

An Update on the Therapeutic Potential of Antimicrobial Peptides against Acinetobacter baumannii Infections


 and
and 
Abstract
1. Introduction

{kind=link}
{kind=link}
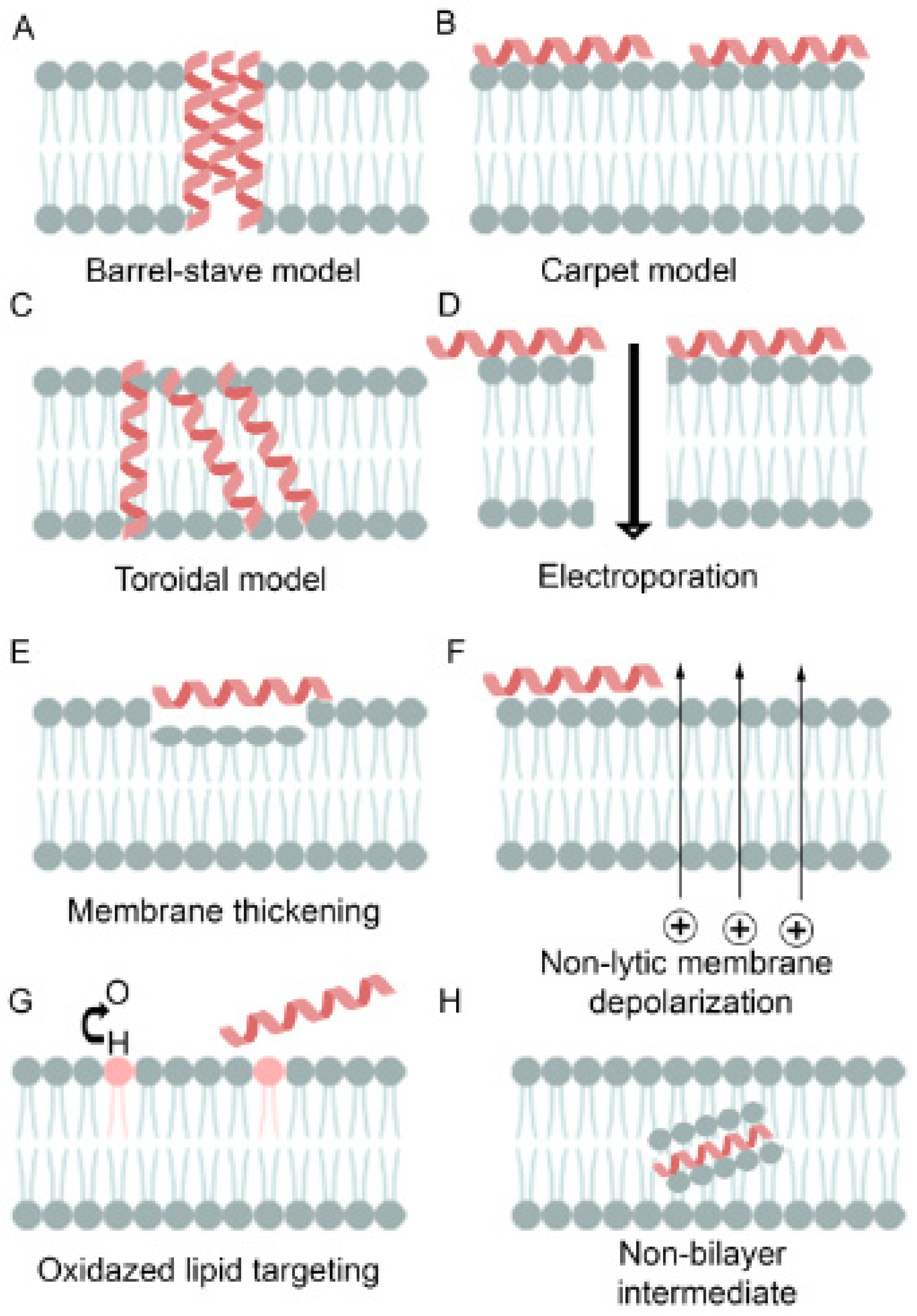{kind=link}
| Antibiotic Resistance Mechanism | Characteristics | Example | Ref. |
|---|---|---|---|
| Target modification or mutation | Mutation or modification of bacterial site will interfere with target matching, thus affecting the effect of antibiotics | Modifying PBPs in MRSA, production of β-lactamases or carbapenemases in genus Klebsiella; fluoroquinolone-resistant S. aureus Mycobacterium tuberculosis resistance to rifampicin is mainly caused by the mutation of the rpoB gene and vancomycin-resistant Enterococcus (VRE) | [29] |
| Reduced permeability | Deletion or damage of Omps is a source of bacterial resistance | Loss of porin D2 from outer cell wall in imipenem-resistant P. aeruginosa | [30] |
| Inactivating enzymes | Inactivating enzymes produced by bacteria, such as antibiotic hydrolases or similar enzymes, can hydrolyze or modify antibiotics inside the cell, rendering their inactivation before reaching the target site | Production of penicillin-inactivating β-lactamase by penicillin-resistant S. aureus, Haemophilus influenzae, and Escherichia coli bacteria, gentamicin-resistant enterococci via enzymatic inactivation of aminoglycosides and carbapenem-producing Enterobacteriaceae | [31] |
| Efflux pumps | Pumping of harmful molecules out of the bacterial cell | Increased efflux of tetracycline, macrolides, clindamycin, or fluoroquinolones in S. aureus | [32] |
| Metabolic enhancement or auxotrophy | Core genome mutations change metabolic pathways and induce antibiotic resistance | The genome of clinically pathogenic E. coli | [33,34] |
| Community cooperative resistance | Most bacteria coexist in communities, collectively resisting antibiotic effects; bacterial biofilms are efficiently protective of biofilm-forming bacterial species | P. aeruginosa, S. aureus, S maltophilia, and other bacteria | [35] |
| Target protective proteins (TPPs) | Bacterial synthetic protein protects antibiotic targets from antibiotics, eliminating their bacteriostatic effects | Clinically isolated S.aureus and other staphylococcus resistance to fusidic acid due to the level acquisition of genes encoding the FusB-type protein | [36] |
| Cell morphology changes | Modulating the body’s relative area via absorption efficiency changes can lead to the dilution of antibiotics entering the bacterial cell | Cells of the commonly used model organism Caulobacter crescentus | [37] |
| Self-repair systems | The multiple antibiotic resistance operon of enteric bacteria manipulates DNA repair and outer membrane integrity, enhancing antibiotic resistance | E. coli multiple antibiotic resistance (mar) loci was recognized as a determinant for cross-resistance to tetracyclines, quinolones, and β-lactams | [38] |
2. Antimicrobial Peptides
2.1. Cathelicidins
2.1.1. Humans
2.1.2. Snake
2.1.3. Alligator
2.1.4. Wallaby
2.1.5. Hoofed Animals
2.2. Defensins
2.2.1. Human α-Defensins
2.2.2. β-Defensins
2.2.3. Insect Defensins
2.3. Frog AMP
2.3.1. Magainin and Pexiganan
2.3.2. Brevinin-2 Related Peptide
2.3.3. Alyteserins
2.3.4. Peptide Glycine–Leucine-Amide
2.3.5. Caerulein Precursor Fragment
2.3.6. Hymenochirins
2.3.7. XT-7
2.3.8. Buforins
2.3.9. Caerin 1.1 and 1.9
2.3.10. Hylin a1
2.4. Fish Piscins
2.5. Hepcidin
2.6. Melittin
2.7. Cecropins
2.8. Mastoparan
2.9. Histatins
2.10. Dermcidin
2.11. Tachyplesin III
2.12. Spider Peptides
2.13. Scorpion
2.14. Lynronne-1
2.15. Hybrid Peptides
3. Resistance to AMPS
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistance Infections Globally: Final Report and Recommendations; The Review on Antimicrobial Resistance; Government of the United Kingdom: London, UK, 2016; 84p. [Google Scholar]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new. antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells, dormancy, and infectious disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Breidenstein, E.B.M.; Hancock, R.E.W. Importance of adaptive and stepwise changes in the rise and spread of antimicrobial resistance. In Antimicrobial Resistance in the Environment; Keen, P., Monforts, M., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; pp. 43–71. ISBN 978-1-118-15623-0. [Google Scholar]
- Olivares, J.; Bernardini, A.; Garcia-Leon, G.; Corona, F.; Sanchez, M.B.; Martinez, J.L. The intrinsic resistome of bacterial pathogens. Front. Microbiol. 2013, 30, 103. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.; Shan, Y. Persister awakening. Mol. Cell 2016, 63, 3–4. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; Clair, G.; Adkins, J.N.; Cheung, A.L.; Lewis, K. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat. Microbiol. 2016, 1, 16051. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Brown Gandt, A.; Rowe, S.E.; Deisinger, J.P.; Conlon, B.P.; Lewis, K. ATP-dependent persister formation in Escherichia coli. mBio 2017, 8, e02267-16. [Google Scholar] [CrossRef] [PubMed]
- Magana, M.; Sereti, C.; Ioannidis, A.; Mitchell, C.A.; Ball, A.R.; Magiorkinis, E.; Chatzipanagiotou, S.; Hamblin, M.R.; Hadjifrangiskou, M.; Tegos, G.P.; et al. Options and limitations in clinical investigation of bacterial biofilms. Clin. Microbiol. Rev. 2018, 31, e00084-16. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.R.; Shan, Y.; Zalis, E.A.; Isabella, V.; Lewis, K. A genetic determinant of persister cell formation in bacterial pathogens. J. Bacteriol. 2018, 200, e00303–e00318. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global, multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pan drug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Zhang, X.D.; Zhao, Q.; Peng, B.; Zheng, J. Analysis of global prevalence of antibiotic resistance in Acinetobacter baumannii infections disclosed a faster increase in OECD countries. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, S.; Biswas, I.; Veeraraghavan, B. Accurate identification of clinically important Acinetobacter spp.: An update. Future Sci. OA 2019, 5, FSO395. [Google Scholar] [CrossRef] [PubMed]
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M.E. Acinetobacter baumannii biofilms: Effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 2018, 11, 2277–2299. [Google Scholar] [CrossRef]
- Cosgaya, C.; Mari-Almirall, M.; van Assche, A.; Fernandez-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; Lievens, B.; et al. Acinetobacter dijkshoorniae sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex mainly recovered from clinical samples in different countries. Int. J. Syst. Evol. Microbiol. 2016, 66, 4105–4111. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; Sedo, O.; Brisse, S.; Higgins, P.G. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus–Acinetobacter baumannii complex isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2015, 63, 934–942. [Google Scholar] [CrossRef]
- Chen, T.L.; Lee, Y.T.; Kuo, S.C.; Yang, S.P.; Fung, C.P.; Lee, S.D. Rapid identification of Acinetobacter baumannii, Acinetobacter nosocomialis, and Acinetobacter pittii with a multiplex PCR assay. J. Med. Microbiol. 2014, 63, 1154–1159. [Google Scholar] [CrossRef]
- Marí-Almirall, M.; Cosgaya, C.; Higgins, P.G.; Van Assche, A.; Telli, M.; Huys, G.; Lievens, B.; Seifert, H.; Dijkshoorn, L.; Roca, I.; et al. MALDI-TOF/MS identification of species from the Acinetobacter baumannii (ab) group revisited: Inclusion of the novel A. seifertii and A. dijkshoorniae species. Clin. Microbiol. Infect. 2017, 23, 210.e1–210.e9. [Google Scholar] [CrossRef] [PubMed]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Garnacho-Montero, J.; Timsit, J.F. Managing Acinetobacter baumannii infections. Curr. Opin. Infect. Dis. 2019, 32, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nature 2017, 543, 15. [Google Scholar] [CrossRef] [PubMed]
- Barth, V.C.J.; Rodrigues, B.Á.; Bonatto, G.D.; Gallo, S.W.; Pagnussatti, V.E.; Ferreira, C.A.S.; de Oliveira, S.D. Heterogeneous persister cells formation in Acinetobacter baumannii. PLoS ONE 2013, 8, e84361. [Google Scholar] [CrossRef] [PubMed]
- Davies, J. Origins and evolution of antibiotic resistance. Microbiologia 1996, 12, 9–16. [Google Scholar] [CrossRef]
- McPhee, J.B.; Tamber, S.; Brazas, M.D.; Lewenza, S.; Hancock, R.E.W. Antibiotic Resistance Due to Reduced Uptake. In Antimicrobial Drug Resistance: Mechanisms of Drug Resistance; Mayers, D.L., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 97–110. [Google Scholar]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef]
- Amaral, L.; Martins, A.; Spengler, G.; Molnar, J. Efflux pumps of Gram-negative bacteria: What they do, how they do it, with what and how to deal with them. Front. Pharmacol. 2014, 4, 168. [Google Scholar] [CrossRef]
- Zampieri, M. The genetic underground of antibiotic resistance. Science 2021, 371, 783–784. [Google Scholar] [CrossRef]
- Mee, M.T.; Collins, J.J.; Church, G.M.; Wang, H.H. Syntrophic exchange in synthetic microbial communities. Proc. Natl. Acad. Sci. USA 2014, 111, E2149–E2156. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef]
- Sharkey, L.K.; Edwards, T.A.; O’Neill, A.J. ABC-F proteins mediate antibiotic resistance through ribosomal protection. mBio 2016, 7, e01975. [Google Scholar] [CrossRef]
- Mickiewicz, K.M.; Kawai, Y.; Drage, L.; Gomes, M.C.; Davison, F.; Pickard, R.; Hall, J.; Mostowy, S.; Aldridge, P.D.; Errington, J. Possible role of L-form switching in recurrent urinary tract infection. Nat. Commun. 2019, 10, 4379. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Lou, H.; Zhu, R.; Zhu, J.; Zhang, D.; Zhao, B.S.; Zeng, S.; Chen, X.; Chan, J.; He, C.; et al. The multiple antibiotic resistance regulator MarR is a copper sensor in Escherichia coli. Nat. Chem. Biol. 2014, 10, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Novović, K.; Jovčić, B. Colistin resistance in Acinetobacter baumannii: Molecular mechanisms and epidemiology. Antibiotics 2023, 12, 516. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, B.; Gajic, I.; Dimkic, I.; Kekic, D.; Zornic, S.; Pozder, T.; Radisavljevic, S.; Opavski, N.; Kojic, M.; Ranin, L. The first nationwide multicenter study of Acinetobacter baumannii recovered in Serbia: Emergence of OXA-72, OXA-23 and NDM-1-producing isolates. Antimicrob. Resist. Infect. Control 2020, 9, 101. [Google Scholar] [CrossRef]
- Isler, B.; Doi, Y.; Bonomo, R.A.; Paterson, D.L. New treatment options against carbapenem-resistant Acinetobacter baumannii infections. Antimicrob. Agents Chemother. 2019, 63, e01110–e01118. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf (accessed on 20 June 2023).
- Domalaon, R.; Zhanel, G.G.; Schweizer, F. Short antimicrobial peptides and peptide scaffolds as promising antibacterial agents. Curr. Top. Med. Chem. 2016, 16, 1217–1230. [Google Scholar] [CrossRef]
- Vrancianu, C.O.; Gheorghe, I.; Czobor, I.B.; Chifiriuc, M.C. Antibiotic resistance profiles, molecular mechanisms and innovative treatment strategies of Acinetobacter baumannii. Microorganisms 2020, 8, 935. [Google Scholar] [CrossRef]
- Fan, L.; Sun, J.; Zhou, M.; Zhou, J.; Lao, X.; Zheng, H.; Xu, H. DRAMP: A comprehensive data repository of antimicrobial peptides. Sci. Rep. 2016, 6, 24482. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sunkara, L.T. Avian antimicrobial host defense peptides: From biology to therapeutic applications. Pharmaceuticals 2014, 7, 220. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.; Ortiz, C.; Guzman, F.; Fernandez-Lafuente, R.; Torres, R. Antimicrobial peptides: Promising compounds against pathogenic microorganisms. Curr. Med. Chem. 2014, 21, 2299. [Google Scholar] [CrossRef] [PubMed]
- Govender, T.; Dawood, A.; Esterhuyse, A.J.; Katerere, D.R. Antimicrobial properties of the skin secretions of frogs. S. Afr. J. Sci. 2012, 108, 25–30. [Google Scholar] [CrossRef][Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta 2016, 1858, 980–987. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug Resist. Updates 2016, 26, 43–57. [Google Scholar] [CrossRef]
- Bobone, S.; Roversi, D.; Giordano, L.; De Zotti, M.; Formaggio, F.; Toniolo, C.; Park, Y.; Stella, L. The lipid dependence of antimicrobial peptide activity is an unreliable experimental test for different pore models. Biochemistry 2012, 51, 10124–10126. [Google Scholar] [CrossRef]
- Roversi, D.; Luca, V.; Aureli, S.; Park, Y.; Mangoni, M.L.; Stella, L. How many antimicrobial peptide molecules kill a bacterium? The case of PMAP-23. ACS Chem. Biol. 2014, 9, 2003–2007. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.W. Action of antimicrobial peptides: Two-state model. Biochemistry 2000, 39, 8347–8352. [Google Scholar] [CrossRef] [PubMed]
- Parchebafi, A.; Tamanaee, F.; Ehteram, H.; Ahmad, E.; Nikzad, H.; Haddad Kashani, H. The dual interaction of antimicrobial peptides on bacteria and cancer cells; mechanism of action and therapeutic strategies of nanostructures. Microb. Cell Fact. 2022, 21, 118. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A.; Ackermann, M.; Huttner, K.M. Small, anionic, and charge-neutralizing propeptide fragments of zymogens are antimicrobial. Antimicrob. Agents Chemother. 1997, 41, 1615–1617. [Google Scholar] [CrossRef] [PubMed]
- Subbalakshmi, C.; Sitaram, N. Mechanism of antimicrobial action of indolicidin. FEMS Microbiol. Lett. 1998, 160, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Sass, V.; Pag, U.; Tossi, A.; Bierbaum, G.; Sahl, H.G. Mode of action of human beta-defensin 3 against Staphylococcus aureus and transcriptional analysis of responses to defensin challenge. Int. J. Med. Microbiol. 2008, 298, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Soehnlein, O.; Kai-Larsen, Y.; Frithiof, R.; Sorensen, O.E.; Kenne, E.; Scharffetter-Kochanek, K.; Eriksson, E.E.; Herwald, H.; Agerberth, B.; Lindbom, L. Neutrophil primary granule proteins HBP and HNP1-3 boost bacterial phagocytosis by human and murine macrophages. J. Clin. Investig. 2008, 118, 3491–3502. [Google Scholar] [CrossRef] [PubMed]
- Funderburg, N.; Lederman, M.M.; Feng, Z.; Drage, M.G.; Jadlowsky, J.; Harding, C.V.; Weinberg, A.; Sieg, S.F. Human -defensin-3 activates professional antigen-presenting cells via Toll-like receptors 1 and 2. Proc. Natl. Acad. Sci. USA 2007, 104, 18631–18635. [Google Scholar] [CrossRef]
- Ehrenstein, G.; Lecar, H. Electrically gated ionic channels in lipid bilayers. Q. Rev. Biophys. 1977, 10, 1–34. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Breukink, E.; de Kruijff, B. The lantibiotic nisin, a special case or not? Biochim. Biophys. Acta 1999, 1462, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Wimley, W.C. Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chem. Biol. 2010, 5, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Rapaport, D.; Shai, Y. Interaction of fluorescently labeled pardaxin and its analogs with lipid bilayers. J. Biol. Chem. 1991, 266, 23769–23775. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y.; Bach, D.; Yanovsky, A. Channel formation properties of synthetic pardaxin and analogs. J. Biol. Chem. 1990, 265, 20202–20209. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, N.; Matsuzaki, K. Polar angle as a determinant of amphipathic α-helix-lipid interactions: A model peptide study. Biophys. J. 2000, 79, 2075–2083. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-H.; Hall, K.N.; Aguilar, M.-I. Antimicrobial peptide structure and mechanism of action: A focus on the role of membrane structure. Curr. Top. Med. Chem. 2016, 16, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.T.J.; Hale, J.D.; Elliot, M.; Hancock, R.E.W.; Straus, S.K. Effect of membrane composition on antimicrobial peptides aurein 2.2 and 2.3 from Australian southern bell frogs. Biophys. J. 2009, 96, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Sparr, E.; Ash, W.L.; Nazarov, P.V.; Rijkers, D.T.S.; Hemminga, M.A.; Tieleman, D.P.; Killian, J.A. Self-association of transmembrane-helices in model membranes. J. Biol. Chem. 2005, 280, 39324–39331. [Google Scholar] [CrossRef]
- Cheng, J.T.J.; Hale, J.D.; Elliott, M.; Hancock, R.E.W.; Straus, S.K. The importance of bacterial membrane composition in the structure and function of aurein 2.2 and selected variants. Biochim. Biophys. Acta Biomembr. 2011, 1808, 622–633. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef]
- Shai, Y. Mode of action of membrane-active antimicrobial peptides. Biopolymers 2002, 66, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.I.; Le Brun, A.P.; Whitwell, T.C.; Sani, M.-A.; James, M.; Separovic, F. The antimicrobial peptide aurein 1.2 disrupts model membranes via the carpet mechanism. Phys. Chem. Chem. Phys. 2012, 14, 15739. [Google Scholar] [CrossRef]
- Sitaram, N.; Nagaraj, R. Interaction of antimicrobial peptides with biological and model membranes: Structural and charge requirements for activity. Biochim. Biophys. Acta 1999, 1462, 29–54. [Google Scholar] [CrossRef] [PubMed]
- Rozek, A.; Friedrich, C.L.; Hancock, R.E. Structure of the bovine antimicrobial peptide indolicidin bound to dodecyl phosphocholine and sodium dodecyl sulfate micelles. Biochemistry 2000, 39, 15765–15774. [Google Scholar] [CrossRef] [PubMed]
- Gee, M.L.; Burton, M.; Grevis-James, A.; Hossain, M.A.; McArthur, S.; Palombo, E.A.; Wade, J.D.; Clayton, A.H. Imaging the action of antimicrobial peptides on living bacterial cells. Sci. Rep. 2013, 3, 1557. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Rangarajan, N.; Weisshaar, J.C. Lights, camera, action! Antimicrobial peptide mechanisms imaging in space and time. Trends Microbiol. 2016, 24, 111–122. [Google Scholar] [CrossRef] [PubMed]
- das Neves, R.C.; Mortari, M.R.; Schwartz, E.F.; Kipnis, A.; Junqueira-Kipnis, A.P. Antimicrobial and antibiofilm effects of peptides from venom of social wasp and scorpion on multidrug-resistant Acinetobacter baumannii. Toxins 2019, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kang, H.K.; Park, E.; Kim, M.K.; Park, Y. Bactericidal activities and action mechanism of the novel antimicrobial peptide Hylin a1 and its analog peptides against Acinetobacter baumannii infection. Eur. J. Pharm. Sci. 2022, 175, 106205. [Google Scholar] [CrossRef]
- Conlon, J.M.; Ahmed, E.; Pal, T.; Sonnevend, A. Potent and rapid bactericidal action of alyteserin-1c and its [E4K] analog against multidrug-resistant strains of Acinetobacter baumannii. Peptides 2010, 31, 1806–1810. [Google Scholar] [CrossRef]
- Mant, C.; Jiang, Z.; Gera, L.; Davis, T.; Nelson, K.L.; Bevers, S.; Hodges, R.S. De novo designed amphipathic α-helical antimicrobial peptides incorporating dab and dap residues on the polar face to treat the gram-negative pathogen, Acinetobacter baumannii. J. Med. Chem. 2019, 62, 3354–3366. [Google Scholar] [CrossRef]
- Barksdale, S.M.; Hrifko, E.J.; van Hoek, M.L. Cathelicidin antimicrobial peptide from Alligator mississippiensis has antibacterial activity against multi-drug resistant Acinetobacter baumanii and Klebsiella pneumonia. Dev. Comp. Immunol. 2017, 70, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Santana, F.L.; Arenas, I.; Haney, E.F.; Estrada, K.; Hancock, R.E.W.; Corzo, G. Identification of a crocodylian β-defensin variant from Alligator mississippiensis with antimicrobial and antibiofilm activity. Peptides 2021, 141, 170549. [Google Scholar] [CrossRef] [PubMed]
- Defraine, V.; Schuermans, J.; Grymonprez, B.; Govers, S.K.; Aertsen, A.; Fauvart, M.; Michiels, J.; Lavigne, R.; Briers, Y. Efficacy of artilysin art-175 against resistant and persistent Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 3480–3488. [Google Scholar] [CrossRef] [PubMed]
- Jaśkiewicz, M.; Neubauer, D.; Kazor, K.; Bartoszewska, S.; Kamysz, W. Antimicrobial activity of selected antimicrobial peptides against planktonic culture and biofilm of Acinetobacter baumannii. Probiotics Antimicrob. Proteins 2019, 11, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.J.; Pitts, R.E.; Pearson, R.A.; King, L.B. The effects of antimicrobial peptides WAM-1 and LL-37 on multidrug-resistant Acinetobacter baumannii. Pathog. Dis. 2018, 76, fty007. [Google Scholar] [CrossRef] [PubMed]
- Falla, T.J.; Karunaratne, D.N.; Hancock, R.E. Mode of action of the antimicrobial peptide indolicidin. J. Biol. Chem. 1996, 271, 19298–19303. [Google Scholar] [CrossRef]
- Wu, M.; Hancock, R.E. Interaction of the cyclic antimicrobial cationic peptide bactenecin with the outer and cytoplasmic membrane. J. Biol. Chem. 1999, 274, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Eales, M.G.; Ferrari, E.; Goddard, A.D.; Lancaster, L.; Sanderson, P.; Miller, C. Mechanistic and phenotypic studies of bicarinalin, BP100 and colistin action on Acinetobacter baumannii. Res. Microbiol. 2018, 169, 296–302. [Google Scholar] [CrossRef]
- Jung, C.-J.; Liao, Y.-D.; Hsu, C.-C.; Huang, T.-Y.; Chuang, Y.-C.; Chen, J.-W.; Kuo, Y.-M.; Chia, J.-S. Identification of potential therapeutic antimicrobial peptides against Acinetobacter baumannii in a mouse model of pneumonia. Sci. Rep. 2021, 11, 7318. [Google Scholar] [CrossRef]
- Luo, X.; Ye, X.; Ding, L.; Zhu, W.; Zhao, Z.; Luo, D.; Liu, N.; Sun, L.; Chen, Z. Identification of the scorpion venom-derived antimicrobial peptide Hp1404 as a new antimicrobial agent against carbapenem-resistant Acinetobacter baumannii. Microb. Pathog. 2021, 157, 104960. [Google Scholar] [CrossRef]
- Sahoo, A.; Swain, S.S.; Behera, A.; Sahoo, G.; Mahapatra, P.K.; Panda, S.K. Antimicrobial peptides derived from insects offer a novel therapeutic option to combat biofilm: A review. Front. Microbiol. 2021, 10, 661195. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Sonnevend, A.; Pál, T.; Vila-Farrés, X. Efficacy of six frog skin-derived antimicrobial peptides against colistin-resistant strains of the Acinetobacter baumannii group. Int. J. Antimicrob. Agents 2012, 39, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghaferi, N.; Kolodziejek, J.; Nowotny, N.; Coquet, L.; Jouenne, T.; Leprince, J.; Vaudry, H.; King, J.D.; Conlon, J.M. Antimicrobial peptides from the skin secretions of the South-East Asian frog Hylarana erythraea (Ranidae). Peptides 2010, 31, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Ahmed, E.; Condamine, E. Antimicrobial properties of brevinin-2-related peptide and its analogs: Efficacy against multidrug-resistant Acinetobacter baumannii. Chem. Biol. Drug Des. 2009, 74, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Jayamani, E.; Rajamuthiah, R.; Larkins-Ford, J.; Fuchs, B.B.; Conery, A.L.; Vilcinskas, A.; Ausubel, F.M.; Mylonakis, E. Insect-derived cecropins display activity against Acinetobacter baumannii in a whole-animal high-throughput Caenorhabditis elegans model. Antimicrob. Agents Chemother. 2015, 59, 1728–1737. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, A.; Cirioni, O.; Del Prete, M.S.; Barchiesi, F.; Paggi, A.M.; Petrelli, E.; Scalise, G. Comparative activities of polycationic peptides and clinically used antimicrobial agents against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. J. Antimicrob. Chemother. 2000, 46, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Kang, N.; Ko, S.J.; Park, J.; Park, E.; Shin, D.W.; Kim, S.H.; Lee, S.A.; Lee, J.I.; Lee, S.H.; et al. Antibacterial and antibiofilm activity and mode of action of Magainin 2 against drug-resistant Acinetobacter baumannii. Int. J. Mol. Sci. 2018, 19, 3041. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: Buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions. Biochem. Biophys. Res. Commun. 1998, 244, 253–257. [Google Scholar] [CrossRef]
- Cirioni, O.; Silvestri, C.; Ghiselli, R.; Orlando, F.; Riva, A.; Gabrielli, E.; Mocchegiani, F.; Cianforlini, N.; Trombettoni, M.M.; Saba, V.; et al. Therapeutic efficacy of buforin II and rifampin in a rat model of Acinetobacter baumannii sepsis. Crit. Care Med. 2009, 37, 1403–1407. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, P.; Xiao, L.; Liu, Y.; Wu, K.; Ni, G.; Li, H.; Wang, T.; Wu, X.; Chen, G.; et al. Caerin 1.1 and 1.9 peptides from australian tree frog inhibit antibiotic-resistant bacteria growth in a murine skin infection model. Microbiol. Spectr. 2021, 9, e0005121. [Google Scholar] [CrossRef]
- Tajbakhsh, M.; Akhavan, M.M.; Fallah, F.; Karimi, A. A recombinant snake cathelicidin derivative peptide: Antibiofilm properties and expression in Escherichia coli. Biomolecules 2018, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Vila-Farres, X.; De La Maria, C.G.; López-Rojas, R.; Pachón, J.; Giralt, E.; Vila, J. In vitro activity of several antimicrobial peptides against colistin-susceptible and colistin-resistant Acinetobacter baumannii. Clin. Microbiol. Infect. 2012, 18, 383–387. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; D’Amato, G.; Silvestri, C.; Del Prete, M.S.; Łukasiak, J.; Scalise, G. Comparative activities of cecropin A, melittin, and cecropin A-melittin peptide CA(1-7)M(2-9)NH2 against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. Peptides 2003, 24, 1315–1318. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G.; Agerberth, B.; Boman, A. Mechanisms of action on Escherichia coli of cecropin P1 and PR-39, two antibacterial peptides from pig intestine. Infect. Immun. 1993, 61, 2978–2984. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, A.; Gupta, K.; Van Hoek, M.L. Characterization of Cimex lectularius (bedbug) defensin peptide and its antimicrobial activity against human skin microflora. Biochem. Biophys. Res. Commun. 2016, 470, 955–960. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cai, Y.; Wang, R.; Liang, B.B.; An, M.M. In-vitro bactericidal activity of colistin against biofilm-associated Pseudomonas aeruginosa and Acinetobacter baumannii. J. Hosp. Infect. 2009, 72, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Al-Ghaferi, N.; Ahmed, E.; Meetani, M.A.; Leprince, J.; Nielsen, P.F. Orthologs of magainin, PGLa, procaerulein-derived, and proxenopsin-derived peptides from skin secretions of the octoploid frog Xenopus amieti (Pipidae). Peptides 2010, 31, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Mechkarska, M.; Ahmed, E.; Leprince, J.; Vaudry, H.; King, J.D.; Takada, K. Purification and properties of antimicrobial peptides from skin secretions of the Eritrea clawed frog Xenopus clivii (Pipidae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 350–354. [Google Scholar] [CrossRef]
- Mechkarska, M.; Ahmed, E.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; King, J.D.; Conlon, J.M. Antimicrobial peptides with therapeutic potential from skin secretions of the Marsabit clawed frog Xenopus borealis (Pipidae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 467–472. [Google Scholar] [CrossRef]
- Kirkpatrick, C.L.; Broberg, C.A.; McCool, E.M.; Lee, W.J.; Chao, A.; McConnell, E.W.; Pritchard, D.A.; Hebert, M.; Fleeman, R.; Adams, J.; et al. The “PepSAVI-MS” pipeline for natural product bioactive peptide discovery. Anal. Chem. 2017, 89, 1194–1201. [Google Scholar] [CrossRef]
- Domhan, C.; Uhl, P.; Kleist, C.; Zimmermann, S.; Umstätter, F.; Leotta, K.; Mier, W.; Wink, M. Replacement of L-amino acids by d-amino acids in the antimicrobial peptide ranalexin and its consequences for antimicrobial activity and biodistribution. Molecules 2019, 24, 2987. [Google Scholar] [CrossRef] [PubMed]
- Sağmak Tartar, A.; Özer Balin, Ş.; Akbulut, A.; Yardim, M.; Aydin, S. Roles of dermcidin, salusin-α, salusin-β and TNF-α in the pathogenesis of human brucellosis. Iran. J. Immunol. 2019, 16, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Farshadzadeh, Z.; Pourhajibagher, M.; Taheri, B.; Ekrami, A.; Modarressi, M.H.; Azimzadeh, M.; Bahador, A. Antimicrobial and anti-biofilm potencies of dermcidin-derived peptide DCD-1L against Acinetobacter baumannii: An in vivo wound healing model. BMC Microbiol. 2022, 22, 25. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Su, P.Y.; Kuo, S.C.; Lauderdale, T.Y.; Shih, C. Adding a c-terminal cysteine (ctc) can enhance the bactericidal activity of three different antimicrobial peptides. Front. Microbiol. 2018, 9, 1440. [Google Scholar] [CrossRef]
- Tejman-Yarden, N.; Robinson, A.; Davidov, Y.; Shulman, A.; Varvak, A.; Reyes, F.; Rahav, G.; Nissan, I. Delftibactin-A, a non-ribosomal peptide with broad antimicrobial activity. Front. Microbiol. 2019, 10, 2377. [Google Scholar] [CrossRef]
- Liu, C.B.; Shan, B.; Bai, H.M.; Tang, J.; Yan, L.Z.; Ma, Y.B. Hydrophilic/hydrophobic characters of antimicrobial peptides derived from animals and their effects on multidrug resistant clinical isolates. Dongwuxue Yanjiu 2015, 36, 41–47. [Google Scholar] [CrossRef]
- Dekan, Z.; Headey, S.J.; Scanlon, M.; Baldo, B.A.; Lee, T.-H.; Aguilar, M.-I.; Deuis, J.R.; Vetter, I.; Elliott, A.G.; Amado, M.; et al. ∆-Myrtoxin-Mp1a is a helical heterodimer from the venom of the jack jumper ant that has antimicrobial, membrane-disrupting, and nociceptive activities. Angew. Chem. Int. Ed. Engl. 2017, 56, 8495–8499. [Google Scholar] [CrossRef]
- Zhao, F.; Lan, X.Q.; Du, Y.; Chen, P.Y.; Zhao, J.; Zhao, F.; Lee, W.-H.; Zhang, Y. King cobra peptide OH-CATH30 as a potential candidate drug through clinic drug-resistant isolates. Zool. Res. 2018, 9, 87–96. [Google Scholar] [CrossRef]
- Mechkarska, M.; Prajeep, M.; Radosavljevic, G.D.; Jovanovic, I.P.; Al Baloushi, A.; Sonnevend, A.; Lukic, M.L.; Conlon, J.M. An analog of the host-defense peptide hymenochirin-1B with potent broad-spectrum activity against multidrug-resistant bacteria and immunomodulatory properties. Peptides 2013, 50, 153–159. [Google Scholar] [CrossRef]
- Bolatchiev, A. Antimicrobial peptides Epinecidin-1 and Beta-Defesin-3 are effective against a broad spectrum of antibiotic-resistant bacterial isolates and increase survival rate in experimental sepsis. Antibiotics 2022, 11, 76. [Google Scholar] [CrossRef]
- Khan, A.; Xu, M.; Wang, T.; You, C.; Wang, X.; Ren, H.; Zhou, H.; Khan, A.; Han, C.; Li, P. Catechol cross-linked antimicrobial peptide hydrogels prevent multidrug-resistant Acinetobacter baumannii infection in burn wounds. Biosci. Rep. 2019, 39, BSR20190504. [Google Scholar] [CrossRef] [PubMed]
- Sacco, F.; Bitossi, C.; Casciaro, B.; Loffredo, M.R.; Fabiano, G.; Torrini, L.; Raponi, F.; Raponi, G.; Mangoni, M.L. The antimicrobial peptide Esc(1-21) synergizes with colistin in inhibiting the growth and in killing multidrug resistant Acinetobacter baumannii strains. Antibiotics 2022, 11, 234. [Google Scholar] [CrossRef] [PubMed]
- Gordya, N.; Yakovlev, A.; Kruglikova, A.; Tulin, D.; Potolitsina, E.; Suborova, T.; Bordo, D.; Rosano, C.; Chernysh, S. Natural antimicrobial peptide complexes in the fighting of antibiotic resistant biofilms: Calliphora vicina medicinal maggots. PLoS ONE 2017, 12, e0173559. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Galadari, S.; Raza, H.; Condamine, E. Design of potent, non-toxic antimicrobial agents based upon the naturally occurring frog skin peptides, ascaphin-8 and peptide XT-7. Chem. Biol. Drug Des. 2008, 72, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, S.H.; Murphy, R.A.; Juul-Madsen, K.; Fredborg, M.; Hvam, M.L.; Axelgaard, E.; Skovdal, S.M.; Meyer, R.L.; Sørensen, U.B.S.; Möller, A.; et al. The immunomodulatory drug glatiramer acetate is also an effective antimicrobial agent that kills gram-negative bacteria. Sci. Rep. 2017, 7, 15653. [Google Scholar] [CrossRef] [PubMed]
- Routsias, J.G.; Karagounis, P.; Parvulesku, G.; Legakis, N.J.; Tsakris, A. In vitro bactericidal activity of human β-defensin 2 against nosocomial strains. Peptides 2010, 31, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Maisetta, G.; Batoni, G.; Esin, S.; Florio, W.; Bottai, D.; Favilli, F.; Campa, M. In vitro bactericidal activity of human beta-defensin 3 against multidrug-resistant nosocomial strains. Antimicrob. Agents Chemother. 2006, 50, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, G.; Wang, S.; Chen, Y.; Gong, Y.; Chen, S.; Xu, Y.; Hu, M.; Wang, X.; Zeng, H.; et al. A simplified derivative of human defensin 5 with potent and efficient activity against multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2018, 62, e01504–e01517. [Google Scholar] [CrossRef]
- Romeo, D.; Skerlavaj, B.; Bolognesi, M.; Gennaro, R. Structure and bactericidal activity of an antibiotic dodecapeptide purified from bovine neutrophils. J. Biol. Chem. 1988, 263, 9573–9575. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, M.K.; Park, Y. Comparative antimicrobial activity of Hp404 peptide and its analogs against Acinetobacter baumannii. Int. J. Mol. Sci. 2021, 22, 5540. [Google Scholar] [CrossRef]
- Neshani, A.; Sedighian, H.; Mirhosseini, S.A.; Ghazvini, K.; Zare, H.; Jahangiri, A. Antimicrobial peptides as a promising treatment option against Acinetobacter baumannii infections. Microb. Pathog. 2020, 146, 104238. [Google Scholar] [CrossRef] [PubMed]
- Serra, I.; Scorciapino, M.A.; Manzo, G.; Casu, M.; Rinaldi, A.C.; Attoub, S.; Mechkarska, M.; Conlon, J.M. Conformational analysis and cytotoxic activities of the frog skin host-defense peptide, hymenochirin-1Pa. Peptides 2014, 61, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Sambanthamoorthy, K.; Palys, T.; Paranavitana, C. The human antimicrobial peptide LL-37 and its fragments possess both antimicrobial and antibiofilm activities against multidrug-resistant Acinetobacter baumannii. Peptides 2013, 49, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, L.; Mahdi, N.; Al-Kakei, S.; Musafer, H.; Al-Joofy, I.; Essa, R.; Zwain, L.; Salman, I.; Mater, H.; Al-Alak, S.; et al. Treatment strategy by lactoperoxidase and lactoferrin combination: Immunomodulatory and antibacterial activity against multidrug-resistant Acinetobacter baumannii. Microb. Pathog. 2018, 114, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Neshani, A.; Zare, H.; Akbari Eidgahi, M.R.; Chichaklu, A.H.; Movaqar, A.; Ghazvini, K. Review of antimicrobial peptides with anti-helicobacter pylori activity. Helicobacter 2019, 24, e12555. [Google Scholar] [CrossRef]
- Tan, H.; Wang, J.; Song, Y.; Liu, S.; Lu, Z.; Luo, H.; Tang, X. Antibacterial potential analysis of novel α-helix peptides in the chinese wolf spider Lycosa sinensis. Pharmaceutics 2022, 14, 2540. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, R.; Wiesner, J.; Marker, A.; Pfeifer, Y.; Bauer, A.; Hammann, P.E.; Vilcinskas, A. Profiling antimicrobial peptides from the medical maggot Lucilia sericata as potential antibiotics for MDR gram-negative bacteria. J. Antimicrob. Chemother. 2019, 74, 96–107. [Google Scholar] [CrossRef]
- Jayawant, E.S.; Hutchinson, J.; Gašparíková, D.; Lockey, C.; Pruñonosa Lara, L.; Guy, C.; Brooks, R.L.; Dixon, A.M. Molecular basis of selectivity and activity for the antimicrobial peptide lynronne-1 informs rational design of peptide with improved activity. Chembiochem 2021, 22, 2430–2439. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef]
- Vila-Farrés, X.; López-Rojas, R.; Pachón-Ibáñez, M.E.; Teixidó, M.; Pachón, J.; Vila, J.; Giralt, E. Sequence-activity relationship, and mechanism of action of mastoparan analogues against extended-drug resistant Acinetobacter baumannii. Eur. J. Med. Chem. 2015, 101, 34–40. [Google Scholar] [CrossRef]
- Al-Khafaji, Z.; Al-Samaree, M. Design of synthetic antimicrobial peptides against resistant Acinetobacter baumannii using computational approach. Int. J. Pharmaceut Sci. Res. 2017, 8, 2033–2039255. [Google Scholar] [CrossRef]
- Lin, C.H.; Lee, M.C.; Tzen, J.T.C.; Lee, H.M.; Chang, S.M.; Tu, W.C.; Lin, C.F. Efficacy of Mastoparan-AF alone and in combination with clinically used antibiotics on nosocomial multidrug-resistant Acinetobacter baumannii. Saudi J. Biol. Sci. 2017, 24, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Ikram, A.; Raza, A.; Saeed, S.; Zafar Paracha, R.; Younas, Z.; Khadim, M.T. Therapeutic potential of novel mastoparan-chitosan nanoconstructs against clinical MDR Acinetobacter baumannii: In silico, in vitro and in vivo studies. Int. J. Nanomed. 2021, 16, 3755–3773. [Google Scholar] [CrossRef] [PubMed]
- Gui, S.; Li, R.; Feng, Y.; Wang, S. Transmission electron microscopic morphological study and flow cytometric viability assessment of Acinetobacter baumannii susceptible to Musca domestica cecropin. ScientificWorldJournal 2014, 2014, 657536. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pashaei, F.; Bevalian, P.; Akbari, R.; Pooshang Bagheri, K. Single dose eradication of extensively drug resistant Acinetobacter spp. In a mouse model of burn infection by melittin antimicrobial peptide. Microb. Pathog. 2019, 127, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Akbar, R.; Hakemi-Vala, M.; Pashaie, F.; Bevalian, P.; Hashemi, A.; Pooshang Bagheri, K. Highly synergistic effects of melittin with conventional antibiotics against multidrug-resistant isolates of Acinetobacter baumannii and Pseudomonas aeruginosa. Microb. Drug Resist. 2019, 25, 193–202. [Google Scholar] [CrossRef]
- Bardbari, A.M.; Arabestani, M.R.; Karami, M.; Keramat, F.; Aghazadeh, H.; Alikhani, M.Y.; Bagheri, K.P. Highly synergistic activity of melittin with imipenem and colistin in biofilm inhibition against multidrug-resistant strong biofilm producer strains of Acinetobacter baumannii. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Shamova, O.V.; Orlov, D.S.; Zharkova, M.S.; Balandin, S.V.; Yamschikova, E.V.; Knappe, D.; Hoffmann, R.; Kokryakov, V.N.; Ovchinnikova, T.V. Minibactenecins ChBac7.Nα and ChBac7. Nβ—Antimicrobial peptides from leukocytes of the goat Capra hircus. Acta Naturae 2016, 8, 136–146. [Google Scholar] [CrossRef]
- Han, H.M.; Ko, S.; Cheong, M.J.; Bang, J.K.; Seo, C.H.; Luchian, T.; Park, Y. Myxinidin2 and myxinidin3 suppress inflammatory responses through STAT3 and MAPKs to promote wound healing. Oncotarget 2017, 8, 87582–87597. [Google Scholar] [CrossRef]
- Irani, N.; Basardeh, E.; Samiee, F.; Fateh, A.; Shooraj, F.; Rahimi, A.; Shahcheraghi, F.; Vaziri, F.; Masoumi, M.; Pazhouhandeh, M.; et al. The inhibitory effect of the combination of two new peptides on biofilm formation by Acinetobacter baumannii. Microb. Pathog. 2018, 121, 310–317. [Google Scholar] [CrossRef]
- Jahangiri, A.; Neshani, A.; Mirhosseini, S.A.; Ghazvini, K.; Zare, H.; Sedighian, H. Synergistic effect of two antimicrobial peptides, Nisin and P10 with conventional antibiotics against extensively drug-resistant Acinetobacter baumannii and colistin-resistant Pseudomonas aeruginosa isolates. Microb. Pathog. 2021, 150, 104700. [Google Scholar] [CrossRef] [PubMed]
- Jenei, S.; Tiricz, H.; Szolomájer, J.; Tímár, E.; Klement, É.; Al Bouni, M.A.; Lima, R.M.; Kata, D.; Harmati, M.; Buzás, K.; et al. Potent chimeric antimicrobial derivatives of the medicago truncatula ncr247 symbiotic peptide. Front. Microbiol. 2020, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.M.; Rathod, B.B.; Tiricz, H.; Howan, D.H.O.; Al Bouni, M.A.; Jenei, S.; Tímár, E.; Endre, G.; Tóth, G.K.; Kondorosi, É. legume plant peptides as sources of novel antimicrobial molecules against human pathogens. Front. Mol. Biosci. 2022, 9, 870460. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.M.; Zorgani, A.; Jalowicki, G.; Kerr, A.; Khaldi, N.; Martins, M. Unlocking NuriPep 1653 from common pea protein: A potent antimicrobial peptide to tackle a pan-drug resistant Acinetobacter baumannii. Front Microbiol 2019, 18, 2086. [Google Scholar] [CrossRef] [PubMed]
- Flamm, R.K.; Rhomberg, P.R.; Simpson, K.M.; Farrell, D.J.; Sader, H.S.; Jones, R.N. In vitro spectrum of pexiganan activity when tested against pathogens from diabetic foot infections and with selected resistance mechanisms. Antimicrob. Agents Chemother. 2015, 59, 1751–1754. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ge, Y.; MacDonald, D.L.; Holroyd, K.J.; Thornsberry, C.; Wexler, H.; Zasloff, M. In vitro antibacterial properties of pexiganan, an analog of magainin. Antimicrob. Agents Chemother. 1999, 43, 782–788. [Google Scholar] [CrossRef] [PubMed]
- McLean, D.T.; McCrudden, M.T.; Linden, G.J.; Irwin, C.R.; Conlon, J.M.; Lundy, F.T. Antimicrobial and immunomodulatory properties of PGLa-AM1, CPF-AM1, and magainin-AM1: Potent activity against oral pathogens. Regul. Pept. 2014, 194–195, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Morroni, G.; Simonetti, O.; Brenciani, A.; Brescini, L.; Kamysz, W.; Kamysz, E.; Neubauer, D.; Caffarini, M.; Orciani, M.; Giovanetti, E.; et al. In vitro activity of Protegrin-1, alone and in combination with clinically useful antibiotics, against Acinetobacter baumannii strains isolated from surgical wounds. Med. Microbiol. Immunol. 2019, 208, 877–883. [Google Scholar] [CrossRef]
- Thandar, M.; Lood, R.; Winer, B.Y.; Deutsch, D.R.; Euler, C.W.; Fischetti, V.A. novel engineered peptides of a phage lysin as effective antimicrobials against multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 671–679. [Google Scholar] [CrossRef]
- De Breij, A.; Riool, M.; Cordfunke, R.A.; Malanovic, N.; De Boer, L.; Koning, R.I.; Ravensbergen, E.; Franken, M.; van der Heijde, T.; Boekema, B.K.; et al. The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Sci. Transl. Med. 2018, 10, eaan4044. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Arafat, K.; Attoub, S.; Sonnevend, A. Analogues of the frog skin peptide alyteserin-2a with enhanced antimicrobial activities against Gram-negative bacteria. J. Pept. Sci. 2012, 18, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, A.K.; Alamri, M.A.; Almasoudi, A.S.; Abbasmanthiri, R.; Mahfoud, M. Evaluation of the in vitro antimicrobial activity of selected Saudi scorpion venoms tested against multidrug-resistant micro-organisms. J. Glob. Antimicrob. Resist. 2017, 10, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wong, E.S.W.; Whitley, J.C.; Li, J.; Stringer, J.M.; Short, K.R.; Renfree, M.B.; Belov, K.; Cocks, B.G. Ancient antimicrobial peptides kill antibiotic-resistant pathogens: Australian mammals provide new options. PLoS ONE 2011, 6, e24030. [Google Scholar] [CrossRef] [PubMed]
- Swedan, S.; Shubair, Z.; Almaaytah, A. Synergism of cationic antimicrobial peptide WLBU2 with antibacterial agents against biofilms of multi-drug resistant Acinetobacter baumannii and Klebsiella pneumoniae. Infect. Drug Resist. 2019, 12, 2019–2030. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Yin, Y.; Wang, G.; Yang, M.; Li, Y.; Zhang, Z.; Lai, R. The antimicrobial peptide ZY4 combats multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii infection. Proc. Natl. Acad. Sci. USA 2019, 116, 26516–26522. [Google Scholar] [CrossRef] [PubMed]
- Andersen, I.K.L.; Thomsen, T.T.; Rashid, J.; Bobak, T.R.; Oddo, A.; Franzyk, H.; Løbner-Olesen, A.; Hansen, P.R. C-locked analogs of the antimicrobial peptide BP214. Antibiotics 2022, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Gopal, R.; Kim, Y.G.; Lee, J.H.; Lee, S.K.; Chae, J.D.; Son, B.K.; Seo, C.H.; Park, Y. Synergistic effects and antibiofilm properties of chimeric peptides against multidrug-resistant Acinetobacter baumannii strains. Antimicrob. Agents Chemother. 2014, 58, 1622–1629. [Google Scholar] [CrossRef]
- Peng, J.; Long, H.; Liu, W.; Wu, Z.; Wang, T.; Zeng, Z.; Guo, G.; Wu, J. Antibacterial mechanism of peptide Cec4 against Acinetobacter baumannii. Infect. Drug Resist. 2019, 12, 417–2428. [Google Scholar] [CrossRef]
- Peng, J.; Wang, Y.; Wu, Z.; Mao, C.; Li, L.; Cao, H.; Qiu, Z.; Guo, G.; Liang, G.; Shen, F. Antimicrobial peptide Cec4 eradicates multidrug-resistant Acinetobacter baumannii in vitro and in vivo. Drug Des. Dev. Ther. 2023, 30, 977–992. [Google Scholar] [CrossRef]
- Hacioglu, M.; Oyardi, O.; Bozkurt-Guzel, C.; Savage, P.B. Antibiofilm activities of ceragenins and antimicrobial peptides against fungal-bacterial mono and multispecies biofilms. J. Antibiot. 2020, 73, 455–462. [Google Scholar] [CrossRef]
- Ostorhazi, E.; Hoffmann, R.; Herth, N.; Wade, J.D.; Kraus, C.N.; Otvos, L., Jr. Advantage of a narrow spectrum host defense (antimicrobial) peptide over a broad spectrum analog in preclinical drug development. Front. Chem. 2018, 6, 359. [Google Scholar] [CrossRef] [PubMed]
- Jariyarattanarach, P.; Klubthawee, N.; Wongchai, M.; Roytrakul, S.; Aunpad, R. Novel D-form of hybrid peptide (D-AP19) rapidly kills Acinetobacter baumannii while tolerating proteolytic enzymes. Sci. Rep. 2022, 12, 15852. [Google Scholar] [CrossRef] [PubMed]
- Gorr, S.U.; Brigman, H.V.; Anderson, J.C.; Hirsch, E.B. The antimicrobial peptide DGL13K is active against drug-resistant gram-negative bacteria and sub-inhibitory concentrations stimulate bacterial growth without causing resistance. PLoS ONE 2022, 17, e0273504. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Hua, X.; Zhou, R.; Cui, J.; Wang, T.; Kong, F.; You, H.; Liu, X.; Adu-Amankwaah, J.; Guo, G.; et al. Antimicrobial and anti-inflammatory activities of MAF-1-derived antimicrobial peptide Mt6 and its D-enantiomer D-Mt6 against Acinetobacter baumannii by targeting cell membranes and lipopolysaccharide interaction. Microbiol. Spectr. 2022, 10, e0131222. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Z.; Li, X.; Fan, Y.; He, G.; Wan, Y.; Yu, C.; Tang, J.; Li, M.; Zhang, X.; et al. In vitro and in vivo activities of antimicrobial peptides developed using an amino acid-based activity prediction method. Antimicrob. Agents Chemother. 2014, 58, 5342–5349. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, Z.; Li, X.; Tian, Y.; Fan, Y.; Yu, C.; Zhou, B.; Liu, Y.; Xiang, R.; Yang, L. Synergistic effects of antimicrobial peptide DP7 combined with antibiotics against multidrug-resistant bacteria. Drug Des. Devel Ther. 2017, 11, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, Z.; Tian, Y.; Yin, Q.; Cheng, X.; Lian, M.; Zhou, B.; Zhang, X.; Yang, L. Efficacy of antimicrobial peptide DP7, designed by machine-learning method, against methicillin-resistant Staphylococcus aureus. Front. Microbiol. 2019, 10, 1175. [Google Scholar] [CrossRef]
- Li, J.; Prats-Ejarque, G.; Torrent, M.; Andreu, D.; Brandenburg, K.; Fernández-Millán, P.; Boix, E. In vivo evaluation of ecp peptide analogues for the treatment of Acinetobacter baumannii infection. Biomedicines 2022, 10, 386. [Google Scholar] [CrossRef]
- Taheri, B.; Mohammadi, M.; Momenzadeh, N.; Farshadzadeh, Z.; Roozbehani, M.; Dehghani, P.; Hajian, S.; Darvishi, S.; Shamseddin, J. Substitution of lysine for isoleucine at the center of the nonpolar face of the antimicrobial peptide, piscidin-1, leads to an increase in the rapidity of bactericidal activity and a reduction in toxicity. Infect. Drug Resist. 2019, 12, 1629–1647. [Google Scholar] [CrossRef]
- Ramalho, S.R.; de Sardi, J.C.O.; Júnior, E.C.; Marchetto, R.; Wender, H.; Vargas, L.F.P.; de Miranda, A.; Almeida, C.V.; de Oliveira Almeida, L.H.; de Oliveira, C.F.R.; et al. The synthetic antimicrobial peptide IKR18 displays anti-infectious properties in Galleria mellonella in vivo model. Biochim. Biophys. Acta Gen. Subj. 2022, 1866, 130244. [Google Scholar] [CrossRef]
- Gong, R.; An, Z.; Zhang, W.; Chen, F.; Wang, K.J. The antimicrobial peptide lj-hep2 from Lateolabrax japonicus exerting activities against multiple pathogenic bacteria and immune protection in vivo. Mar. Drugs 2022, 20, 651. [Google Scholar] [CrossRef]
- Brito, J.C.M.; Willliam, G.L.; Resende, J.M.; Assis, D.C.S.; Boff, D.; Cardoso, V.N.; Amaral, F.A.; Souza-Fagundes, E.M.; Fernandes, S.O.A.; Lima, M.E. Pegylated LyeTx I-b peptide is effective against carbapenem-resistant Acinetobacter baumannii in an in vivo model of pneumonia and shows reduced toxicity. Int. J. Pharm. 2021, 609, 121156. [Google Scholar] [CrossRef] [PubMed]
- Eshtiaghi, S.; Nazari, R.; Fasihi-Ramandi, M. Molecular docking, anti-biofilm & antibacterial activities and therapeutic index of mCM11 peptide on Acinetobacter baumannii strains. Curr. Microbiol. 2023, 80, 191. [Google Scholar] [CrossRef] [PubMed]
- Denardi, L.B.; de Arruda Trindade, P.; Weiblen, C.; Ianiski, L.B.; Stibbe, P.C.; Pinto, S.C.; Santurio, J.M. In vitro activity of the antimicrobial peptides h-Lf1-11, MSI-78, LL-37, fengycin 2B, and magainin-2 against clinically important bacteria. Braz. J. Microbiol. 2022, 53, 171–177. [Google Scholar] [CrossRef]
- Howan, D.H.O.; Jenei, S.; Szolomajer, J.; Endre, G.; Kondorosi, É.; Tóth, G.K. Enhanced antibacterial activity of substituted derivatives of NCR169C peptide. Int. J. Mol. Sci. 2023, 24, 2694. [Google Scholar] [CrossRef] [PubMed]
- Jayathilaka, E.H.T.T.; Rajapaksha, D.C.; Nikapitiya, C.; De Zoysa, M.; Whang, I. Antimicrobial and anti-biofilm peptide octominin for controlling multidrug-resistant Acinetobacter baumannii. Int. J. Mol. Sci. 2021, 22, 5353. [Google Scholar] [CrossRef] [PubMed]
- Jayathilaka, E.H.T.T.; Nikapitiya, C.; De Zoysa, M.; Whang, I. Antimicrobial peptide octominin-encapsulated chitosan nanoparticles enhanced antifungal and antibacterial activities. Int. J. Mol. Sci. 2022, 23, 15882. [Google Scholar] [CrossRef]
- Rajapaksha, D.C.; Jayathilaka, E.H.T.T.; Edirisinghe, S.L.; Nikapitiya, C.; Lee, J.; Whang, I.; De Zoysa, M. Octopromycin: Antibacterial and antibiofilm functions of a novel peptide derived from Octopus minor against multidrug-resistant Acinetobacter baumannii. Fish Shellfish Immunol. 2021, 117, 82–94. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, F.W.; Wang, W.X.; Zhao, Y.Y.; Sun, S.Y.; Yu, J.H.; Vitek, M.P.; Li, G.F.; Ma, R.; Wang, S.; et al. Apolipoprotein E mimetic peptide COG1410 combats pandrug-resistant Acinetobacter baumannii. Front. Microbiol. 2022, 13, 934765. [Google Scholar] [CrossRef]
- Nagarajan, D.; Roy, N.; Kulkarni, O.; Nanajkar, N.; Datey, A.; Ravichandran, S.; Thakur, C.T.S.; Aprameya, I.V.; Sarma, S.P.; Chakravortty, D.; et al. Ω76: A designed antimicrobial peptide to combat carbapenem- and tigecycline-resistant Acinetobacter baumannii. Sci. Adv. 2019, 5, eaax 1946. [Google Scholar] [CrossRef]
- Mandel, S.; Michaeli, J.; Nur, N.; Erbetti, I.; Zazoun, J.; Ferrari, L.; Felici, A.; Cohen-Kutner, M.; Bachnoff, N. OMN6 a novel bioengineered peptide for the treatment of multidrug resistant Gram-negative bacteria. Sci. Rep. 2021, 11, 6603. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, J.; Mandel, S.; Maximov, S.; Zazoun, J.; Savoia, P.; Kothari, N.; Valmont, T.; Ferrari, L.; Duncan, L.R.; Hawser, S.; et al. In vitro and in vivo antimicrobial activity of the novel peptide OMN6 against multidrug-resistant Acinetobacter baumannii. Antibiotics 2022, 11, 1201. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Jang, A.; Yoon, Y.K.; Kim, Y. development of novel peptides for the antimicrobial combination therapy against carbapenem-resistant Acinetobacter baumannii Infection. Pharmaceutics 2021, 13, 1800. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Chen, E.H.; Yang, S.Y.; Kuo, P.S.; Jan, H.M.; Yang, T.C.; Hsieh, M.Y.; Lee, K.T.; Lin, C.H.; Chen, R.P. A systematic study of the stability, safety, and efficacy of the de novo designed antimicrobial peptide PepD2 and its modified derivatives against Acinetobacter baumannii. Front. Microbiol. 2021, 12, 678330. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Morales, J.; Guardiola, S.; Ballesté-Delpierre, C.; Giralt, E.; Vila, J. A new synthetic protegrin as a promising peptide with antibacterial activity against MDR Gram-negative pathogens. J. Antimicrob. Chemother. 2022, 77, 3077–3085. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.; Lapuebla, A.; Landman, D.; Quale, J. In vitro and in vivo activity of a novel antisense peptide nucleic acid compound against multidrug-resistant Acinetobacter baumannii. Microb. Drug Resist. 2019, 25, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, M.; Choi, J.; Jang, A.; Yoon, Y.K.; Kim, Y. Antiseptic 9-Meric peptide with potency against carbapenem-resistant Acinetobacter baumannii infection. Int. J. Mol. Sci. 2021, 22, 12520. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.F.; Hamed, M.I.; Panitch, A.; Seleem, M.N. Targeting methicillin-resistant Staphylococcus aureus with short salt-resistant synthetic peptides. Antimicrob. Agents Chemother. 2014, 58, 4113–4122. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Brezden, A.; Mohammad, H.; Chmielewski, J.; Seleem, M.N. A short D-enantiomeric antimicrobial peptide with potent immunomodulatory and antibiofilm activity against multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Sci. Rep. 2017, 7, 6953. [Google Scholar] [CrossRef]
- Sharma, D.; Choudhary, M.; Vashistt, J.; Shrivastava, R.; Bisht, G.S. Cationic antimicrobial peptide and its poly-N-substituted glycine congener: Antibacterial and antibiofilm potential against A. baumannii. Biochem. Biophys. Res. Commun. 2019, 518, 472–478. [Google Scholar] [CrossRef]
- Ali, M.; van Gent, M.E.; de Waal, A.M.; van Doodewaerd, B.R.; Bos, E.; Koning, R.I.; Cordfunke, R.A.; Drijfhout, J.W.; Nibbering, P.H. Physical and functional characterization of plga nanoparticles containing the antimicrobial peptide SAAP-148. Int. J. Mol. Sci. 2023, 24, 2867. [Google Scholar] [CrossRef] [PubMed]
- Farzi, N.; Oloomi, M.; Bahramali, G.; Siadat, S.D.; Bouzari, S. Antibacterial properties and efficacy of LL-37 fragment GF-17D3 and scolopendin A2 peptides against resistant clinical strains of Staphylococcus aureus, Pseudomonas aeruginosa, and Acinetobacter baumannii in vitro and in vivo model studies. Probiotics Antimicrob. Proteins, 2023; ahead-of-print. [Google Scholar] [CrossRef] [PubMed]
- Mourtada, R.; Herce, H.D.; Yin, D.J.; Moroco, J.A.; Wales, T.E.; Engen, J.R.; Walensky, L.D. Design of stapled antimicrobial peptides that are stable, nontoxic and kill antibiotic-resistant bacteria in mice. Nat. Biotechnol. 2019, 37, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Heulot, M.; Jacquier, N.; Aeby, S.; Le Roy, D.; Roger, T.; Trofimenko, E.; Barras, D.; Greub, G.; Widmann, C. The anticancer peptide TAT-RasGAP317-326 exerts broad antimicrobial activity. Front. Microbiol. 2017, 8, 994. [Google Scholar] [CrossRef] [PubMed]
- Vizzarro, G.; Jacquier, N. In vitro synergistic action of TAT-RasGAP317-326 peptide with antibiotics against Gram-negative pathogens. J. Glob. Antimicrob. Resist. 2022, 31, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, T.; Hargraves, S.; Georgieva, M.; Widmann, C.; Jacquier, N. The antimicrobial peptide TAT-RasGAP317-326 inhibits the formation and expansion of bacterial biofilms in vitro. J. Glob. Antimicrob. Resist. 2021, 25, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Hazam, P.K.; Cheng, C.C.; Hsieh, C.Y.; Lin, W.C.; Hsu, P.H.; Chen, T.L.; Lee, Y.T.; Chen, J.Y. Development of bactericidal peptides against multidrug-resistant Acinetobacter baumannii with enhanced stability and low toxicity. Int. J. Mol. Sci. 2022, 23, 2191. [Google Scholar] [CrossRef] [PubMed]
- Dalla Torre, C.; Sannio, F.; Battistella, M.; Docquier, J.D.; De Zotti, M. Peptaibol analogs show potent antibacterial activity against multidrug resistant opportunistic pathogens. Int. J. Mol. Sci. 2023, 24, 7997. [Google Scholar] [CrossRef]
- Zeng, P.; Yi, L.; Xu, J.; Gao, W.; Xu, C.; Chen, S.; Chan, K.F.; Wong, K.Y. Investigation of antibiofilm activity, antibacterial activity, and mechanistic studies of an amphiphilic peptide against Acinetobacter baumannii. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183600. [Google Scholar] [CrossRef]
- Wang, X.; Duan, H.; Li, M.; Xu, W.; Wei, L. Characterization and mechanism of action of amphibian-derived wound-healing-promoting peptides. Front. Cell Dev. Biol. 2023, 11, 1219427. [Google Scholar] [CrossRef]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of antimicrobial peptides. A review. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef] [PubMed]
- Dowzicky, M.J.; Chmelařová, E. Antimicrobial susceptibility of gram-negative and gram-positive bacteria collected from eastern Europe: Results from the tigecycline evaluation and surveillance trial (T.E.S.T.), 2011–2016. J. Glob. Antimicrob. Resist. 2019, 17, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Björstad, A.; Askarieh, G.; Brown, K.L.; Christenson, K.; Forsman, H.; Onnheim, K.; Li, H.N.; Teneberg, S.; Maier, O.; Hoekstra, D.; et al. The host defense peptide LL-37 selectively permeabilizes apoptotic leukocytes. Antimicrob. Agents Chemother. 2009, 53, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- De, Y.; Chen, Q.; Schmidt, A.P.; Anderson, G.M.; Wang, J.M.; Wooters, J.; Oppenheim, J.J.; Chertov, O. LL-37 the neutrophil granule-and epithelial cell-derived cathelicidin utilizes formyl peptide receptor-like 1 (FPRL1) as a receptor to chemoattract human peripheral blood neutrophils monocytes and T cells. J. Exp. Med. 2000, 192, 1069–1074. [Google Scholar] [CrossRef]
- Nijnik, A.; Hancock, R.E.W. Host defense peptides: Antimicrobial and immunomodulatory activity and potential applications for tackling antibiotic-resistant infections. Emerg. Health Threats J. 2009, 2, e1. [Google Scholar] [CrossRef] [PubMed]
- Neshani, A.; Zare, H.; Eidgahi, M.R.A.; Kakhki, R.K.; Safdari, H.; Khaledi, A.; Ghazvini, K. LL-37: A review of antimicrobial profile against sensitive and antibiotic-resistant human bacterial pathogens. Gene Rep. 2019, 17, 100519. [Google Scholar] [CrossRef]
- Esfandiyari, R.; Halabian, R.; Behzadi, E.; Sedighian, H.; Jafari, R.; Fooladi, A.A.I. Performance evaluation of antimicrobial peptide ll-37 and hepcidin and β-defensin-2 secreted by mesenchymal stem cells. Heliyon 2019, 5, e02652. [Google Scholar] [CrossRef]
- Haisma, E.M.; de Breij, A.; Chan, H.; van Dissel, J.T.; Drijfhout, J.W.; Hiemstra, P.S.; El Ghalbzouri, A.; Nibbering, P.H. LL-37-derived peptides eradicate multidrug-resistant Staphylococcus aureus from thermally wounded human skin equivalents. Antimicrob. Agents Chemother. 2014, 58, 4411–4419. [Google Scholar] [CrossRef]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Mietzner, T.A.; Montelaro, R.C. Rational design of engineered cationic antimicrobial peptides consisting exclusively of arginine and tryptophan, and their activity against multidrug-resistant pathogens. Antimicrob. Agents Chemother. 2013, 57, 2511–2521. [Google Scholar] [CrossRef]
- Liu, C.; Shan, B.; Qi, J.; Ma, Y. Systemic responses of multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii following exposure to the antimicrobial peptide cathelicidin-BF imply multiple intracellular targets. Front. Cell Infect. Microbiol. 2017, 7, 466. [Google Scholar] [CrossRef]
- Zhao, H.; Gan, T.X.; Liu, X.D.; Jin, Y.; Lee, W.H.; Shen, J.H.; Zhang, Y. Identification and characterization of novel reptile cathelicidins from elapid snakes. Peptides 2008, 29, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Samuel, R.L.; Massiah, M.A.; Gillmor, S.D. The structure and behavior of the NA-CATH antimicrobial peptide with liposomes. Biochim. Biophy Acta 2015, 1848, 2394–2405. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bishop, B.M.; Juba, M.L.; Devine, M.C.; Barksdale, S.M.; Rodriguez, C.A.; Chung, M.C.; Russo, P.S.; Vliet, K.A.; Schnur, J.M.; van Hoek, M.L. Bioprospecting the American alligator (Alligator mississippiensis) host defense peptidome. PLoS ONE 2015, 10, e0117394. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.A.; Digby, M.R.; Lefévre, C.; Nicholas, K.R.; Deane, E.M.; Williamson, P. Identification characterization and expression of cathelicidin in the pouch young of Tammar wallaby (Macropus eugenii). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 149, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Selsted, M.E.; Novotny, M.J.; Morris, W.L.; Tang, Y.Q.; Smith, W.; Cullor, J.S. Indolicidin is a novel bactericidal tridecapeptide amide from neutrophils. J. Biol. Chem. 1992, 267, 4292–4295. [Google Scholar] [CrossRef] [PubMed]
- Végh, A.G.; Nagy, K.; Bálint, Z.; Kerényi, A.; Rákhely, G.; Váró, G.; Szegletes, Z. Effect of antimicrobial peptide-amide: Indolicidin on biological membranes. J. Biomed. Biotechnol. 2011, 2011, 670589. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Chen, C.; Jou, M.L.; Lee, A.Y.L.; Lin, Y.C.; Yu, Y.P.; Huang, W.T.; Wu, S.H. Structural and DNA-binding studies on the bovine antimicrobial peptide indolicidin: Evidence for multiple conformations involved in binding to membranes and DNA. Nucleic Acids Res. 2005, 33, 4053–4064. [Google Scholar] [CrossRef] [PubMed]
- Marchand, C.; Krajewski, K.; Lee, H.F.; Antony, S.; Johnson, A.A.; Amin, R.; Kvaratskhelia, M.; Pommier, Y. Covalent binding of the natural antimicrobial peptide indolicidin to DNA abasic sites. Nucleic Acids Res. 2006, 34, 5157–5165. [Google Scholar] [CrossRef]
- Knutelski, S.; Awad, M.; Łukasz, N.; Bukowski, M.; Śmiałek, J.; Suder, P.; Dubin, G.; Mak, P. Isolation, identification, and bioinformatic analysis of antibacterial proteins and peptides from immunized hemolymph of red palm weevil Rhynchophorus ferrugineus. Biomolecules 2021, 11, 83. [Google Scholar] [CrossRef]
- Schneider, J.J.; Unholzer, A.; Schaller, M.; Schäfer-Korting, M.; Korting, H.C. Human defensins. J. Mol. Med. 2005, 83, 587–595. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Lu, W. α-Defensins in human innate immunity. Immunol. Rev. 2012, 45, 84–112. [Google Scholar] [CrossRef] [PubMed]
- Wanniarachchi, Y.A.; Kaczmarek, P.; Wan, A.; Nolan, E.M. Human defensin 5 disulfide array mutants: Disulfide bond deletion attenuates antibacterial activity against Staphylococcus aureus. Biochemistry 2011, 37, 8005–8017. [Google Scholar] [CrossRef] [PubMed]
- Marcelino-Pérez, G.; Ruiz-Medrano, R.; Gallardo-Hernández, S.; Xoconostle-Cázares, B. Adsorption of recombinant human β-defensin 2 and two mutants on mesoporous silica nanoparticles and its effect against Clavibacter michiganensis subsp. Michiganensis. Nanomaterials 2021, 11, 2144. [Google Scholar] [CrossRef] [PubMed]
- Schibli, D.J.; Hunter, H.N.; Aseyev, V.; Starner, T.D.; Wiencek, J.M.; McCray, P.B.; Tack, B.F.; Vogel, H.J. The solution structures of the human beta-defensins lead to a better understanding of the potent bactericidal activity of HBD3 against Staphylococcus aureus. J. Biol. Chem. 2002, 277, 8279–8289. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, T.; Spielmann, M.; Zuhaili, B.; Fossum, M.; Metzig, M.; Koehler, T.; Steinau, H.-U.; Yao, F.; Onderdonk, A.B.; Steinstraesser, L.; et al. Human beta defensin-3 promotes wound healing in infected diabetic wounds. J. Gene Med. 2009, 11, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Zerweck, J.; Strandberg, E.; Kukharenko, O.; Reichert, J.; Bürck, J.; Wadhwani, P.; Ulrich, A.S. Molecular mechanism of synergy between the antimicrobial peptides PGLa and magainin 2. Sci. Rep. 2017, 7, 13153. [Google Scholar] [CrossRef]
- Tamba, Y.; Yamazaki, M. Magainin 2-induced pore formation in the lipid membranes depends on its concentration in the membrane interface. J. Phys. Chem. B 2009, 113, 486–4852. [Google Scholar] [CrossRef]
- Maloy, W.L.; Kari, U.P. Structure-activity studies on magainins and other host defense peptides. Biopolymers 1995, 37, 105–122. [Google Scholar] [CrossRef]
- Gottler, L.M.; Ramamoorthy, A. Structure membrane orientation mechanism and function of pexiganan-a highly potent antimicrobial peptide designed from magainin. Biochim. Biophys. Acta 2009, 1788, 1680–1686. [Google Scholar] [CrossRef]
- Ramamoorthy, A.; Thennarasu, S.; Lee, D.K.; Tan, A.; Maloy, L. Solid-state NMR investigation of the membrane-disrupting mechanism of antimicrobial peptides MSI-78 and MSI-594 derived from magainin 2 and melittin. Biophys. J. 2006, 91, 206–216. [Google Scholar] [CrossRef]
- Fuchs, P.C.; Barry, A.L.; Brown, S.D. In vitro antimicrobial activity of MSI-78 a magainin analog. Antimicrob. Agents Chemother. 1998, 42, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Bevier, C.R.; Sonnevend, A.; Kolodziejek, J.; Nowotny, N.; Nielsen, P.F.; Conlon, J.M. Purification and characterization of antimicrobial peptides from the skin secretions of the mink frog (Rana septentrionalis). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 139, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Abraham, B.; Sonnevend, A.; Jouenne, T.; Cosette, P.; Leprince, J.; Vaudry, H.; Bevier, C.R. Purification and characterization of antimicrobial peptides from the skin secretions of the carpenter frog Rana virsgatipes (Ranidae, Aquarana). Reg. Peptides 2005, 13, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Savelyeva, A.; Ghavami, S.; Davoodpour, P.; Asoodeh, A.; Los, M.J. An overview of Brevinin superfamily: Structure function and clinical perspectives. Adv. Exp. Med. Biol. 2014, 818, 197–212. [Google Scholar] [PubMed]
- Popovic, S.; Urbán, E.; Lukic, M.; Conlon, J.M. Peptides with antimicrobial and anti-inflammatory activities that have therapeutic potential for treatment of acne vulgaris. Peptides 2012, 34, 275–282. [Google Scholar] [CrossRef]
- Popovic, S.; Djurdjevic, P.; Zaric, M.; Mijailovic, Z.; Avramovic, D.; Baskic, D. Effects of host defense peptides B2RP Brevinin-2GU D-Lys-Temporin Lys-XT-7 and DLys-Ascaphin-8 on peripheral blood mononuclear cells: Preliminary study. Period. Biol. 2017, 119, 113–118. [Google Scholar] [CrossRef]
- Conlon, J.M.; Demandt, A.; Nielsen, P.F.; Leprince, J.; Vaudry, H.; Woodhams, D.C. The alyteserins: Two families of antimicrobial peptides from the skin secretions of the midwife toad Alytes obstetricans (Alytidae). Peptides 2009, 30, 1069–1073. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2015, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Conlon, J.M.; Prajeep, M.; Mechkarska, M.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; King, J.D. Characterization of the host-defense peptides from skin secretions of Merlin’s clawed frog Pseudhymenochirus merlini: Insights into phylogenetic relationships among the Pipidae. Comp. Biochem. Physiol. Part. D Genom. Proteonomics 2013, 8, 352–357. [Google Scholar] [CrossRef]
- Ali, M.F.; Soto, A.; Knoop, F.C.; Conlon, J.M. Antimicrobial peptides isolated from skin secretions of the diploid frog Xenopus tropicalis (Pipidae). Biochim. Biophys. Acta 2001, 1550, 81–89. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, M.S.; Kim, S.C. A novel antimicrobial peptide from Bufo bufo gargarizans. Biochem. Biophy Res. Commun. 1996, 218, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Apponyi, M.A.; Pukala, T.L.; Brinkworth, C.S.; Maselli, V.M.; Bowie, J.H.; Tyler, M.J.; Booker, G.W.; Wallace, J.C.; Carver, J.A.; Separovic, F. Host-defence peptides of Australian anurans: Structure, mechanism of action and evolutionary significance. Peptides 2004, 25, 1035–1054. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.S.; Ferreira, T.C.G.; Cilli, E.M.; Crusca JR, E.; Mendes-Giannini, M.J.S.; Sebben, A.; Ricart, C.A.O.; Sousa, M.V.; Fontes, W. Hylin a1, the first cytolytic peptide isolated from the arboreal South American frog Hypsiboas albopunctatus (“spotted treefrog”). Peptides 2009, 30, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Ross, N.W.; Mackinnon, S.L. Comparison of the biochemical composition of normal epidermal mucus and extruded slime of hagfish (Myxine glutinosa L.). Fish. Shellfish. Immunol. 2008, 25, 625–632. [Google Scholar] [CrossRef]
- Ellis, A.E. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Silphaduang, U.; Noga, E.J. Peptide antibiotics in mast cells of fish. Nature 2001, 414, 268–269. [Google Scholar] [CrossRef] [PubMed]
- Noga, E.J.; Silphaduang, U.; Park, N.G.; Seo, J.K.; Stephenson, J.; Kozlowicz, S. Piscidin 4, a novel member of the piscidin family of antimicrobial peptides. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 299–305. [Google Scholar] [CrossRef]
- Peng, K.C.; Lee, S.H.; Hour, A.L.; Pan, C.Y.; Lee, L.H.; Chen, J.Y. Five diferent piscidins from Nile tilapia, Oreochromis niloticus: Analysis of their expressions and biological functions. PLoS ONE 2012, 7, e50263. [Google Scholar] [CrossRef]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef]
- Tao, R.; Tong, Z.; Lin, Y.; Xue, Y.; Wang, W.; Kuang, R.; Wang, P.; Tian, Y.; Ni, L. Antimicrobial and antibiofilm activity of pleurocidin against cariogenic microorganisms. Peptides 2011, 32, 1748–1754. [Google Scholar] [CrossRef]
- Lee, J.; Lee, D.G. Structure–antimicrobial activity relationship between pleurocidin and its enantiomer. Exp. Mol. Med. 2008, 40, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Patrzykat, A.; Friedrich, C.L.; Zhang, L.; Mendoza, V.; Hancock, R.E.W. Sublethal concentrations of pleurocidin-derived antimicrobial peptides inhibit macromolecular synthesis in Escherichia coli. Antimicrob. Agents Chemother. 2002, 46, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, D.G. Influence of the hydrophobic amino acids in the N- and Cterminal regions of pleurocidin on antifungal activity. J. Microbiol. Biotechnol. 2010, 20, 1192–1195. [Google Scholar] [CrossRef] [PubMed]
- Hazam, P.K.; Cheng, C.C.; Lin, W.C.; Hsieh, C.Y.; Chen, J.Y. Strategic modification of low-active natural antimicrobial peptides confers ability to neutralize pathogens in vitro and in vivo. Eur. J. Med. Chem. 2022, unpublished. [Google Scholar]
- Hazam, P.K.; Chen, J.Y. Therapeutic utility of the antimicrobial peptide Tilapia Piscidin 4 (TP4). Aquac. Rep. 2020, 17, 100409. [Google Scholar] [CrossRef]
- Krause, A.; Neitz, S.; Mägert, H.J.; Schulz, A.; Forssmann, W.G.; Schulz-Knappe, P.; Adermann, K. LEAP-1, a novel highly disulfide-bonded human peptide, exhibits antimicrobial activity. FEBS Lett. 2000, 480, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J. Biol. Chem. 2001, 276, 7806–7810. [Google Scholar] [CrossRef]
- Barton, J.C.; Acton, R.T. Hepcidin, iron, and bacterial infection. Vitam. Horm. 2019, 110, 223–242. [Google Scholar] [CrossRef]
- Shike, H.; Lauth, X.; Westerman, M.E.; Ostland, V.E.; Carlberg, J.M.; Van Olst, J.C.; Shimizu, C.; Bulet, P.; Burns, J.C. Bass hepcidin is a novel antimicrobial peptide induced by bacterial challenge. Eur. J. Biochem. 2002, 269, 2232–2237. [Google Scholar] [CrossRef]
- Padhi, A.; Verghese, B. Evidence for positive Darwinian selection on the hepcidin gene of Perciform and Pleuronectiform fishes. Mol. Divers. 2007, 11, 119–130. [Google Scholar] [CrossRef]
- Hilton, K.B.; Lambert, L.A. Molecular evolution and characterization of hepcidin gene products in vertebrates. Gene 2008, 415, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.J.; Cai, J.J.; Cai, L.; Qu, H.D.; Yang, M.; Zhang, M. Cloning and expression of a hepcidin gene from a marine fish (Pseudosciaena crocea) and the antimicrobial activity of its synthetic peptide. Peptides 2009, 30, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Neves, J.V.; Caldas, C.; Vieira, I.; Ramos, M.F.; Rodrigues, P.N. Multiple hepcidins in a teleost fish, Dicentrarchus labrax: Different hepcidins for differen roles. J. Immunol. 2015, 195, 2696–2709. [Google Scholar] [CrossRef] [PubMed]
- Fennell, J.F.; Shipman, W.H.; Cole, L.J. Antibacterial action of melittin, a polypeptide from bee venom. Proceed Soc. Exp. Biol. Med. 1968, 127, 707–710. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.V.; de Barros, G.; Pinto, E.G.; Tempone, A.G.; Orsi, R.O.; Dos Santos, L.D.; Calvi, S.; Ferreira, R.S., Jr.; Pimenta, D.C.; Barraviera, B. Melittin induces in vitro death of Leishmania (Leishmania) infantum by triggering the cellular innate immune response. J. Venom. Anim. Toxins Trop. Dis. 2016, 22, 1. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kwon, O.; An, H.J.; Park, K.K. Antifungal effects of bee venom components on Trichophyton rubrum: A novel approach of bee venom study for possible emerging antifungal agent. Ann. Dermatol. 2018, 30, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Chaturvedi, P.K.; Chun, S.N.; Lee, Y.G.; Ahn, W.S. Honeybee venom possesses anticancer and antiviral effects by differential inhibition of HPV E6 and E7 expression on cervical cancer cell line. Oncol. Rep. 2015, 33, 1675–1682. [Google Scholar] [CrossRef]
- Van den Bogaart, G.; Guzman, J.V.; Mika, J.T.; Poolman, B. On the mechanism of pore formation by melittin. J. Biol. Chem. 2008, 283, 33854–33857. [Google Scholar] [CrossRef]
- Rangel, K.; Lechuga, G.C.; Almeida Souza, A.L.; Carvalho, J.P.R.S.; Villas-Bôas, M.H.S.; De Simone, S.G. Pan-drug resistant Acinetobacter baumannii but not other strains are resistant to the bee venom peptide melittin. Antibiotics 2020, 149, 178. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Vaeck, M.; Tempst, P. Apidaecins: Antibacterial peptides from honeybees. EMBO J. 1989, 8, 2387–2391. [Google Scholar] [CrossRef]
- Borovsky, D.; Rougé, P.; Shatters, R.G., Jr. Bactericidal properties of proline-rich Aedes aegypti trypsin modulating oostatic factor (AeaTMOF). Life 2022, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial peptides, a mini review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.; Hultmark, D.; Engström, Å.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Leung, K.; Chen, H.M. The combined effects of antibacterial peptide cecropin a and anticancer agents on leukemia cells. Anticancer Res. 2002, 22, 2811–2816. [Google Scholar] [PubMed]
- Lim, J.; Hong, J.; Jung, Y.; Ha, J.; Kim, H.; Myung, H.; Song, M.J. Bactericidal effect of Cecropin A fused endolysin on drug-resistant gram-negative pathogens. Microbiol. Biotechnol. 2022, 32, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.P.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599–607. [Google Scholar] [CrossRef]
- Burian, M.; Schittek, B. The secrets of dermcidin action. Int. J. Med. Microbiol. 2015, 305, 283–286. [Google Scholar] [CrossRef]
- Zeth, K.; Méndez-Vilas, A. Structure and Mechanism of Human Antimicrobial Peptide Dermcidin and Its Antimicrobial Potential; Formatex Research Center: Badajoz, Spain, 2013; Volume 2. pp. 1333–1342.
- Liu, C.; Qi, J.; Shan, B.; Ma, Y. Tachyplesin causes membrane instability that kills multidrug-resistant bacteria by inhibiting the 3-ketoacyl carrier protein reductase FabG. Front. Microbiol. 2018, 9, 825. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.J.; Liu, Y.Y.; Li, H.; Guo, L.X.; Liu, Z.H.; Shi, X.L.; Hu, M. In vitro potential of Lycosin-I as an alternative antimicrobial drug for treatment of multidrug-resistant Acinetobacter baumannii infections. Antimicrob. Agents Chemother. 2014, 58, 6999–7002. [Google Scholar] [CrossRef]
- Cao, L.; Dai, C.; Li, Z.; Fan, Z.; Song, Y.; Wu, Y.; Cao, Z.; Li, W. Antibacterial activity and mechanism of a scorpion venom peptide derivative in vitro and in vivo. PLoS ONE 2012, 7, e40135. [Google Scholar] [CrossRef] [PubMed]
- Oyama, L.B.; Girdwood, S.E.; Cookson, A.R.; Fernandez-Fuentes, N.; Privé, F.; Vallin, H.E.; Wilkinson, T.J.; Golyshin, P.N.; Golyshina, O.V.; Mikut, R.; et al. The rumen microbiome: An underexplored resource for novel antimicrobial discovery. NPJ Biofilms Microbiomes 2017, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.; Lee, E.; Jeon, D.; Park, Y.G.; Bang, J.K.; Park, Y.S.; Shin, S.Y.; Kim, Y. Peptoid-substituted hybrid antimicrobial peptide derived from papiliocin and magainin 2 with enhanced bacterial selectivity and anti-inflammatory activity. Biochemistry 2015, 54, 3921–3931. [Google Scholar] [CrossRef] [PubMed]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A novel, rationally designed, hybrid antimicrobial peptide, inspired by cathelicidin and aurein, exhibits membrane-active mechanisms against Pseudomonas aeruginosa. Sci. Rep. 2021, 10, 9117. [Google Scholar] [CrossRef] [PubMed]
- Ferre, R.; Melo, M.N.; Correia, A.D.; Feliu, L.; Bardaji, E.; Planas, M.; Castanho, M. Synergistic effects of the membrane actions of cecropin-melittin antimicrobial hybrid peptide BP100. Biophys. J. 2009, 96, 1815–1827. [Google Scholar] [CrossRef]
- Oddo, A.; Thomsen, T.T.; Kjelstrup, S.; Gorey, C.; Franzyk, H.; Frimodt-Møller, N.; Løbner-Olesen, A.; Hansen, P.R. An all-D amphipathic undecapeptide shows promising activity against colistin-resistant strains of Acinetobacter baumannii and a dual mode of action. Antimicrob. Agents Chemother. 2016, 60, 592–599. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem. Pharmacol. 2017, 133, 117. [Google Scholar] [CrossRef]
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moghaddam, M.M.; Mirnejad, R. Antimicrobial peptides: Features action and their resistance mechanisms in bacteria. Microb. Drug Resist. 2018, 24, 747. [Google Scholar] [CrossRef]
- Joo, H.S.; Fu, C.I.; Otto, M. Bacterial strategies of resistance to antimicrobial peptides. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150292. [Google Scholar] [CrossRef]
- Omardien, S.; Brul, S.; Zaat, S.A.J. Antimicrobial activity of cationic antimicrobial peptides against gram-positive: Current progress made in understanding the mode of action and the response of bacteria. Front. Cell Dev. Biol. 2016, 4, 111. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Tomida, J.; Kawamura, Y. MexXY multidrug efflux system of Pseudomonas aeruginosa. Front. Microbiol. 2012, 3, 408. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, B.; Gorr, S.U. Antimicrobial peptides: Mechanisms of action and resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Jeannot, K.; Bolard, A.; Plésiat, P. Resistance to polymyxins in gram-negative organisms. Int. J. Antimicrob. Agents 2017, 49, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Harris, P.N.A. Colistin resistance: A major breach in our last line of defense. Lancet Infect. Dis. 2016, 16, 132–133. [Google Scholar] [CrossRef] [PubMed]
- Macnair, C.R.; Stokes, J.M.; Carfrae, L.A.; Fiebig-Comyn, A.A.; Coombes, B.K.; Mulvey, M.R.; Brown, E.D. Overcoming mcr-1 mediated colistin resistance with colistin in combination with other antibiotics. Nat. Commun. 2018, 9, 458. [Google Scholar] [CrossRef]
- Gheorghe, I.; Saviuc, C.; Ciubuca, B.; Lazar, V.; Chifiriuc, M.C. Chapter 8—Nano drug delivery. In Nanomaterials for Drug Delivery and Therapy; Grumezescu, A.M., Ed.; William Andrew Publishing: Norwich, NY, USA, 2019; pp. 225–244. ISBN 978-0-12-816505-8. [Google Scholar]
- Sun, B.; Wibowo, D.; Middelberg, A.P.J.; Zhao, C.X. Cost-effective downstream processing of recombinantly produced pexiganan peptide and its antimicrobial activity. AMB Express 2018, 8, 6. [Google Scholar] [CrossRef]
- Falciani, C.; Lozzi, L.; Pollini, S.; Luca, V.; Carnicelli, V.; Brunetti, J.; Lelli, B.; Bindi, S.; Scali, S.; Di Giulio, A.; et al. Isomerization of an antimicrobial peptide broadens antimicrobial spectrum to gram-positive bacterial pathogens. PLoS ONE 2012, 7, e46259. [Google Scholar] [CrossRef]
- Hirt, H.; Gorr, S.U. Antimicrobial peptide GL13K is effective in reducing biofilms of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 4903–4910. [Google Scholar] [CrossRef]
- de la Fuente-Núñez, C.; Cardoso, M.H.; de Souza Cândido, E.; Franco, O.L.; Hancock, R.E. Synthetic antibiofilm peptides. Biochim. Biophys. Acta 2016, 1858, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Kraus, D.; Peschel, A. Molecular mechanisms of bacterial resistance to antimicrobial peptides. Curr. Top. Microbiol. Immunol. 2006, 306, 231–250. [Google Scholar] [CrossRef]
- Verma-Gaur, J.; Qu, Y.; Harrison, P.F.; Lo, T.L.; Quenault, T.; Dagley, M.J.; Bellousoff, M.; Powell, D.R.; Beilharz, T.H.; Traven, A. Integration of posttranscriptional gene networks into metabolic adaptation and biofilm maturation in Candida albicans. PLoS Genet. 2015, 11, e1005590. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.H. Lipopolysaccharide: Basic biochemistry, intracellular signaling, and physiological impacts in the gut. Intest. Res. 2014, 12, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.; Atilano, M.L.; Pombinho, R.; Covas, G.; Gallo, R.L.; Filipe, S.R.; Sousa, S.; Cabanes, D. L-Rhamnosylation of Listeria monocytogenes wall teichoic acids promotes resistance to antimicrobial peptides by delaying interaction with the membrane. PLoS Pathog. 2015, 11, e1004919. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, M.R.; Casella, L.G.; Jones, J.W.; Adams, M.D.; Zurawski, D.V.; Hazlett, K.R.; Doi, Y.; Ernst, R.K. Unique structural modifications are present in the lipopolysaccharide from colistin-resistant strains of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2013, 57, 4831–4840. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Ortega, C.; Olivares, J.; Martínez, J.L. RND multidrug efflux pumps: What are they good for? Front. Microbiol. 2013, 4, 7. [Google Scholar] [CrossRef]
- Delmar, J.A.; Su, C.C.; Yu, E.W. Bacterial multidrug efflux transporters. Annu. Rev. Biophys. 2014, 43, 93–117. [Google Scholar] [CrossRef]



| Mechanism of AMP | Mode of Action | Reference |
|---|---|---|
| Direct killing: Membrane target | Electrostatic interactions and hydrophobic interactions (peptide and bacterial cell surface), membrane rupture-bound types of machinery and bacterial lysis—bilayer disruption | [54,55,56,57,58,59] |
| Direct killing: Non-membrane target | Action on the bacterial cell wall, activation of autolysin, intracellular targets: inhibition of protein/nucleic acid synthesis, disruption of enzymatic activities and bacterial lysis | [54,55,56,57,60] |
| Immune modulation | Chemotaxis, activation of immunocytes, microbial killing; anti-endotoxin activity, suppression of toll-like receptors (TLRs) and/or cytokine-mediated production of proinflammatory cytokines and preventing excessive and harmful proinflammatory responses, controls the inflammation | [57,61,62] |
| Peptide | Source | Sequence (nº Amino Acid) | Structure | MIC against A. baumannii (μg/mL) | Ref. | |
|---|---|---|---|---|---|---|
| ATBS | MDR | |||||
| Agelaia-MPI | Agelaia pallipes pallipes | INWLKLGKAIIDAL (14aa) | AH | 6.25 | 12.5–25 | [81] |
| aHylin a1-15K | Hypsiboas albopunctatus (American frog) | IAKAILPLALKALKKLIK-NH2 (19aa) | AH | 1–2 * | 1–2 * | [82] |
| Alytesirin-1c | Frog skin peptide | GLKEIFKAGLGSLVKGIAAHVAS-NH2 (23aa) | AH | — | 11.3–22.6 | [83] |
| α-Helical-26 (A12L/A20L) | D- and L-diastereomeric peptides | Ac KWKSFLKTFKSLKKTVLHTL LKAISS-NH2 (26aa) | AH | — | 0.5–1.0 | [84] |
| AM-CATH21 | GLFKKLRRKIKKGFKKIFKRL (21aa) | AH | 42 | 10 | [85] | |
| AM-CATH28 | American alligator | KIKKGFKKIFKRLPPIGVGVSIPLAGKR (28aa) | AH | 28 | 10 | |
| AM-CATH36 | GLFKKLRRKIKKGFKKIFKRLPPIG VGVSIPLAGKR (36aa) | AH | 5.2 | 5.2 | ||
| Am23SK | Alligator mississippiensis | SCRFSGGYCIWNWERCRSGHFLVALCPFRKRCCK (34aa) | AH | — | 2 | [86] |
| Artlysin Art-175 | Pseudomonas aeruginosa bacteriophage | Comprises a modified variant of endolysin KZ144 with an N-terminal fusion to SMAP-29 | NF | — | 4–20 | [87] |
| Aurein 1,2 | Frog skin peptide | GLFDIIKKIAESF (13aa) | AH | 16 | — | [88] |
| Bactenecin | Bovine neutrophil granules, Caprine | LCRIVVIRVCR (12aa) | B-turn structure Ciclyc | 64 | — | [89,90,91] |
| Bicarinalin (YRTX-Tb1a) | Tetramorium bicarinatum venom | KIKIPWGKVKDFLVGGMKAV (20aa) | AH | — | 4 | [92] |
| BMAP-27 | Bovine myeloid | GRFKRFRKKFKKLFKKLSPVIPLLHLG (27aa) | AH | 8–16 | 4–16 | [93] |
| BmKn1 | Heterometrus petersii (Scorpion venom gland) | FIGAVAGLLSKIF (13aa) | AH | >40 | — | [94] |
| BmKn2 | Mesobuthus martensii Karsch (Scorpion) | FIGAIARLLSKIF-NH2 (13aa) | AH | 10 | 5–10 | [95] |
| B2RP-Era | Frog skin peptide | GVIKSVLKGVAKTVALGML-NH2 (19aa) | AH | 8–32 | 8–64 | [96,97] |
| Brevinina 2 (B2RP) | Frog skin peptide | GIWDTIKSMGKVFAGKILQNL-NH2 (21aa) | AH | 29 | 7–13.9 | [98] |
| BR003-cecropin A | Aedes aegypti | GGLKKLGKKLEGAGKRVFNAAEK ALPVVAGAKALRK (36aa) | AH | 5 | 5 | [99] |
| Buforin II | Frog skin peptide | TRSSRAGLQFPVGRVHRLLRK (21aa) | AH | 8–19.5 | 0.25–39 | [100,101,102,103] |
| Caerin 1.1 | GLLSVLGSVAKHVLPHVVPVIAEHL-NH2 (25aa) | AH | 7.5 | — | [104] | |
| Caerin 1.9 | Australian tree frog | GLFGVLGSIAKHVLPHVVPVIAEKL-NH2 | 3.75 | — | ||
| Caerin 1.1 + Caerin 1.9 | GLLSVLGSVAKHVLPHVVPVIAEHL-NH2+ GLFGVLGSIAKHVLPHVVPVIAEKL-NH2 | 0.9375–1.875 | — | |||
| CATH-BF derivative (Cath-A and OH- | Bungarus fasciatus (Snake venom) | KFFRKLKKSVKKRAKEFFKKPRVI GVSIPF(30aa) | AH | — | 8–32 | [105] |
| Cecropin A | Hyalophora cecropia (Cecropia moth) | KWKLFKKIEKVGQNIRDGIIKAGP AVAVVGQATQIAK (37aa) | AH | 32 | 0.5–32 | [106,107] |
| Cecropin P1 | Ascaris suum (Pig) | SWLSKTAKKLENSAKKRISEGIAIA IQGGPR (31aa) | AH | 1.6 | — | [106,108] |
| Citropin 1.1. | Litora genus (Australian tree frog) | GLFDVIKKVASVIGGL-NH2 (16aa) | AH | 16 | — | [88] |
| CL defensin | Cimex Lectularius (Bedbug) | ATCDLFSFQSKWVTPNHAACAAHCTARGNRGGRCKKAVCHCRK (43aa) | AH, antiparallel BS; N-terminal loop | — | — | [109] |
| Colistin (Polymyxin E) | Bacillus colistinus | C52H98N16O13 (cyclic compound) | BS n and B-tur | Antibiofilm, side effects | — | [110] |
| Con10 | Opisthacanthus cayaporum (Scorpion venoms) | FWSFLVKAASKILPSLIGGGDDNKSSS (27aa) | AH | 12.5 | 12.5 | [81] |
| CPF-AM1 | Frog skin peptide | GLGSVLGKALKIGANLL (19aa) | AH | 16–128 | 4–128 | [96,111,112] |
| CPF-B1 | Frog skin peptide | GLGSLLGKAFKIGLKTVGKMMGGAPREQ (28aa) | AH | — | 11.4–22.8 | [113] |
| CPF-C1 | Frog skin peptide | GFGSLLGKALRLGANVL (17aa) | AH | 5 | — | [112] |
| Ctriporin | Heterometrus petersii (Scorpion venom gland) | FLWGLIPGAISAVTSLIKK (19aa) | AH | 20 | 20–40 | [94] |
| Cy02 (cyclotide) | Viola odorata | GIPCGESCVWIPCISSAIGCSCKSKVCYRN (30aa) | BSs | — | 15 * | [114] |
| Danalexin | American bulfrog (Rana catesbeiana) | LGGLIKIVPAMICAVTKKC (19aa) | AH | — | 4–16 | [115] |
| DCD-1 L | Eccrine sweat glands | SSLLEKGLDGAKKAVGGLGKLGKDAVEDLESVGKGAVHDVKDVLD SVL (48aa) | AH | 16 | — | [116,117] |
| D-150-177C, HBcARD derivative peptide | Hepatitis B virus | RRRGRSPRRRTPSPRRRRSQSPRR RRSC (28aa) | AH | 16 | 16–32 | [118] |
| Delfibactin A | Gram-negative bactéria Delfia spp. | C40H68N14O18 | NF | — | 16 | [119] |
| [D4K] B2RP | Frog skin peptide | GIWKTIKSMGKVFAGKILQNL-NH2 (21aa) | AH | 4–16 | 4–16 | [96,120] |
| D-Myrtoxin-Mp 1a (Mp1a) | Myrmecia pilosula (Venom) | IDWKKVDWKKVSKKTCKVMXKACKEL-NH2 (26aa) | AH | 0.025 * | — | [121] |
| DOH-CATH30 | King cobra (Snake venon) | KFFKKLKNSVKKRAKKFFKKPRVIGVSIPF (30aa) | AH | — | 1.56–12.5 | [122] |
| [E4k] Alytesirin-1c | Frog skin peptide | GLKEIFKAGLGSLVKGIAAHVAS-NH2 (23aa) | AH | 4–16 | 4–16 | [96,120] |
| [E6k,D9k] Hymenochirin-1B | Frog skin peptide | LKLSPKTKDTLKKVLKGAIKGAIA IASMA-NH2 (29aa) | AH | — | 4.9 | [123] |
| Epi-122–42 | Epinephelus coioides (Orange-spotted grouper) | GFIFHIIKGLFHAGKMIHGLV (21aa) | NF | — | 4–32 | [124] |
| Epsilon-poly L-lysine (EPL)-catechol | Streptomyces albulus derived | Complex | NF | — | Reducing bacterial burden in vivo | [125] |
| Esc(1-21) | Frog-skin | GIFSKLAGKKIKNLLISGLKG-NH2 (21aa) | AH | — | 17.5–35 | [126] |
| Esc(1-21) + Colistina | GIFSKLAGKKIKNLLISGLKG-NH2 (21aa) + Colistina | AH | — | 1.1–4.4 | ||
| FLIP 7 | Calliphora vicina (Medicinal Maggots) | ATCDLLSGTGANHSACAAHCLLRGNRGGYCNGKAVCVCRN (40aa) | AH | — | 125–416 biofilm bactéria sensitivity | [127] |
| [G4K] XT7 | Frog skin peptide | GLLGPLLKIAAKVGSNLL-NH2 (18aa) | AH | 4–32 | 4–64 | [96,128] |
| Glatiramer acetate | Homo sapiens | EAYKAAEKAYAAKEAAKEAAKAKAEKKAAYAKAKAAKYEKKAKKAAAEYKKK (52aa) | NF | Reduct viable cells | Reduct viable cells | [129] |
| HBD-2 | The epithelial lining of the respiratory/urinary tracts | GIGDPVTCLKSGAICHPVFCPRRYKQIGTCGLPGTKCCKKP (41aa) | Beta | 3.90–9.35 | 3.25–4.5 | [130] |
| HBD-3 | The epithelial lining of the respiratory/urinary tracts | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK (45aa) | AH + BS | 4 | 4 | [131] |
| HBD-3 | Epithelial cells | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK (45aa) | AH + BS | — | 4–16 | [124] |
| HD5d5 | Homo sapiens (Polymorphonuclear neutrophil) | ARARCRRGRAARRRRLRGVCRIRGRLRRLAAR (32aa) | AH | 40 | 40 | [132] |
| Histatin-8 | Homo sapiens | KFHEKHHSHRGY (12aa) | AH | 8 | — | [133] |
| HNP-1 | Homo sapiens (Polymorphonuclear | ACYCRIPACIAGERRYGTCIYQGRLWAFCC (30aa) | AH | 50 | — | [106] |
| HNP-2 | (neutrophil) | CYCRIPACIAGERRYGTCIYQGRLWAFCC (29aa) | 50 | — | [106] | |
| Hp1404 | Heterometrus petersii (Scorpion venom gland) | GILGKLWEGVKSIF (14aa) | AH | 5 | 5–10 | [94,134] |
| Hp1404 analogs | GILGKLWEGVKSIF (14aa) analogs | 3.13–25 * | — | |||
| Hp l404 analogs (A, K, V, L, I, W) | Heterometrus petersii (Scorpion venom gland) | GILGKLWEGVKSIF-NH2 (14aa) | AH | 3.13–12.5 | 3.13–16.25 | [134,135] |
| Hylin a1 | Hypsiboas albopunctatus (American frog) | IFGAILPLALGALKNLIK-NH2 (18aa) | AH | 2 * | 2–8 * | [82] |
| Hylin a1-11K | IAKAILPLALKALKNLIK-NH2 (19aa) | 1–2* | 1–2 * | |||
| Hymenochirin-1 Pa | Frog skin peptide | LKLSPKTKDTLKKVLKGAIKGAIAIASMA-NH2 (29aa) | AH | — | — | [136] |
| Im4 | Heterometrus petersii (Scorpion venom gland) | FIGMIPGLIGGLISAIK (17aa) | AH | >40 | — | [94] |
| Im5 | FLGSLFSIGSKLLPGVIKLFQRKKQ (25aa) | AH | 2.5 | 2.5–10 | ||
| Indolicidin | Cytoplasmic granules of the bovine neutrophils | LPWKWPWWPWRR-NH2 (13aa) | Other structure | 4 | 2–64 | [89,106,133] |
| KS-12 | KRIVQRIKDFLR (12aa) | AH | 256 | 64–256 | [137] | |
| KR-20 | Homo sapiens | KRIVQRIKDFLRNLVPRTES (20aa) | 64 | 16–32 | ||
| KR-30 | KSKEKIGKEFKRIVQRIKDFLRNLV PRTES (30aa) | 16 | 8–16 | |||
| Lactoferrin (Lf) | Camel (Colostrum milk) | Large protein | complex | — | Significant clearance of A. baumannii | [138] |
| Lactoperoxidase (Lpo) | rates in lung | |||||
| Latarcin 2a | Pleuronectes americanos (Winter flounder) | H-GLFGKLIKKFGRKAISYA VKKARGKH-OH (26aa) | AH | 16 | 8–64 | [93] |
| LL-37 | Homo sapiens | LLGDFFRKSKEKIGKEFKRIVQRIK DFLRNLVPRTES (37aa) | AH | 32 | 16–32 | [137,139] |
| LS-AMP-E1 | Lycosa sinensis (Chinese wolf spider) | AGMKNIIDAIKKKLGGKL (18aa) | AH | — | 25–100 * | [140] |
| LS-AMP-F1 | TGLGKIGYLMKKLLSKAKV (19aa) | AH | — | 3.1–12.5 * | [141] | |
| LS-sarcotoxin | Lucilla serricata | GWLKKIGKKIERVGQHTRDATIQTIGVAQQAANVAATLK-NH2 (39aa) | AH | 4 | 4–8 | [141] |
| LS-stomoxyn | GFRKRFNKLVKKVKHTIKETANVSKDVAIVAGSGVAVGAAM-NH2 (41aa) | AH | 8 | 4–16 | ||
| Lynronne-1 | Bovine rumen microbiome | LPRRNRWSKIWKKVVTVFS (19aa) | AH | 4 | — | [142] |
| Magainin-1 | Frog skin peptide | GIGKFLHSAGKFGKAFVGEIMKS (23aa) | AH | — | 256 | [100,101] |
| Magainin-2 | Frog skin peptide | GIGKFLHSAKKFGKAFVGEIMNS (23aa) | AH | 9.8–64 | 4.9–64 | [100,101,143] |
| Mastoparan | Vespula lewisii (Hornet venom) | INLKALAALAKKIL (14aa) | AH | 4 | — | [106,144,145] |
| Mastoparan-AF (EMP-AF) | Vespa affinis (Hornet venom) | INLKAIAALAKKLF-NH2 (14aa) | AH | 2–16 | 2–16 | [146] |
| Mastoparan-Chitosan Nanoconstruct | Vespula lewissi (Wasp venom) | INLKALAALAKKIL-NH2 (14aa) | AH | — | 2–4 | [147] |
| Maximin H2 | Oreochromis niloticus (Nile Tilapia) | ILGPVLSMVGSALGGLIKKI-NH2 (20aa) | AH | 64 | 16–128 | [93] |
| Mdc | Housefly larvae | GWLKKIGKKIERVGQHTRDATIQ TIGVAQQANAVAATLKG (40aa) | D-helix | 4 | 4 | [148] |
| Melittin | Apis mellifera (European honeybee) | GIGAVLKVLTTGLPALISWIKRKRQQ (26aa) | AH | 0.25–4 | 0.25–25 | [106,149,150] |
| Melittin with colistin (COL) | Apis mellifera (European honeybee) | GIGAVLKVLTTGLPALISWIKRKR QQ (26aa) + COL | AH | 0.37–0.5 | 0.19–0.37 | [151] |
| Melittin with imipenem (IPM) | GIGAVLKVLTTGLPALISWIKRKR QQ (26aa) + IPM | AH | 0.31–0.37 | 0.12–0.25 | ||
| Mini-ChBac7.5 Nα | Capra hircus (Domestic goat) | RRLRPRRPRLPRPRPRPRPRPR (22aa) | AH | — | 2 * | [152] |
| Mini-ChBac7.5 Nβ | RRLRPRRPRLPRPRPRPRPRP (21aa) | AH | — | 4 * | ||
| Myxinidin 2 | Myxine glutinosa L (Atlantic hagfish) | KIKWILKYWKWS (12aa) | AH | — | 12.5 | [153] |
| Myxinidin 3 | RIRWILRYWRWS (12aa) | BS | — | 6.3 | ||
| N10 | Blood biopanning | ACKDVNTSMCGGK (13aa) | AH | 500 | 500 | [154] |
| NA-CATH | Naja atra (Snake venom) | KRFKKFFKKLKNSVKKRAKKFFKK PKVIGVTFPF (34aa) | AH | 10 | 10 | [85] |
| NB2 | Biofilm biopanning | ACERSIRTVCGGK (13aa) | AH | 500 | 500 | [154] |
| NDBP5.8 | Opisthacanthus cayaporum (Scorpion venoms) | GILGKIWEGVKSLI (14aa) | AH | >25 | >25 | [81] |
| Nisin | Lactococcus lactis (Probiotic bacterium) | MSTKDFNLDLVSVSKKDSGASPRITSISLCTPGGKTGALNGCNMKTATCHCSIHVSK (34aa) | NF | 128 | 64–128 | [155] |
| Nisin + P10 combined | Lactococcus lactis (Probiotic bacterium) + Synthetic derivated | MSTKDFNLDLVSVSKKDSGASPRITSISLCTPGGKTGALNGCNMKTATCHCSIHVSK (34aa) + LAREYKKIVEKLKRWLRQVLRTLR (24aa) | NF | 32 | 16–32 | |
| Nodule-specific cysteine-rich (NCR) peptide and its derivatives | Medicago trunculata | RNGCIVDPRCPYQQCRRPLYCRRR (24aa) | AH | 1.6–25 MBC | — | [156,157] |
| NRC12 | Flatfish Genes | GWKKWFNRAKKVGKTVGGLAVDHYL-NH2 (25aa) | AH | 16 | 8–32 | [93] |
| Nuripep 1653 | Derived from the P54 nutrient reservoir protein (aa 271–292) pea protein from Pisum sativum | VRGLAPKKSLWPFGGPFKSPFN (22aa) | AH | — | 12 | [158] |
| OH-CATH30 | King cobra (Snake venom) | KFFKKLKNSVKKRAKKFFKKPRVI GVSIPF(30aa) | AH | 10 | 10 | [122] |
| Pexiganan | Frog skin peptide | GIGKFLKKAKKFGKAFVKILKK (22aa) | AH | 1–8 | 1–8 | [88,159,160] |
| PGLa-AM1 | Frog skin peptide | GMASKAGSVLGKVAKVALKAAL-NH2 (22aa) | AH | 16–128 | 16–128 | [96,161] |
| Pilosulin | Ant venom (toxin pilosulin) | GLGSVFGRLARILGRVIPKV-NH2 (20aa) | AH | 16 | 8–16 | [93] |
| Pleurocidin | Pleuronectes americanus (Winter flounder) | GWGSFFKKAAHVGKHVGKAALTHYL-NH2 (25aa) | AH | 16 | 8–32 | [93] |
| Polydin-I | Polybia dimorpha (Social wasp) | AVAGEKLWLLPHLLKMLLTPTP (22aa) | AH | >25 | >25 | [81] |
| Polybia-MPII | Pseudopolybia vespiceps testacea | INWLKLGKMVIDAL (14aa) | AH | 12.5 | 25 | [81] |
| Protegrin-1 | Cimex lectularius | RGGRLCYCRRRFCVCVGR-NH2 (18aa) | AH | — | 2–8 | [162] |
| P307SQ-8C | Hepatitis B virus | NAKDYKGAAAEFPKWNKAGGRV LAGLVKRRKSQSRESQC (39aa) | NF | 125 | 62.5–125 | [163] |
| Ranalexin | Rana catesbeiana (American bulfrog) | LGGLIKIVPAMICAVTKKC (19aa) | AH | — | 4–18 | [115] |
| SAAP-148 | Homo sapiens | LKRVWKRVFKLLKRYWRQLKKPVR (24aa) | AH | — | 6 | [164] |
| Spiniferin | Heterometrus petersii (Scorpion venom gland) | ILGEIWKGIKDIL (13aa) | AH | >40 | — | [94] |
| [S7K, G11K] Alytesirrin-2a | Frog skin peptide | ILGKLLKTAAKLLSNL-NH2 (16aa) | AH | — | 8 | [165] |
| SMAP29 | Sheep myeloid | RGLRRLGRKIAHGVKKYGPTVLRIIRIAG (29aa) | AH | 8 | 4–32 | [93] |
| Tachyplesin III | Tachypleus gigas and Carcinoscorpius rotundicauda (Horseshoe crabs) | KWCFRVCYRGICYRKCR-NH2 (17aa) | BS 2 dissulfite bridges | — | 8–16 | [120] |
| Temporin A | Rana temporaria (European red frog) | FLPLIGRVLSGIL-NH2 (13aa) | AH | 128 | — | [88] |
| TP4 | Oreochromis niloticus (Nile tilapia) | FIHHIIGGLFSAGKAIHRLIRRRRR (25aa) | AH | 16 | 8–32 | [93] |
| Venon cocktail proteins | Leiurus quinquestriatus (Scorpion venom) | Cocktail | NF | — | 50.6% of inhibition (20 mg/mL of venom) | [166] |
| VsCT1 | Heterometrus petersii (Scorpion venom gland) | FLKGIIDTVSNWL (13aa) | AH | >40 | — | [94] |
| WAM-1 | Macropus eugenii (Tammar wallaby) | KRGFGKKLRKRLKKFRNSIK KRLKNFNVVIPIPLPG (36aa) | AH | 8.12 | 4–64 | [89,167] |
| WLBU2- arginine-rich amphiphilic peptide | Skin wounds | RRWVRRVRRWVRRVVRVVRRWVRR (24aa) | NF | ~7.484 | ~7.484 | [168] |
| ZY4 cathelicidin-BF-15 derived | Bungarus fasciatus (Snake venom) | VCKRWKKWKR KWKKWCV-NH2 (17aa) | Cyclic SH-bridge | — | 4.6–9.4 | [169] |
| Peptide | Source | Sequence (nº Amino Acid) | Structure | MIC against A. baumannii (μg/mL) | Ref. | |
|---|---|---|---|---|---|---|
| ATBS | MDR | |||||
| AS-CATH8 | Synthetic peptide | KRVNWAKVGRTALKLLPYIFG (21aa) | AH | 0.6 | — | [86] |
| BmKn2-7 | FIKRIARLLRKIF-NH2 (13aa) | AH | 5 | 5–10 | [95] | |
| BmKn2-7R | Synthetic peptide | FIRRIARLLRRIF-NH2 (13aa) | AH | 2.5 | 2.5–5 | |
| BmKn2-7K | FIKKIAKLLKKIF-NH2 (13aa) | AH | 2.5 | 2.5–5 | ||
| BP100 | KKLFKKILKYL (11aa) | AH | — | 4 | [170] | |
| BP214 | Hybrid peptide | KKLFKKILRYL (11aa) | AH | 2 | — | |
| BP214 analogs | KKLFKKILRYL (11aa) analogs | AH | >64 | — | ||
| CA(1–8)-ME(1–12) (CAME) | Chimeric peptide | KWKLFKKIGIGAVLKVLTTG-NH2 (20aa) | AH | 3.12 | 3.12–12.5 | [171] |
| CA(1–8)-MA(1–12) (CAMA) | KWKLFKKIGIGKFLHSAKKF-NH2 (20aa) | AH | 12.5 | 3.12–12.5 | ||
| Cecropin-4 | Synthetic peptide | GWLKKIGKKIERVGQNTRDATIQ AIGVAQQAANVAATLKG (40aa) | AH | 4 | 4 | [172,173] |
| Cecropin A (1–8) melittin (1–10) (CAME) | Hybrid peptide | KWKLFKKIGIGAVLKVLTTG-NH2 (20aa) | AH | 32 | 8–32 | [93] |
| Ceragenins; CSA-192; CSA-131; D-150-177C; HBcARDderivative | Cholic acid synthetic mimics | Steroids compounds | NF | — | — | [174] |
| Chex1-Arg20 amide (ARV-1502) | NA | RPNKPRPYLPRPRPPRPVR-NH2 (19aa) | NF | — | Reduction of bacterial load | [175] |
| D-AP19 | Hybrid peptide | RLFRRVKKVAGKIAKRIWK-NH2 (19aa) | NF | 7.81 | 3.91–15.63 | [176] |
| DGL 13K | Synthetic derived D-enantiomers of GL13K derived from the salivary protein BPIFA2 | GKIIKLKASLKLL-NH2 (13aa) | NF | — | 8–32 | [177] |
| D-Mt6 | Synthetic peptide | KFKKTAKWLIKSAWLLLKSLALKMK (25aa) | AH | 8 | — | [178] |
| DP7 | Computationally designed | VQWRIRVAVIRK (12aa) | AH | — | 4–16 | [179,180,181] |
| ECPep-D | Synthetic peptide | RPFTRAQWFAIQHISPRTIAMRAINNYRWR (30aa) | NF | 37.57 | — | [182] |
| ECPep-2D-Orn | Synthetic peptide | OPFTOAQWFAIQHISPOTIAMOAINNYOWO (30aa) | NF | 17.53 | — | [182] |
| GW-A2 | GAKYAKIIYNYLKKIANALW (20aa) | AH | 32 | 8–32 | [93] | |
| GW-H1a | Synthetic peptide | GYNYAKKLANLAKKFANALW-NH2 (20aa) | AH | 32 | 8–32 | |
| GW-Q6 | GIKIAKKAITIAKKIAKIYW (20aa) | AH | 16 | 8–16 | ||
| HP(2–9)-MA(1–12) (HPMA) | Chimeric peptide | AKKVFKRLGIGKFLHSAKKF-NH2 (20aa) | AH | 6.25 | 3.12–6.25 | [171] |
| HP(2–9)-ME(1–12) (HPME) | AKKVFKRLGIGAVLKVLTTG (20aa) | AH | 6.25 | 3.12–12.5 | ||
| I16K-piscidin-1 and analogs | Hybrid striped bass Morone saxatilis x M. chrysops | FFHHIFRGIVHVGKTIHRLVTG (22aa) | NF | — | 3.1 | [183] |
| IKR18 | Computationally designed | IKRQYKRFFKLFKWFLKK (18aa) | AH | 1 | — | [184] |
| LJ-hep2(66–86) | Synthetic peptide | IKCKFCCGCCTPGVCGVCCRF (21aa) | NF | — | 1.5–3 | [185] |
| LyeTx I-bPEG | Synthetic peptide | WLTALKFLGKNLGKLAKQQCAKL (PEG) (24aa) | AH | — | — | [186] |
| mCM11, cecropin–melittin 11 | Synthetic peptide | NH2-WRLFRRILRVL-NH2 (11aa) | AH | 32 | <4–>512 | [187] |
| MSI-78 | Synthetic peptide, magainin analog | GIGLPLLLALLPGLAPVLILLL-NH2 (22aa) | AH | — | 5 | [188] |
| Mt6 | Synthetic peptide | KKFKKTAKWLIKSAWLLLKSLALKMK (26aa) | AH | 8 | — | [178] |
| NCR169C and its substitution derivatives | Synthetic peptide | KSKKPLFKIWKCVENVCVLWYK | AH | 1.6–12.5 MBC | — | [189] |
| Octominin, Octominin-CNPs | Synthetic derived, defensin 3 of Octopus minor | GWLIRGAIHAGKAIHGLIHRRRH (23aa) | AH | — | 5 | [190,191] |
| Octopromycin | Synthetic peptide | N-RRLIRTDTGPIIYDYFKDQLLKKGMVI LRESMKNLKGM-C (38aa) | AH | — | 50 | [192] |
| OG1410 | ApoE-based synthetic peptide | acetyl-ASAib-LRKL-Aib-KRLL-amide | AH | 16 | 16 | [193] |
| Omega 76-shuft1 | Computationally designed | AFLLKKKKGIIFFEKAKKGK (20aa) | AH | — | 4–16 | [194] |
| Omiganan | Synthetic peptide | ILRWPWWPWRRK-NH2 (12aa) | AH | 32 | — | [88] |
| r-Omiganan | KRRWPWWPWRLI-NH2 (12aa) | AH | 16 | — | ||
| OMN6 | Synthetic peptide | H-M-C-KWKLFKKIEKVGQNIRDGIIKA-GP-AVAVVGQATQIAK-C-NH2 (40aa) | AH | 8 | 4–8 | [195,196] |
| ′Ω17 family peptides | Computationally designed | RKKAIKLVKKLVKKLKKALK (20aa) | AH | 2 | 1–8 | [194] |
| ′Ω76 family peptides | FLKAIKKFGKEFKKIGAKLK (20aa) | AH | 4 | 2–8 | ||
| P10 | Synthetic derivated | LAREYKKIVEKLKRWLRQVLRTLR (24aa) | NF | 4 | 8–32 | [155] |
| P10 + Nisin combined | Synthetic derivated + Lactococcus lactis (Probiotic bacterium) | LAREYKKIVEKLKRWLRQVLRTLR (24aa) + MSTKDFNLDLVSVSKKDSGASPRITSISLCTPGGKTGALNGCNMKTATCHCSIHVSK (34aa) | NF | 1 | 4–16 | |
| PapMA | Hybrid peptide | RWKIFKKIPKFLHSAKKF-NH2 (18aa) | AH | 32 | 16–32 | [197] |
| pepD2 | Computationally designed | WKKLKKLLKKLKKL-NH2 (14aa) | AH | 8 | - | [198] |
| PLP-3 | Synthetic peptides derived from the innate immune system of vertebrates | ~RRPVCVVPLPRVPCLRRR~ | B- hairpin | 1–2 | 1–2 | [199] |
| PNA (RXR)4 XB | Peptide nucleic acid conjugated to (RXR)4 Phosphorodiamidate Morpholino Oligomers | RXRRXRRXRRXRXB (14aa) | NF | — | 1.25 * | [200] |
| Pro9-3 | Computationally designed | RLWLAIWRR-NH2 (9aa) | AH | 16 | 8–64 | [201] |
| Pro9-3D | RLWLAIWRR-NH2 (9aa) | AH | 8 | 4–16 | ||
| RR | Computationally designed | WLRRIKAWLRR (11aa) | AH | — | 25–99 | [202,203] |
| RR2 | WIRRIKKWIRRVHK (14aa) | AH | — | 3–6 | ||
| RR-4 | WLRRIKAWLRRIKA (14aa) | AH | — | 3–6 | ||
| R-Pro9-3 | Computationally designed | RRWIALWLR-NH2 (9aa) | AH | 16 | 8–32 | [201] |
| R-Pro9-3D | RRWIALWLR-NH2 (9aa) | AH | 8 | 4–16 | ||
| S4A | NA | IOWAGOLFOLFO-NH2 (12aa) | AH | 100 | 50 | [204] |
| SAAP-148 NPs | Synthetic peptide | LKRVWKRVFKLLKRYWRQLKKPVR (24aa) + NPs | AH | — | — | [205] |
| Scolopendin A2 | Synthetic peptide | AGLQFKVGRIGRLLRK (16aa) | NF | — | 16 | [206] |
| SPO | NA | NINONWNANGNONLNFNONLNFNO-NH2 (22aa) | AH | 100 | 50 | [204] |
| Stapled AMP Mag (i + 4)1, 15(A9 K, B21A, N22 K, S23 K) | NA, based on magainin two structure | Mag(i + 4)1,15(A9K,B21A,N22K,S23K) | complex | — | — | [207] |
| TAT-RasGAP317–326 anticancer peptide | Chimeric (cell penetrating sequence + Src homology sequence) | G48RKKRRQRRR57 + W317MWVTNLRTD326 | AH | Growth inhibitory effect | — | [208,209] |
| TAT-RasGAP317–326 | Chimeric peptide | G48RKKRRQRRR57 (10aa) | NF | — | — | [210] |
| TP2-5 | Computationally designed | KKCIAKAILKKAKKLLKKLVNP (22aa) | AH | 3.125 | 1.56–3.125 | [211] |
| TP2-6 | KKCIAKAILKKAKKLLKDLVNP (22aa) | AH | 3.125 | 3.125–12.5 | ||
| Trichogin analogs | Synthetic peptide | 1-Oct-Aib-Gly-Leu-Aib-Gly-Gly-Leu-Aib-Gly-Ile-Lol | >128 | — | [212] | |
| zp3 | Synthetic peptide | GIIAGIIIKIKK-NH2 (12aa) | AH | 4 | — | [213] |
| Mechanism | Gram-Negative Bacteria | Reference |
|---|---|---|
| Degradation or sequestration by secreted proteins | Proteolytic degradation | [314,315,316] |
| Impedance by exopolymers or biofilm matrix molecules | Alginate, polysialic acid | [304,317,318] |
| Cytoplasmic outer membrane alteration | Increased IM rigidity by PG acylation | [319] |
| Surface modification | Repulsion by lipid A phosphate modification increased OM rigidity by lipid A acylation. O-antigen of LPS | [320,321] |
| Multidrug efflux pump | Export via efflux pumps (RND family) | [322,323] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangel, K.; Lechuga, G.C.; Provance, D.W., Jr.; Morel, C.M.; De Simone, S.G. An Update on the Therapeutic Potential of Antimicrobial Peptides against Acinetobacter baumannii Infections. Pharmaceuticals 2023, 16, 1281. https://doi.org/10.3390/ph16091281
Rangel K, Lechuga GC, Provance DW Jr., Morel CM, De Simone SG. An Update on the Therapeutic Potential of Antimicrobial Peptides against Acinetobacter baumannii Infections. Pharmaceuticals. 2023; 16(9):1281. https://doi.org/10.3390/ph16091281
Chicago/Turabian StyleRangel, Karyne, Guilherme Curty Lechuga, David W. Provance, Jr., Carlos M. Morel, and Salvatore G. De Simone. 2023. "An Update on the Therapeutic Potential of Antimicrobial Peptides against Acinetobacter baumannii Infections" Pharmaceuticals 16, no. 9: 1281. https://doi.org/10.3390/ph16091281
APA StyleRangel, K., Lechuga, G. C., Provance, D. W., Jr., Morel, C. M., & De Simone, S. G. (2023). An Update on the Therapeutic Potential of Antimicrobial Peptides against Acinetobacter baumannii Infections. Pharmaceuticals, 16(9), 1281. https://doi.org/10.3390/ph16091281