, N.; Oksenych, V.; Kamyshnyi, O. Optimization of the Search for Neuroprotectors among Bioflavonoids. Pharmaceuticals 2024, 17, 877. https://doi.org/10.3390/ph17070877
Belenichev I, Ryzhenko V, Popazova O, Bukhtiyarova N, Gorchakova N, Oksenych V, Kamyshnyi O. Optimization of the Search for Neuroprotectors among Bioflavonoids. Pharmaceuticals. 2024; 17(7):877. https://doi.org/10.3390/ph17070877
Chicago/Turabian StyleBelenichev, Igor, Victor Ryzhenko, Olena Popazova, Nina Bukhtiyarova, Nadia Gorchakova, Valentyn Oksenych, and Oleksandr Kamyshnyi. 2024. "Optimization of the Search for Neuroprotectors among Bioflavonoids" Pharmaceuticals 17, no. 7: 877. https://doi.org/10.3390/ph17070877
APA StyleBelenichev, I., Ryzhenko, V., Popazova, O., Bukhtiyarova, N., Gorchakova, N., Oksenych, V., & Kamyshnyi, O. (2024). Optimization of the Search for Neuroprotectors among Bioflavonoids. Pharmaceuticals, 17(7), 877. https://doi.org/10.3390/ph17070877