
Repurposing Terfenadine as a Novel Antigiardial Compound

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Terfenadine Inhibits Growth and Cell Viability of Giardia lamblia Trophozoites
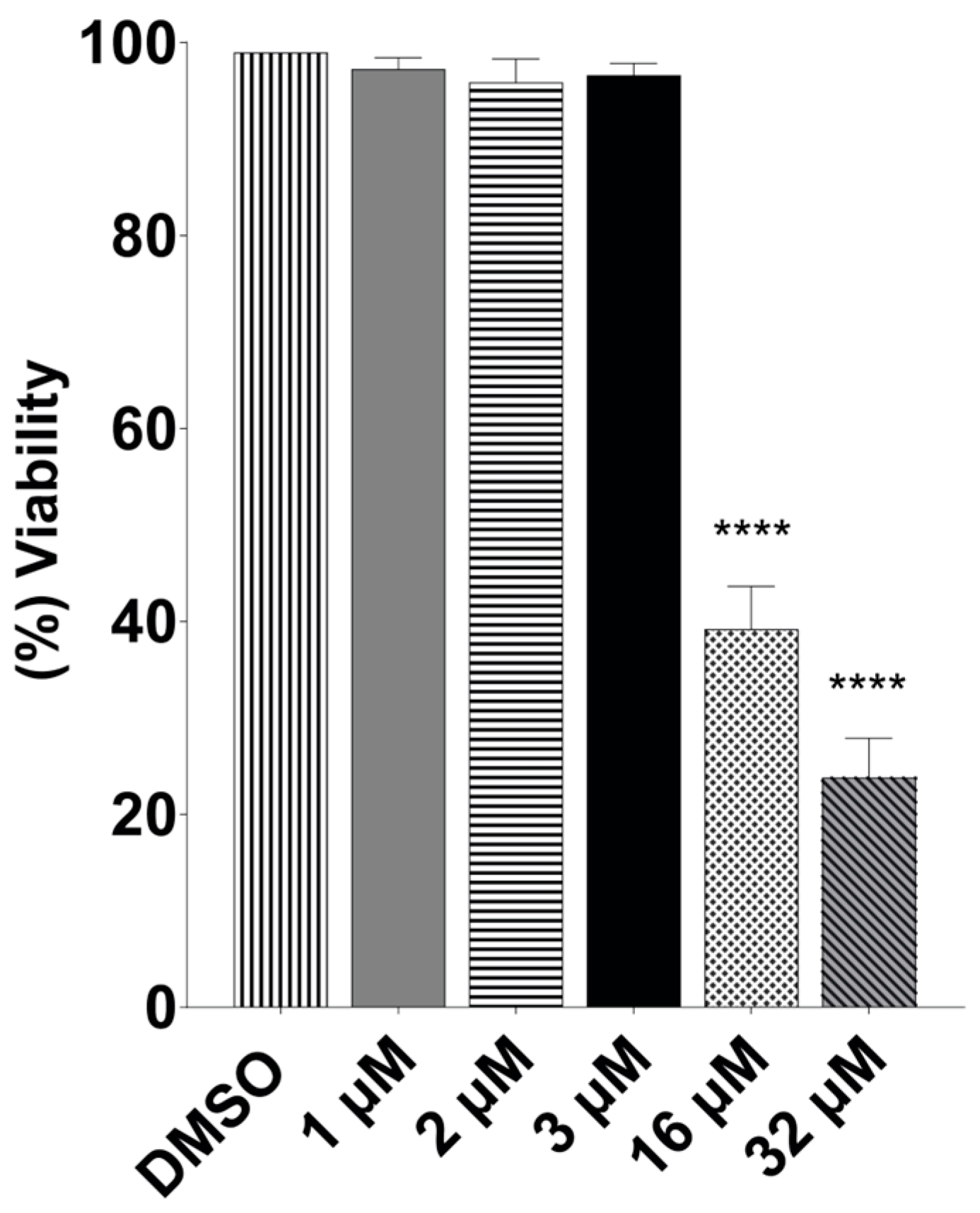
2.2. Terfenadine Did Not Affect Caco-2 Cells but Was Selective for Giardia
2.3. Terfenadine Inhibited Giardia lamblia Adhesion to Caco-2 Cells
2.4. Terfenadine Altered the Morphology of Giardia lamblia Trophozoites
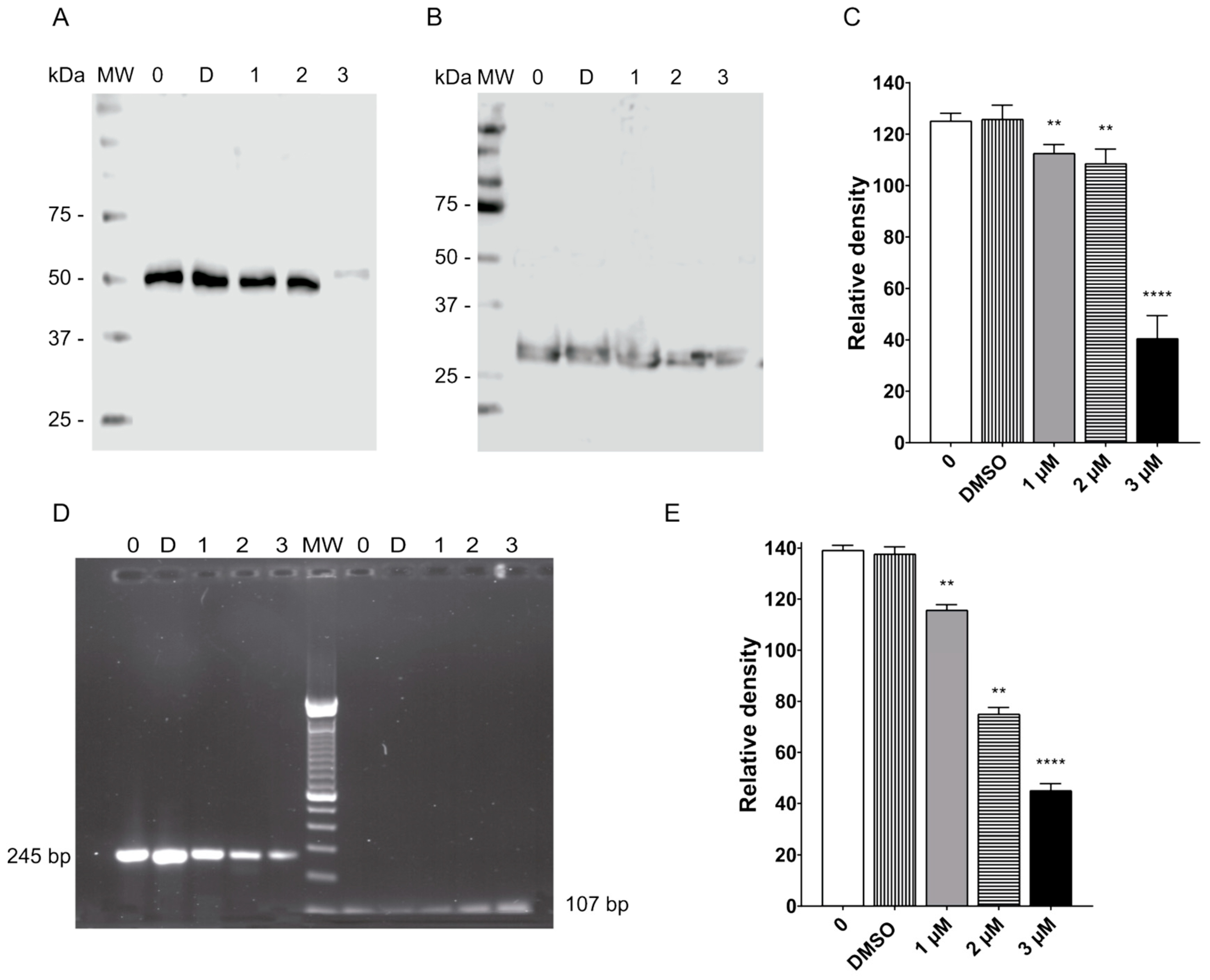
2.5. Morphological Changes by Terfenadine Involved Remodeling of Tubulin on Giardia lamblia Trophozoites
2.6. Terfenadine Caused Downreglation of Tubulin in Giardia lamblia Trophozoites
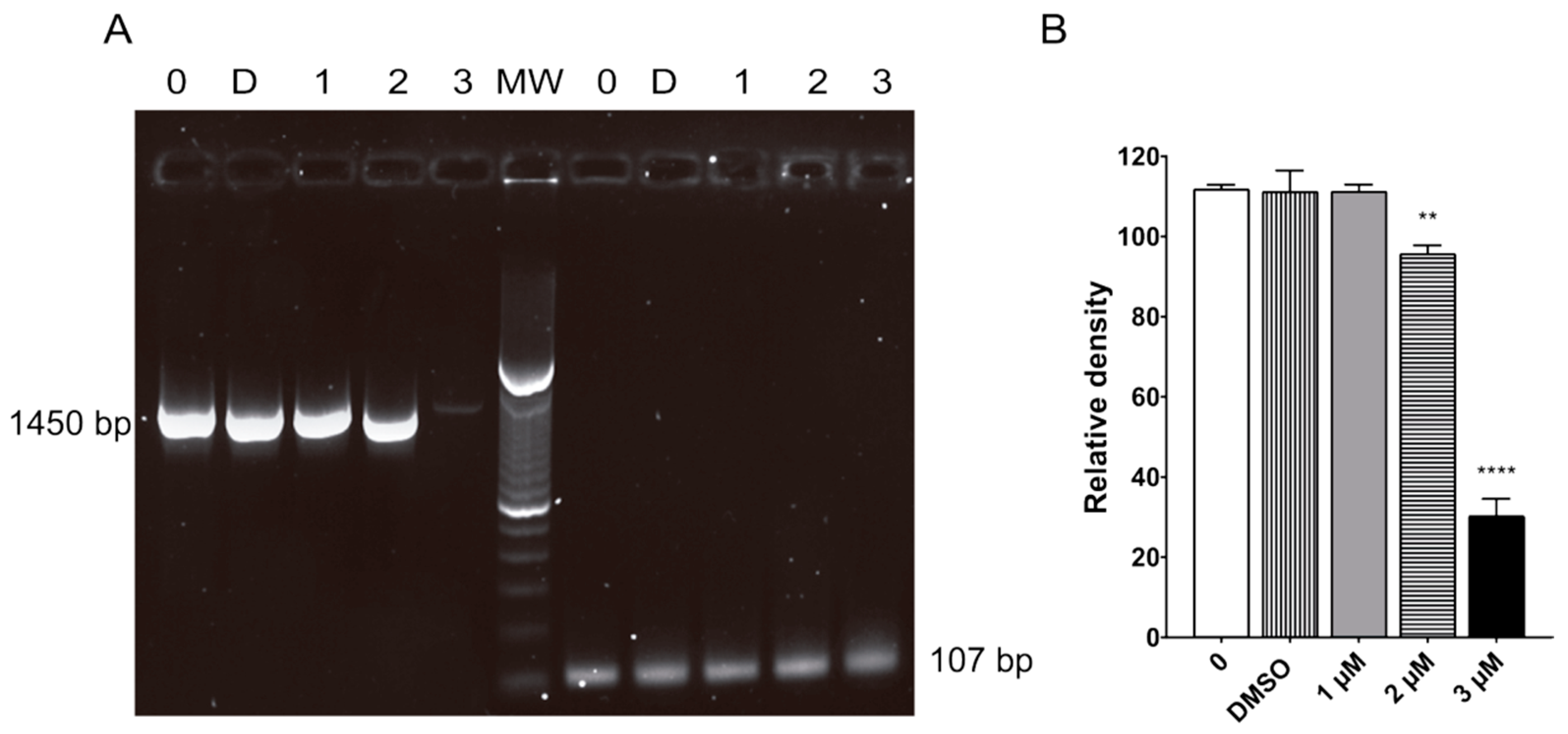
2.7. Terfenadine Induced Alterations in GiK Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture of Giardia lamblia Trophozoites and Caco-2 Cell Line
4.2. Growth Inhibition Assay
4.3. Terfenadine Cytotoxicity Assay
4.4. Selectivity Index Calculation
4.5. Cell Viability Assay
4.6. Cell Adhesion Assay
4.7. Scanning Electron Microscopy (SEM)
4.8. Immunofluorescence
4.9. SDS-PAGE and Western Blot Assay
4.9.1. Protein Extracts and SDS-PAGE
4.9.2. Western Blot
4.10. RT-PCR
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D-glucose | DIBICO 2001-E |
| NaCl | Sigma-Aldrich S7653 |
| K2HPO4 | Sigma-Aldrich P5655 |
| KH2PO4 | Sigma-Aldrich P5505 |
| Peptone | Sigma-Aldrich G8270 |
| L-ascorbic acid | Sigma-Aldrich A5960 |
| L-cysteine | Sigma-Aldrich C6852 |
| Ferric ammonium citrate | Sigma-Aldrich F5879 |
| Dehydrated bovine bile | Sigma-Aldrich B3883 |
Appendix B
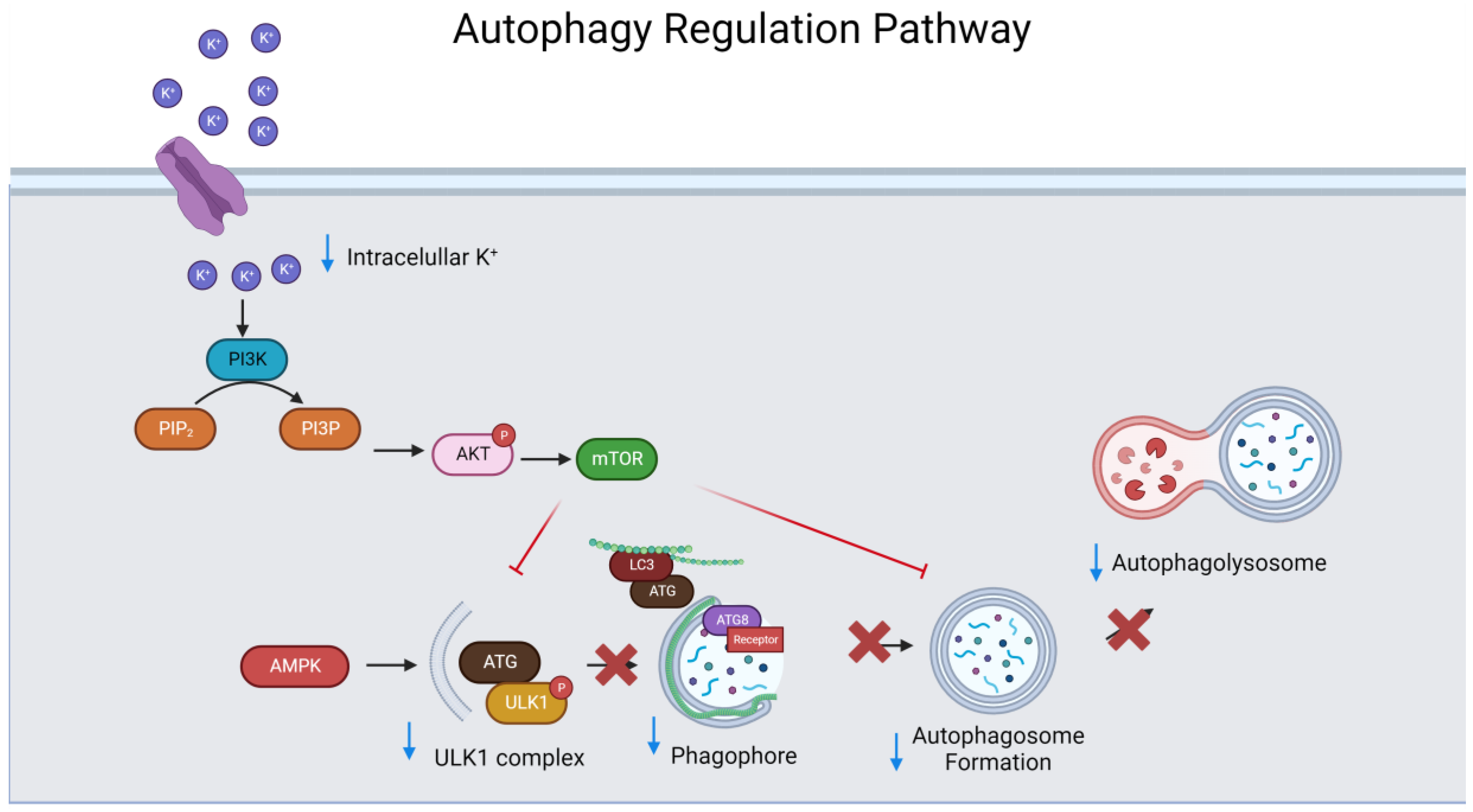
Appendix C
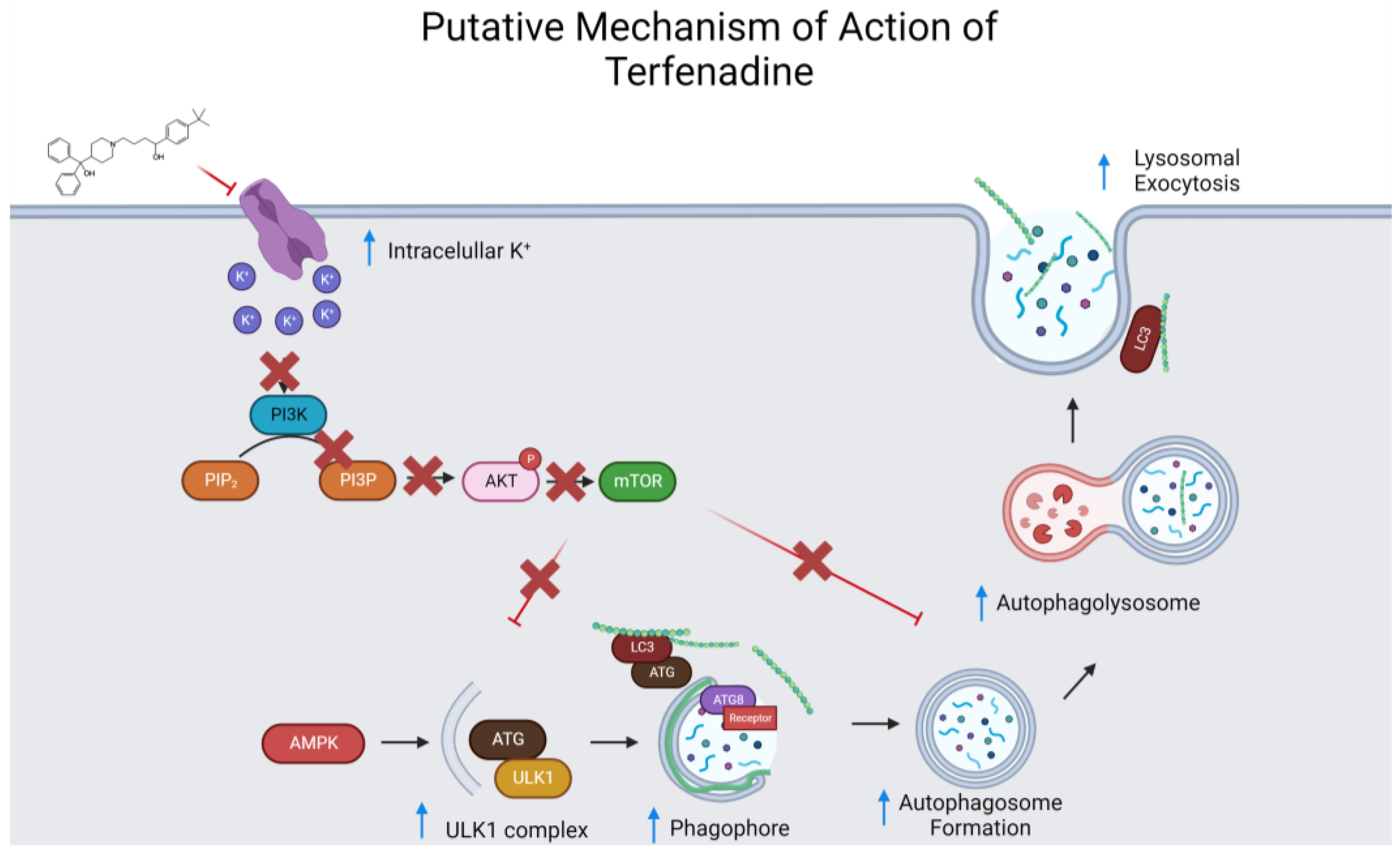
References
- Lalle, M.; Hanevik, K. Treatment-refractory giardiasis: Challenges and solutions. Infect. Drug Resist. 2018, 11, 1921–1933. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.C.; Leung, A.A.M.; Wong, A.H.C.; Sergi, C.M.; Kam, J.K.M. Giardiasis: An Overview. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Halliez, M.C.; Buret, A.G. Extra-intestinal and long term consequences of Giardia duodenalis infections. World J. Gastroenterol. 2013, 19, 8974–8985. [Google Scholar] [CrossRef] [PubMed]
- Painter, J.E.; Collier, S.A.; Gargano, J.W. Association between Giardia and arthritis or joint pain in a large health insurance cohort: Could it be reactive arthritis? Epidemiol. Infect. 2017, 145, 471–477. [Google Scholar] [CrossRef]
- Watkins, R.R.; Eckmann, L. Treatment of giardiasis: Current status and future directions. Curr. Infect. Dis. Rep. 2014, 16, 396. [Google Scholar] [CrossRef]
- Vivancos, V.; González-Alvarez, I.; Bermejo, M.; Gonzalez-Alvarez, M. Giardiasis: Characteristics, Pathogenesis and New Insights About Treatment. Curr. Top. Med. Chem. 2018, 18, 1287–1303. [Google Scholar] [CrossRef]
- Hernández Ceruelos, A.; Romero-Quezada, L.C.; Ruvalcaba Ledezma, J.C.; López Contreras, L. Therapeutic uses of metronidazole and its side effects: An update. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 397–401. [Google Scholar] [CrossRef]
- Piloiu, C.; Dumitrascu, D.L. Albendazole-Induced Liver Injury. Am. J. Ther. 2021, 28, e335–e340. [Google Scholar] [CrossRef]
- Tinidazole. In LiverTox: Clinical and Research Information on Drug-Induced Liver Injury; National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2020.
- Chen, Y.; Li, P.; Chen, T.; Liu, H.; Wang, P.; Dai, X.; Zou, Q. Ronidazole Is a Superior Prodrug to Metronidazole for Nitroreductase-Mediated Hepatocytes Ablation in Zebrafish Larvae. Zebrafish 2023, 20, 95–102. [Google Scholar] [CrossRef]
- Ansell, B.R.; McConville, M.J.; Ma’ayeh, S.Y.; Dagley, M.J.; Gasser, R.B.; Svärd, S.G.; Jex, A.R. Drug resistance in Giardia duodenalis. Biotechnol. Adv. 2015, 33, 888–901. [Google Scholar] [CrossRef]
- Argüello-García, R.; Leitsch, D.; Skinner-Adams, T.; Ortega-Pierres, M.G. Drug resistance in Giardia: Mechanisms and alternative treatments for Giardiasis. Adv. Parasitol. 2020, 107, 201–282. [Google Scholar] [CrossRef]
- Leitsch, D. Drug Resistance in the Microaerophilic Parasite Giardia lamblia. Curr. Trop. Med. Rep. 2015, 2, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Villanueva, A.; Rufino-Gonzalez, Y.; Mendez, S.T.; Torres-Arroyo, A.; Ponce-Macotela, M.; Martinez-Gordillo, M.N.; Reyes-Vivas, H.; Oria-Hernandez, J. Disulfiram as a novel inactivator of Giardia lamblia triosephosphate isomerase with antigiardial potential. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Maganda, V.Y.; Rangel-Castañeda, I.A.; Suárez-Rico, D.O.; Cortés-Zárate, R.; Hernández-Hernández, J.M.; Pérez-Rangel, A.; Chiquete-Félix, N.; León-Ávila, G.; González-Pozos, S.; Gaona-Bernal, J.; et al. Antigiardial Activity of Acetylsalicylic Acid Is Associated with Overexpression of HSP70 and Membrane Transporters. Pharmaceuticals 2020, 13, 440. [Google Scholar] [CrossRef] [PubMed]
- Tejman-Yarden, N.; Miyamoto, Y.; Leitsch, D.; Santini, J.; Debnath, A.; Gut, J.; McKerrow, J.H.; Reed, S.L.; Eckmann, L. A reprofiled drug, auranofin, is effective against metronidazole-resistant Giardia lamblia. Antimicrob. Agents Chemother. 2013, 57, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Mertens, R.T.; Gukathasan, S.; Arojojoye, A.S.; Olelewe, C.; Awuah, S.G. Next Generation Gold Drugs and Probes: Chemistry and Biomedical Applications. Chem. Rev. 2023, 123, 6612–6667. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Castañeda, I.A.; Castillo-Romero, A.; León-Ávila, G.; Zermeño-Ruiz, M.; Hernández-Hernández, J.M. Drug repositioning: Antiprotozoal activity of terfenadine against Entamoeba histolytica trophozoites. Parasitol. Res. 2021, 121, 303–309. [Google Scholar] [CrossRef]
- Palomo-Ligas, L.; Gutiérrez-Gutiérrez, F.; Ochoa-Maganda, V.Y.; Cortés-Zárate, R.; Charles-Niño, C.L.; Castillo-Romero, A. Identification of a novel potassium channel (GiK) as a potential drug target in Giardia lamblia: Computational descriptions of binding sites. PeerJ 2019, 7, e6430. [Google Scholar] [CrossRef]
- Information NCoB. Terfenadine Pubchem: NCBI. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Terfenadine (accessed on 15 September 2023).
- Mørch, K.; Hanevik, K. Giardiasis treatment: An update with a focus on refractory disease. Curr. Opin. Infect. Dis. 2020, 33, 355–364. [Google Scholar] [CrossRef]
- Tejman-Yarden, N.; Eckmann, L. New approaches to the treatment of giardiasis. Curr. Opin. Infect. Dis. 2011, 24, 451–456. [Google Scholar] [CrossRef]
- Ehrenkaufer, G.; Li, P.; Stebbins, E.E.; Kangussu-Marcolino, M.M.; Debnath, A.; White, C.V.; Moser, M.S.; DeRisi, J.; Gisselberg, J.; Yeh, E.; et al. Identification of anisomycin, prodigiosin and obatoclax as compounds with broad-spectrum anti-parasitic activity. PLoS Negl. Trop. Dis. 2020, 14, e0008150. [Google Scholar] [CrossRef]
- Roy, M.; Dumaine, R.; Brown, A.M. HERG, a primary human ventricular target of the nonsedating antihistamine terfenadine. Circulation 1996, 94, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.; Maurer, M.; Taglialatela, M.; Church, M.K. Cardiac safety of second-generation H(1) -antihistamines when updosed in chronic spontaneous urticaria. Clin. Exp. Allergy 2019, 49, 1615–1623. [Google Scholar] [CrossRef]
- Honig, P.K.; Wortham, D.C.; Zamani, K.; Conner, D.P.; Mullin, J.C.; Cantilena, L.R. Terfenadine-ketoconazole interaction. Pharmacokinetic and electrocardiographic consequences. JAMA 1993, 269, 1513–1518. [Google Scholar] [CrossRef]
- Gibson, J.P.; Huffmann, K.W.; Newberne, J.W. Preclinical safety studies with terfenadine. Arzneimittelforschung 1982, 32, 1179–1184. [Google Scholar] [PubMed]
- Simons, F.E.; Watson, W.T.; Simons, K.J. The pharmacokinetics and pharmacodynamics of terfenadine in children. J. Allergy Clin. Immunol. 1987, 80, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Ngo, E.; Spigset, O.; Lupattelli, A.; Panchaud, A.; Annaert, P.; Allegaert, K.; Nordeng, H. Antihistamine use during breastfeeding with focus on breast milk transfer and safety in humans: A systematic literature review. Basic Clin. Pharmacol. Toxicol. 2022, 130, 171–181. [Google Scholar] [CrossRef]
- Singh, N.; Puri, S.K. Causal prophylactic activity of antihistaminic agents against Plasmodium yoelii nigeriensis infection in Swiss mice. Acta Trop. 1998, 69, 255–260. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef]
- Certad, G.; Viscogliosi, E.; Chabé, M.; Cacciò, S.M. Pathogenic Mechanisms of Cryptosporidium and Giardia. Trends Parasitol. 2017, 33, 561–576. [Google Scholar] [CrossRef]
- Benchimol, M.; Gadelha, A.P.; de Souza, W. Unusual Cell Structures and Organelles in Giardia intestinalis and Trichomonas vaginalis Are Potential Drug Targets. Microorganisms 2022, 10, 2176. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.V.F.; de Souza, W.; Vögerl, K.; Bracher, F.; Benchimol, M.; Gadelha, A.P.R. In vitro effects of the 4-[(10H-phenothiazin-10-yl)methyl]-N-hydroxybenzamide on Giardia intestinalis trophozoites. Acta Trop. 2022, 232, 106484. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Diaz, H.; Canizalez-Roman, A.; Nepomuceno-Mejia, T.; Gallardo-Vera, F.; Hornelas-Orozco, Y.; Nazmi, K.; Bolscher, J.G.; Carrero, J.C.; Leon-Sicairos, C.; Leon-Sicairos, N. Parasiticidal effect of synthetic bovine lactoferrin peptides on the enteric parasite Giardia intestinalis. Biochem. Cell Biol. 2017, 95, 82–90. [Google Scholar] [CrossRef]
- Choi, S.; Houdek, X.; Anderson, R.A. Phosphoinositide 3-kinase pathways and autophagy require phosphatidylinositol phosphate kinases. Adv. Biol. Regul. 2018, 68, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, S.; Oniku, A.E.; Topping, K.; Mamhoud, Z.N.; Paget, T.A. Programmed cell death in Giardia. Parasitology 2012, 139, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Abuammar, H.; Bhattacharjee, A.; Simon-Vecsei, Z.; Blastyák, A.; Csordás, G.; Páli, T.; Juhász, G. Ion Channels and Pumps in Autophagy: A Reciprocal Relationship. Cells 2021, 10, 3537. [Google Scholar] [CrossRef]
- Nicolau-Galmés, F.; Asumendi, A.; Alonso-Tejerina, E.; Pérez-Yarza, G.; Jangi, S.M.; Gardeazabal, J.; Arroyo-Berdugo, Y.; Careaga, J.M.; Díaz-Ramón, J.L.; Apraiz, A.; et al. Terfenadine induces apoptosis and autophagy in melanoma cells through ROS-dependent and -independent mechanisms. Apoptosis 2011, 16, 1253–1267. [Google Scholar] [CrossRef]
- Yichoy, M.; Duarte, T.T.; De Chatterjee, A.; Mendez, T.L.; Aguilera, K.Y.; Roy, D.; Roychowdhury, S.; Aley, S.B.; Das, S. Lipid metabolism in Giardia: A post-genomic perspective. Parasitology 2011, 138, 267–278. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Ali, S.M.; Kim, D.H.; Guertin, D.A.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton. Curr. Biol. 2004, 14, 1296–1302. [Google Scholar] [CrossRef]
- Mackeh, R.; Perdiz, D.; Lorin, S.; Codogno, P.; Poüs, C. Autophagy and microtubules—New story, old players. J. Cell Sci. 2013, 126, 1071–1080. [Google Scholar] [CrossRef]
- Trisciuoglio, D.; Degrassi, F. The Tubulin Code and Tubulin-Modifying Enzymes in Autophagy and Cancer. Cancers 2021, 14, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, L. Recent progress in autophagic lysosome reformation. Traffic 2017, 18, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Popova, J.S.; Greene, A.K.; Wang, J.; Rasenick, M.M. Phosphatidylinositol 4,5-bisphosphate modifies tubulin participation in phospholipase Cbeta1 signaling. J. Neurosci. 2002, 22, 1668–1678. [Google Scholar] [CrossRef]
- Melgari, D.; Barbier, C.; Dilanian, G.; Rücker-Martin, C.; Doisne, N.; Coulombe, A.; Hatem, S.N.; Balse, E. Microtubule polymerization state and clathrin-dependent internalization regulate dynamics of cardiac potassium channel: Microtubule and clathrin control of K(V)1.5 channel. J. Mol. Cell. Cardiol. 2020, 144, 127–139. [Google Scholar] [CrossRef]
- Camacho, J.; Sánchez, A.; Stühmer, W.; Pardo, L.A. Cytoskeletal interactions determine the electrophysiological properties of human EAG potassium channels. Pflug. Arch. 2000, 441, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Vitre, B.; Coquelle, F.M.; Heichette, C.; Garnier, C.; Chrétien, D.; Arnal, I. EB1 regulates microtubule dynamics and tubulin sheet closure in vitro. Nat. Cell Biol. 2008, 10, 415–421. [Google Scholar] [CrossRef]
- Keister, D.B. Axenic culture of Giardia lamblia in TYI-S-33 medium supplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 487–488. [Google Scholar] [CrossRef]
- Bastidas, O. Cell Counting with Neubauer Chamber, Basic Hemocytometer Usage; Technical Note-Neubauer Chamber Cell Counting; Celeromics: Valencia, Spain, 2013; pp. 1–6. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Krzywik, J.; Mozga, W.; Aminpour, M.; Janczak, J.; Maj, E.; Wietrzyk, J.; Tuszyński, J.A.; Huczyński, A. Synthesis, Antiproliferative Activity and Molecular Docking Studies of Novel Doubly Modified Colchicine Amides and Sulfonamides as Anticancer Agents. Molecules 2020, 25, 1789. [Google Scholar] [CrossRef]
- Rigothier, M.C.; Coconnier, M.H.; Servin, A.L.; Gayral, P. A new in vitro model of Entamoeba histolytica adhesion, using the human colon carcinoma cell line Caco-2: Scanning electron microscopic study. Infect. Immun. 1991, 59, 4142–4146. [Google Scholar] [CrossRef]
- Palomo-Ligas, L.; Estrada-Camacho, J.; Garza-Ontiveros, M.; Vargas-Villanueva, J.R.; Gutiérrez-Gutiérrez, F.; Nery-Flores, S.D.; Cañas Montoya, J.A.; Ascacio-Valdés, J.; Campos-Muzquiz, L.G.; Rodriguez-Herrera, R. Polyphenolic extract from Punica granatum peel causes cytoskeleton-related damage on Giardia lamblia trophozoites in vitro. PeerJ 2022, 10, e13350. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Haniu, H.; Komori, N. Determination of Protein Molecular Weights on SDS-PAGE. Methods Mol. Biol. 2019, 1855, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Ward, H.D.; Lev, B.I.; Kane, A.V.; Keusch, G.T.; Pereira, M.E. Identification and characterization of taglin, a mannose 6-phosphate binding, trypsin-activated lectin from Giardia lamblia. Biochemistry 1987, 26, 8669–8675. [Google Scholar] [CrossRef] [PubMed]
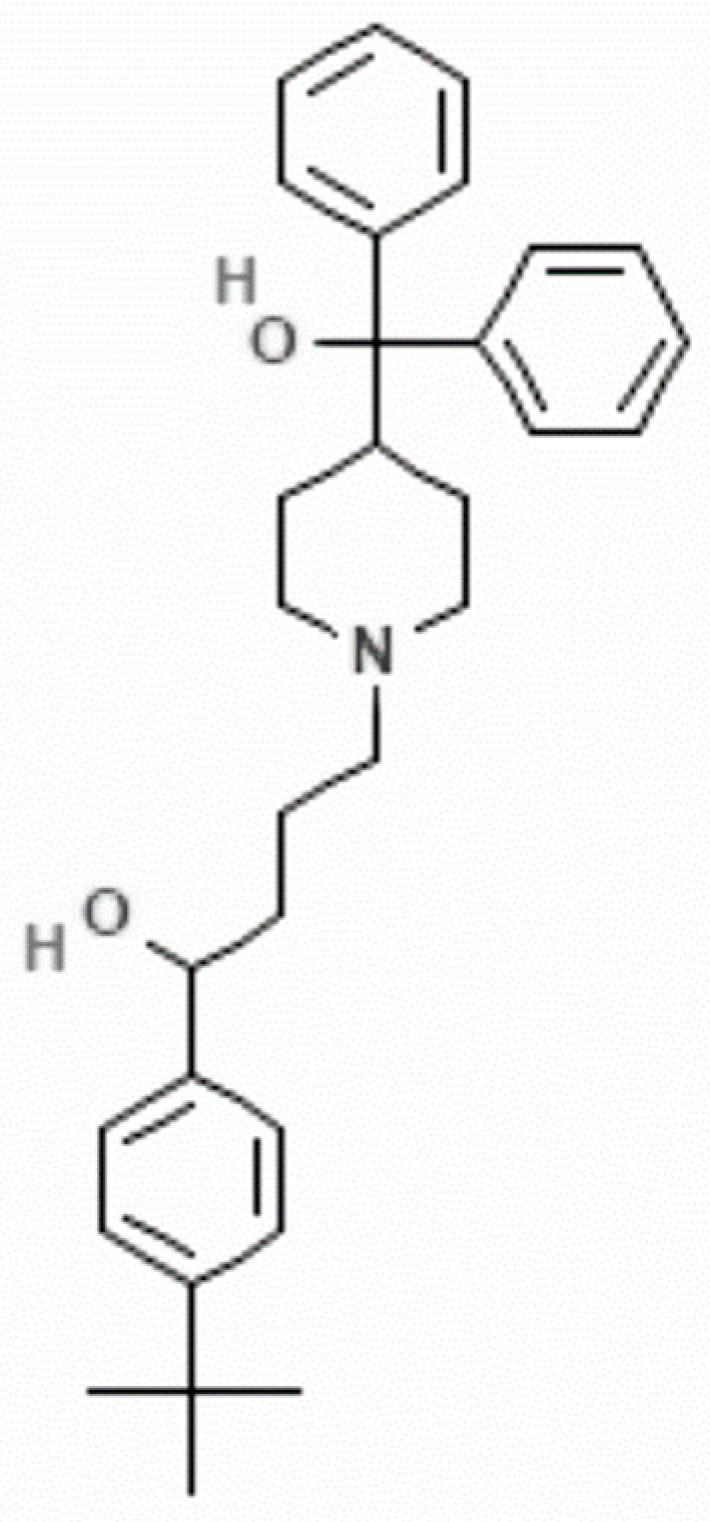




| IC50 on Giardia | CC50 on Caco-2 Cells | Selectivity Index CC50 on Caco-2 Cells/IC50 on Giardia |
|---|---|---|
| 1.6 μM | 16 μM | 10.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez-Rico, D.O.; Munguía-Huizar, F.J.; Cortés-Zárate, R.; Hernández-Hernández, J.M.; González-Pozos, S.; Perez-Rangel, A.; Castillo-Romero, A. Repurposing Terfenadine as a Novel Antigiardial Compound. Pharmaceuticals 2023, 16, 1332. https://doi.org/10.3390/ph16091332
Suárez-Rico DO, Munguía-Huizar FJ, Cortés-Zárate R, Hernández-Hernández JM, González-Pozos S, Perez-Rangel A, Castillo-Romero A. Repurposing Terfenadine as a Novel Antigiardial Compound. Pharmaceuticals. 2023; 16(9):1332. https://doi.org/10.3390/ph16091332
Chicago/Turabian StyleSuárez-Rico, Daniel Osmar, Francisco Javier Munguía-Huizar, Rafael Cortés-Zárate, José Manuel Hernández-Hernández, Sirenia González-Pozos, Armando Perez-Rangel, and Araceli Castillo-Romero. 2023. "Repurposing Terfenadine as a Novel Antigiardial Compound" Pharmaceuticals 16, no. 9: 1332. https://doi.org/10.3390/ph16091332
APA StyleSuárez-Rico, D. O., Munguía-Huizar, F. J., Cortés-Zárate, R., Hernández-Hernández, J. M., González-Pozos, S., Perez-Rangel, A., & Castillo-Romero, A. (2023). Repurposing Terfenadine as a Novel Antigiardial Compound. Pharmaceuticals, 16(9), 1332. https://doi.org/10.3390/ph16091332