


Proteomic Profiling Identifies Distinct Regulation of Proteins in Obese Diabetic Patients Treated with Metformin
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics and Demographics of the Study Population
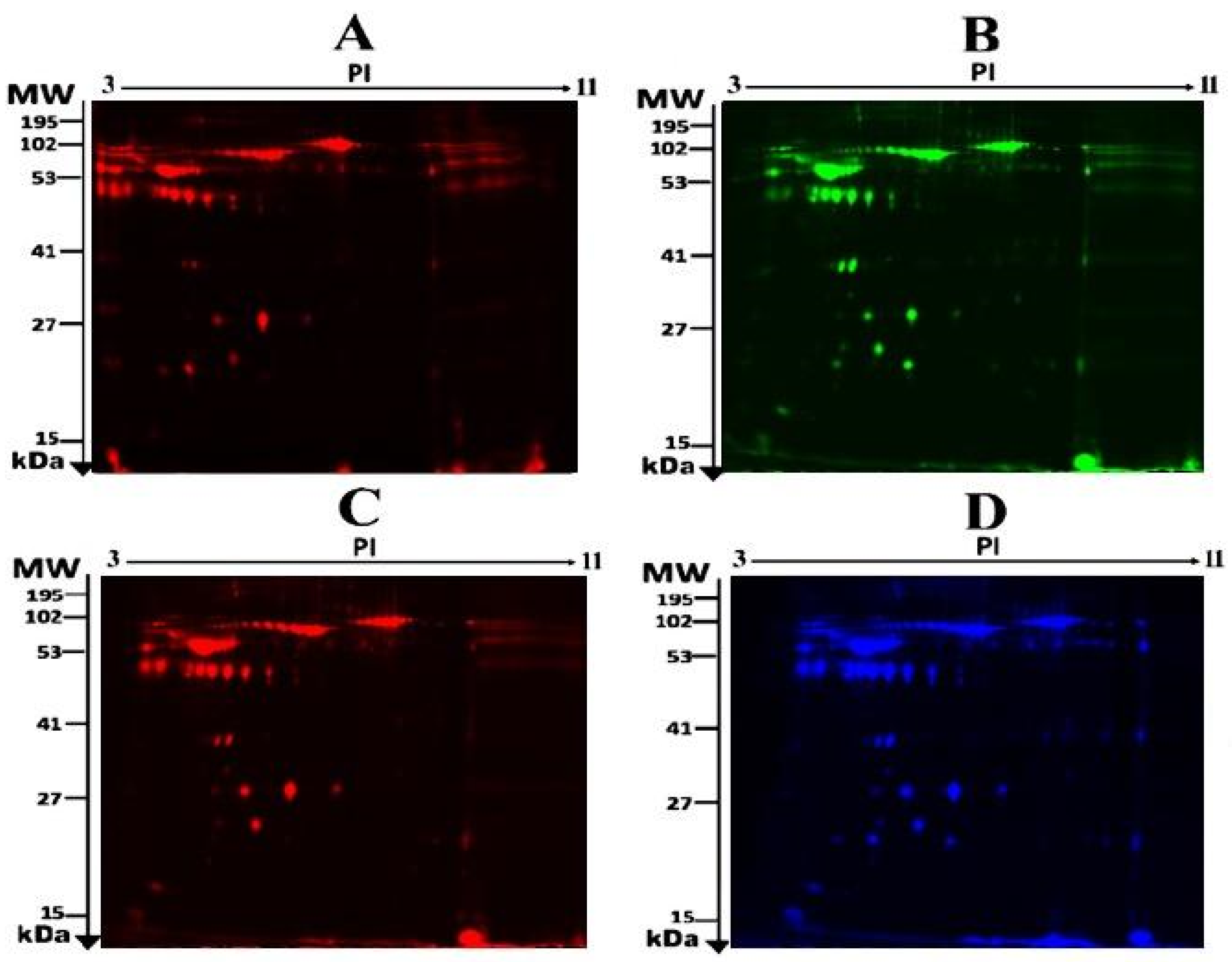
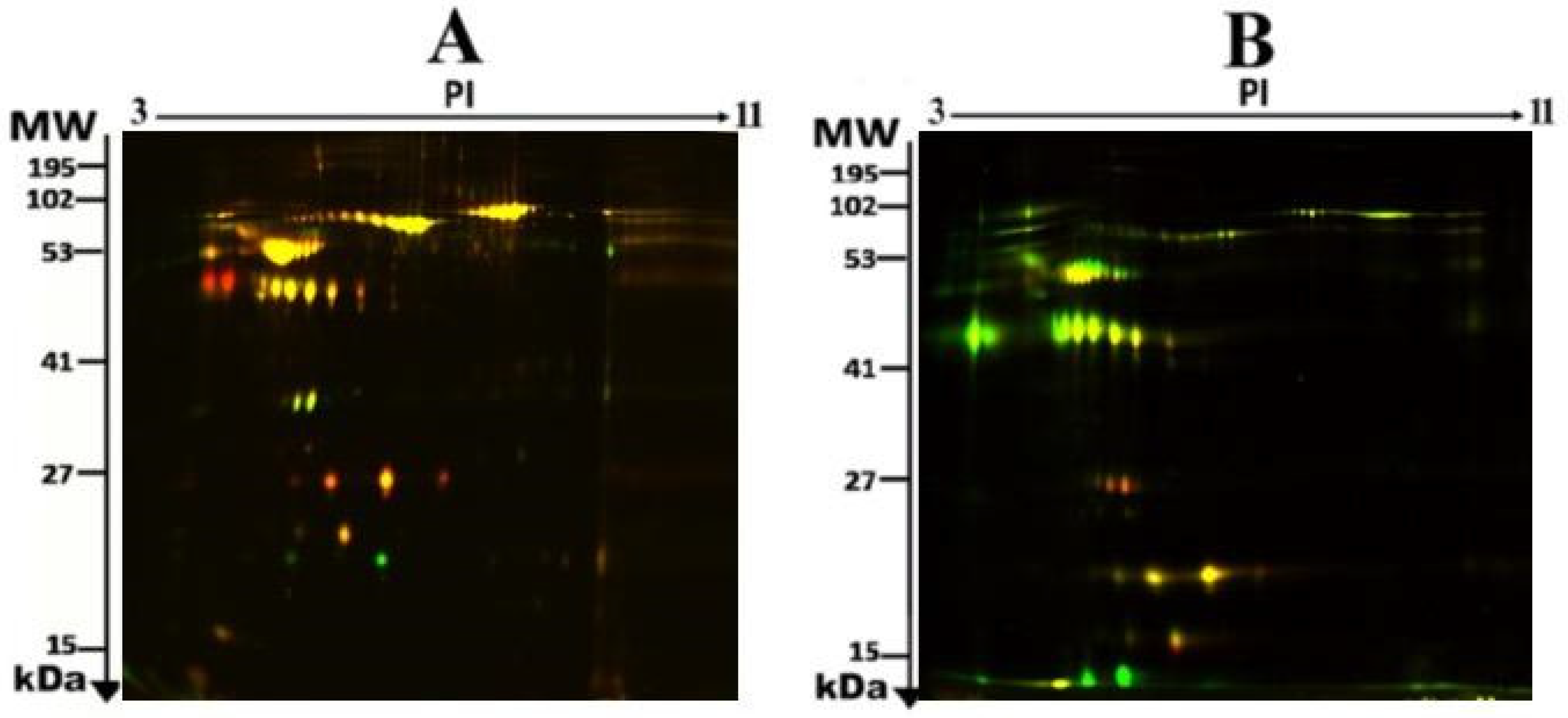
2.2. Proteomic Analysis and Identification of Differentially Expressed Proteins
2.3. Principal Component Analysis:
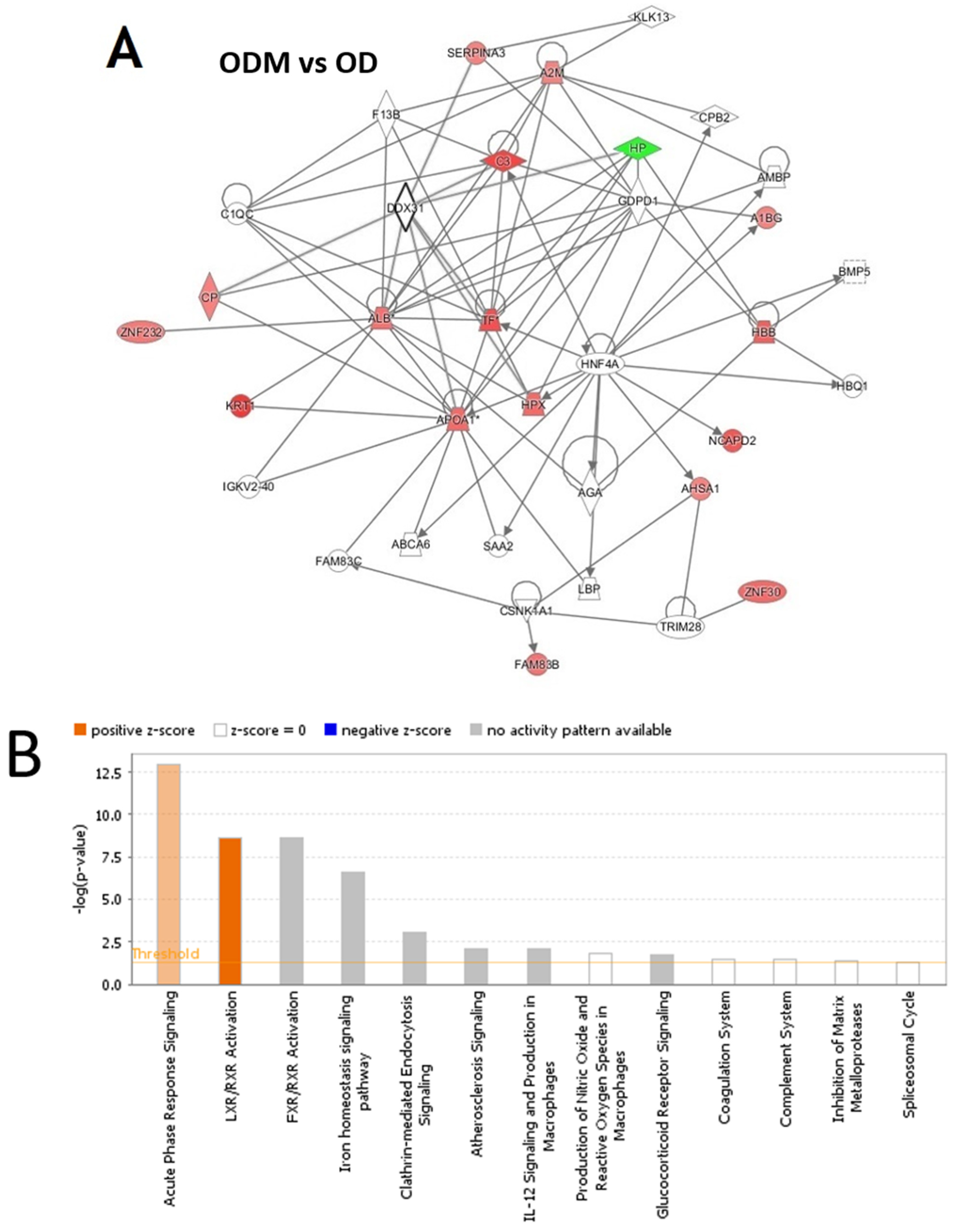
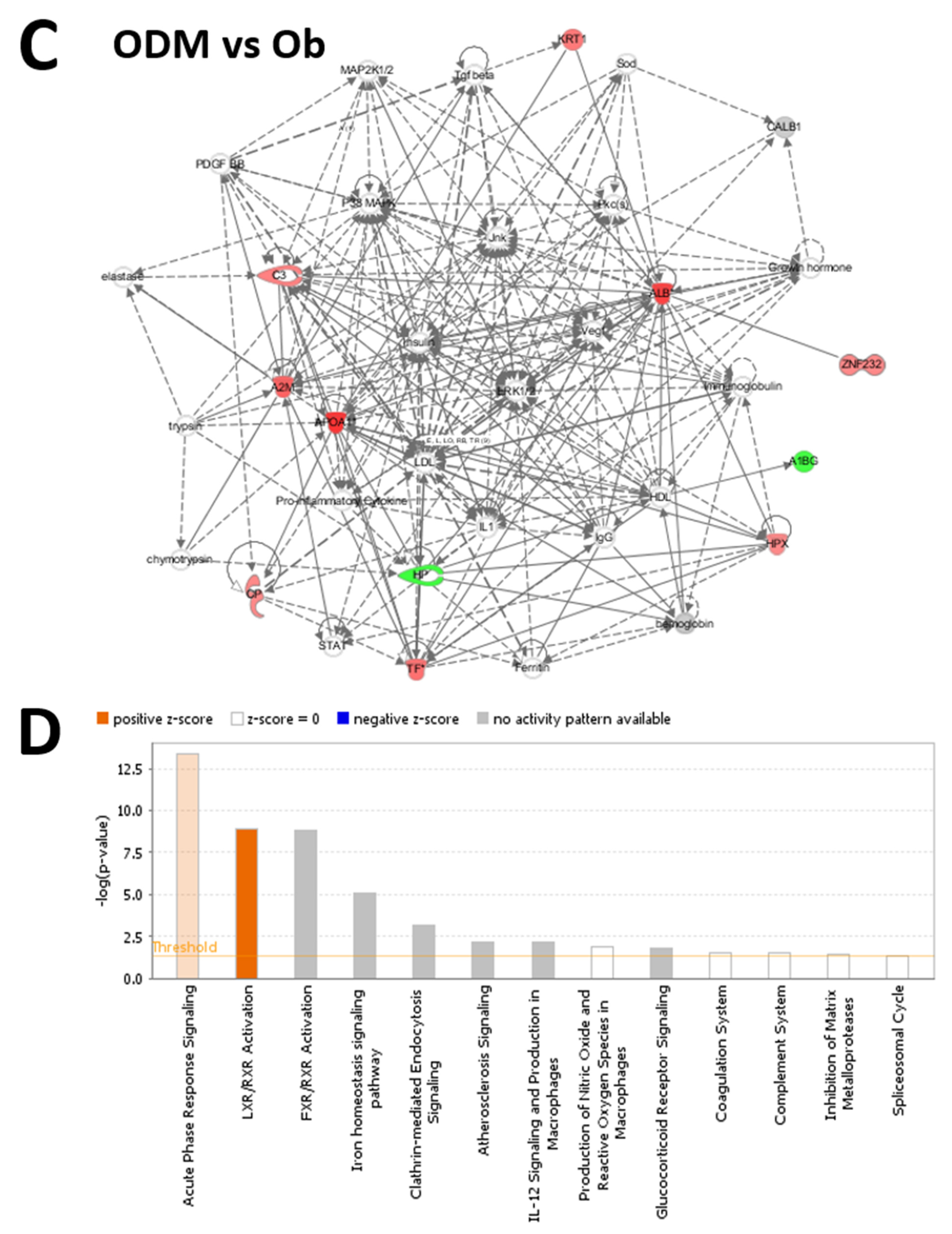
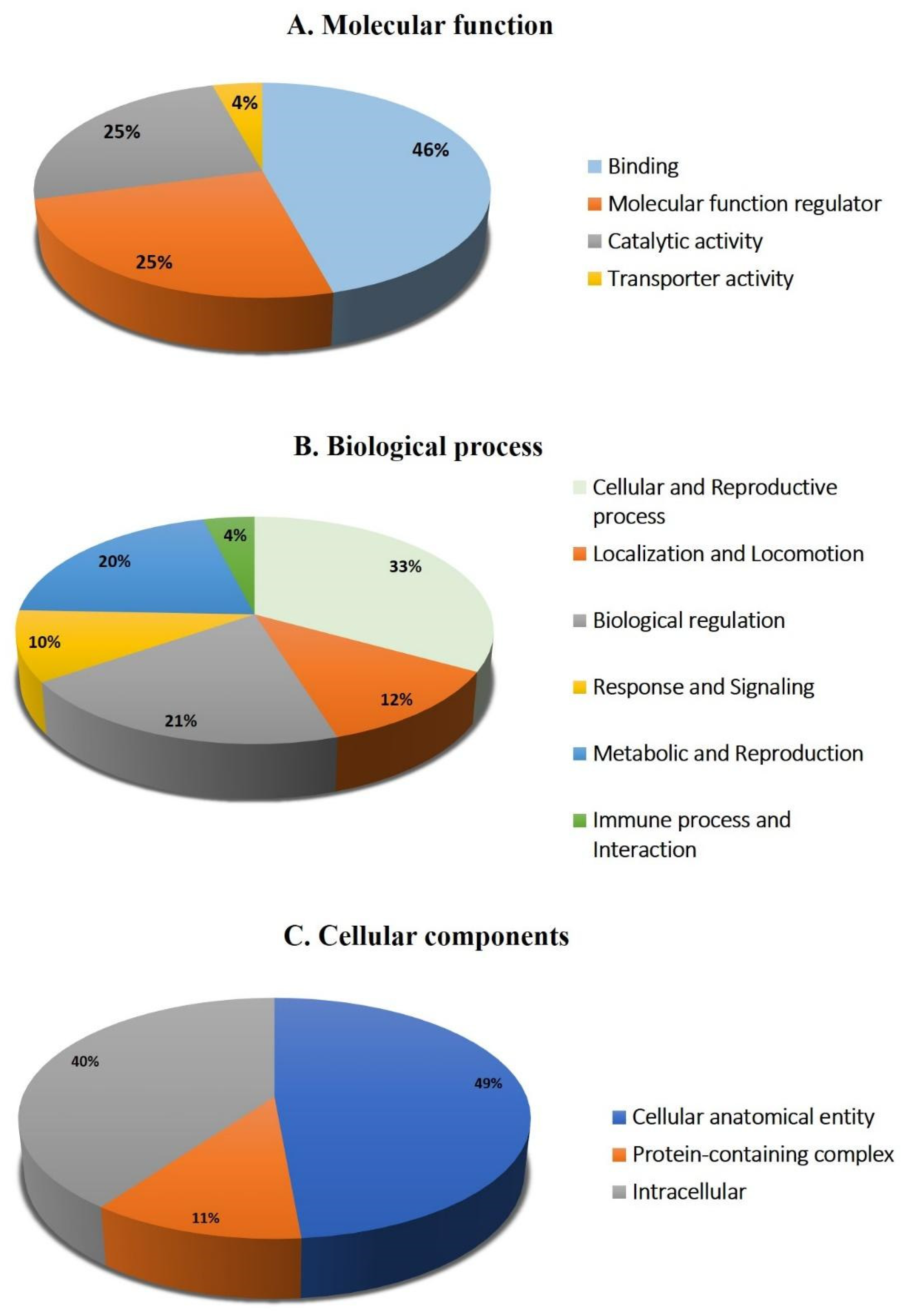
2.4. Network Pathway Analysis and Functional Classification of Proteins
3. Discussion
4. Materials and Methods
4.1. Ethical Considerations and Informed Consent
4.2. Study Subjects
4.3. Biochemical Analysis
4.4. Sample Preparation and Protein Extraction
4.5. Protein Labeling with Cyanine dyes
4.6. 2D-DIGE and Image Scanning
4.7. Colloidal Coomassie Blue Staining of the Preparative Gel
4.8. Protein Digestion and MALDI Analysis
4.9. Principal Component Analysis
4.10. Bioinformatics Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stanaway, J.D.; Afshin, A.; Gakidou, E.; Lim, S.S.; Abate, D.; Abate, K.H.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F. Global, regional, and national comparative risk assessment of 84 behavioural, environmental and occupational, and metabolic risks or clusters of risks for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1923–1994. [Google Scholar]
- Frühbeck, G.; Toplak, H.; Woodward, E.; Yumuk, V.; Maislos, M.; Oppert, J.-M. Obesity: The gateway to ill health-an EASO position statement on a rising public health, clinical and scientific challenge in Europe. Obes. Facts 2013, 6, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Duncan, B.B.; Schmidt, M.I.; Pankow, J.S.; Ballantyne, C.M.; Couper, D.; Vigo, A.; Hoogeveen, R.; Folsom, A.R.; Heiss, G. Low-grade systemic inflammation and the development of type 2 diabetes: The atherosclerosis risk in communities study. Diabetes 2003, 52, 1799–1805. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic adipose tissue inflammation linking obesity to insulin resistance and type 2 diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Golay, A. Metformin and body weight. Int. J. Obes. 2008, 32, 61–72. [Google Scholar] [CrossRef]
- Boden, G. 45Obesity, insulin resistance and free fatty acids. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 139. [Google Scholar] [CrossRef]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef]
- Benomar, Y.; Taouis, M. Molecular mechanisms underlying obesity-induced hypothalamic inflammation and insulin resistance: Pivotal role of resistin/TLR4 pathways. Front. Endocrinol. 2019, 10, 140. [Google Scholar] [CrossRef]
- Jiang, L.; Shi, K.; Guo, Y.-k.; Ren, Y.; Li, Z.-l.; Xia, C.-c.; Li, L.; Liu, X.; Xie, L.-j.; Gao, Y. The additive effects of obesity on myocardial microcirculation in diabetic individuals: A cardiac magnetic resonance first-pass perfusion study. Cardiovasc. Diabetol. 2020, 19, 52. [Google Scholar] [CrossRef]
- Group, U.P.D.S. Effect of intensive blood-glucose control with metformin on complications in overweight patients with type 2 diabetes (UKPDS 34). Lancet 1998, 352, 854–865. [Google Scholar]
- Hundal, R.S.; Krssak, M.; Dufour, S.; Laurent, D.; Lebon, V.; Chandramouli, V.; Inzucchi, S.E.; Schumann, W.C.; Petersen, K.F.; Landau, B.R. Mechanism by which metformin reduces glucose production in type 2 diabetes. Diabetes 2000, 49, 2063–2069. [Google Scholar] [CrossRef] [PubMed]
- Flory, J.; Lipska, K. Metformin in 2019. JAMA 2019, 321, 1926–1927. [Google Scholar] [CrossRef] [PubMed]
- Hundal, R.S.; Inzucchi, S.E. Metformin: New understandings, new uses. Drugs 2003, 63, 1879–1894. [Google Scholar] [CrossRef]
- Yerevanian, A.; Soukas, A.A. Metformin: Mechanisms in human obesity and weight loss. Curr. Obes. Rep. 2019, 8, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Razazan, A.; Nagpal, R.; Jain, S.; Wang, B.; Mishra, S.P.; Wang, S.; Justice, J.; Ding, J.; McClain, D.A. Metformin reduces aging-related leaky gut and improves cognitive function by beneficially modulating gut microbiome/goblet cell/mucin axis. J. Gerontol. Ser. A 2020, 75, e9–e21. [Google Scholar] [CrossRef]
- Chen, Z.-Z.; Gerszten, R.E. Metabolomics and proteomics in type 2 diabetes. Circ. Res. 2020, 126, 1613–1627. [Google Scholar] [CrossRef]
- Kim, S.W.; Choi, J.-W.; Yun, J.W.; Chung, I.-S.; Cho, H.C.; Song, S.-E.; Im, S.-S.; Song, D.-K. Proteomics approach to identify serum biomarkers associated with the progression of diabetes in Korean patients with abdominal obesity. PLoS ONE 2019, 14, e0222032. [Google Scholar] [CrossRef]
- Geyer, P.E.; Wewer Albrechtsen, N.J.; Tyanova, S.; Grassl, N.; Iepsen, E.W.; Lundgren, J.; Madsbad, S.; Holst, J.J.; Torekov, S.S.; Mann, M. Proteomics reveals the effects of sustained weight loss on the human plasma proteome. Mol. Syst. Biol. 2016, 12, 901. [Google Scholar] [CrossRef]
- Aleidi, S.M.; Dahabiyeh, L.A.; Gu, X.; Al Dubayee, M.; Alshahrani, A.; Benabdelkamel, H.; Mujammami, M.; Li, L.; Aljada, A.; Abdel Rahman, A.M. Obesity connected metabolic changes in type 2 diabetic patients treated with metformin. Front. Pharmacol. 2021, 11, 616157. [Google Scholar] [CrossRef]
- Morgantini, C.; Natali, A.; Boldrini, B.; Imaizumi, S.; Navab, M.; Fogelman, A.M.; Ferrannini, E.; Reddy, S.T. Anti-inflammatory and antioxidant properties of HDLs are impaired in type 2 diabetes. Diabetes 2011, 60, 2617–2623. [Google Scholar] [CrossRef]
- Navab, M.; Hama, S.Y.; Anantharamaiah, G.; Hassan, K.; Hough, G.P.; Watson, A.D.; Reddy, S.T.; Sevanian, A.; Fonarow, G.C.; Fogelman, A.M. Normal high density lipoprotein inhibits three steps in the formation of mildly oxidized low density lipoprotein: Steps 2 and 3. J. Lipid Res. 2000, 41, 1495–1508. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Real, J.M.; López-Bermejo, A.; Ricart, W. Cross-talk between iron metabolism and diabetes. Diabetes 2002, 51, 2348–2354. [Google Scholar] [CrossRef] [PubMed]
- González-Domínguez, Á.; Visiedo-García, F.M.; Domínguez-Riscart, J.; González-Domínguez, R.; Mateos, R.M.; Lechuga-Sancho, A.M. Iron metabolism in obesity and metabolic syndrome. Int. J. Mol. Sci. 2020, 21, 5529. [Google Scholar] [CrossRef] [PubMed]
- Memişoǧulları, R.; Bakan, E. Levels of ceruloplasmin, transferrin, and lipid peroxidation in the serum of patients with Type 2 diabetes mellitus. J. Diabetes Its Complicat. 2004, 18, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Van Campenhout, A.; Van Campenhout, C.; Lagrou, A.R.; Moorkens, G.; De Block, C.; Manuel-y-Keenoy, B. Iron-binding antioxidant capacity is impaired in diabetes mellitus. Free. Radic. Biol. Med. 2006, 40, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, Y.; Yang, Y.; Xu, Z.; Tong, J.; Li, Z.; Zhou, X.; Li, C. Transcriptomic and proteomic analysis of potential therapeutic target genes in the liver of metformin-treated Sprague-Dawley rats with type 2 diabetes mellitus. Int. J. Mol. Med. 2018, 41, 3327–3341. [Google Scholar] [CrossRef]
- Kawasaki, N.; Asada, R.; Saito, A.; Kanemoto, S.; Imaizumi, K. Obesity-induced endoplasmic reticulum stress causes chronic inflammation in adipose tissue. Sci. Rep. 2012, 2, 799. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Xiao, H. Metformin actions on the liver: Protection mechanisms emerging in hepatocytes and immune cells against NASH-related HCC. Int. J. Mol. Sci. 2021, 22, 5016. [Google Scholar] [CrossRef]
- Ren, H.; Shao, Y.; Wu, C.; Ma, X.; Lv, C.; Wang, Q. Metformin alleviates oxidative stress and enhances autophagy in diabetic kidney disease via AMPK/SIRT1-FoxO1 pathway. Mol. Cell. Endocrinol. 2020, 500, 110628. [Google Scholar] [CrossRef]
- MacKellar, M.; Vigerust, D.J. Role of haptoglobin in health and disease: A focus on diabetes. Clin. Diabetes 2016, 34, 148–157. [Google Scholar] [CrossRef]
- Jing, Y.; Wu, F.; Li, D.; Yang, L.; Li, Q.; Li, R. Metformin improves obesity-associated inflammation by altering macrophages polarization. Mol. Cell. Endocrinol. 2018, 461, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.; Romero, M.; Vazquez, T.; Lechner, S.; Blomberg, B.B.; Frasca, D. Metformin improves in vivo and in vitro B cell function in individuals with obesity and Type-2 Diabetes. Vaccine 2017, 35, 2694–2700. [Google Scholar] [CrossRef] [PubMed]
- Gabuza, K.B.; Sibuyi, N.R.S.; Mobo, M.P.; Madiehe, A.M. Differentially expressed serum proteins from obese Wistar rats as a risk factor for obesity-induced diseases. Sci. Rep. 2020, 10, 12415. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ambrosi, J.; Catalán, V.; Diez-Caballero, A.; Martínez-Cruz, L.A.; Gil, M.J.; García-Foncillas, J.; Cienfuegos, J.A.; Salvador, J.; Mato, J.M.; Frühbeck, G. Gene expression profile of omental adipose tissue in human obesity. FASEB J. 2004, 18, 215–217. [Google Scholar] [CrossRef]
- Sooy, K.; Schermerhorn, T.; Noda, M.; Surana, M.; Rhoten, W.B.; Meyer, M.; Fleischer, N.; Sharp, G.W.; Christakos, S. Calbindin-D28k Controls [Ca2+] iand Insulin Release: EVIDENCE OBTAINED FROM CALBINDIN-D28k KNOCKOUT MICE AND β CELL LINES. J. Biol. Chem. 1999, 274, 34343–34349. [Google Scholar] [CrossRef]
- Yang, L.; Lu, P.; Yang, X.; Li, K.; Qu, S. Annexin A3, a calcium-dependent phospholipid-binding protein: Implication in cancer. Front. Mol. Biosci. 2021, 8, 716415. [Google Scholar] [CrossRef]
- Purvis, G.S.; Collino, M.; Loiola, R.A.; Baragetti, A.; Chiazza, F.; Brovelli, M.; Sheikh, M.H.; Collotta, D.; Cento, A.; Mastrocola, R. Identification of AnnexinA1 as an endogenous regulator of RhoA, and its role in the pathophysiology and experimental therapy of type-2 diabetes. Front. Immunol. 2019, 10, 571. [Google Scholar] [CrossRef]
- Ishii, H.; Yoshida, M.; Hajjar, K.A.; Tanaka, A.; Yasukochi, Y.; Shimokado, K.; Numano, F. Potential role of recombinant annexin II in diabetic vascular injury. Ann. N. Y. Acad. Sci. 2001, 947, 308–311. [Google Scholar] [CrossRef]
- Pietrani, N.T.; Ferreira, C.N.; Rodrigues, K.F.; Perucci, L.O.; Carneiro, F.S.; Bosco, A.A.; Oliveira, M.C.; Pereira, S.S.; Teixeira, A.L.; Alvarez-Leite, J.I. Proresolving protein Annexin A1: The role in type 2 diabetes mellitus and obesity. Biomed. Pharmacother. 2018, 103, 482–489. [Google Scholar] [CrossRef]
- Meadows, S.M.; Cleaver, O. Annexin A3 regulates early blood vessel formation. PLoS ONE 2015, 10, e0132580. [Google Scholar] [CrossRef]
- Watanabe, T.; Ito, Y.; Sato, A.; Hosono, T.; Niimi, S.; Ariga, T.; Seki, T. Annexin A3 as a negative regulator of adipocyte differentiation. J. Biochem. 2012, 152, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Burdin, D.V.; Kolobov, A.A.; Brocker, C.; Soshnev, A.A.; Samusik, N.; Demyanov, A.V.; Brilloff, S.; Jarzebska, N.; Martens-Lobenhoffer, J.; Mieth, M. Diabetes-linked transcription factor HNF4α regulates metabolism of endogenous methylarginines and β-aminoisobutyric acid by controlling expression of alanine-glyoxylate aminotransferase 2. Sci. Rep. 2016, 6, 35503. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.; Ræder, H.; Eide, S.A.; Midthjell, K.; Hveem, K.; Søvik, O.; Molven, A.; Njølstad, P.l.R. Studies in 3523 Norwegians and meta-analysis in 11,571 subjects indicate that variants in the hepatocyte nuclear factor 4α (HNF4A) P2 region are associated with type 2 diabetes in scandinavians. Diabetes 2007, 56, 3112–3117. [Google Scholar] [CrossRef]
- Abu-Farha, M.; Tiss, A.; Abubaker, J.; Khadir, A.; Al-Ghimlas, F.; Al-Khairi, I.; Baturcam, E.; Cherian, P.; Elkum, N.; Hammad, M. Proteomics analysis of human obesity reveals the epigenetic factor HDAC4 as a potential target for obesity. PLoS ONE 2013, 8, e75342. [Google Scholar] [CrossRef] [PubMed]
- Fliser, E.; Jerkovič, K.; Vidovič, T.; Gorenjak, M. Investigation of unusual high serum indices for lipemia in clear serum samples on Siemens analysers Dimension. Biochem. Medica 2012, 22, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Lin, S.Y.; Yeh, Y.Y.; Hsiao, H.H.; Wu, C.Y.; Chen, S.T.; Wang, A.H.J. A modified protein precipitation procedure for efficient removal of albumin from serum. Electrophoresis 2005, 26, 2117–2127. [Google Scholar] [CrossRef]
- Masood, A.; Benabdelkamel, H.; Ekhzaimy, A.A.; Alfadda, A.A. Plasma-based proteomics profiling of patients with hyperthyroidism after antithyroid treatment. Molecules 2020, 25, 2831. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Moustafa, A.; Sallam, R.; Bassas, A.; Duncan, M. Proteomic analysis of mature adipo cytes from obese patients in relation to aging. Exp. Gerontol. 2013, 48, 1196–1203. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ODM (n = 7) | OD (n = 8) | OB (n = 10) | |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| Age (y) | 48.71 ± 11.34 | 50 ± 9.78 | 40.4 ± 6.00 ‡ |
| Gender (F/M) | 6/1 | 1/7 | 9/1 |
| BMI (kg/m2) | 47.31 ± 6.88 | 30.74 ± 1.12 * | 43.27 ± 6.78 ‡ |
| Glucose (mM) | 11.04 ± 3.57 | 12.2 ± 4.73 | 5.3 ± 0.43 *,‡ |
| HbA1c (%) | 9.19 ± 1.59 | 9.45 ± 3.09 | 5.75 ± 0.25 *,‡ |
| Total Chol mmol/L | 4.21 ± 0.92 † | 5.78 ± 0.75 | 4.932 ± 0.70 |
| LDL (mmol/L) | 4.21 ± 0.92 ‡ | 5.78 ± 0.75 * | 4.932 ± 0.70 |
| HDL (mmol/L) | 2.40 ± 0.67 | 3.7 ± 0.62 | 3.18 ± 0.76 † |
| Trig. (mM) | 1.06 ± 0.27 | 0.96 ± 0.18 | 1.187 ± 0.15 † |
| S. No. | Protein Name | MASCOT ID a | p-Value b (ANOVA) | Ratio ODM/Ob | Exp d | Ratio ODM/OD c | Exp d | Ratio OD/Ob | Exp d |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Calbindin | CALB1_HUMAN | 0.01 | 1 | NS | 2 | UP | −1.89 | DOWN |
| 2 | Apolipoprotein A–I | APOA1_HUMAN | 0.01 | 2.6 | UP | 1.5 | UP | - | NS |
| 3 | Albumin | ALBU_HUMAN | 0.01 | 1.7 | UP | 1.5 | UP | 1.1 | NS |
| 4 | Haptoglobin | HPT_HUMAN | 0.01 | −1.55 | DOWN | −1.6 | DOWN | 1 | NS |
| 5 | ANXA3_HUMAN | ANXA3_HUMAN | 0.01 | 1.8 | UP | 1.1 | NS | 1.6 | UP |
| 6 | Alpha-2-macroglobulin | A2MG_HUMAN | 0.01 | 2.0 | UP | 1.5 | UP | 1.5 | UP |
| 7 | Albumin | ALBU_HUMAN | 0.02 | 2.4 | UP | 1.7 | UP | 1.4 | NS |
| 8 | Hemopexin | HEMO_HUMAN | 0.02 | 1.5 | UP | 1.8 | UP | - | NS |
| 9 | Condensin complex subunit 1 | CND1_HUMAN | 0.02 | 1.7 | UP | 2 | UP | - | NS |
| 10 | Pre-mRNA-splicing factor ISY1 homolog | ISY1_HUMAN | 0.02 | 1.1 | NS | 1.7 | UP | −1.5 | DOWN |
| 11 | Hemoglobin subunit beta | HBB_HUMAN | 0.02 | - | NS | 1.9 | UP | −2.0 | DOWN |
| 12 | Albumin | ALBU_HUMAN | 0.02 | 1.6 | UP | 1.7 | UP | - | NS |
| 13 | Complement C3 | CO3_HUMAN | 0.02 | 1.6 | UP | 2.2 | UP | - | NS |
| 14 | Albumin | ALBU_HUMAN | 0.03 | 1.5 | UP | 1.7 | UP | - | NS |
| 15 | Serotransferrin | TRFE_HUMAN | 0.03 | 1.5 | UP | 2 | UP | −1.6 | DOWN |
| 16 | Zinc finger protein 232 | ZN232_HUMAN | 0.03 | 1.5 | UP | 1.6 | UP | - | NS |
| 17 | Serotransferrin | TRFE_HUMAN | 0.04 | 1.5 | UP | 2.1 | UP | −1.5 | DOWN |
| 18 | Keratin, type II cytoskeletal 1 | K2C1_HUMAN | 0.04 | 1.7 | UP | 2.4 | UP | −1.5 | DOWN |
| 19 | Activator of 90 kDa heat shock protein ATPase homolog 1 | AHSA1_HUMAN | 0.04 | - | NS | 1.5 | UP | −1.6 | DOWN |
| 20 | Zinc finger protein 30 | ZNF30_HUAMN | 0.05 | 1.0 | NS | 1.8 | UP | −1.7 | DOWN |
| 21 | Serotransferrin | TRFE_HUMAN | 0.05 | 1.8 | UP | 1.7 | UP | 1.1 | NS |
| 22 | Alpha-1B-glycoprotein | A1BG_HUMAN | 0.05 | −1.5 | DOWN | 1.5 | UP | −1.6 | DOWN |
| 23 | PC4 and SFRS1-interacting protein | PSIP1_HUMAN | 0.06 | −1.5 | DOWN | −1.5 | DOWN | −1.5 | DOWN |
| 24 | Clathrin heavy-chain linker domain-containing protein 1 | CB063_HUMAN | 0.05 | 1.2 | NS | 1.7 | UP | −1.5 | DOWN |
| 25 | Zinc finger protein 69 | ZNF69_HUMAN | 0.05 | 1.5 | UP | 1.6 | UP | - | NS |
| 26 | Protein FAM83B | FA83B_HUMAN | 0.05 | 1.6 | UP | 1.7 | UP | 1 | NS |
| 27 | Ceruloplasmin | CERU_HUMAN | 0.05 | 1.5 | UP | 1.5 | UP | 1 | NS |
| 28 | Alpha-1-antichymotrypsin | AACT_HUMAN | 0.05 | −1.8 | DOWN | 1.5 | UP | −2.1 | DOWN |
| 29 | Apolipoprotein A-I | APOA1_HUMAN | 0.05 | 1.8 | UP | 1.8 | UP | - | NS |
| 30 | Coronin-1A | COR1A_HUMAN | 0.05 | 1.6 | UP | 1.5 | UP | 1 | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alshahrani, A.; Aljada, A.; Masood, A.; Mujammami, M.; Alfadda, A.A.; Musambil, M.; Alanazi, I.O.; Al Dubayee, M.; Abdel Rahman, A.M.; Benabdelkamel, H. Proteomic Profiling Identifies Distinct Regulation of Proteins in Obese Diabetic Patients Treated with Metformin. Pharmaceuticals 2023, 16, 1345. https://doi.org/10.3390/ph16101345
Alshahrani A, Aljada A, Masood A, Mujammami M, Alfadda AA, Musambil M, Alanazi IO, Al Dubayee M, Abdel Rahman AM, Benabdelkamel H. Proteomic Profiling Identifies Distinct Regulation of Proteins in Obese Diabetic Patients Treated with Metformin. Pharmaceuticals. 2023; 16(10):1345. https://doi.org/10.3390/ph16101345
Chicago/Turabian StyleAlshahrani, Awad, Ahmad Aljada, Afshan Masood, Muhammad Mujammami, Assim A. Alfadda, Mohthash Musambil, Ibrahim O. Alanazi, Mohammed Al Dubayee, Anas M. Abdel Rahman, and Hicham Benabdelkamel. 2023. "Proteomic Profiling Identifies Distinct Regulation of Proteins in Obese Diabetic Patients Treated with Metformin" Pharmaceuticals 16, no. 10: 1345. https://doi.org/10.3390/ph16101345
APA StyleAlshahrani, A., Aljada, A., Masood, A., Mujammami, M., Alfadda, A. A., Musambil, M., Alanazi, I. O., Al Dubayee, M., Abdel Rahman, A. M., & Benabdelkamel, H. (2023). Proteomic Profiling Identifies Distinct Regulation of Proteins in Obese Diabetic Patients Treated with Metformin. Pharmaceuticals, 16(10), 1345. https://doi.org/10.3390/ph16101345