
Discovery of Putative Dual Inhibitor of Tubulin and EGFR by Phenotypic Approach on LASSBio-1586 Homologs

 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
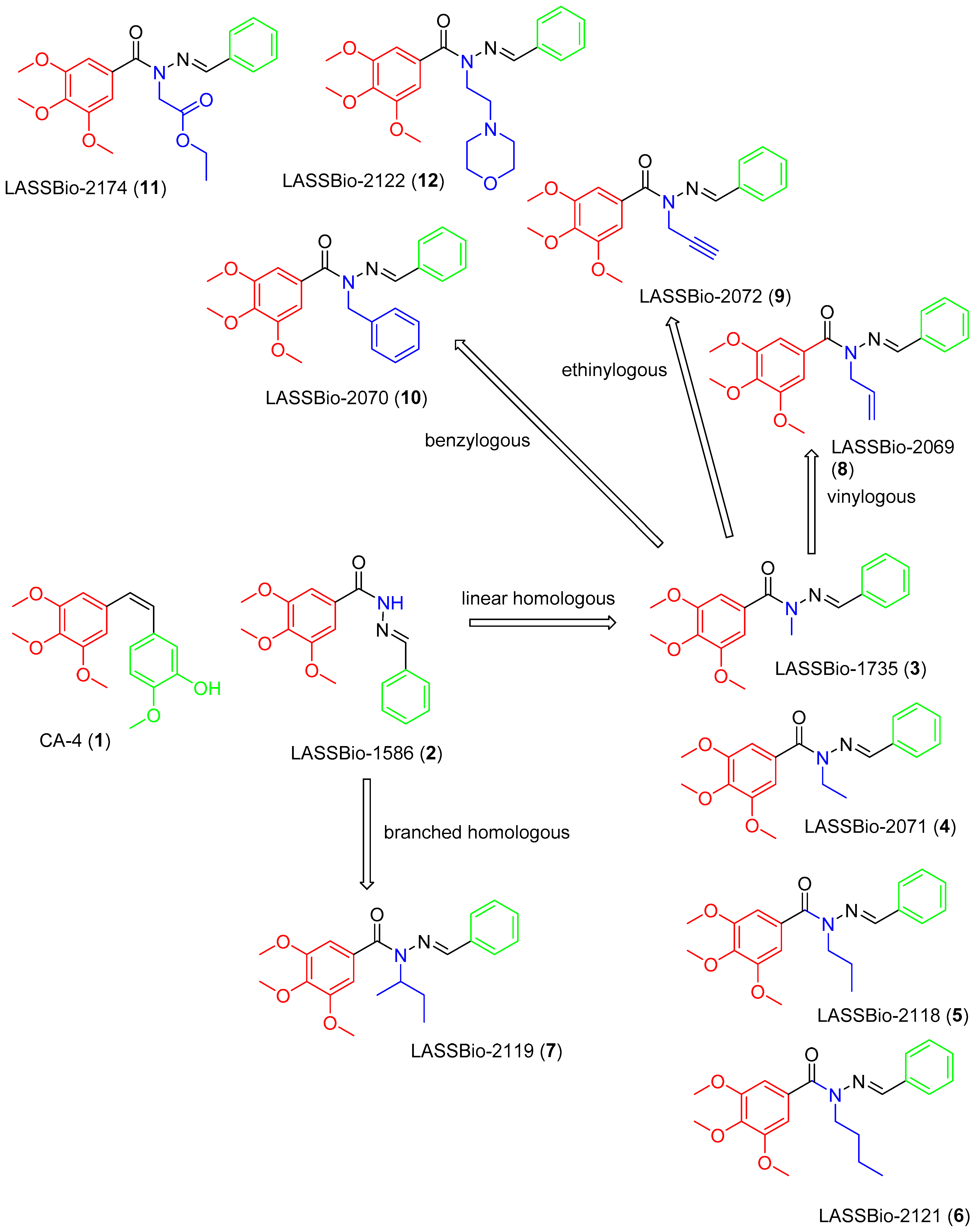
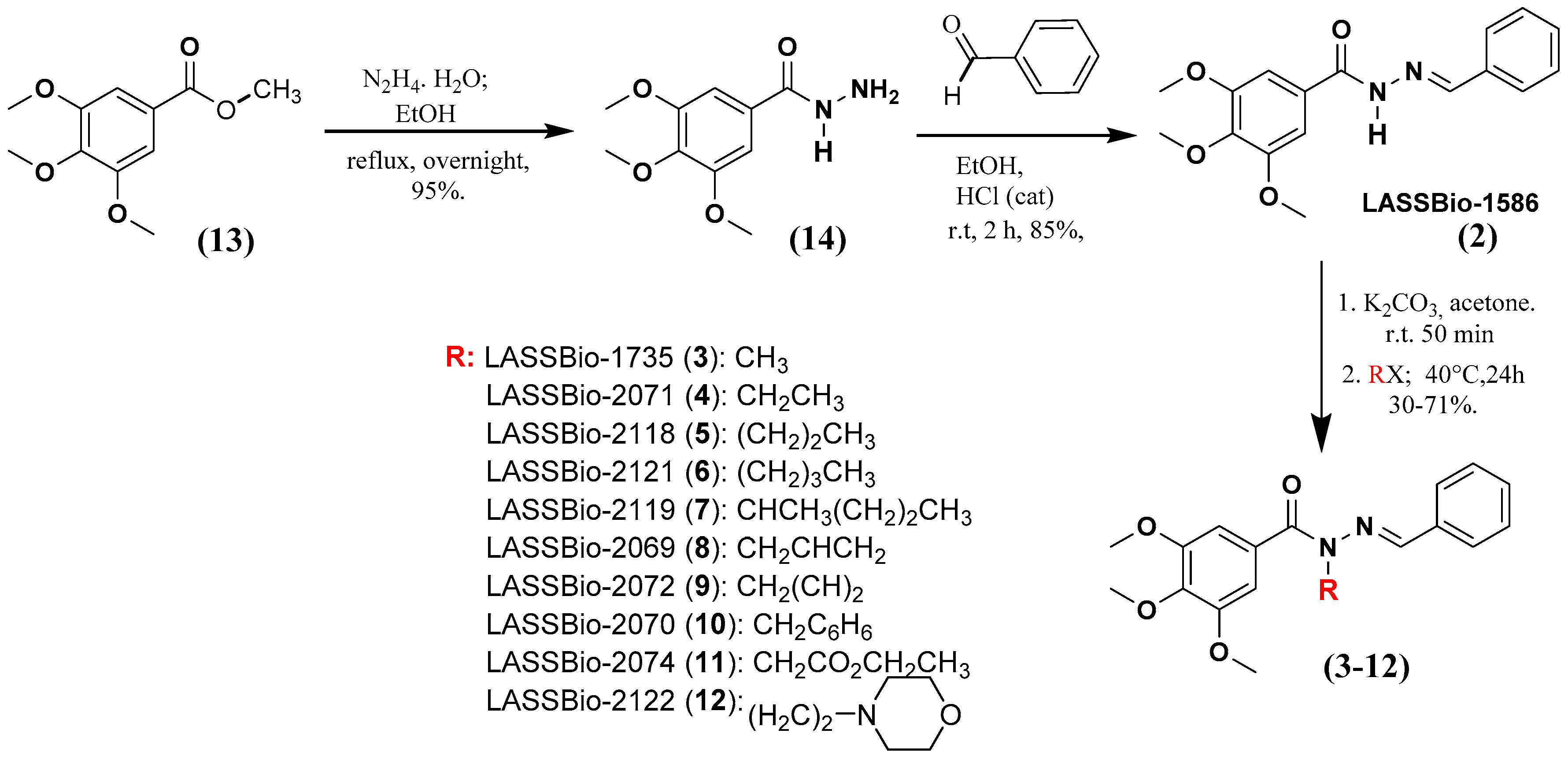
2.1. Chemistry
2.2. Pharmacological Experiments
2.2.1. Cellular Cytotoxicity
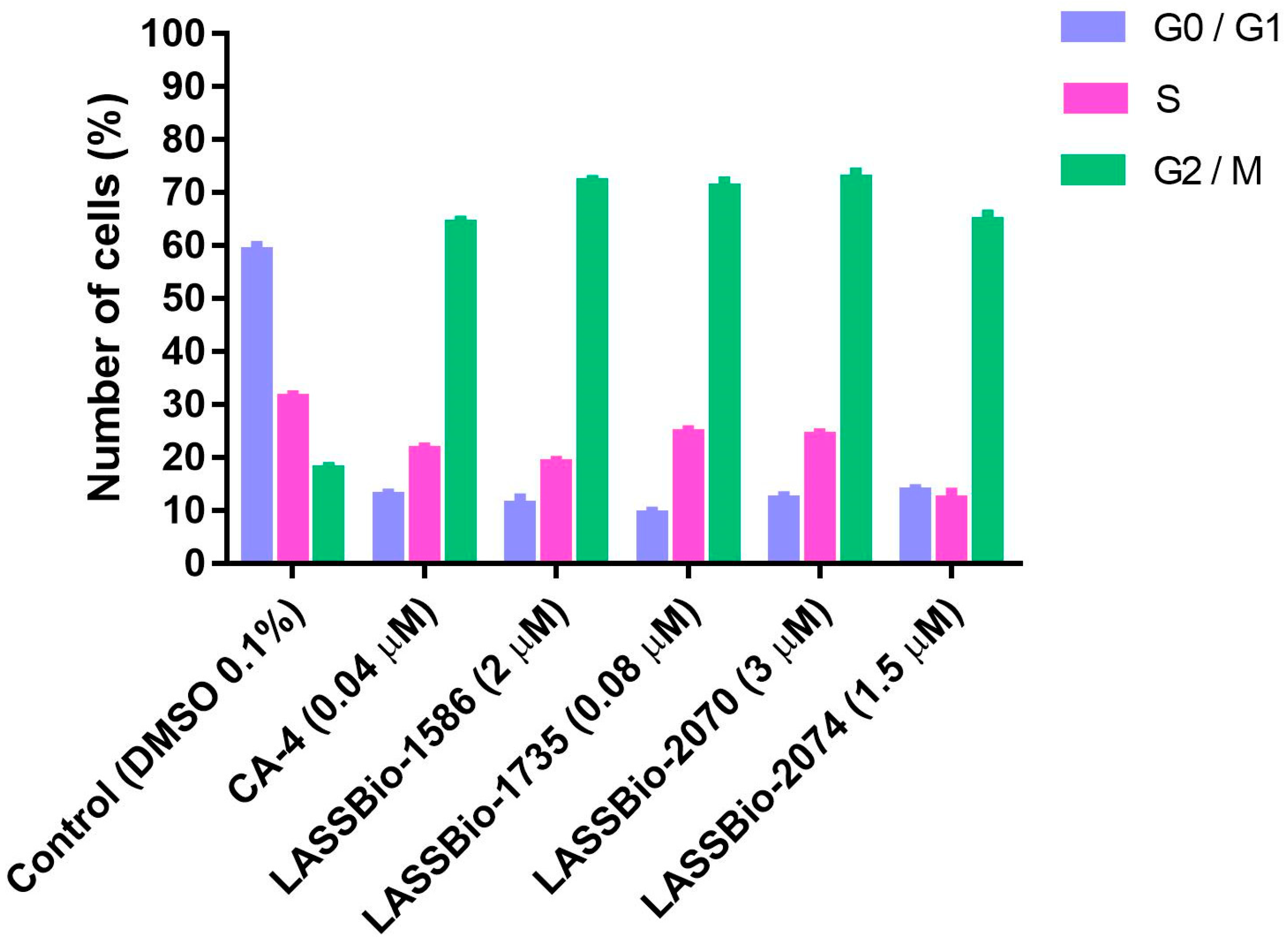
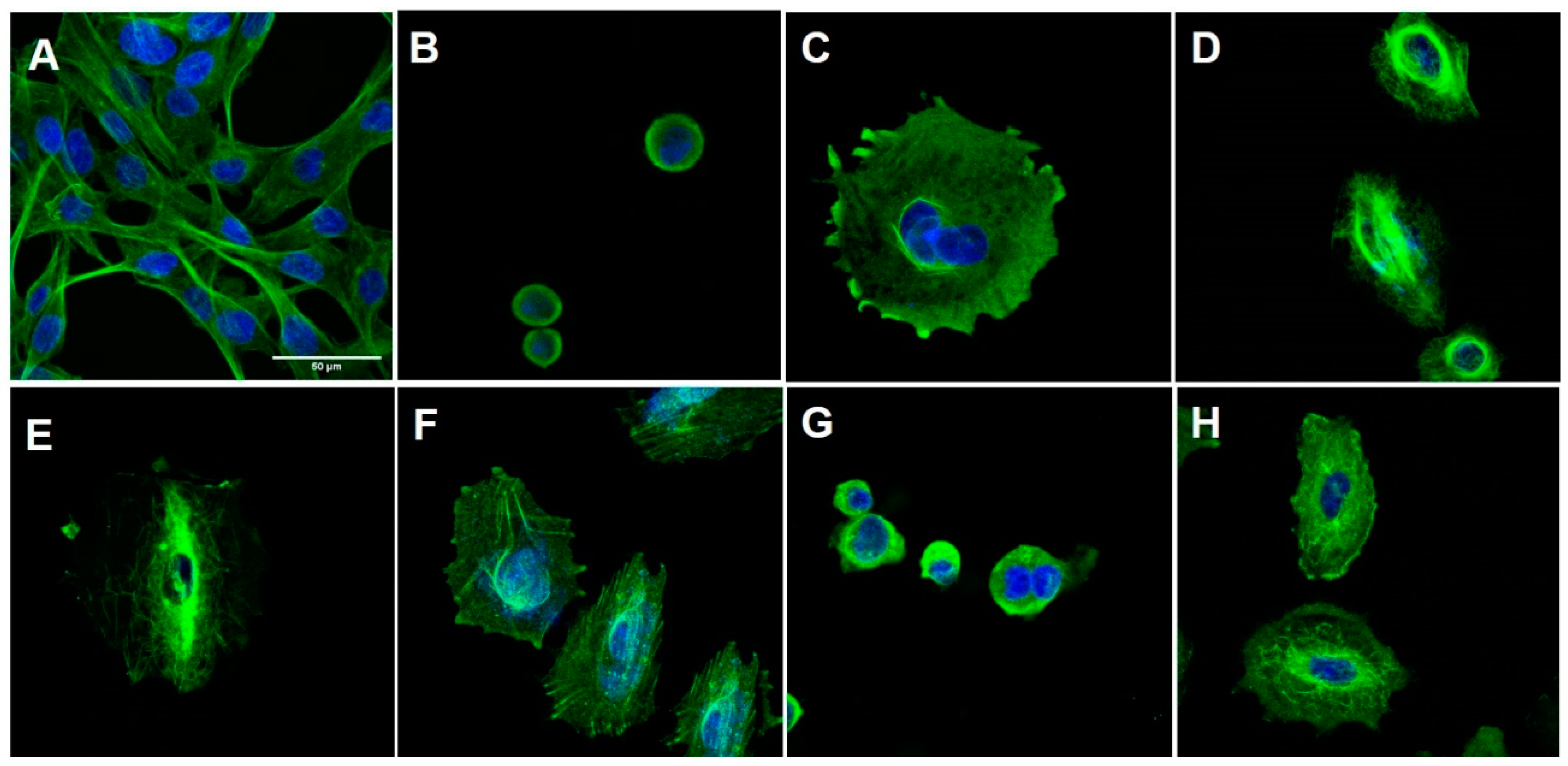
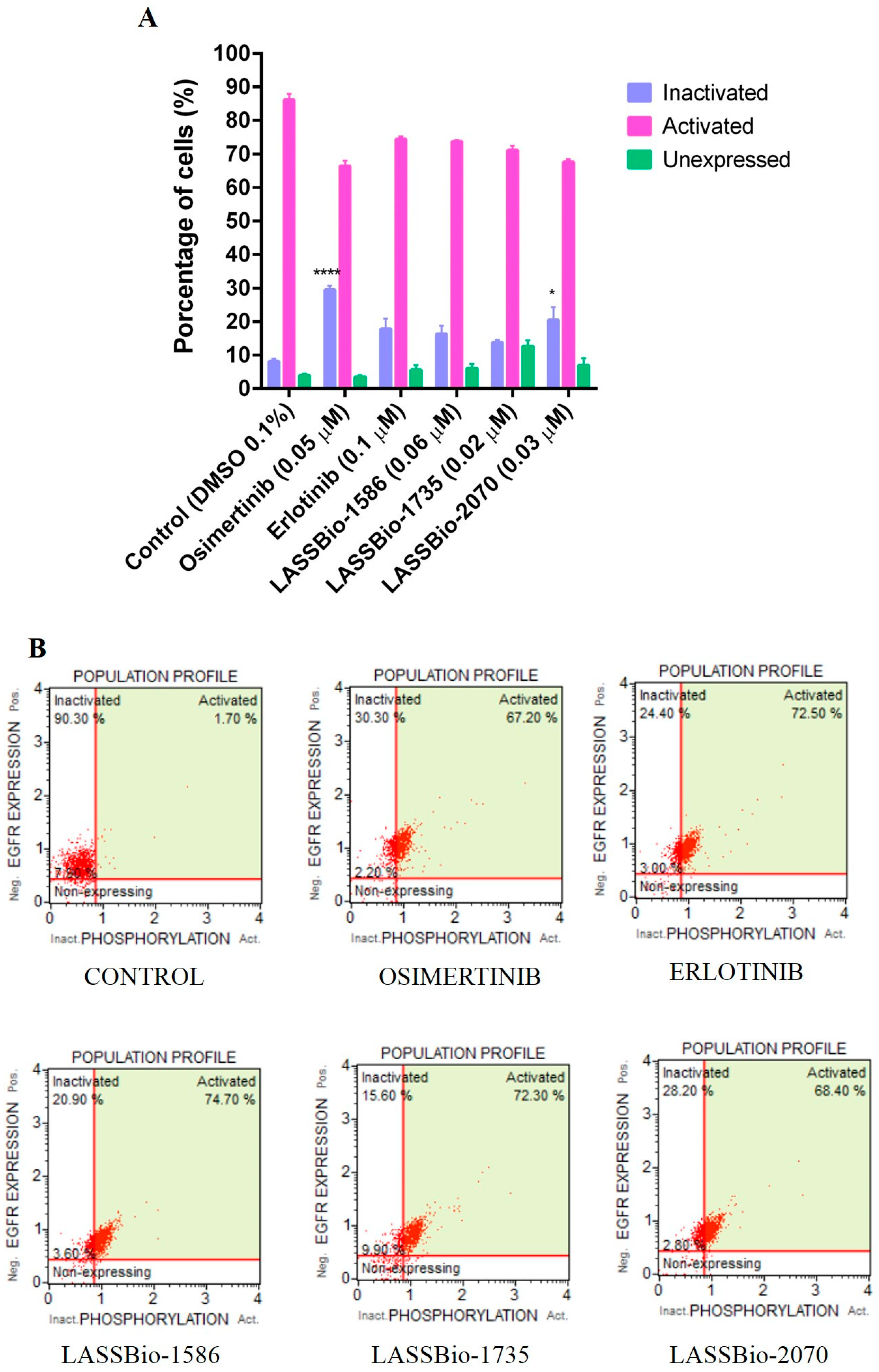
2.2.2. Flow Cytometry Analysis
2.3. Molecular Docking Studies
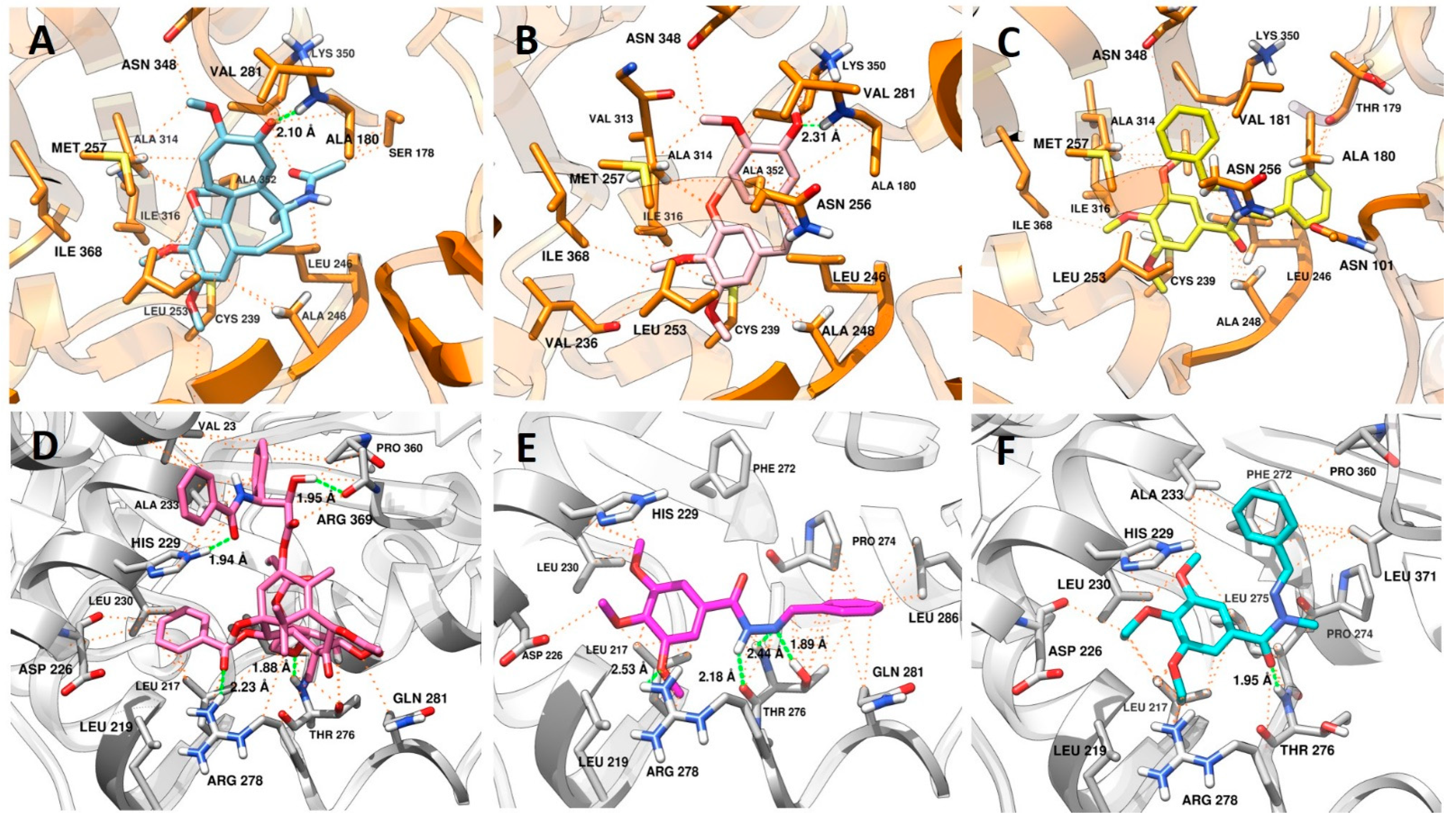
2.3.1. Docking in the Colchicine Binding Site
2.3.2. Docking in the Taxol Binding Site
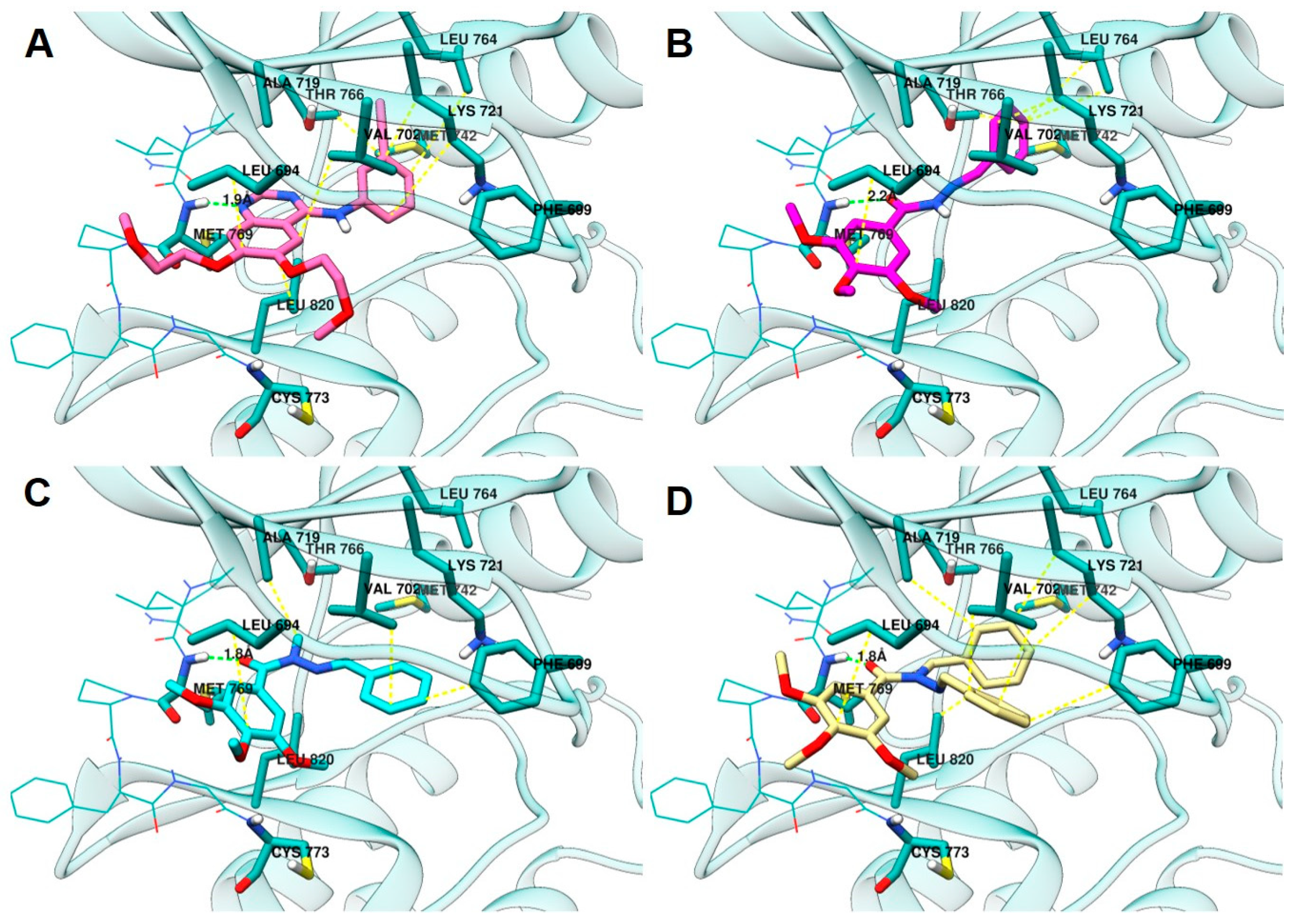
2.3.3. Docking in the EGFR
3. Material and Methods
3.1. Synthesis and Characterization
3.1.1. Procedure for the Preparation of (E)-N’-benzylidene-3,4,5-trimetoxybenzohydrazide (2; LASSBio-1586)
3.1.2. General Procedure for the Preparation of the N-Alkylated Acylhydrazones (3–12)
3.2. Cell Culture
3.3. Cell Viability Assay
3.4. Flow Cytometry Analysis
3.5. Immunofluorescence and Digital Image Acquisition
3.6. Molecular Docking Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Florian, S.; Mitchison, T.J. Anti-microtubule drugs. Methods Mol. Biol. 2016, 1413, 403–421. [Google Scholar] [CrossRef] [PubMed]
- Wordeman, L.; Vicente, J.J. Microtubule targeting agents in disease: Classic drugs, novel roles. Cancers 2021, 13, 5650. [Google Scholar] [CrossRef]
- Stanton, R.A.; Gernert, K.M.; Nettles, J.H.; Aneja, R. Drugs that target dynamic microtubules: A new molecular perspective. Med. Res. Rev. 2011, 31, 443–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, W. Resistance to anti-tubulin agents: From vinca alkaloids to epothilones. Cancer Drug Resist. 2019, 2, 82–106. [Google Scholar] [CrossRef]
- Yeung, S.C.; She, M.; Yang, H.; Pan, J.; Sun, L.; Chaplin, D. Combination chemotherapy including combretastatin A4 phosphate and paclitaxel is effective against anaplastic thyroid cancer in a nude mouse xenograft model. J. Clin. Endocrinol. Metab. 2007, 92, 2902–2909. [Google Scholar] [CrossRef]
- Nainwal, L.M.; Alam, M.M.; Shaquiquzzaman, M.; Marella, A.; Kamal, A. Combretastatin-based compounds with therapeutic characteristics: A patent review. Expert Opin. Ther. Pat. 2019, 29, 703–731. [Google Scholar] [CrossRef] [PubMed]
- do Amaral, D.A.; Cavalcanti, B.C.; Bezerra, D.P.; Ferreira, P.M.P.; Castro, R.P.; Sabino, J.R.; Machado, C.M.L.; Chammas, R.; Pessoa, C.; Sant’Anna, C.M.R.; et al. Docking, synthesis and antiproliferative activity of N-acylhydrazone derivatives designed as combretastatin A4 analogues. PLoS ONE 2014, 9, e85380. [Google Scholar] [CrossRef] [Green Version]
- Thota, S.; Rodrigues, D.A.; Pinheiro, P.S.M.; Lima, L.M.; Fraga, C.A.M.; Barreiro, E.J. N-acylhydrazones as drugs. Bioorg. Med. Chem. Lett. 2018, 28, 2797–2806. [Google Scholar] [CrossRef]
- Negi, A.S.; Gautam, Y.; Alam, S.; Chanda, D.; Luqman, S.; Sarkar, J.; Khan, F.; Konwar, R. Natural antitubulin agents: Importance of 3,4,5-trimethoxyphenyl fragment. Bioorg. Med. Chem. 2015, 23, 373–389. [Google Scholar] [CrossRef]
- Lima, L.M.; Alves, M.A.; do Amaral, D.N. Homologation: A versatile molecular modification strategy to drug discovery. Curr. Top. Med. Chem. 2019, 19, 1734–1750. [Google Scholar] [CrossRef]
- Mosman, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bargou, R.C.; Jürchott, K.; Wagener, C.; Bergmann, S.; Metzner, S.; Bommert, K.; Mapara, K.M.Y.; Winzer, K.J.; Dietel, M.; Dörken, B.; et al. Nuclear localization and increased levels of transcription factor YB-1 in primary human breast cancers are associated with intrinsic MDR1 gene expression. Nat. Med. 1997, 3, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Tsou, S.; Hou, M.; Hsu, L.; Chen, Y.; Chen, Y. Gain-of-function p53 mutant with 21-bp deletion confers susceptibility to multidrug resistance in MCF-7 cells. Int. J. Mol. Med. 2016, 37, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokharel, D.; Roseblade, A.; Oenarto, V.; Lu, J.F.; Bebawy, M. Proteins regulating the intercellular transfer and function of P-glycoprotein in multidrug-resistant cancer. Ecancermedicalscience 2017, 11, 768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borenfreund, E.; Puerner, J.A. Toxicity determined in vitro by morphological alterations and neutral red absorption. Toxicol. Lett. 1985, 24, 119–124. [Google Scholar] [CrossRef]
- Kars, M.D.; Işeri, O.D.; Gündüz, U. A microarray based expression profiling of paclitaxel and vincristine resistant MCF-7 cells. Eur. J. Pharmacol. 2011, 657, 4–9. [Google Scholar] [CrossRef]
- Ashraf, M.; Shaik, T.B.; Malik, M.S.; Syed, R.; Mallipeddi, P.L.; Vardhan, M.; Kamal, A. Design and synthesis of cis-restricted benzimidazole and benzothiazole mimics of combretastatin A-4 as antimitotic agents with apoptosis inducing ability. Bioorg. Med. Chem. Lett. 2016, 26, 4527–4535. [Google Scholar] [CrossRef]
- Shan, Y.; Zhang, J.; Liu, Z.; Wang, M.; Dong, Y. Developments of combretastatin A-4 derivatives as anticancer agents. Curr. Med. Chem. 2011, 18, 523–538. [Google Scholar] [CrossRef]
- Barreca, M.; Stathis, A.; Barraja, P.; Bertoni, F. An overview on anti-tubulin agents for the treatment of lymphoma patients. Pharmacol. Therapeut. 2020, 211, 107552. [Google Scholar] [CrossRef]
- Gigant, B.; Wang, C.; Ravelli, R.B.G.; Roussi, F.; Steinmetz, M.O.; Curmi, P.A.; Sobel, A.; Knossow, M. Structural basis for the regulation of tubulin by vinblastine. Nature 2005, 435, 519–522. [Google Scholar] [CrossRef]
- Steinmetz, M.O.; Prota, A.E. Microtubule-targeting agents: Strategies to hijack the cytoskeleton. Trends Cell Biol. 2018, 28, 776–792. [Google Scholar] [CrossRef] [PubMed]
- Sebahar, P.R.; Willardsen, A.; Anderson, M.B. Anticancer agents: VTA or VDA. Curr. Bioact. Compd. 2009, 5, 79–97. [Google Scholar] [CrossRef]
- Akhmanova, A.; Steinmetz, M.O. Control of microtubule organization and dynamics: Two ends in the limelight. Nat. Rev. Mol. Cell Biol. 2015, 16, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Pinard, P.V.; Fuentes, N.F.; Burd, B.; Angeletti, R.; Fiser, A.; Horwitz, S.B.; Orr, G.A. Insights into the mechanism of microtubule stabilization by Taxol. Proc. Natl. Acad. Sci. USA 2006, 103, 10166–10173. [Google Scholar] [CrossRef] [Green Version]
- Owens, J. Determining druggability. Nat. Rev. Drug Discov. 2007, 6, 187. [Google Scholar] [CrossRef]
- Agoni, C.; Olotu, F.A.; Ramharack, P.; Soliman, M.E. Druggability and drug-likeness concepts in drug design: Are biomodelling and predictive tools having their say? J. Mol. Model. 2020, 26, 120. [Google Scholar] [CrossRef] [PubMed]
- Salum, L.B.; Mascarello, A.; Canevarolo, R.R.; Altei, W.F.; Laranjeira, A.B.; Neuenfeldt, P.D.; Stumpf, T.R.; Delatorre, L.D.C.; Vollmer, L.L.; Daghestani, H.N.; et al. N-(1′-naphthyl)-3,4,5-trimethoxybenzohydrazide as microtubule destabilizer: Synthesis, cytotoxicity, inhibition of cell migration and in vivo activity against acute lymphoblastic leukemia. Eur. J. Med. Chem. 2015, 96, 504–518. [Google Scholar] [CrossRef]
- Wu, Y.L.; Tsuboi, M.; He, J.; John, T.; Grohe, C.; Majem, M.; Goldman, J.W.; Laktionov, K.; Kim, S.W.; Kato, T.; et al. Osimertinib in resected EGFR-mutated non–small-cell lung cancer. N. Engl. J. Med. 2020, 383, 1711–1723. [Google Scholar] [CrossRef]
- McLoughlin, E.C.; O´Boyle, N.M. Colchicine-binding site inhibitors from chemistry to clinic: A review. Pharmaceuticals 2020, 13, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Sun, H.; Xu, S.; Zhu, Z.; Xu, J. Tubulin inhibitors targeting the colchicine binding site: A perspective of privileged structures. Future Med. Chem. 2017, 9, 1756–1794. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yan, W.; Yu, Y.; Wang, Y.; Yang, T.; Xue, L.; Yuan, X.; Long, C.; Liu, Z.; Chen, X.; et al. The compound millepachine and its derivatives inhibit tubulin polymerization by irreversibly binding to the colchicine-binding site in β-tubulin. J. Biol. Chem. 2018, 293, 9461–9472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Miller, D.D.; Li, W. Molecular interactions at the colchicine binding site in tubulin: An X-ray crystallography perspective. Drug Dis. Today 2022, 27, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Deng, S.; Albadari, N.; Yun, M.K.; Zhang, S.; Li, Y.; Ma, D.; Parke, D.N.; Yang, L.; Seagroves, T.N.; et al. Design, synthesis, and biological evaluation of stable colchicine-binding site tubulin inhibitors 6-aryl-2-benzoyl-pyridines as potential anticancer agents. J. Med. Chem. 2021, 64, 12049–12074. [Google Scholar] [CrossRef]
- Debs, G.E.; Cha, M.; Liu, X.; Sindelar, C.V. Dynamic and asymmetric fluctuations in the microtubule wall captured by high-resolution cryoelectron microscopy. Proc. Natl. Acad. Sci. USA 2020, 117, 16976–16984. [Google Scholar] [CrossRef] [PubMed]
- Pawar, D.M.; Khali, A.A.; Hooks, D.R.; Collins, K.; Elliott, T.; Stafford, J.; Smith, L.; Noe, E.A. E and Z conformations of esters, thiol esters, and amides. J. Am. Chem. Soc. 1998, 120, 2108–2112. [Google Scholar] [CrossRef]
- Field, J.J.; Díaz, J.F.; Miller, J.H. The binding sites of microtubule-stabilizing agents. Chem. Biol. 2013, 20, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Sept, D. Taxol allosterically alters the dynamics of the tubulin dimer and increases the flexibility of microtubules. Biophys. J. 2008, 95, 3252–3258. [Google Scholar] [CrossRef] [Green Version]
- Korb, O.; Stützle, T.; Exner, T.E. Empirical scoring functions for advanced protein−ligand docking with PLANTS. J. Chem. Inf. Model. 2009, 49, 84–96. [Google Scholar] [CrossRef]
- Kümmerle, A.E.; Raimundo, J.M.; Leal, C.M.; da Silva, G.S.; Balliano, T.L.; Pereira, M.A.; de Simone, C.A.; Sudo, R.T.; Zapata-Sudo, G.; Fraga, C.A.M. Studies towards the identification of putative bioactive conformation of potent vasodilator arylidene N-acylhydrazone derivatives. Eur. J. Med. Chem. 2009, 44, 4004–4009. [Google Scholar] [CrossRef]
- Barreiro, E.J.; Kümmerle, A.E.; Fraga, C.A.M. The methylation effect in medicinal chemistry. Chem. Rev. 2011, 111, 5215–5246. [Google Scholar] [CrossRef]
- Heppner, D.E.; Günther, M.; Wittlinger, F.; Laufer, S.A.; Eck, M.J. Structural basis for EGFR mutant inhibition by trisubstituted imidazole inhibitors. J. Med. Chem. 2020, 63, 4293–4305. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhou, Z.; Sun, X.; Yang, Z.; Zheng, P.; Xu, S.; Zhu, W. The new opportunities in medicinal chemistry of fourth-generation EGFR inhibitors to overcome C797S mutation. Eur. J. Med. Chem. 2021, 210, 112995. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Pawara, R.; Ansari, A.; Surana, S. Recent updates on third generation EGFR inhibitors and emergence of fourth generation EGFR inhibitors to combat C797S resistance. Eur. J. Med. Chem. 2017, 142, 32–47. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, R.; Refolo, M.G.; Lippolis, C.; Carella, N.; Messa, C.; Cavallini, A.; Carr, B.I. Modulation of Regorafenib effects on HCC cell lines by epidermal growth fator. Cancer Chemother. Pharmacol. 2015, 75, 1237–1245. [Google Scholar] [CrossRef] [Green Version]
- Moskot, M.; Gabig-Cimińska, M.; Jakóbkiewicz-Banecka, J.; Węsierska, M.; Bocheńska, K.; Węgrzyn, G. Cell cycle is disturbed in mucopolysaccharidosis type II fibroblasts, and can be improved by genistein. Gene 2016, 585, 100–103. [Google Scholar] [CrossRef]
- Filgueiras, M.C.; Morrot, A.; Soares, P.M.G.; Costa, M.L.; Mermelstein, C. Effects of 5-fluorouracil in nuclear and cellular morphology, proliferation, cell cycle, apoptosis, cytoskeletal and caveolar distribution in primary cultures of smooth muscle cells. PLoS ONE 2013, 8, e63177. [Google Scholar] [CrossRef] [Green Version]
- Stamos, J.; Sliwkowski, M.X.; Eigenbrot, C. Structure of the epidermal growth factor receptor kinase domain alone and in complex with a 4-anilinoquinazoline inhibitor. J. Biol. Chem. 2002, 277, 46265–46272. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.E.; Ayaz, P.; Zhu, S.J.; Zhao, P.; Liang, L.; Zhang, C.H.; Wu, Y.C.; Li, J.L.; Choi, H.G.; Huang, X.; et al. Structural basis of AZD9291 selectivity for EGFR T790M. J. Med. Chem. 2020, 63, 8502–8511. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | ||||||
|---|---|---|---|---|---|---|
| Compounds | R | Overall Yields (%) | Physical Appearance | Melting Point (°C) | Determined Purity (%) | Retention Time in HPLC (min) |
| LASSBio-1586 (2) | H | 70 | white powder | 138.33 | 99.94 | 9.63 |
| LASSBio-1735 (3) | Methyl | 71 | white powder | 110.56 | 99.90 | 9.15 |
| LASSBio-2071 (4) | Ethyl | 60 | white powder | 83.94 | 99.14 | 9.27 |
| LASSBio-2118 (5) | N-propyl | 28 | yellow oil | - | 98.04 | 9.55 |
| LASSBio-2121 (6) | N-butyl | 26 | yellow oil | - | 99.4 | 10.86 |
| LASSBio-2119 (7) | Sec-butyl | 60 | yellow oil | - | 99.8 | 12.40 |
| LASSBio-2069 (8) | Allyl | 71 | white powder | 64.47 | 98.23 | 6.37 |
| LASSBio-2072 (9) | Propynyl | 70 | white powder | 133.91 | 97.61 | 8.83 |
| LASSBio-2070 (10) | Benzyl | 46 | white powder | 130.12 | 98.33 | 6.38 |
| LASSBio-2074 (11) | Methylene Carbethoxy | 65 | white powder | 91.67 | 98.00 | 9.69 |
| LASSBio-2122 (12) | Morpholine Ethyl | 36 | white powder | 147.64 | 98.34 | 9.27 |
| Compounds | CC50 (µM) (CI) | |||||||
|---|---|---|---|---|---|---|---|---|
| HL-60 | H1975 | H292 | LoVo | MCF-7 | PC-3 | PC-9 | GM16000 | |
| CA-4 (1) | 0.077 (0.029–0.201) | 0.035 (0.016–0.079) | 0.013 (0.006–0.02) | 0.039 (0.015–0.079) | 8.58 (5.407- 10.61) | 0.096 (0.072–0.2829) | 0.01 (0.008–0.020) | 0.064 (0.023–0.176) |
| Pelitinib | 2.079 (1.246–3.468) | 0.512 (0.433–0.605) | 0.07835 (0.05–0.012) | 0.037 (0.014–0.069) | 3.91 (3.103–4.927) | 2.485 (1.809–3.413) | 0.0591 (0.037–0.069) | 2.749 (2.223–3.399) |
| LASSBio-1586 (2) | 2.127 (1.784–2.535) | 1.568 (1.271–1.935) | 1.398 (1.048–1.864) | 0.039 (0.015–0.069) | 7.53 (5.217–10.87) | 1.619 (1.295–2.023) | 0.587 (0.512–0.672) | 1.507 (1.103–2.058) |
| LASSBio-1735 (3) | 0.039 (0.023–0.065) | 0.079 (0.043–0.143) | 0.250 (0.174–0.357) | 0.149 (0.092–0.243) | 0.276 (0.137–0.557) | 0.201 (0.117–0.346) | 0.090 (0.045- 0.179) | 0.131 (0.068–0.254) |
| LASSBio-2071 (4) | 0.231 (0.113- 0.475) | 4.861 (1.504–5.721) | 2.546 (1.443–4.491) | 0.301 (0.162–0.559) | 1.238 (0.266–5.756) | 1.660 (1.248–2.207) | 0.384 (0.205–0.718) | 5.127 (2.344–11.21) |
| LASSBio-2118 (5) | 0.301 (0.190–0.478) | 0.775 (0.397–1.512) | 3.722 (2.177–6.364) | 0.031 (0.005–0.192) | 0.144 (0.030–0.670) | 0.432 (0.250–0.746) | 0.136 (0.071- 0.258) | 0.843 (0.391–1.818) |
| LASSBio-2121 (6) | 2.171 (1.678–2.809) | 2.935 (1.035–4.323) | 9.553 (6.323–14.43) | 0.5 (0.256–0.969) | 0.500 (0.187–1.333) | 0.696 (0.445–1.088) | 0.096 (0.050–0.184) | 0.961 (0.385–2.397) |
| LASSBio-2119 (7) | ND * | ND * | ND * | ND * | ND * | ND * | ND * | ND * |
| LASSBio-2069 (8) | 1.348 (0.956–1.901) | 3.739 (1.948–4.741) | 0.778 (0.440–1.378) | 0.298 (0.137–0.650) | 1.596 (0.589–4.324) | 0.462 (0.240–0.889) | 0.387 (0.224–0.667) | 0.830 (0.317–2.174) |
| LASSBio-2072 (9) | 0.158 (0.088–0.283) | 3.979 (1.195–5.253) | 3.144 (1.825–5.416) | 0.399 (0.202–0.786) | 2.318 (1.297–8.07) | 2.250 (1.556–3.252) | 0.460 (0.250–0.846) | 0.612 (0.329–1.138) |
| LASSBio-2070 (10) | 0.172 (0.092- 0.322) | 2.791 (1.374–4.81) | 0.883 (0.395–1.974) | 0.208 (0.100–0.431) | 1.7 (0.586–4.932) | 0.361 (0.171–0.761) | 0.337 (0.173–0.654) | 1.046 (0.498–2.196) |
| LASSBio-2074 (11) | 0.126 (0.061–0.262) | 1.204 (0.602–4.013) | 0.7503 (0.325–1.729) | 0.106 (0.039–0.285) | 0.869 (0.175–2.305) | 1.361 (0.772–2.397) | 0.202 (0.112–0.364) | 4.194 (1.767–9.955) |
| LASSBio-2122 (12) | 4.150 (2.761–6.238) | 2.482 (1.574–4.881) | 2.791 (1.362–5.719) | 0.421 (0.225–0.787) | 4.114 (2.546–6.649) | 2.396 (1.385–4.146) | 0.871 (0.496–1.528) | 5.649 (4.905–6.505) |
| Compounds | SI | ||||||
|---|---|---|---|---|---|---|---|
| GM16000/ HL-60 | GM16000/ H1975 | GM16000/ H292 | GM16000/ LoVo | GM16000/ MCF-7 | GM16000/ PC-3 | GM16000/ PC-9 | |
| CA-4 (1) | 0.8 | 1.8 | 4.5 | 1.6 | 0.007 | 0.7 | 6.4 |
| Pelitinib | 1.3 | 5.4 | 35.3 | 74.3 | 0.7 | 1.1 | 46.5 |
| LASSBio-1586 (2) | 0.7 | 1.0 | 1.0 | 39.0 | 0.2 | 0.9 | 2.6 |
| LASSBio-1735 (3) | 3.4 | 1.7 | 0.5 | 0.9 | 0.5 | 0.6 | 1.4 |
| LASSBio-2071 (4) | 22.2 | 1.0 | 2.0 | 17.0 | 4.1 | 3.0 | 13.3 |
| LASSBio-2118 (5) | 2.8 | 1.1 | 0.2 | 27.2 | 5.9 | 1.9 | 6.2 |
| LASSBio-2121 (6) | 0.4 | 0.3 | 0.1 | 2.0 | 1.9 | 1.4 | 10.0 |
| LASSBio-2069 (8) | 0.6 | 0.2 | 1.0 | 2.8 | 0.5 | 1.8 | 2.1 |
| LASSBio-2072 (9) | 4.0 | 0.2 | 0.2 | 1.5 | 0.3 | 0.3 | 1.3 |
| LASSBio-2070 (10) | 6.0 | 0.4 | 1.2 | 5.0 | 0.6 | 2.9 | 3.1 |
| LASSBio-2074 (11) | 33.3 | 3.5 | 5.6 | 39.5 | 4.8 | 3.0 | 20.8 |
| LASSBio-2122 (12) | 1.4 | 2.3 | 2.0 | 13.4 | 1.4 | 2.3 | 6.5 |
| Compounds | CC50 (µM) (CI) | |||||
|---|---|---|---|---|---|---|
| HL-60 | H1975 | H292 | LoVo | PC-9 | GM16000 | |
| CA-4 (1) | 1.299 (0.639–2.635) | 0.058 (0.024–0.140) | 0.091 (0.057–0.143) | 0.1795 (0.073–0.439) | 0.025 (0.011–0.055) | 0.016 (0.004–0.07) |
| Pelitinib | 3.419 (2.938–3.980) | 2.569 (1.993–3.311) | 0.480 (0.343–0.731) | 1.697 (1.386–2.078) | 0.325 (0.258–0.411) | 0.899 (0.6–1.35) |
| LASSBio-1586 (2) | 3.592 (3.099–4.163) | 2.017 (0.054–0.171) | 2.186 (1.666–2.870) | 1.720 (1.266–2.338) | 0.940 (0.761–1.163) | 0.763 (0.48–1.215) |
| LASSBio-1735 (3) | 8.768 (7.584–10.14) | 0.096 (1.600–2.542) | 0.152 (0.082–0.281) | 0.479 (0.273–0.841) | 1.341 (1.032–1.744) | 0.059 (0.019–0.181) |
| LASSBio-2071 (4) | 6.033 (4.064–8.957) | 4.905 (3.573–6.732) | 0.300 (0.191–0.980) | 3.052 (1.547–6.019) | 0.881 (0.544–1.427) | 1.996 (1.134–3.513) |
| LASSBio-2118 (5) | 1.457 (0.759–2.796) | 1.124 (0.557–2.266) | 0.181 (0.052–0.629) | 0.740 (0.209–2.622) | 0.263 (0.133–0.52) | 1.096 (0.60–2.00) |
| LASSBio-2121 (6) | 2.606 (1.605–4.231) | 1.713 (1.041–2.818) | 0.215 (0.081–0.567) | 2.004 (0.832–4.823) | 0.327 (0.173–0.618) | 1.196 (0.623–2.296) |
| LASSBio-2069 (8) | 0.526 (0.272–1.019) | 1.650 (0.813–3.349) | 0.154 (0.062–0.385) | 0.808 (0.278–2.348) | 0.276 (0.156–0.490) | 0.095 (0.404–2.963) |
| LASSBio-2072 (9) | 5.848 (4.131–8.278) | 5.342 (3.556–8.027) | 0.450 (0.176–1.150) | 3.052 (1.547–6.019) | 1.273 (0.669–2.420) | 2.524 (1.697–3.754) |
| LASSBio-2070 (10) | 6.291 (5.159–7.673) | 0.903 (0.512–1.594) | 0.147 (0.056–0.383) | 2.125 (0.954–4.734) | 0.244 (0.135–0.443) | 0.935 (0.442–1.974) |
| LASSBio-2074 (11) | 5.688 (3.618–8.945) | 4.827 (2.988–7.799) | 0.593 (0.199–1.769) | 2.639 (1.160–4.003) | 1.405 (0.636–3.098) | 3.911 (1.774–6.21) |
| LASSBio-2122 (12) | 5.094 (3.643–7.122) | 6.367 (4.708–8.609) | 0.412 (0.159–1.068) | 1.052 (0.450–2.455) | 1.499 (0.928–2.421) | 3.8 (1.658–5.52) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbosa, G.; Gelves, L.G.V.; Costa, C.M.X.; Franco, L.S.; de Lima, J.A.L.; Aparecida-Silva, C.; Teixeira, J.D.; Mermelstein, C.d.S.; Barreiro, E.J.; Lima, L.M. Discovery of Putative Dual Inhibitor of Tubulin and EGFR by Phenotypic Approach on LASSBio-1586 Homologs. Pharmaceuticals 2022, 15, 913. https://doi.org/10.3390/ph15080913
Barbosa G, Gelves LGV, Costa CMX, Franco LS, de Lima JAL, Aparecida-Silva C, Teixeira JD, Mermelstein CdS, Barreiro EJ, Lima LM. Discovery of Putative Dual Inhibitor of Tubulin and EGFR by Phenotypic Approach on LASSBio-1586 Homologs. Pharmaceuticals. 2022; 15(8):913. https://doi.org/10.3390/ph15080913
Chicago/Turabian StyleBarbosa, Gisele, Luis Gabriel Valdivieso Gelves, Caroline Marques Xavier Costa, Lucas Silva Franco, João Alberto Lins de Lima, Cristiane Aparecida-Silva, John Douglas Teixeira, Claudia dos Santos Mermelstein, Eliezer J. Barreiro, and Lidia Moreira Lima. 2022. "Discovery of Putative Dual Inhibitor of Tubulin and EGFR by Phenotypic Approach on LASSBio-1586 Homologs" Pharmaceuticals 15, no. 8: 913. https://doi.org/10.3390/ph15080913
APA StyleBarbosa, G., Gelves, L. G. V., Costa, C. M. X., Franco, L. S., de Lima, J. A. L., Aparecida-Silva, C., Teixeira, J. D., Mermelstein, C. d. S., Barreiro, E. J., & Lima, L. M. (2022). Discovery of Putative Dual Inhibitor of Tubulin and EGFR by Phenotypic Approach on LASSBio-1586 Homologs. Pharmaceuticals, 15(8), 913. https://doi.org/10.3390/ph15080913