1. Introduction
Acinetobacter baumannii is an opportunistic pathogen, associated with most hospital-derived Acinetobacter infections [
1].
A. baumannii infection is frequent in immunocompromised patients with skin and soft tissue diseases, surgical sites, bloodstream infections, and urinary tract infections [
2]. Aerobic, non-fermentative, Gram-negative, non-motile bacillus,
A. baumannii, harbors multiple virulence factors to survive in a wide range of adverse conditions and persists on surfaces for a long time in an inactive form. The possession of an intrinsic efflux pump, genetic adaptation, biofilm formation, and the presence of several beta-lactamase inhibitors are the main defense mechanisms of
A. baumannii for the development of antibiotic resistance [
3]. In recent studies,
A. baumannii was found to be resistant to multiple antibiotics, including carbapenem, which is the last line of drugs for controlling it [
4]. At present, combination antibiotic therapy is the final solution used in the medical fraternity for controlling multidrug resistant (MDR)
A. baumannii, which makes treatment procedures more complex and has more incidences of side effects [
5]. The development of alternatives to antibiotics is the only solution for overcoming the challenges of MDR
A. baumannii [
6].
Tackling antibiotic resistance has become a major challenge in the healthcare system, and thus far, multiple compounds, such as probiotics, prebiotics, organic acids, antimicrobial peptides (AMPs), nanomaterials, and bacteriophages, have been tested as possible candidates [
7,
8,
9]. Most of AMPs are natural antimicrobial compounds synthesized by organisms from archaea to mammals as a group of compounds involved in innate immunity. AMPs play a key role in defending the host against pathogenic bacteria, fungi, protozoa, and viruses [
10]. AMPs are diverse within and across species, sharing common features consisting of 10 to 50 amino acid (AA) residues, water solubility, and high hydrophobicity with a positive charge (majority of AMPs). Depending on their secondary structure, AMPs can be divided into α-helices, β-sheets, and extenders [
11]. AMPs possess multiple modes of action, from initial membrane disruptions to DNA and RNA damage, induction of reactive oxygen species (ROS) generation, and denaturation of proteins and enzymes [
12]. Since AMPs acquire multiple modes of action and are not constrained to a single pathogen species, the possibility of the development of AMP resistance by the pathogen is at the lowest possible level [
13]. Moreover, the majority of newly discovered and synthesized AMPs have shown extremely low or no toxicity towards humans and other tested animals [
14]. Considering the characteristics and functional features of AMPs with low toxicity, AMPs have potential for the next generation of antimicrobials.
Marine invertebrates are deficient in adaptive immunity and primarily depend on innate immunity, which consists of physical and chemical compounds, including AMPs [
15].
Octopus minor is considered a rich source of novel AMPs, and we synthesized octominin and octopromycin in previous studies [
16,
17]. Octominin, which is based on defense protein-3, showed potent bactericidal and fungicidal activities against
A. baumannii and
Candida albicans, respectively, whereas octopromycin, which is based on proline-rich 5 protein, showed high antibacterial activity against
A. baumannii [
16,
17,
18]. Prohibitin is a highly conserved class of proteins that inhibits cellular proliferation and has other roles in nuclear signaling, mitochondrial integrity, cell division, and cellular membrane metabolism [
19]. We found the prohibitin-2 gene in the
O. minor transcriptome, which was selected for the development of octoprohibitin as a novel AMP. Initially, we tested the antibacterial activity of octoprohibitin against
A. baumannii and its detailed modes of action, including morphological and structural changes of the surface, membrane permeability alterations, ROS, and DNA binding ability. Furthermore, we tested octoprohibitin as an antibiofilm agent by inhibition of biofilm formation and biofilm eradication assays. Ultimately, an
A. baumannii-infected zebrafish model was used to determine in vivo efficacy of Octoprohibitin for controlling the infection.
3. Discussion
AMPs may be a one of major solutions for the control of MDR pathogens such as
A. baumannii, since they contain multiple modes of action, are effective against a wide range of pathogenic species, are able to improve host immunity, and have low toxicity [
23]. Marine invertebrates are rich in several AMPs because they rely mainly on innate immunity. In several previous studies, we studied
O. minor for its AMPs and synthesized octominin and octopromycin, which showed potent antimicrobial activities against both fungi and bacteria [
16,
17]. In the present study, the AA sequence of the prohibitin-2 cDNA sequence was selected as a template for designing and synthesizing a novel AMP named octoprohibitin. The
A. baumannii strain used in this study was screened for antibiotic sensitivity and was found to be resistant to multiple antibiotics in our previous study [
24]. Initially, octoprohibitin showed slight antimicrobial activity against
A. baumannii at a pH of 7.4. Some AMPs exhibit pH-dependent antimicrobial activity. Esculentin-2EM, an alpha-helical peptide isolated from
Glandirana emeljanovi, showed a higher rate of antimicrobial activity against Gram-positive and Gram-negative bacteria at pH 8 than that at pH 6 [
24]. We examined the optimum pH for the highest antibacterial activity of octoprohibitin and found more efficient bactericidal activity at pH 8 and 9. Since the optimum pH range for
A. baumannii growth is 5–8, we selected pH 8 for
A. baumannii culturing for further experiments with octoprohibitin treatments [
25]. Potent antimicrobial activity of octoprohibitin was detected at MIC and MBC values of 200 and 400 µg/mL, respectively. The bactericidal or bacteriostatic activity of the antimicrobial agent is determined by the MBC/MIC ratio, and a compound is considered bactericidal if the ratio is below 4.0, and bacteriostatic if the ratio is higher than 4.0 [
26]. Octoprohibitin is considered bactericidal rather than bacteriostatic because of its MBC/MIC ratio of 2.0. Moreover, a time-kill kinetic assay and bacterial viability test verified concentration-dependent antibacterial activity, similar to our previous finding with octominin and octopromycin against
A. baumannii [
17,
18].
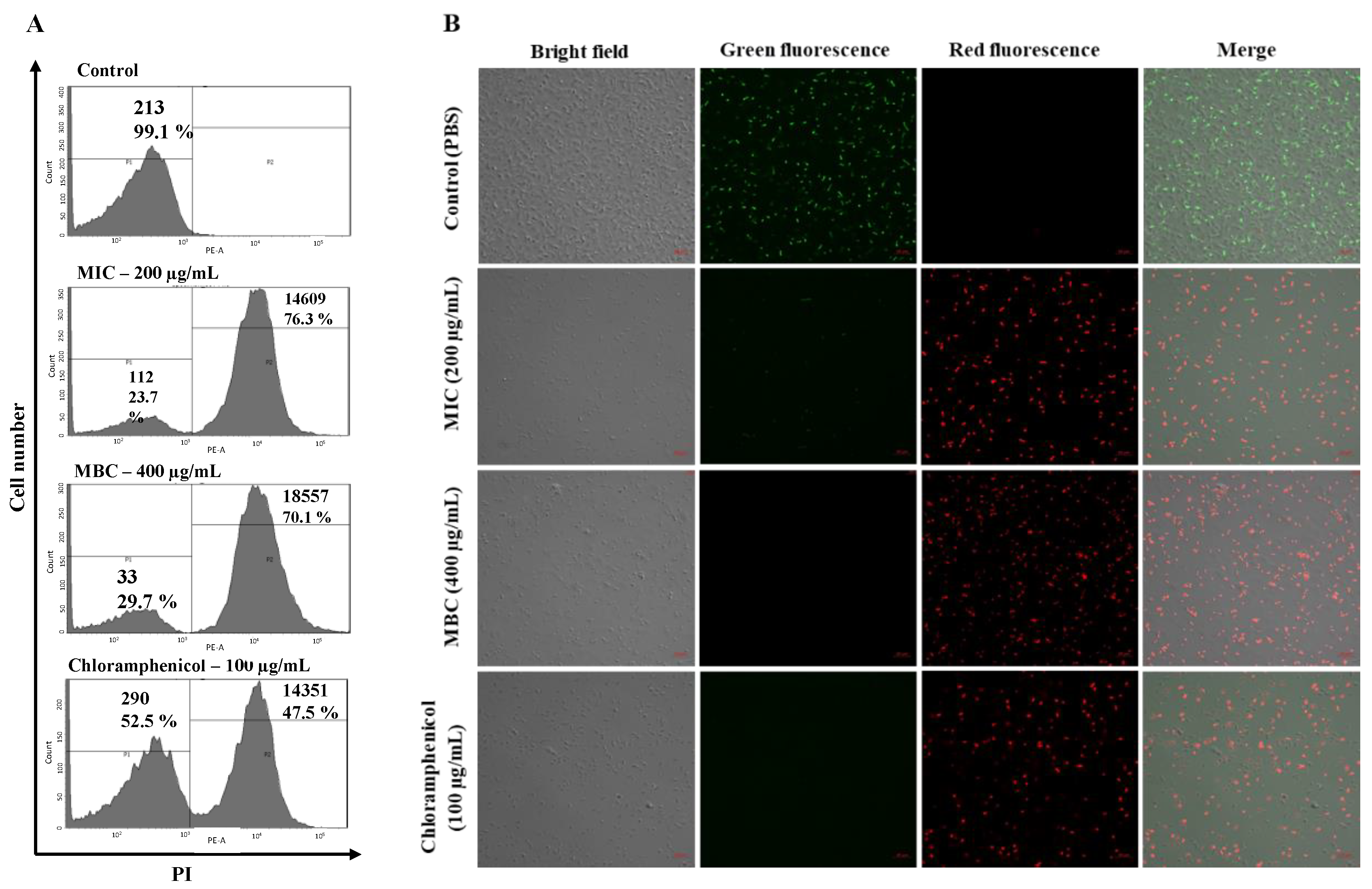
Cationic AMPs mainly act by disrupting the bacterial membrane. The positive charge of AMPs facilitates efficient binding to the negatively charged bacterial membrane and completes the activity through different models, such as forming pores and ion channels to induce overflow of bacterial content (barrel stave model) or disintegrating the cell membrane by changing the surface tension (carpet model) [
27]. Since octoprohibitin possesses a +10 charge and amphiphilic nature, the main predicted mode of action against
A. baumannii is morphological and structural damage to the bacterial surface and membrane permeability alterations. Micrographs of FE-SEM analysis confirmed that the surface morphology of the bacteria was altered by the formation of pores on the membrane and significant damage to the surface by octoprohibitin treatment. In addition, Mwangi et al. suggested that cathelicidin-based AMP ZY4 induces permeabilization of the
A. baumannii membrane with a similar pattern of damage by examining the cell surface using FE-SEM [
28]. According to the toroidal-pore model, cationic AMPs aggregate inside the cell membrane and alter the phospholipid monolayer of bacteria to produce nanoscale pores, which facilitates permeability alterations [
29]. A PI uptake assay of octoprohibitin-treated
A. baumannii confirmed a significant amount of membrane permeability alterations similar to the activity of octominin to induce membrane perturbation in
A. baumannii [
18]. Morphological alterations with pore formation and membrane permeability changes together might result in the leakage of the intracellular content of
A. baumannii to the outside and cause cellular apoptosis or necrosis, ultimately causing bacterial death.
Several studies have shown that AMPs can penetrate the bacterial cell cytoplasm and induce further bactericidal activities by interrupting biochemical processes inside the cell [
30]. Since octoprohibitin induces alterations in membrane surface morphology and permeability, we hypothesized that octoprohibitin could translocate into the cytoplasm and induce further antimicrobial actions. A study on LL-37, a cationic AMP, induced superoxide and peroxide production in
Escherichia coli under aerobic conditions [
31]. Increased ROS inside bacterial cells induces damage to proteins, lipids, and nucleotides, and this negative impact on bacteria is tolerable up to a certain level due to the activation of antioxidative stress response genes. Once an antimicrobial agent is capable of producing lethal stress with a significant amount of ROS, which is not borne by defense mechanisms, bacterial death is self-driven [
32]. The latest findings suggest that generated ROS can cause eventual cell death by surging the self-amplification of ROS, even after the removal of the antimicrobial agent [
33]. Flow cytometry and confocal microscopy analysis confirmed that octoprohibitin could increase the level of ROS inside
A. baumannii, which could be a potential antimicrobial action that has not been well discussed previously with other AMPs in detail. However, further experiments are essential to determine the exact mechanism underlying ROS formation by octoprohibitin in
A. baumannii.
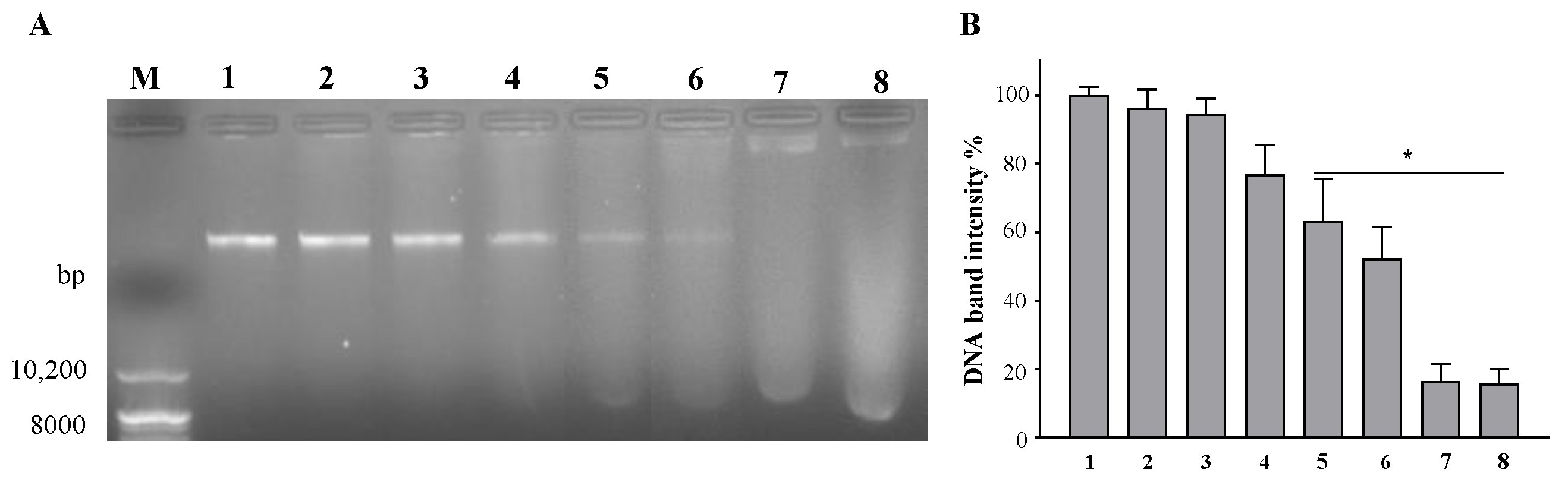
Since we expected octoprohibitin translocation into
A. baumannii, we predicted its cationic charge to facilitate possible binding to negatively charged genomic DNA for further expansion of the antimicrobial activity. Several AMPs depend on their antimicrobial activity by binding to bacterial DNA. BF2-A and BF2-C, which are buforin II-based AMPs isolated from Asian toads, can inhibit nucleic acid metabolism [
34]. The ostrich-derived β-defensins ostricacin-1 (Osp-1) and ostricacin-2 (Osp-2) show antimicrobial activity by inhibiting protein synthesis and enzymatic activity as a result of their binding affinity to DNA [
35]. Microcin-based B17 AMP can bind to DNA and block DNA replication by inhibiting the gyrase-DNA complex [
36]. The gel retardation assay is the initial step in investigating the DNA-binding ability of an AMP. Since octoprohibitin showed concentration-dependent DNA binding and possible genomic DNA degradation in
A. baumannii, it may contain further DNA-based antimicrobial activities. Further investigations are essential to determine the exact mechanism by which octoprohibitin binds to and disrupts the genomic DNA of
A. baumannii.A. baumannii has a remarkable capacity to form biofilms on a wide range of surfaces, from abiotic stainless steel to host epithelial cells, and it has been identified as one of the major factors for antibiotic resistance [
37]. Antibiofilm AMPs can act adversely at different stages of biofilm formation through various mechanisms, including downregulation of quorum sensing, killing of early-stage biofilm, and inhibition of adhesion [
38]. Octoprohibitin might also have generated similar actions to inhibit the formation of
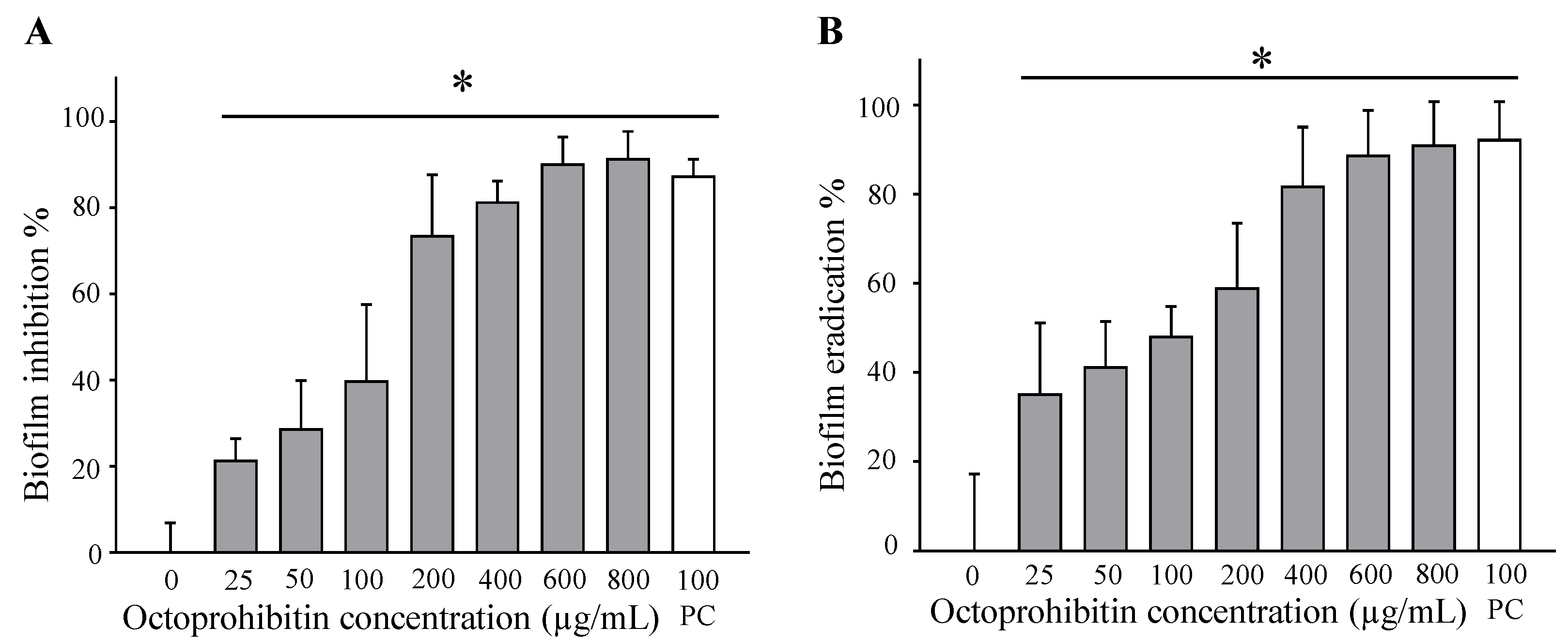
A. baumannii biofilms with an MBIC of 1000 µg/mL. Nevertheless, eradication of pre-formed biofilms by antimicrobials is considered challenging because biofilms consist of various defense mechanisms. To date, inadequate diffusion of antibiotics, efflux mechanisms, enzyme-facilitated deactivation, heterogenic function, slow development rate or persistent cells, and biofilm phenotype adaptive mechanisms have been identified [
39,
40]. We hypothesized that octoprohibitin could evade these defense mechanisms because it produced a potential disruption of the pre-formed
A. baumannii biofilm in a concentration-dependent manner. Although octoprohibitin had significant antibiofilm effects in CV staining, further experiments are essential to define the exact inhibition and eradication mechanism of
A. baumannii and its biofilm.
AMPs are considered to possess cell selectivity by differentiating the microbial membrane structure, and act only against pathogenic microorganisms without generating toxicity against host cells [
41]. Several AMPs have been tested in vitro for their efficiency in antimicrobial activity, but their application in vivo is limited because of their toxicity to eukaryotic cells, susceptibility to proteolytic degradation, and the development of allergies [
42,
43]. Since octoprohibitin is positively charged and hydrophobic, we projected its selectivity towards bacterial cells and lowered its interaction with eukaryotic cells for the least toxic effects. Since the hemolysis concentration (600 µg/mL) and HEK293T viability reduction concentration (800 µg/mL) were higher than the MIC (200 µg/mL) and MBC (400 µg/mL), we confirmed the safety of octoprohibitin for application in zebrafish to determine its effectiveness in vivo against
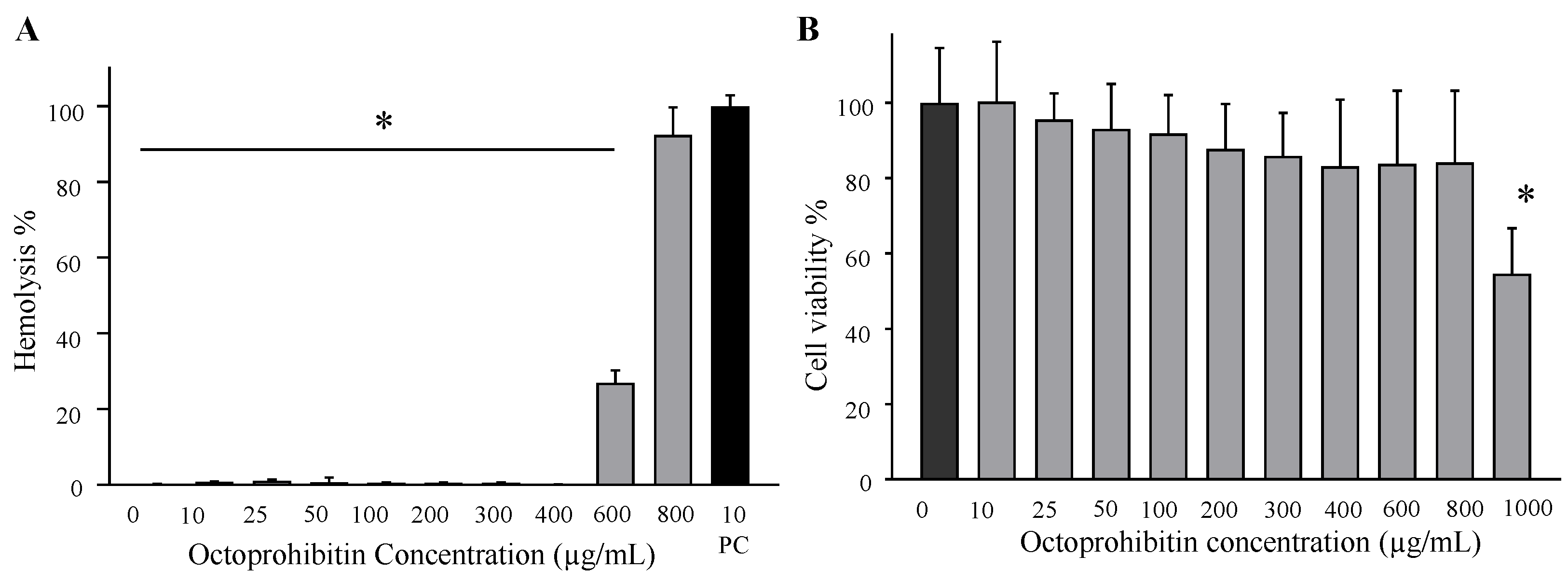
A. baumannii infection.
In our previous study, we developed a zebrafish model of
A. baumannii infection and successfully tested it with octopromycin [
17]. Because we could confirm the low toxicity of octoprohibitin in vitro, the zebrafish model was used to observe how octoprohibitin treatment controls
A. baumannii infection in vivo. As expected, octoprohibitin demonstrated its antimicrobial activity in zebrafish against
A. baumannii infection significantly (
p < 0.05), showing high CS% compared to that of the control group. Since visible pathological signs of hemorrhage were observed in untreated fish in the gill and abdomen area, we expanded our study to determine alterations in tissue levels by histopathology analysis. Although
A. baumannii was not well tested in the zebrafish model previously, we identified a significant level of tissue alterations in the gut, kidney, and gills. Generally, these tissues show necrosis, reduced cell volume, and possible hemorrhages. In particular, hyperplasia of goblet cells in the gut tissue and hyperplasia and clubbing of secondary lamella in the gill confirmed the inflammatory conditions caused by
A. baumannii in the PBS-treated control group. However, the low level of tissue damage in the octoprohibitin-treated group was confirmed by the survival data and suggested a high level of control zebrafish from
A. baumannii infection. IDR-1, which is a derivative of bovine bactenecin, does not possess direct antimicrobial properties in vitro but was found to be protective in several mouse models infected with Gram-negative and Gram-positive pathogenic bacteria [
44]. Likewise, some AMPs exhibit immunomodulatory actions in the host, such as altered chemokine and cytokine release, leukocyte activation, mast cell degranulation, anti-endotoxin generation, and chemotaxis strengthening immunity [
45]. As octoprohibitin was able to control
A. baumannii infection in zebrafish, we can conclude that protection is either by directly killing the bacteria or indirectly modulating the immunity of the host, and/or by both actions. Further experiments are essential to determine the exact mode of action of octoprohibitin in controlling
A. baumannii infection in vivo.
Even though we have tested octoprohibitin antibacterial activity against Gram-negative A. baumannii, owing to its vast mode of action, such as membrane permeabilization, ROS generation, DNA binding, and antibiofilm activities, octoprohibitin may produce antibacterial activities against other Gram-negative and Gram-positive bacteria, antiviral activities, and antifungal activities. Additionally, owing to the ROS generation properties of octoprohibitin, it may be developed as an anticancer drug. However, further studies on octoprohibitin are essential for elaborating its applications in other fields.
4. Materials and Methods
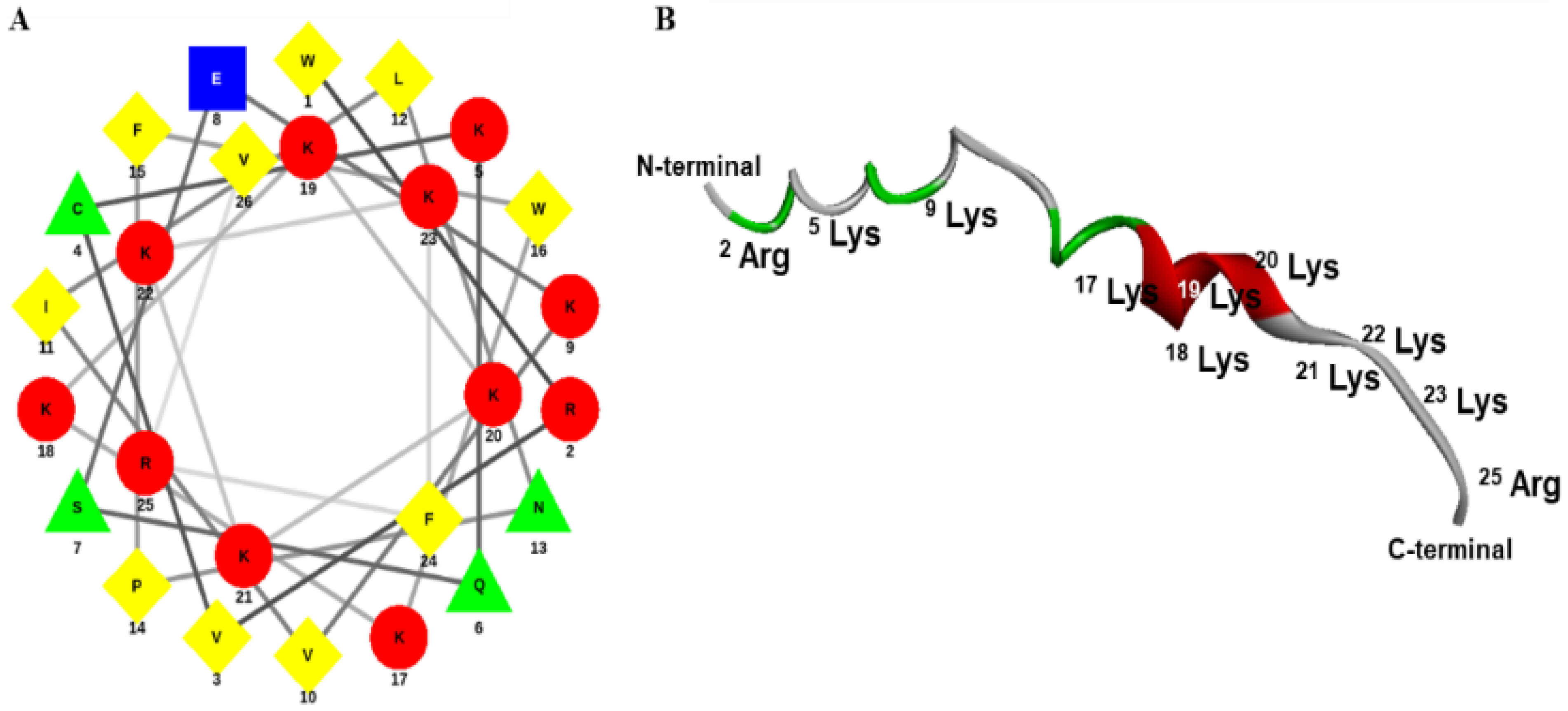
4.1. Design, Synthesis, and Characterization of Octoprohibitin
The transcriptome database of
O. minor was screened for AMPs, and the prohibitin-2 cDNA sequence was considered a candidate for designing a novel AMP. The prohibitin-2 cDNA sequence was submitted to the National Center for Biotechnology Information (NCBI) (
https://www.ncbi.nlm.nih.gov/, accessed on 7 January 2022) database with accession number MW939430. To design octoprohibitin, the C-terminal region of the prohibitin-2 AA sequence was selected as a template. The novel AMP, octoprohibitin (WRVCKQSEKVILNPFWKKKKKKKFRV), which contains 26 AA residues, was synthesized by a solid-phase peptide synthesis technique (AnyGen Co., Gwangju, Korea), and purified by reverse-phase high-performance liquid chromatography using a SHIMADZU C18 analytical column (Shimadzu HPLC LabSolution, Kyoto, Japan). Mass analysis was conducted using linear MALDI-TOF mass spectrometry (AXIMA Assurance, MALDI-TOF; Shimadzu, Kyoto, Japan). The three-dimensional structure of octoprohibitin was generated using APPTEST (
https://research.timmons.eu/apptest, accessed on 7 January 2022), and the images were visualized using Discovery Studio Visualizer (Version 21.1.0.20298, Biovia, CA, USA). The helical wheel projection of octoprohibitin was derived by Netwheel, Peptides Helical Wheel, and Net projection maker (
http://lbqp.unb.br/etWheels/, accessed on 7 January 2022) [
18].
4.2. Analysis of Octoprohibitin Antibacterial Activity against A. baumannii
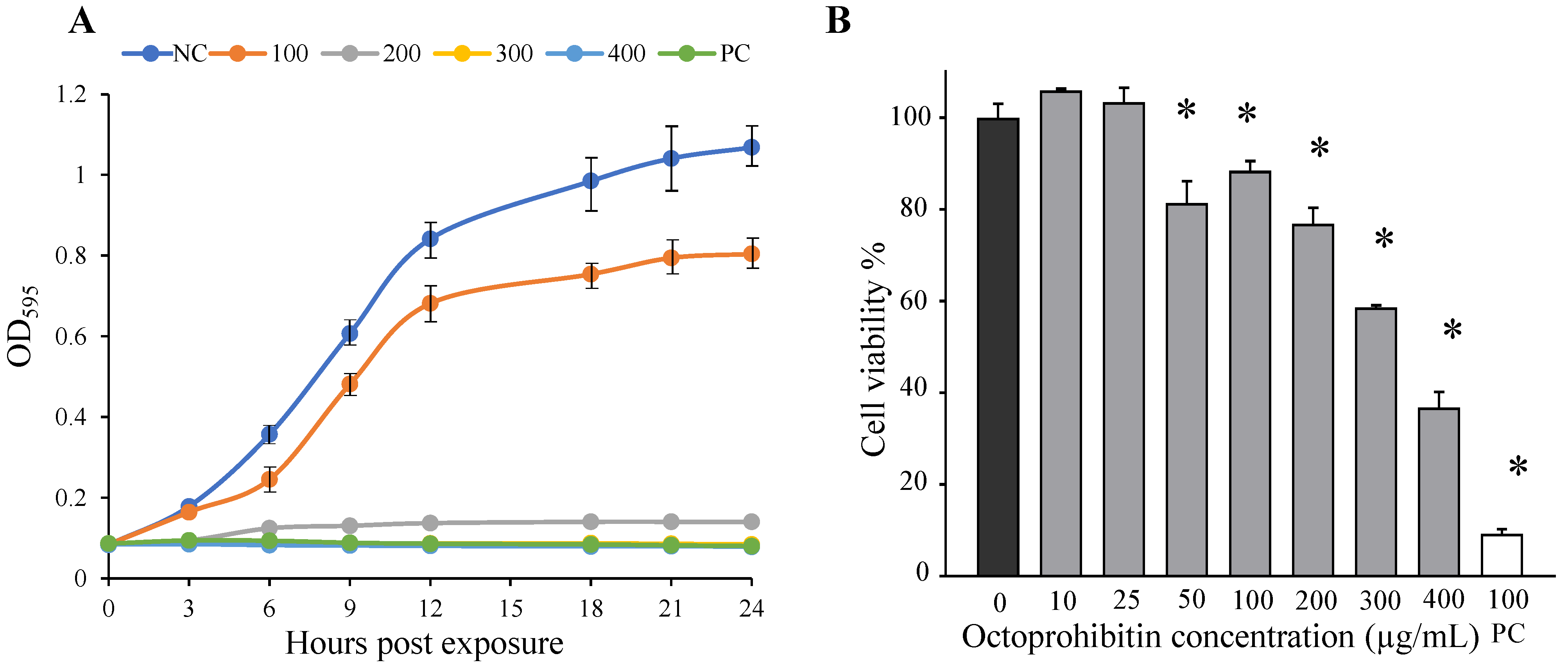
We conducted an initial screening of octoprohibitin activity against A. baumannii at pH 7.4 using the broth microdilution method. Since it showed relatively lower activity, we changed the pH of the bacterial culture broth to 7, 8, and 9 and conducted the sensitivity test similarly, and found that pH 8 was the most effective pH level. All experiments were conducted in LB broth and LB agar at pH 8. The antibacterial activity of octoprohibitin was determined by time-kill kinetics, MIC, MBC, and bacterial viability using standard assays.
MIC and time-kill kinetics assays were conducted using the broth microdilution method, whereas MBC was determined using the agar plating method according to the CLSI guidelines (M07-A10). Broth microdilution was conducted on A. baumannii (1 × 106 colony-forming units per milliliter; CFU/mL) in a 96-well plate (200 µL/well), and octoprohibitin was treated with different concentrations (0–500 µg/mL). The plate was incubated at 25 °C for 24 h, and bacterial growth was evaluated at 3 h intervals by measuring the optical density at 595 nm (OD595) using a microplate spectrophotometer (Bio-Rad, Saint Louis, USA). Growth kinetics are illustrated for each treatment level in a line graph with OD595 vs. time. The lowest octoprohibitin concentration that did not cause any OD595 changes for 24 h (no bacterial growth) was selected as the MIC. To determine the MBC, octoprohibitin treated at ≥MIC bacterial broths (100 µL) from 96-well plates was spread on LB agar plates and incubated at 25 °C for 24 h. The lowest octoprohibitin concentration that did not produce colonies on the agar plate was selected as the MBC against A. baumannii. The same procedure was conducted using chloramphenicol (100 µg/mL) as a positive control to determine MIC and MBC.
To determine the reduction in bacterial viability with octoprohibitin treatment, the MTT assay was conducted according to the method described by Dananjaya et al. [
46] Briefly,
A. baumannii (1 × 10
6 CFU/mL) was treated with octoprohibitin (0–400 µg/mL) and incubated at 25 °C for 4 h. Bacteria were isolated by centrifugation at 1500×
g for 10 min and washed with 1 × PBS. The bacterial cells were then treated with 10 µL of MTT reagent (5 µg/mL) (Sigma Aldrich, St. Louis, MO, USA). After incubation for 30 min, 50 µL of dimethyl sulfoxide (DMSO) was added to the mixture (Sigma Aldrich, St. Louis City, USA) to dissolve the formazan, and the absorbance was measured using a microplate spectrophotometer at OD
595. 4.3. FE-SEM Analysis for Morphological and Structural Changes in Octoprohibitin-Treated A. baumannii
To determine whether octoprohibitin induces morphological and ultrastructural alterations on
A. baumannii surfaces, FE-SEM analysis was conducted as described by Jayathilaka et al. [
13]. In brief,
A. baumannii (1 × 10
6 CFU/mL) was treated with the MIC (200 µg/mL) and MBC (400 µg/mL) of octoprohibitin, 1 × PBS as the negative control, and chloramphenicol (100 µg/mL) as the positive control. After incubation at 25 °C for 9 h, the bacterial cells were collected by centrifugation at 1500 ×
g for 10 min and washed with 1 × PBS. The isolated bacterial cells were pre-fixed with 2.5% glutaraldehyde (Pharmacia, Uppsala, Sweden) for 20 min. Each sample was washed with 1 × PBS and dehydrated using a series of ethanol concentrations (30, 50, 70, 80, 90, and 100%). Cells were coated with platinum using an ion sputter (E_1030, Hitachi, Japan) and observed under FE-SEM (MERLIN™, Carl Zeiss, Germany).
4.4. Analysis of Membrane Permeability Alterations and ROS Generation
Octoprohibitin-treated
A. baumannii was subjected to PI uptake assay and H
2DCFDA staining to determine permeability alterations and ROS generation, respectively. FCM was conducted to determine the fluorescence level, and CFMC was conducted to capture stained cell images, according to a previously described method [
17,
46]. Briefly,
A. baumannii (1 × 10
6 CFU/mL) was treated with octoprohibitin at MIC (200 µg/mL) and MBC (400 µg/mL), 1 × PBS was used as the negative control, and chloramphenicol (100 µg/mL) was used as the positive control. Bacterial cells were collected after incubation at 25 °C for 3 and 9 h for FCM and CFMC, respectively. Isolated cells were washed with 1 × PBS and resuspended in 1 mL of PBS.
For PI uptake assay, cells were treated with 50 µg/mL PI (Sigma Aldrich, Saint Louis, USA) and 40 µg/mL FDA (Sigma Aldrich, Saint Louis, USA) at room temperature (26 ± 2 °C) for 30 min in the dark. Similarly, samples for ROS generation were stained with H2DCFDA (50 μg/mL) (Invitrogen, Waltham, MA, USA) at room temperature for 30 min in the dark. Excess staining was removed by centrifugation and 1 × PBS washing.
To determine the fluorescence level, cell samples were resuspended in 1 mL of 1 × PBS, and the intensity was observed using flow cytometry (FACS Calibur, Becton Dickinson, Franklin Lakes, NJ, USA). For the PI uptake assay, the red fluorescence level was measured, and the green fluorescence level was measured for ROS generation.
To capture micrographs of fluorescent cells, bacterial samples were resuspended in 50 µL of 1 × PBS, and 5 µL was placed on a glass slide. Fluorescent cells were observed under CFMC with a scan head integrated into an Axiovert 200 M inverted microscope (Carl Zeiss, Jena, Germany). The red fluorescence of PI was observed at excitation and emission wavelengths of 535 and 617 nm, respectively. Green fluorescence of FDA and H2DCFDA was observed at the excitation and emission wavelengths of 488 and 535 nm, respectively.
4.5. Agarose Gel Retardation Assay for DNA Binding Affinity
Octoprohibitin-treated genomic DNA of
A. baumannii was subjected to an agarose gel retardation assay to determine the DNA-binding ability according to the method described by Jayathilaka et al. [
18]. Genomic DNA was isolated from
A. baumannii according to the manufacturer’s protocol using an AccuPrep
® Genomic DNA Extraction Kit (Bioneer, Daejeon, South Korea). Isolated DNA was quantified using a NanoDrop One
C Microvolume UV–Vis spectrophotometer (Thermo Scientific™, Waltham, MA, USA). Octoprohibitin treatment was performed on 200 ng of genomic DNA at increasing ratios (0:1, 0.25:1, 0.5:1, 1:1, 2.5:1, 5:1, 7.5:1, and 10:1) and incubated at 37 °C for 4 h. The total amount of DNA was loaded onto a 1% agarose gel with 2 µL of loading buffer (Takara Bio Inc. Kusatsu, Shiga, Japan) and electroporated in a mini-submarine electrophoresis system (Mupid
®-2plus, Takara Bio Inc. Kusatsu, Shiga, Japan) for 40 min. A photograph of the gel was captured using a gel imaging system (MaXidoc
Tm G2, DAIHAN
®, Republic of Korea). The intensity of each gel band was quantified using ImageJ software (ImageJ, ver. 1.6, Rasband, WS, USA), and normalized to the intensity of the untreated gel band.
4.6. Biofilm Inhibition and Eradication Activity Assay of Octoprohibitin
Biofilm formation inhibition and eradication assays were conducted on
A. baumannii planktonic and its biofilm, respectively, with CV staining according to the method described by Kim et al. [
47].
A. baumannii was cultured in LB broth (pH 8) supplemented with 0.1% glucose to enhance and stabilize biofilm formation. For the biofilm inhibition assay,
A. baumannii (1 × 10
6 CFU/mL) broth was added to a 96-well plate (100 µL/well), treated with octoprohibitin at different concentrations (0–1000 µg/mL) and chloramphenicol (100 µg/mL) as a positive control, and incubated for 24 h. For the biofilm eradication assay, initially,
A. baumannii (1 × 10
6 CFU/mL) broth (100 µL/well) was added and allowed to form biofilms on the wall of the 96-well plate. After biofilm formation with 24 h incubation at 25 °C, the supernatant was removed and replaced with fresh media. Octoprohibitin (0–800 µg/mL) or chloramphenicol (100 µg/mL) was added to the biofilm and incubated for 24 h at 25 °C.
After octoprohibitin treatment, the biofilm formation inhibition and biofilm eradication assay plates were used for CV staining. The supernatant was removed, and the wells were washed with 1 × PBS to remove planktonic bacteria. Absolute methanol (100 µL/well) was used to fix the biofilms for 10 min. After removal of methanol, 100 µL of 0.1% CV (Sigma-Aldrich, Burlington, MA, USA) was used to stain the biofilm at room temperature for 30 min. The CV was then removed, and the wells were washed with 1 × PBS to remove unbound CV. Biofilm-bound CV was dissolved by adding absolute ethanol (100 µL/well) and mixing. The absorbance of each well was measured using a microplate spectrophotometer at OD595. The biofilm formation inhibition and eradication percentages were calculated. Biofilm formation inhibition/eradication % = [1 − (Ab test/Ab negative control)] × 100%, where Ab test represents the absorbance value of octoprohibitin or chloramphenicol; Ab negative control represents the absorbance of the negative control (PBS). Ninety percent of the biofilm formation inhibition concentration and eradication concentration were calculated as MBIC and MBEC, respectively.
4.7. Hemolysis Assay and Cytotoxicity Assay for Determination of Octoprohibitin Toxicity
To determine the toxicity level of octoprohibitin, a hemolysis assay on mouse RBCs and a cytotoxicity test on HEK293T cells (American Type Culture Collection ATCC-11268) were conducted. For the hemolysis assay, RBCs were collected by centrifuging mouse blood at 1500× g and washed with 1 × PBS. Then, RBCs were suspended in 1 × PBS and treated with octoprohibitin at different concentrations (0–800 µg/mL), with 1 × PBS as the negative control and 1% (w/v) Triton x-100 (Sigma Aldrich, Saint Louis, MO, USA) as the positive control. After incubation at 37 °C for 4 h, the supernatant was separated by centrifugation at 1500× g and the absorbance was measured using a microplate spectrophotometer at 490 nm. The hemolysis percentage was calculated by normalizing with the 100% hemolysis absorbance using 1% Triton X-100. Hemolysis (%) = ((Ab test − Ab PBS)/(Ab triton− Ab PBS)) × 100%, where Ab test represents the absorbance of octoprohibitin-treated RBCs supernatant, Ab PBS represents the absorbance of PBS-treated RBCs supernatant, and Ab triton represents the absorbance of Triton X-100-treated RBCs supernatant.
An MTT assay was conducted to determine the change in viability of HEK293T cells treated with octoprohibitin. Briefly, HEK293T cells were seeded in Dulbecco’s modified Eagle’s medium (Invitrogen, Carlsbad, CA, USA) with 1% antibiotic/antimycotic solution (Glibco, Carlsbad, CA, USA) and 10% fetal bovine serum (Hyclone, Carlsbad, CA, USA). Then, the cells were seeded in a 96-well plate at a cell density of 1.5 × 106 cells/well and incubated at 37 °C in a 5% CO2 atmosphere. After 24 h of incubation, the supernatant was removed and replaced with a fresh medium. Octoprohibitin treatment was conducted in a concentration series (0–800 µg/mL) and incubated at 37 °C in a 5% CO2 atmosphere for 24 h. Culture media was replaced with 90 µL fresh media and 10 µL of MTT reagent (5 µg/mL). After incubation at 37 °C for 4 h, 50 µL DMSO was added to each well and mixed. Solubilized formazan was quantified by absorbance measurement using a microplate spectrophotometer at OD595.
4.8. In Vivo Octoprohibitin Efficacy Assay against A. baumannii-Infected Zebrafish
In vivo zebrafish experiments were performed according to the institutional animal ethics guidelines and under the supervision of the Committee of Chungnam National University (202103-CNU-072). Bacteria (
A. baumannii) challenge and octoprohibitin treatment were conducted intraperitoneally in zebrafish according to the method described by Liyanage et al. [
48]. Briefly, zebrafish were divided into three groups (30 fish per group in triplicate) and labeled; 1) only PBS-treated (negative control), 2)
A. baumannii challenged and 1 × PBS-treated, and 3)
A. baumannii challenged and octoprohibitin-treated. First, zebrafish were anesthetized using 160 μg/mL buffered tricaine (ethyl 3-aminobenzoate methanesulfonate; Sigma-Aldrich, USA) and injected with an infective dose of
A. baumannii (20 µL of 2.1 × 10
11 CFU/mL). Then, treatment was conducted with either 10 µL of octoprohibitin (10 µg/mL) or 10 µL of 1 × PBS. The negative control group was injected with 30 µL 1 × PBS. Fish were placed in tanks and maintained at 28 °C while observing mortality every 6 h for 48 h. The pathological signs were monitored in the remaining fish at 48 hpt. Finally, CS% was calculated.
For histopathological analysis, three zebrafish per group were sacrificed for collection of gut, kidney, and gill tissues at the end of the experiment. Histological analysis was performed according to the method described by Liyanage et al. [
48]. The collected tissues were processed, embedded, and sectioned to a thickness of 4 µm, and glass slides were prepared for each section. Hematoxylin and eosin staining was performed, and the slides were observed under a light microscope (Leica 3000 LED, Wetzlar, Germany).


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








