
IL-13Rα2 Is Involved in Resistance to Doxorubicin and Survival of Osteosarcoma Patients

 , , ,
, , ,  , ,
, , 

Abstract

1. Introduction
2. Results
2.1. Higher Expression of IL-13Rα2 Is Linked to Reduced Survival in Osteosarcoma Patients
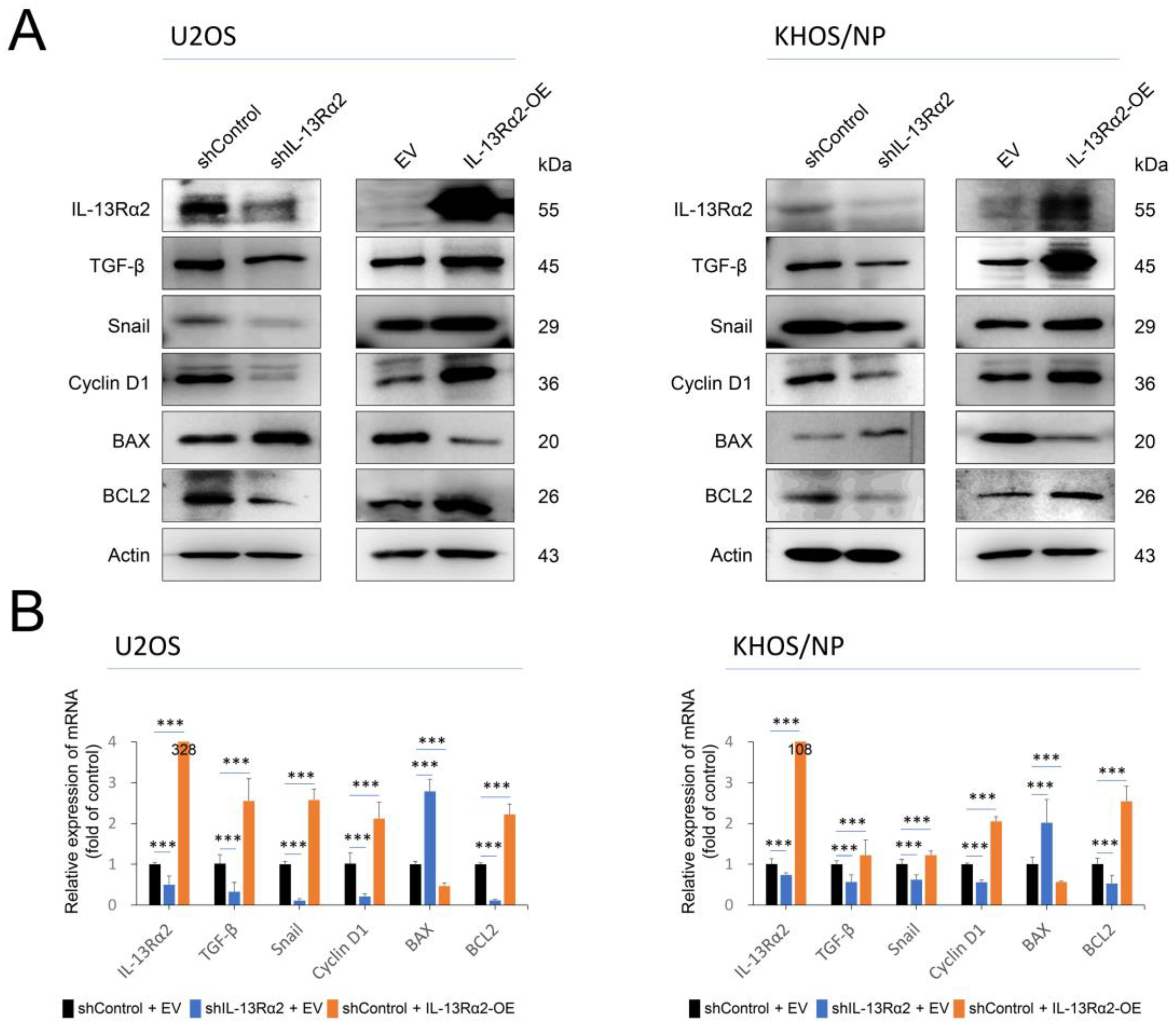
2.2. IL-13Rα2 Is Involved in the Proliferation and Invasion Activity of Osteosarcoma Cells
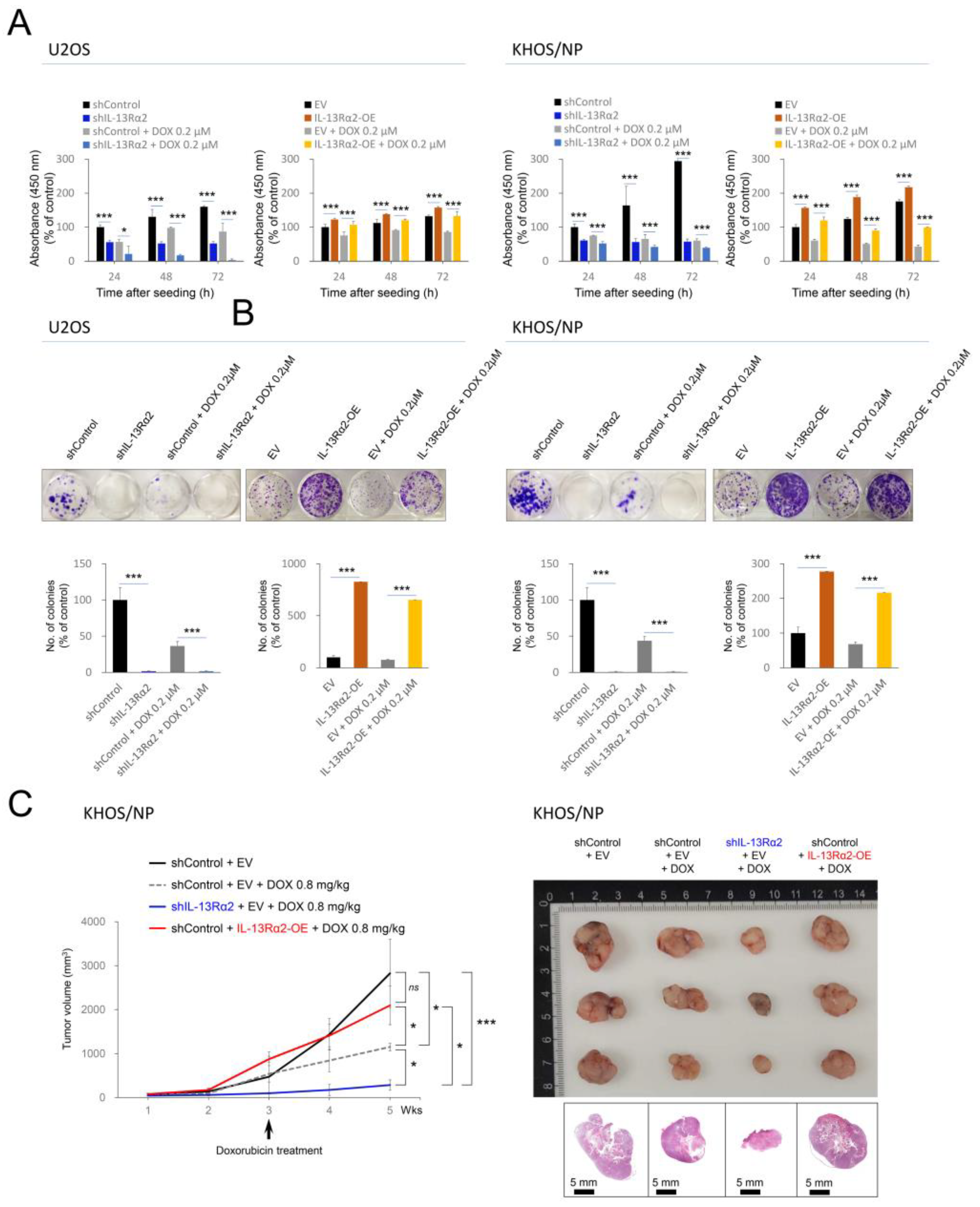
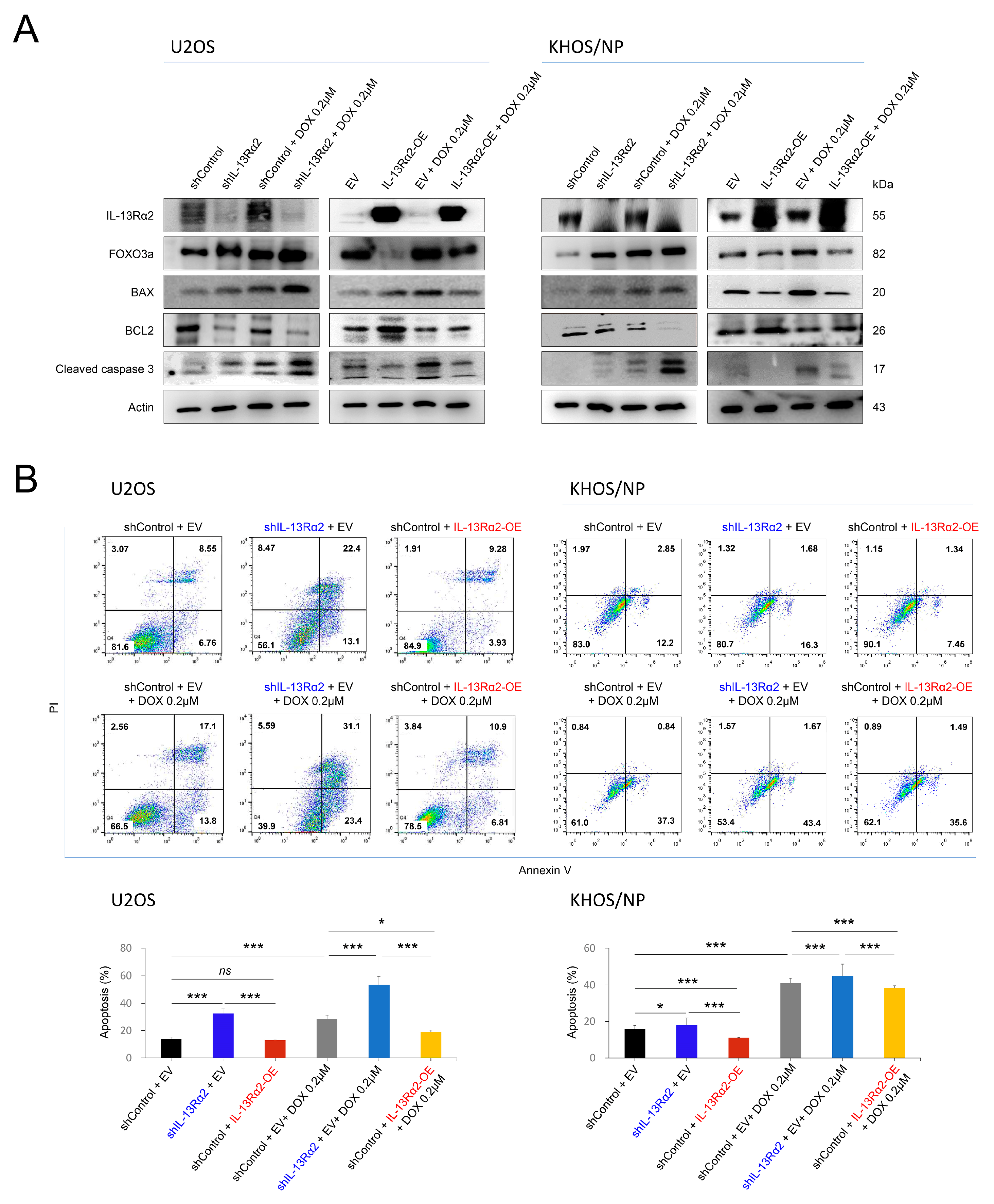
2.3. IL-13Rα2 Is Involved in Resistance to the Cytotoxic Effect of Doxorubicin and Silencing of IL-13Rα2 Sensitized Osteosarcoma Cells to Doxorubicin
3. Discussion
4. Materials and Methods
4.1. Osteosarcoma Patients and Tissue Samples
4.2. Immunohistochemical Staining and Scoring of Tissue Microarrays
4.3. Cell Lines, Reagents, and Transfection
4.4. Cell Proliferation and Colony-Forming Assays
4.5. Migration and Invasion Assay
4.6. Western Blot
4.7. Flow Cytometric Analysis for Apoptosis
4.8. Quantitative Reverse-Transcription Polymerase Chain Reaction
4.9. In Vivo Tumorigenic Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aman, M.J.; Tayebi, N.; Obiri, N.I.; Puri, R.K.; Modi, W.S.; Leonard, W.J. cDNA cloning and characterization of the human interleukin 13 receptor alpha chain. J. Biol. Chem. 1996, 271, 29265–29270. [Google Scholar] [CrossRef] [PubMed]
- Caput, D.; Laurent, P.; Kaghad, M.; Lelias, J.M.; Lefort, S.; Vita, N.; Ferrara, P. Cloning and characterization of a specific interleukin (IL)-13 binding protein structurally related to the IL-5 receptor alpha chain. J. Biol. Chem. 1996, 271, 16921–16926. [Google Scholar] [CrossRef] [PubMed]
- Zurawski, S.M.; Vega, F., Jr.; Huyghe, B.; Zurawski, G. Receptors for interleukin-13 and interleukin-4 are complex and share a novel component that functions in signal transduction. EMBO J. 1993, 12, 2663–2670. [Google Scholar] [CrossRef]
- Choi, W.K.; Hussein, U.K.; Ahmed, A.G.; Zhang, J.; Kim, K.M.; Ahn, A.R.; Park, H.S.; Park, S.H.; Cho, D.H.; Jang, K.Y. Co-operative roles of IL-4Ralpha and IL-13Ralpha1 in the progression of ovarian carcinomas and the survival of ovarian carcinoma patients. Cancer Commun. 2023, 43, 850–854. [Google Scholar] [CrossRef]
- Hage, T.; Sebald, W.; Reinemer, P. Crystal structure of the interleukin-4/receptor alpha chain complex reveals a mosaic binding interface. Cell 1999, 97, 271–281. [Google Scholar] [CrossRef]
- Zhang, J.G.; Hilton, D.J.; Willson, T.A.; McFarlane, C.; Roberts, B.A.; Moritz, R.L.; Simpson, R.J.; Alexander, W.S.; Metcalf, D.; Nicola, N.A. Identification, purification, and characterization of a soluble interleukin (IL)-13-binding protein. Evidence that it is distinct from the cloned Il-13 receptor and Il-4 receptor alpha-chains. J. Biol. Chem. 1997, 272, 9474–9480. [Google Scholar] [CrossRef]
- Karmele, E.P.; Pasricha, T.S.; Ramalingam, T.R.; Thompson, R.W.; Gieseck, R.L., 3rd; Knilans, K.J.; Hegen, M.; Farmer, M.; Jin, F.; Kleinman, A.; et al. Anti-IL-13Ralpha2 therapy promotes recovery in a murine model of inflammatory bowel disease. Mucosal Immunol. 2019, 12, 1174–1186. [Google Scholar] [CrossRef]
- Furue, M.; Ulzii, D.; Nakahara, T.; Tsuji, G.; Furue, K.; Hashimoto-Hachiya, A.; Kido-Nakahara, M. Implications of IL-13Ralpha2 in atopic skin inflammation. Allergol. Int. 2020, 69, 412–416. [Google Scholar] [CrossRef]
- Lumsden, R.V.; Worrell, J.C.; Boylan, D.; Walsh, S.M.; Cramton, J.; Counihan, I.; O’Beirne, S.; Medina, M.F.; Gauldie, J.; Fabre, A.; et al. Modulation of pulmonary fibrosis by IL-13Rα2. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 308, L710–L718. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, J.; Yang, Y.Z.; Wang, F.; Jiang, H.; Chen, H.D.; Wu, H.Y.; Sai, K.; Hu, W.M. IL13RA2 is overexpressed in malignant gliomas and related to clinical outcome of patients. Am. J. Transl. Res. 2020, 12, 4702–4714. [Google Scholar]
- Kioi, M.; Kawakami, M.; Shimamura, T.; Husain, S.R.; Puri, R.K. Interleukin-13 receptor alpha2 chain: A potential biomarker and molecular target for ovarian cancer therapy. Cancer 2006, 107, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, M.; Kawakami, K.; Kasperbauer, J.L.; Hinkley, L.L.; Tsukuda, M.; Strome, S.E.; Puri, R.K. Interleukin-13 receptor alpha2 chain in human head and neck cancer serves as a unique diagnostic marker. Clin. Cancer Res. 2003, 9, 6381–6388. [Google Scholar] [PubMed]
- Okamoto, H.; Yoshimatsu, Y.; Tomizawa, T.; Kunita, A.; Takayama, R.; Morikawa, T.; Komura, D.; Takahashi, K.; Oshima, T.; Sato, M.; et al. Interleukin-13 receptor α2 is a novel marker and potential therapeutic target for human melanoma. Sci. Rep. 2019, 9, 1281. [Google Scholar] [CrossRef] [PubMed]
- Barderas, R.; Bartolomé, R.A.; Fernandez-Aceñero, M.J.; Torres, S.; Casal, J.I. High expression of IL-13 receptor α2 in colorectal cancer is associated with invasion, liver metastasis, and poor prognosis. Cancer Res. 2012, 72, 2780–2790. [Google Scholar] [CrossRef]
- Joshi, B.H.; Plautz, G.E.; Puri, R.K. Interleukin-13 receptor alpha chain: A novel tumor-associated transmembrane protein in primary explants of human malignant gliomas. Cancer Res. 2000, 60, 1168–1172. [Google Scholar]
- Kang, M.A.; Lee, J.; Lee, C.M.; Park, H.S.; Jang, K.Y.; Park, S.H. IL13Rα2 Is Involved in the Progress of Renal Cell Carcinoma through the JAK2/FOXO3 Pathway. J. Pers. Med. 2021, 11, 284. [Google Scholar] [CrossRef]
- Márquez-Ortiz, R.A.; Contreras-Zárate, M.J.; Tesic, V.; Alvarez-Eraso, K.L.F.; Kwak, G.; Littrell, Z.; Costello, J.C.; Sreekanth, V.; Ormond, D.R.; Karam, S.D.; et al. IL13Rα2 Promotes Proliferation and Outgrowth of Breast Cancer Brain Metastases. Clin. Cancer Res. 2021, 27, 6209–6221. [Google Scholar] [CrossRef]
- Bartolomé, R.A.; Martín-Regalado, Á.; Jaén, M.; Zannikou, M.; Zhang, P.; de Los Ríos, V.; Balyasnikova, I.V.; Casal, J.I. Protein Tyrosine Phosphatase-1B Inhibition Disrupts IL13Rα2-Promoted Invasion and Metastasis in Cancer Cells. Cancers 2020, 12, 500. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Suzuki, A.; Leland, P.; Joshi, B.H.; Puri, R.K. Identification of a novel role of IL-13Rα2 in human Glioblastoma multiforme: Interleukin-13 mediates signal transduction through AP-1 pathway. J. Transl. Med. 2018, 16, 369. [Google Scholar] [CrossRef]
- Guarino, M. Src signaling in cancer invasion. J. Cell Physiol. 2010, 223, 14–26. [Google Scholar] [CrossRef]
- Fichtner-Feigl, S.; Strober, W.; Kawakami, K.; Puri, R.K.; Kitani, A. IL-13 signaling through the IL-13alpha2 receptor is involved in induction of TGF-beta1 production and fibrosis. Nat. Med. 2006, 12, 99–106. [Google Scholar] [CrossRef] [PubMed]
- WHO Classification of Tumours Editorial Board. Soft Tissue and Bone Tumours, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Steliarova-Foucher, E.; Colombet, M.; Ries, L.A.G.; Moreno, F.; Dolya, A.; Bray, F.; Hesseling, P.; Shin, H.Y.; Stiller, C.A. International incidence of childhood cancer, 2001–2010: A population-based registry study. Lancet Oncol. 2017, 18, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.; Gianferante, D.M.; Zhu, B.; Mirabello, L. Osteosarcoma: A Surveillance, Epidemiology, and End Results program-based analysis from 1975 to 2017. Cancer 2022, 128, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Broadhead, M.L.; Clark, J.C.; Myers, D.E.; Dass, C.R.; Choong, P.F. The molecular pathogenesis of osteosarcoma: A review. Sarcoma 2011, 2011, 959248. [Google Scholar] [CrossRef]
- Weichselbaum, R.; Little, J.B.; Nove, J. Response of human osteosarcoma in vitro to irradiation: Evidence for unusual cellular repair activity. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1977, 31, 295–299. [Google Scholar] [CrossRef]
- Anninga, J.K.; Gelderblom, H.; Fiocco, M.; Kroep, J.R.; Taminiau, A.H.; Hogendoorn, P.C.; Egeler, R.M. Chemotherapeutic adjuvant treatment for osteosarcoma: Where do we stand? Eur. J. Cancer 2011, 47, 2431–2445. [Google Scholar] [CrossRef]
- Hattinger, C.M.; Fanelli, M.; Tavanti, E.; Vella, S.; Riganti, C.; Picci, P.; Serra, M. Doxorubicin-resistant osteosarcoma: Novel therapeutic approaches in sight? Future Oncol. 2017, 13, 673–677. [Google Scholar] [CrossRef]
- Von Hoff, D.D.; Layard, M.W.; Basa, P.; Davis, H.L., Jr.; Von Hoff, A.L.; Rozencweig, M.; Muggia, F.M. Risk factors for doxorubicin-induced congestive heart failure. Ann. Intern. Med. 1979, 91, 710–717. [Google Scholar] [CrossRef]
- Mohr, A.; Marques Da Costa, M.E.; Fromigue, O.; Audinot, B.; Balde, T.; Droit, R.; Abbou, S.; Khneisser, P.; Berlanga, P.; Perez, E.; et al. From biology to personalized medicine: Recent knowledge in osteosarcoma. Eur. J. Med. Genet. 2024, 69, 104941. [Google Scholar] [CrossRef]
- Liang, H.; Cui, M.; Tu, J.; Chen, X. Advancements in osteosarcoma management: Integrating immune microenvironment insights with immunotherapeutic strategies. Front. Cell Dev. Biol. 2024, 12, 1394339. [Google Scholar] [CrossRef]
- Yu, S.; Yao, X. Advances on immunotherapy for osteosarcoma. Mol. Cancer 2024, 23, 192. [Google Scholar] [CrossRef] [PubMed]
- Jaén, M.; Martín-Regalado, Á.; Bartolomé, R.A.; Robles, J.; Casal, J.I. Interleukin 13 receptor alpha 2 (IL13Rα2): Expression, signaling pathways and therapeutic applications in cancer. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188802. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Bak, Y.; Oh, J.W.; Hong, J.; Lee, S.; Hong, J.T.; Yoon, D.Y. Inhibition of IL-13 and IL-13Rα2 Expression by IL-32θ in Human Monocytic Cells Requires PKCδ and STAT3 Association. Int. J. Mol. Sci. 2019, 20, 1949. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, P.; Lundberg, P.; Persson, E.; Johansson, A.; Lundgren, I.; Lie, A.; Conaway, H.H.; Lerner, U.H. Inhibition of hormone and cytokine-stimulated osteoclastogenesis and bone resorption by interleukin-4 and interleukin-13 is associated with increased osteoprotegerin and decreased RANKL and RANK in a STAT6-dependent pathway. J. Biol. Chem. 2006, 281, 2414–2429. [Google Scholar] [CrossRef] [PubMed]
- Khalmuratova, R.; Shin, H.W.; Kim, D.W.; Park, J.W. Interleukin (IL)-13 and IL-17A contribute to neo-osteogenesis in chronic rhinosinusitis by inducing RUNX2. EBioMedicine 2019, 46, 330–341. [Google Scholar] [CrossRef]
- Xu, W.; Chao, R.; Xie, X.; Mao, Y.; Chen, X.; Chen, X.; Zhang, S. IL13Ralpha2 as a crucial receptor for Chi3l1 in osteoclast differentiation and bone resorption through the MAPK/AKT pathway. Cell Commun. Signal. 2024, 22, 81. [Google Scholar] [CrossRef]
- Lin, C.; Liu, H.; Zhang, H.; He, H.; Li, H.; Shen, Z.; Qin, J.; Qin, X.; Xu, J.; Sun, Y. Interleukin-13 receptor alpha2 is associated with poor prognosis in patients with gastric cancer after gastrectomy. Oncotarget 2016, 7, 49281–49288. [Google Scholar] [CrossRef]
- Kwon, H.J.; Choi, J.E.; Bae, Y.K. Interleukin-13 receptor alpha 2 expression in tumor cells is associated with reduced disease-free survival in patients with luminal subtype invasive breast cancer. Tumour Biol. 2018, 40, 1010428318783657. [Google Scholar] [CrossRef]
- Han, J.; Puri, R.K. Analysis of the cancer genome atlas (TCGA) database identifies an inverse relationship between interleukin-13 receptor α1 and α2 gene expression and poor prognosis and drug resistance in subjects with glioblastoma multiforme. J. Neurooncol. 2018, 136, 463–474. [Google Scholar] [CrossRef]
- Kumar, A.; Bellayr, I.H.; Singh, H.S.; Puri, R.K. IL-13Rα2 gene expression is a biomarker of adverse outcome in patients with adrenocortical carcinoma. PLoS ONE 2021, 16, e0246632. [Google Scholar] [CrossRef]
- Roberts, A.B.; Wakefield, L.M. The two faces of transforming growth factor beta in carcinogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 8621–8623. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Joshi, B.; Nakajima, A.; Puri, R.K. A novel role of interleukin-13 receptor alpha2 in pancreatic cancer invasion and metastasis. Cancer Res. 2009, 69, 8678–8685. [Google Scholar] [CrossRef] [PubMed]
- Jaen, M.; Bartolome, R.A.; Aizpurua, C.; Martin-Regalado, A.; Imbaud, J.I.; Casal, J.I. Inhibition of Liver Metastasis in Colorectal Cancer by Targeting IL-13/IL13Ralpha2 Binding Site with Specific Monoclonal Antibodies. Cancers 2021, 13, 1731. [Google Scholar] [CrossRef]
- Ni, M.; Qin, B.; Xie, L.; Zhang, X.; Yang, J.; Lv, H.; Yang, M.; Zhang, M. IL-13 Contributes to Drug Resistance of NK/T-Cell Lymphoma Cells by Regulating ABCC4. Biomed. Res. Int. 2018, 2018, 2606834. [Google Scholar] [CrossRef]
- Jou, E.; Rodriguez-Rodriguez, N.; Ferreira, A.-C.F.; Jolin, H.E.; Clark, P.A.; Sawmynaden, K.; Ko, M.; Murphy, J.E.; Mannion, J.; Ward, C. An innate IL-25–ILC2–MDSC axis creates a cancer-permissive microenvironment for Apc mutation–driven intestinal tumorigenesis. Sci. Immunol. 2022, 7, eabn0175. [Google Scholar] [CrossRef]
- Cristiani, C.M.; Capone, M.; Garofalo, C.; Madonna, G.; Mallardo, D.; Tuffanelli, M.; Vanella, V.; Greco, M.; Foti, D.P.; Viglietto, G.; et al. Altered Frequencies and Functions of Innate Lymphoid Cells in Melanoma Patients Are Modulated by Immune Checkpoints Inhibitors. Front. Immunol. 2022, 13, 811131. [Google Scholar] [CrossRef]
- Ducarouge, B.; Redavid, A.R.; Victoor, C.; Chira, R.; Fonseca, A.; Hervieu, M.; Berge, R.; Lengrand, J.; Vieugue, P.; Neves, D.; et al. Netrin-1 blockade inhibits tumor associated Myeloid-derived suppressor cells, cancer stemness and alleviates resistance to chemotherapy and immune checkpoint inhibitor. Cell Death Differ. 2023, 30, 2201–2212. [Google Scholar] [CrossRef]
- Nagai, T.; Terada, N.; Fujii, M.; Nagata, Y.; Nakahara, K.; Mukai, S.; Okasho, K.; Kamiyama, Y.; Akamatsu, S.; Kobayashi, T. Identification of the α2 chain of interleukin-13 receptor as a potential biomarker for predicting castration resistance of prostate cancer using patient-derived xenograft models. Cancer Rep. 2023, 6, e1701. [Google Scholar] [CrossRef]
- Jiramongkol, Y.; Lam, E.W. FOXO transcription factor family in cancer and metastasis. Cancer Metastasis Rev. 2020, 39, 681–709. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, J.; Kang, M.A.; Jang, K.Y.; Kim, J.R. Mitoxantrone induces apoptosis in osteosarcoma cells through regulation of the Akt/FOXO3 pathway. Oncol. Lett. 2018, 15, 9687–9696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ha, S.H.; Moon, Y.J.; Hussein, U.K.; Song, Y.; Kim, K.M.; Park, S.H.; Park, H.S.; Park, B.H.; Ahn, A.R.; et al. Inhibition of SIRT6 potentiates the anti-tumor effect of doxorubicin through suppression of the DNA damage repair pathway in osteosarcoma. J. Exp. Clin. Cancer Res. 2020, 39, 247. [Google Scholar] [CrossRef] [PubMed]
- Mrowczynski, O.D.; Payne, R.A.; Bourcier, A.J.; Mau, C.Y.; Slagle-Webb, B.; Shenoy, G.; Madhankumar, A.B.; Abramson, S.B.; Wolfe, D.; Harbaugh, K.S.; et al. Targeting IL-13Ralpha2 for effective treatment of malignant peripheral nerve sheath tumors in mouse models. J. Neurosurg. 2019, 131, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Knudson, K.M.; Hwang, S.; McCann, M.S.; Joshi, B.H.; Husain, S.R.; Puri, R.K. Recent Advances in IL-13Rα2-Directed Cancer Immunotherapy. Front. Immunol. 2022, 13, 878365. [Google Scholar] [CrossRef]
- Balyasnikova, I.V.; Wainwright, D.A.; Solomaha, E.; Lee, G.; Han, Y.; Thaci, B.; Lesniak, M.S. Characterization and immunotherapeutic implications for a novel antibody targeting interleukin (IL)-13 receptor alpha2. J. Biol. Chem. 2012, 287, 30215–30227. [Google Scholar] [CrossRef]
- Allred, D.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar]
- Zhang, J.; Song, Y.; Ahn, A.-R.; Park, H.S.; Park, S.-H.; Moon, Y.J.; Kim, K.M.; Jang, K.Y. PAK4 Is Involved in the Stabilization of PD-L1 and the Resistance to Doxorubicin in Osteosarcoma and Predicts the Survival of Diagnosed Patients. Cells 2024, 13, 1444. [Google Scholar] [CrossRef]
- Maurya, D.K. ColonyCountJ: A user-friendly image J Add-on program for quantification of different colony parameters in clonogenic assay. J. Clin. Toxicol. 2017, 7, 4. [Google Scholar]
- O’Connell, M.R.; Sarkar, S.; Luthra, G.K.; Okugawa, Y.; Toiyama, Y.; Gajjar, A.H.; Qiu, S.; Goel, A.; Singh, P. Epigenetic changes and alternate promoter usage by human colon cancers for expressing DCLK1-isoforms: Clinical Implications. Sci. Rep. 2015, 5, 14983. [Google Scholar] [CrossRef]
- Vincenzi, F.; Rotondo, J.C.; Pasquini, S.; Di Virgilio, F.; Varani, K.; Tognon, M. A3 adenosine and P2X7 purinergic receptors as new targets for an innovative pharmacological therapy of malignant pleural mesothelioma. Front. Oncol. 2021, 11, 679285. [Google Scholar] [CrossRef]
- DeLong, E.R.; DeLong, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef]
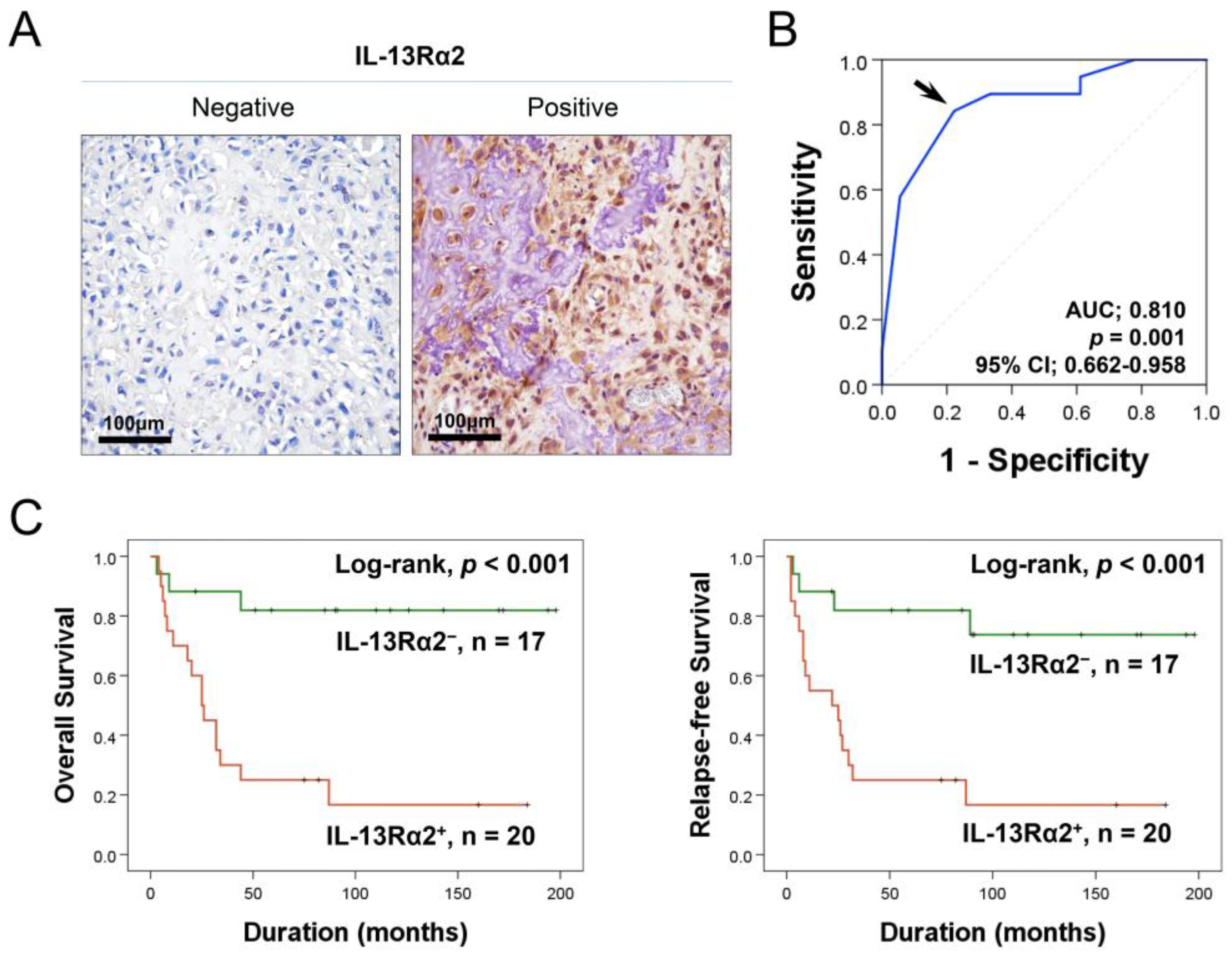
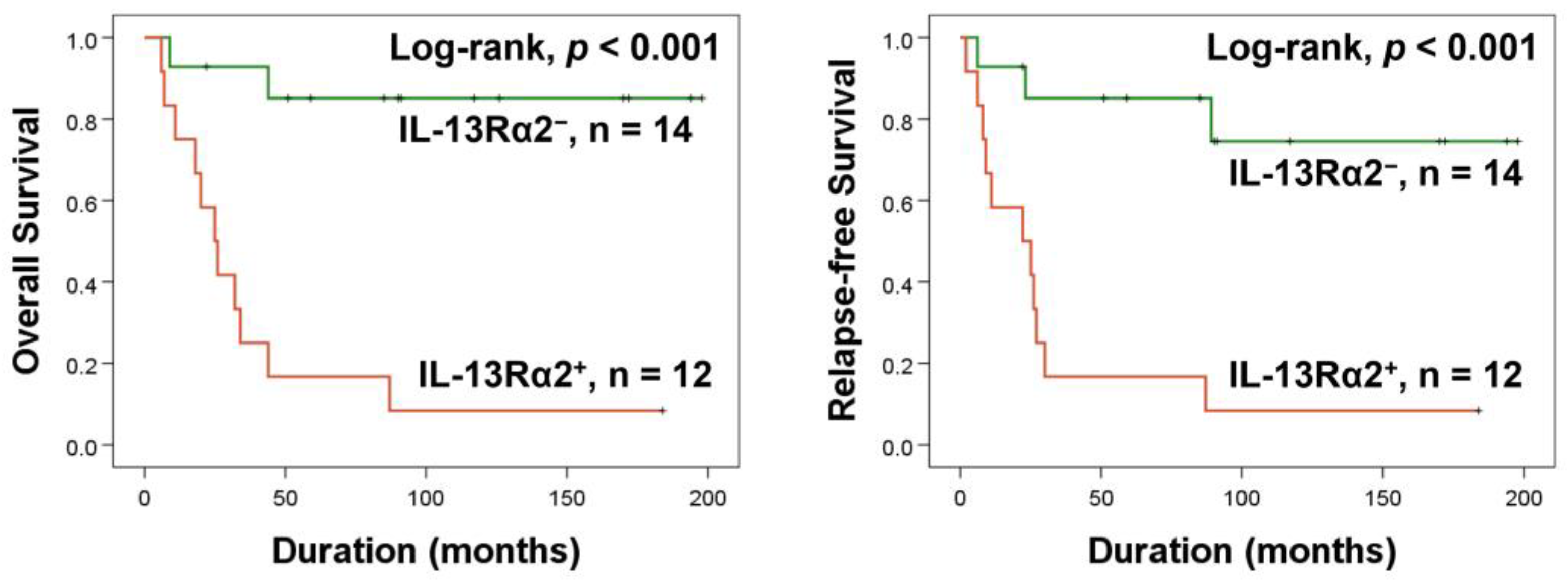
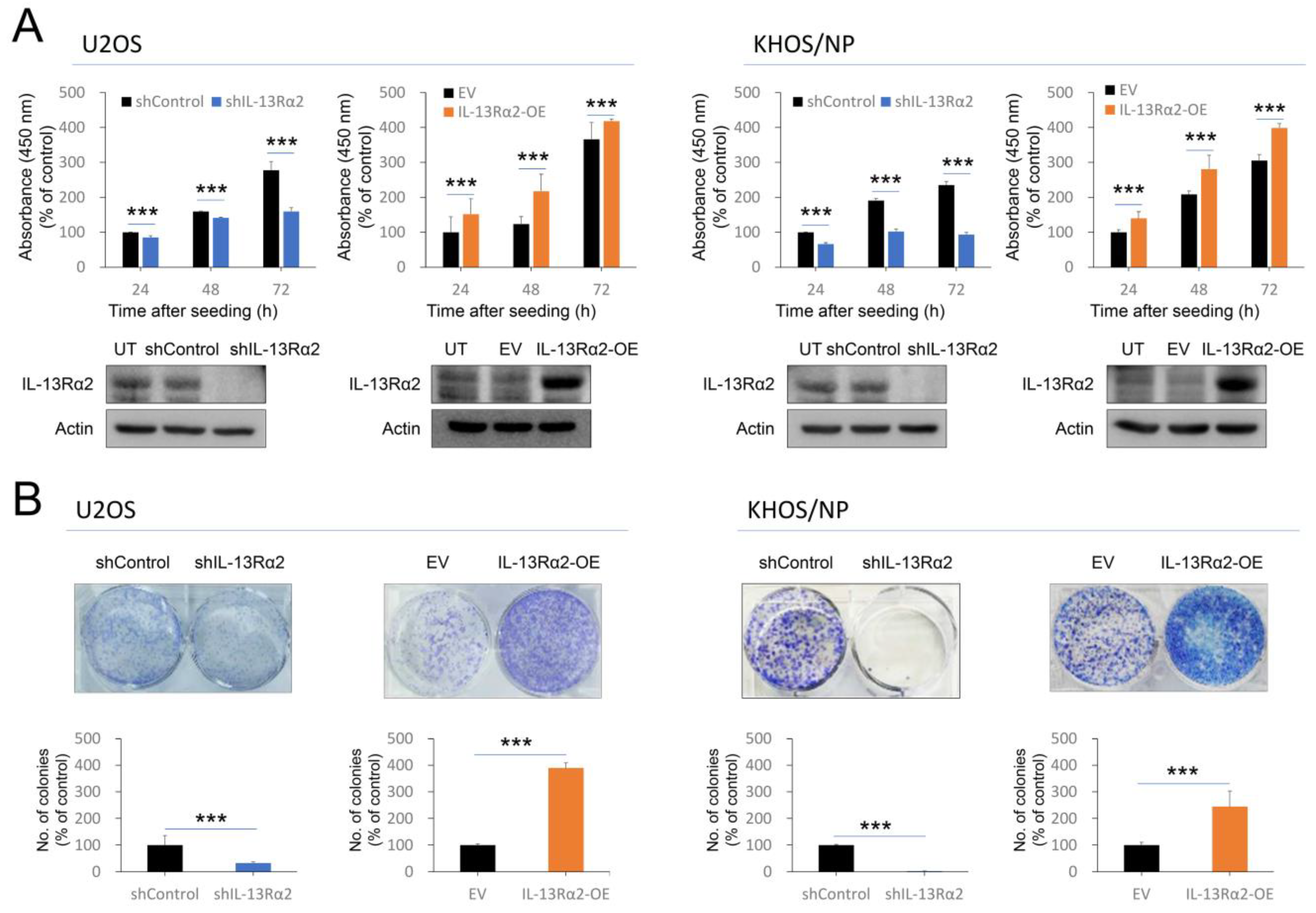
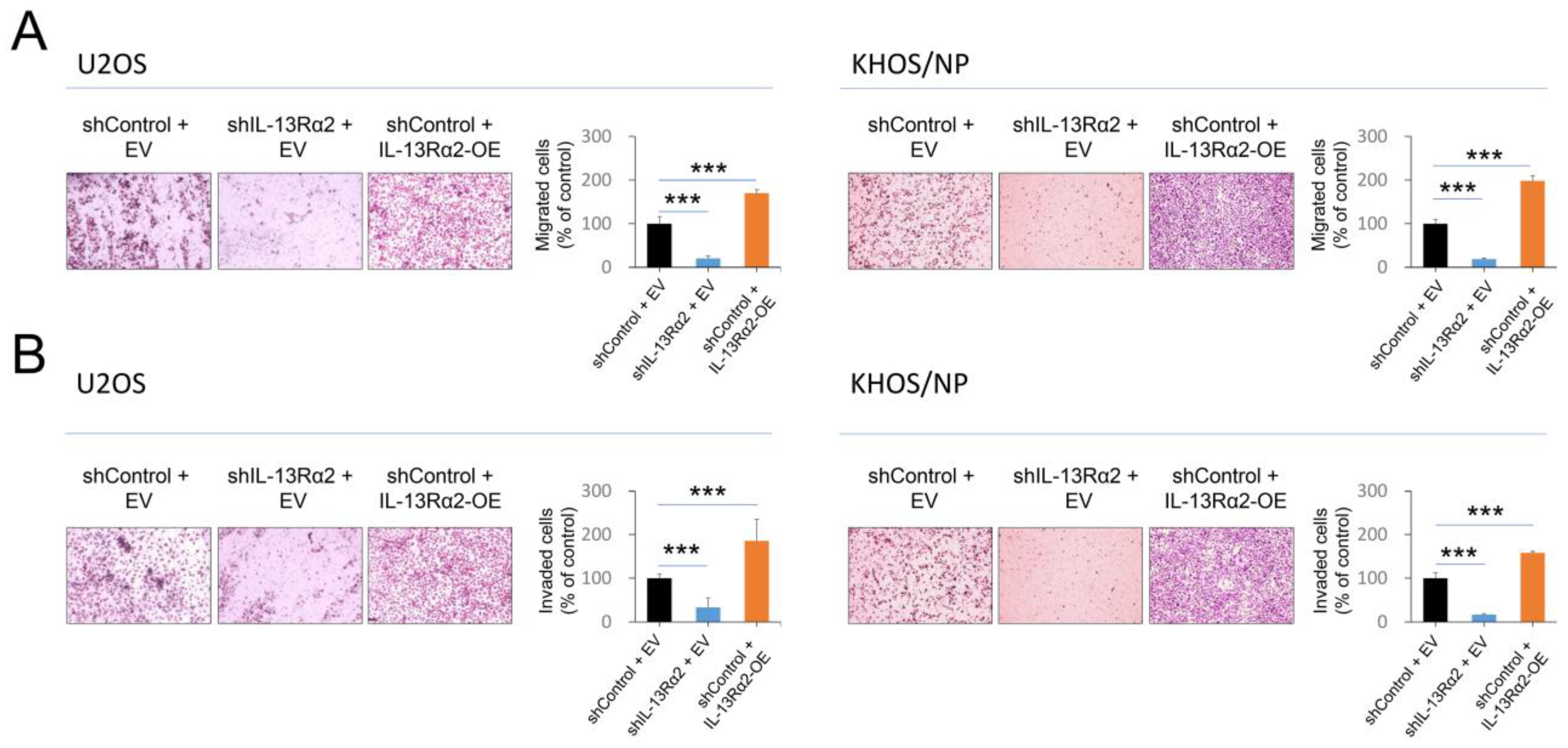

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | No. | IL-13Rα2 | ||
|---|---|---|---|---|
| Positive | p * | |||
| Age, years | <30 | 24 | 12 (50%) | 0.501 |
| ≥30 | 13 | 8 (62%) | ||
| Sex | Male | 25 | 17 (68%) | 0.014 |
| Female | 12 | 3 (25%) | ||
| Tumor size | ≤8 cm | 19 | 8 (42%) | 0.134 |
| >8 cm | 18 | 12 (67%) | ||
| Stage | I and II | 26 | 12 (46%) | 0.138 |
| III and IV | 11 | 8 (73%) | ||
| T category | 1 | 17 | 7 (41%) | 0.316 |
| 2 | 16 | 10 (63%) | ||
| 3 and 4 | 4 | 3 (75%) | ||
| N category | N0 | 34 | 18 (53%) | 0.647 |
| N1 | 3 | 2 (67%) | ||
| M category | M0 | 29 | 14 (48%) | 0.179 |
| M1 | 8 | 6 (75%) |
| Characteristics | No. | OS | RFS | ||
|---|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | ||
| Age, years, ≥30 (vs. <30) | 13/37 | 2.593 (1.047–6.419) | 0.039 | 2.944 (1.217–7.123) | 0.017 |
| Sex, male (vs. female) | 25/37 | 0.833 (0.299–2.323) | 0.727 | 0.686 (0.248–1.899) | 0.468 |
| Tumor size, ≥8 cm (vs. <8 cm) | 18/37 | 3.319 (0.253–8.790) | 0.016 | 2.987 (1.179–7.564) | 0.021 |
| Stage, III and IV (vs. I and II) | 11/37 | 3.115 (1.236–7.853) | 0.016 | 3.652 (1.472–9.062) | 0.005 |
| T category, 1 | 17/37 | 1 | 0.059 | 1 | 0.020 |
| 2 | 16/37 | 3.309 (1.144–9.569) | 0.027 | 3.383 (1.168–9.801) | 0.025 |
| 3 and 4 | 4/37 | 3.998 (0.950–16.828) | 0.059 | 6.014 (1.589–22.760) | 0.008 |
| N category, N1 (vs. N0) | 3/37 | 4.031 (0.838–19.400) | 0.082 | 2.636 (0.571–12.177) | 0.214 |
| M category, M1 (vs. M0) | 8/37 | 3.812 (1.408–10.316) | 0.008 | 3.282 (1.206–8.926) | 0.020 |
| IL-13Rα2, positive (vs. negative) | 20/37 | 7.191 (2.063–25.067) | 0.002 | 5.668 (1.848–17.385) | 0.002 |
| Characteristics | OS | RFS | ||
|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | |
| Age, years, ≥30 (vs. <30) | 3.159 (1.267–7.873) | 0.014 | ||
| Stage, III and IV (vs. I and II) | 2.865 (1.130–7.262) | 0.027 | ||
| IL-13Rα2, positive (vs. negative) | 7.191 (2.063–25.067) | 0.002 | 6.161 (1.909–19.883) | 0.002 |
| Characteristics | No. | OS | RFS | |||
|---|---|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | |||
| Univariate analysis | ||||||
| IL-13Rα2, positive (vs. negative) | 12/26 | 11.985 (2.600–55.254) | 0.001 | 8.208 (2.203–30.583) | 0.002 | |
| Multivariate analysis | ||||||
| Stage, III and IV (vs. I and II) | 3.210 (1.052–9.789) | 0.040 | ||||
| IL-13Rα2, positive (vs. negative) | 11.985 (2.600–55.254) | 0.001 | 7.461 (1.946–28.605) | 0.003 | ||
| Gene | Primer Sequence | Accession Number |
|---|---|---|
| IL-13RA2 (IL-13Rα2) | Forward 5′-TGACGGAATTTGGAGTGAGTGG-3′ Reverse 5′-CCAAATGGTAGCCAGAAACGTAGC-3′ | NM_000640.2 |
| TGFB1 (TGF-β1) | Forward 5′-CCCACAACGAAATCTATGACAA-3′ Reverse 5′-AAGATAACCACTCTGGCGAGTG-3′ | NM_000660.7 |
| SNAL1 (snail) | Forward 5′-ACCCCACATCCTTCTCACTG-3′ Reverse 5′-TACAAAAACCCACGCAGACA-3′ | NM_005985.3 |
| CCND1 (cyclin D1) | Forward 5′-GAGGAAGAGGAGGAGGAGGA-3′ Reverse 5′-GAGATGGAAGGGGGAAAGAG-3′ | NM_053056.2 |
| BCL2 | Forward 5′-ATCGCCCTGTGGATGACTGAGT-3′ Reverse 5′-GCCAGGAGAAATCAAACAGAGGC-3′ | NM_000633.3 |
| BAX | Forward 5′-TGGCAGCTGACATGTTTTCTGAC-3′ Reverse 5′-TCACCCAACCACCCTGGTCTT-3′ | NM_138761.4 |
| GAPDH | Forward 5′-AACAGCGACACCCACTCCTC-3′ Reverse 5′-GGAGGGGAGATTCAGTGTGGT-3′ | NM_001256799.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karamikheirabad, M.; Zhang, J.; Ahn, A.-R.; Park, H.S.; Park, S.-H.; Moon, Y.J.; Kim, K.M.; Jang, K.Y. IL-13Rα2 Is Involved in Resistance to Doxorubicin and Survival of Osteosarcoma Patients. Pharmaceuticals 2024, 17, 1526. https://doi.org/10.3390/ph17111526
Karamikheirabad M, Zhang J, Ahn A-R, Park HS, Park S-H, Moon YJ, Kim KM, Jang KY. IL-13Rα2 Is Involved in Resistance to Doxorubicin and Survival of Osteosarcoma Patients. Pharmaceuticals. 2024; 17(11):1526. https://doi.org/10.3390/ph17111526
Chicago/Turabian StyleKaramikheirabad, Maryam, Junyue Zhang, Ae-Ri Ahn, Ho Sung Park, See-Hyoung Park, Young Jae Moon, Kyoung Min Kim, and Kyu Yun Jang. 2024. "IL-13Rα2 Is Involved in Resistance to Doxorubicin and Survival of Osteosarcoma Patients" Pharmaceuticals 17, no. 11: 1526. https://doi.org/10.3390/ph17111526
APA StyleKaramikheirabad, M., Zhang, J., Ahn, A.-R., Park, H. S., Park, S.-H., Moon, Y. J., Kim, K. M., & Jang, K. Y. (2024). IL-13Rα2 Is Involved in Resistance to Doxorubicin and Survival of Osteosarcoma Patients. Pharmaceuticals, 17(11), 1526. https://doi.org/10.3390/ph17111526