
Bromodomain and Extra-Terminal Inhibitor BMS-986158 Reverses Latent HIV-1 Infection In Vitro and Ex Vivo by Increasing CDK9 Phosphorylation and Recruitment


Abstract
:1. Introduction
2. Results
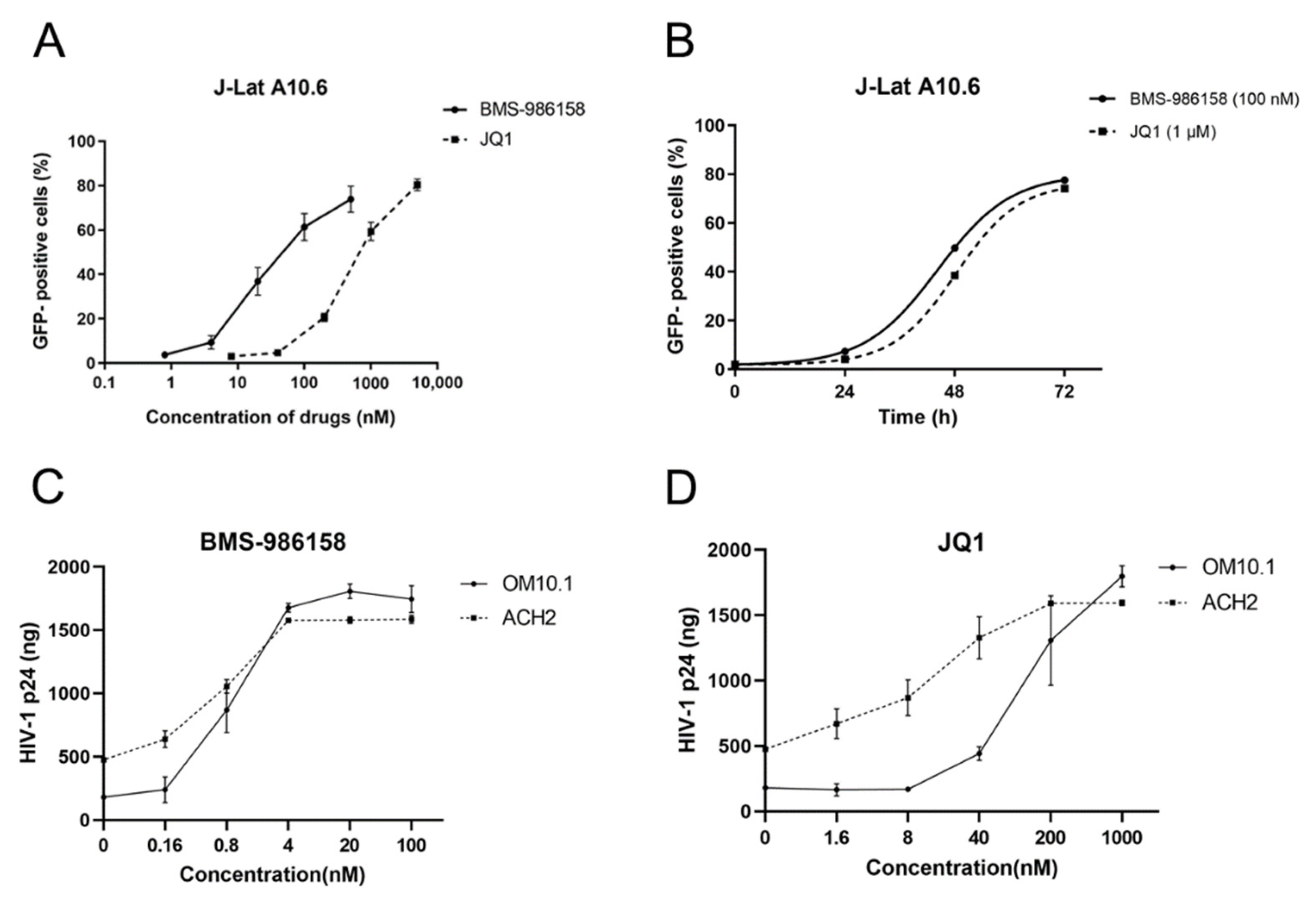
2.1. BMS-986158 Reverses HIV-1 Latency In Vitro and Ex Vivo
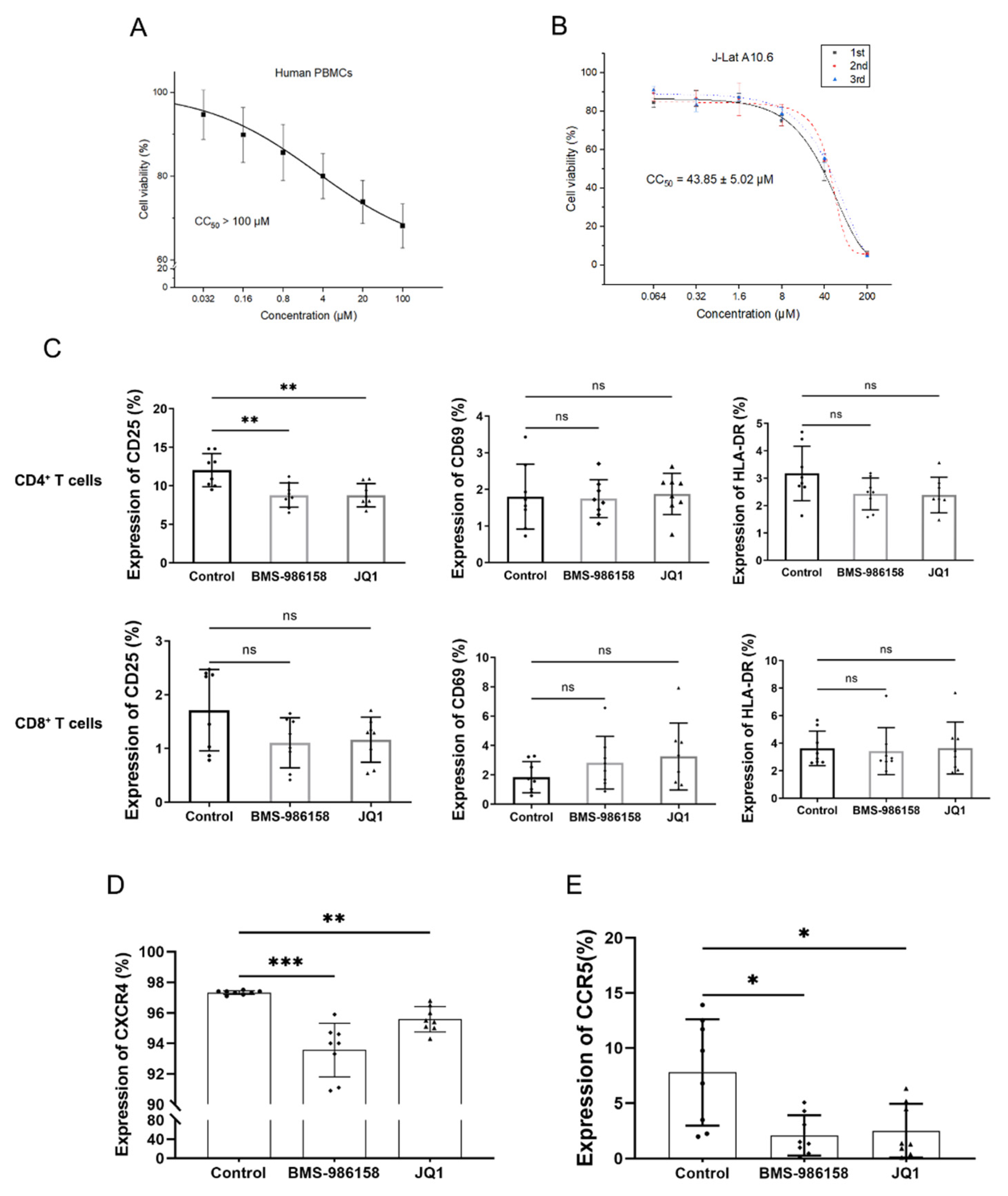
2.2. BMS-986158 Displays Low Toxicity in PBMCs and J-Lat Cells
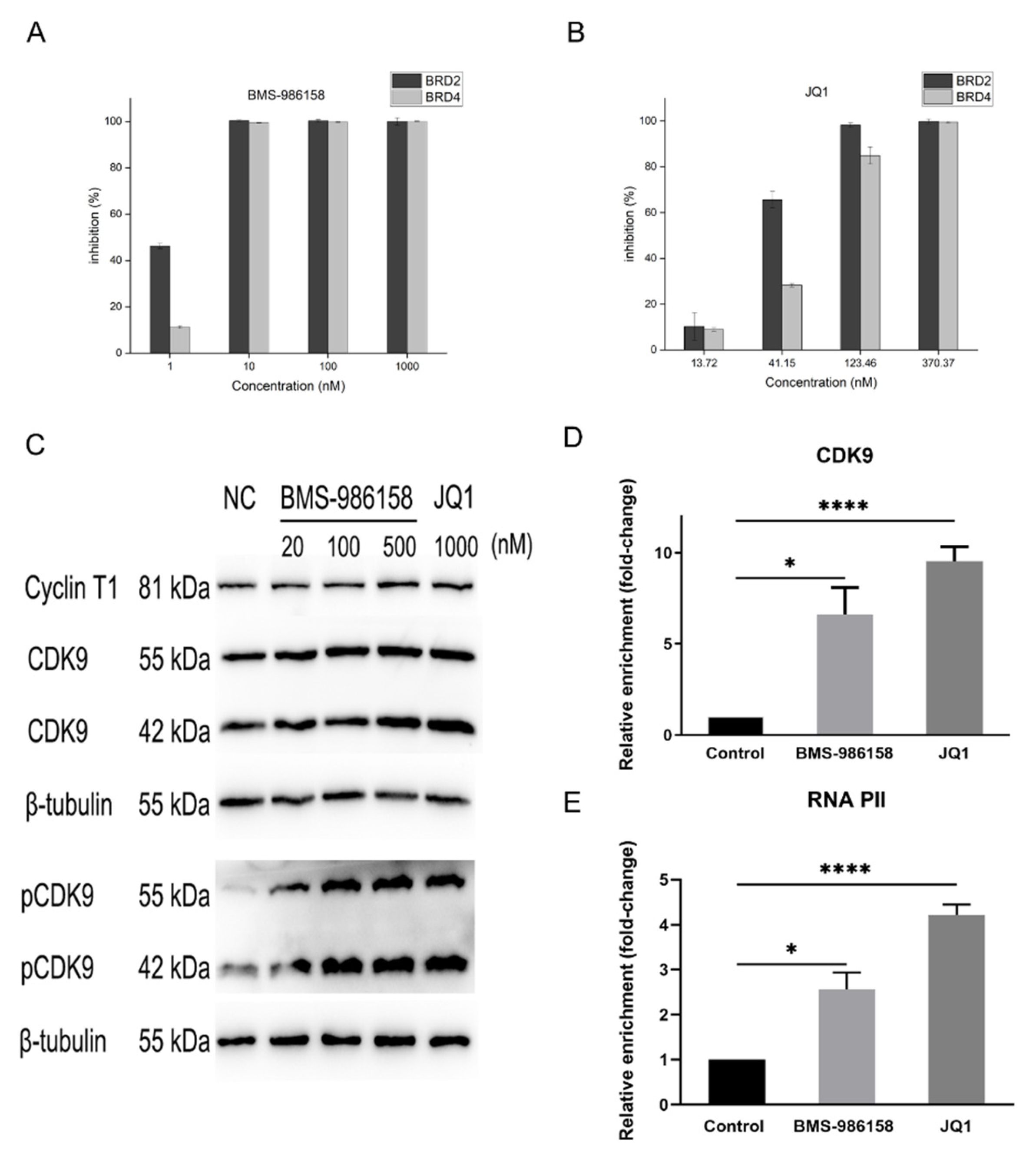
2.3. BMS-986158 Shows a Strong Inhibitory Effect on BRD4 Protein
2.4. BMS-986158 Promotes HIV Transcription via Activation of P-TEFb
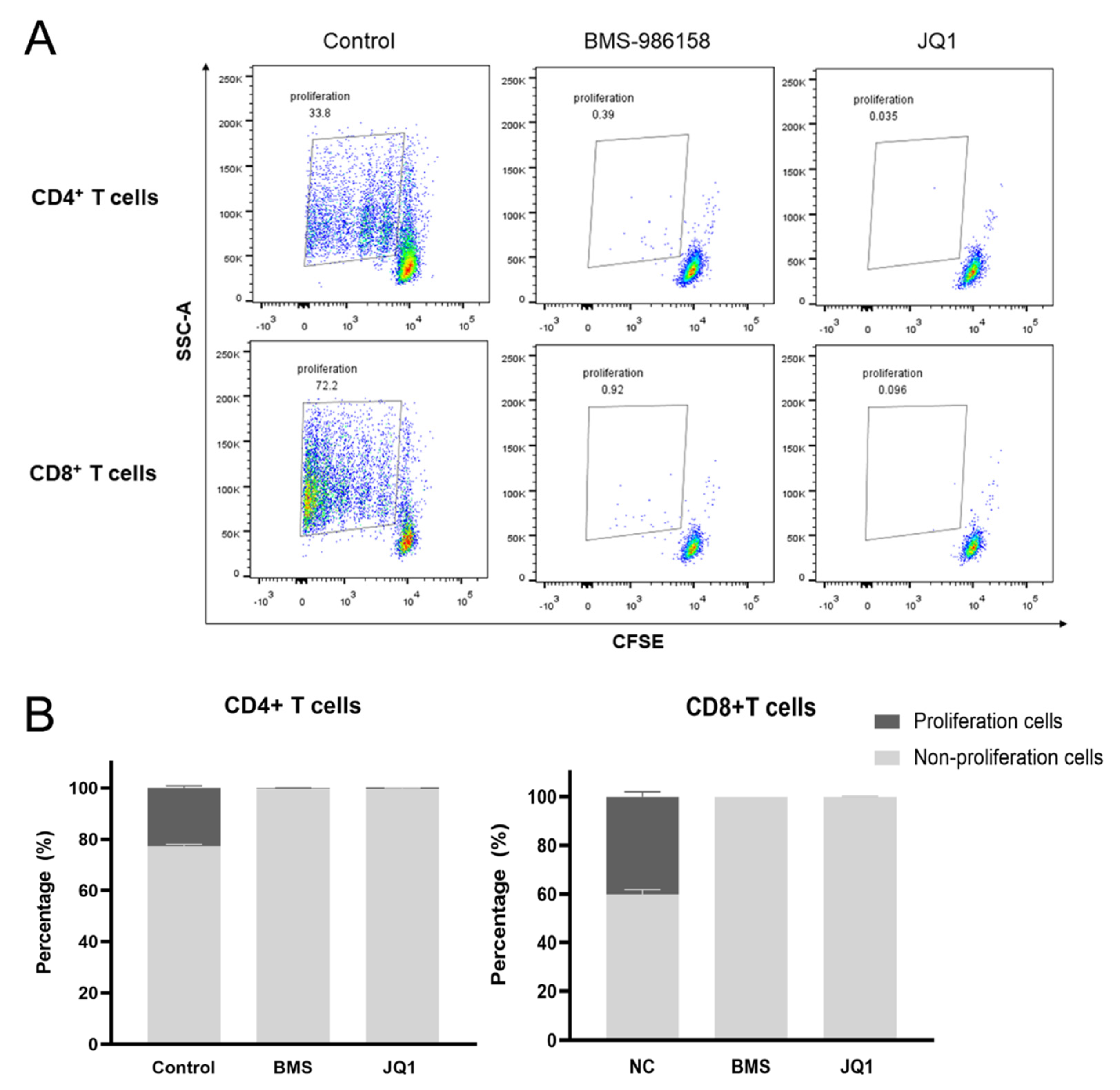
2.5. BMS-986158 Inhibits Cell Cycle Progression
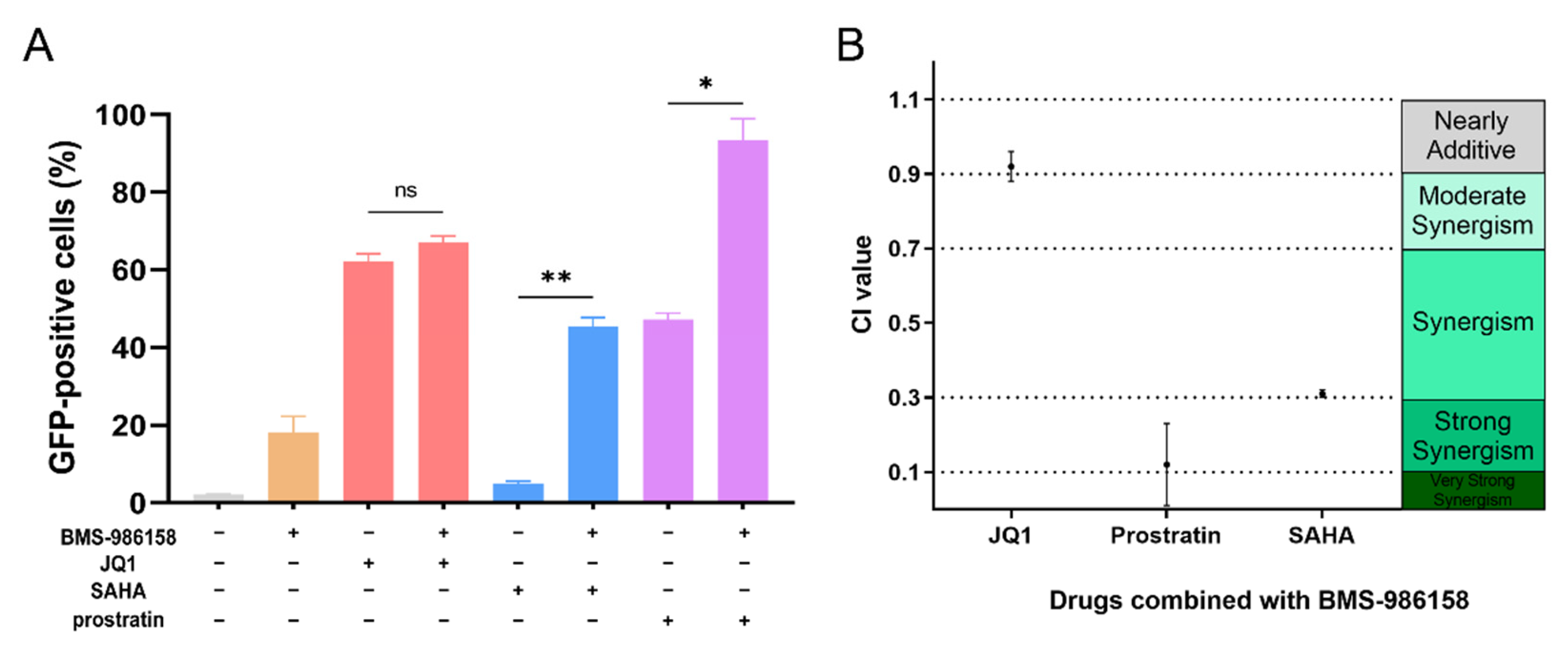
2.6. BMS-986158 Synergizes with SAHA and Prostratin to Reactivate Latent HIV-1
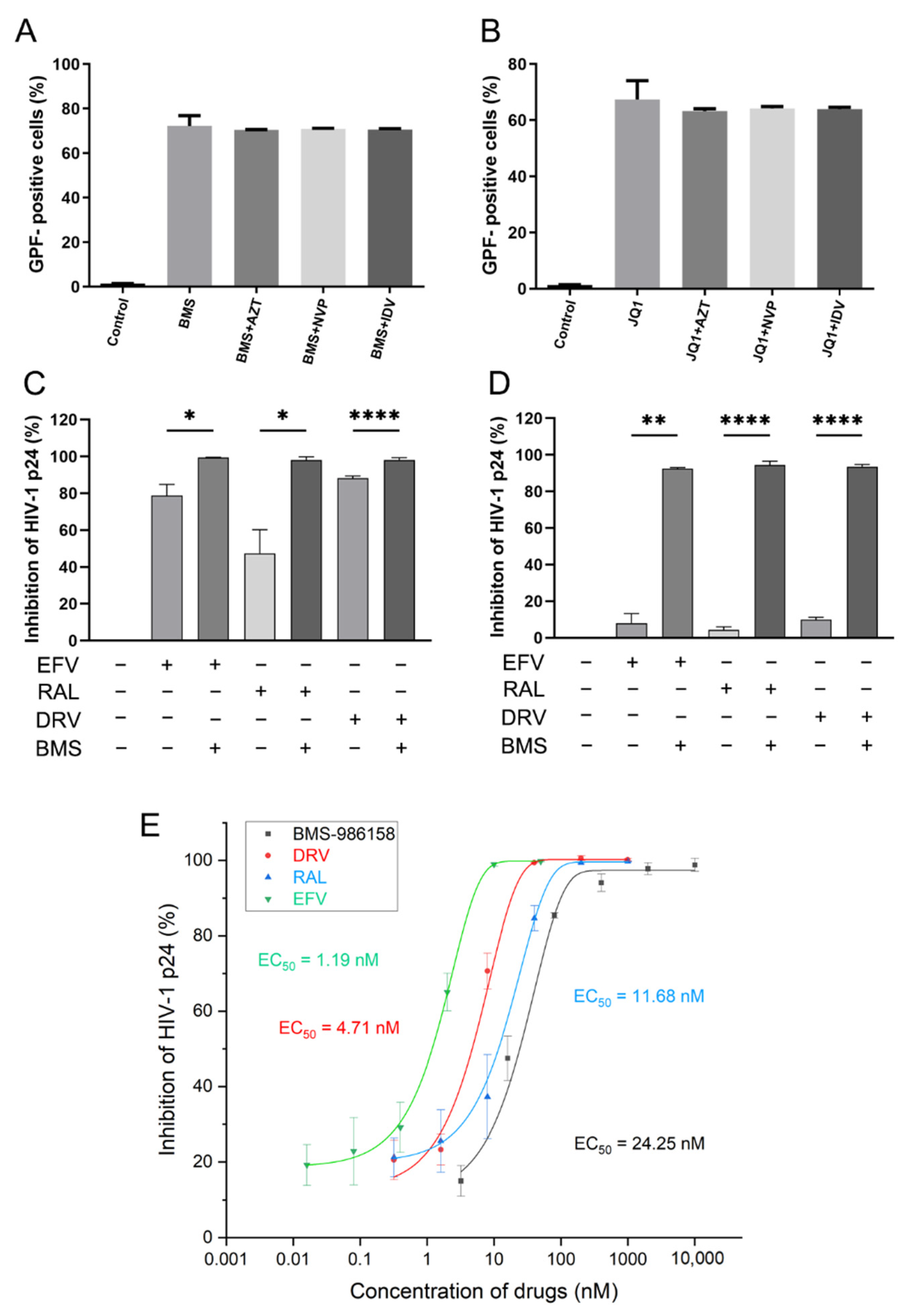
2.7. Effects of BMS-986158 on Latent HIV-1 Reactivation Are Not Impacted by Anti-HIV Drugs
2.8. BMS-986158 Enhances Antiviral Activity of Anti-HIV Drugs
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Compounds
4.3. Cell Lines and Culture
4.4. Measurement of HIV-1 Latency Reversal In Vitro
4.5. Acquisition of PBMCs
4.6. Measurement of HIV-1 Latency Reversal Ex Vivo
4.7. Cell Viability Assays
4.8. Cell Proliferation Assay
4.9. BRD2(1,2) and BRD4(1,2) Homogeneous Time-Resolved Fluorescence (HTRF) Assay
4.10. Antiviral Activity Assay
4.11. ELISA for HIV-1 p24
4.12. Western Blotting
4.13. Chromatin Immunoprecipitation (ChIP) Assay
4.14. Combination Reactivation Assay
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davey, R.T., Jr.; Bhat, N.; Yoder, C.; Chun, T.W.; Metcalf, J.A.; Dewar, R.; Natarajan, V.; Lempicki, R.A.; Adelsberger, J.W.; Miller, K.D.; et al. HIV-1 and T cell dynamics after interruption of highly active antiretroviral therapy (HAART) in patients with a history of sustained viral suppression. Proc. Natl. Acad. Sci. USA 1999, 96, 15109–15114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mata, R.C.; Viciana, P.; de Alarcon, A.; Lopez-Cortes, L.F.; Gomez-Vera, J.; Trastoy, M.; Cisneros, J.M. Discontinuation of antiretroviral therapy in patients with chronic HIV infection: Clinical, virologic, and immunologic consequences. AIDS Patient Care STDS 2005, 19, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Finzi, D.; Hermankova, M.; Pierson, T.; Carruth, L.M.; Buck, C.; Chaisson, R.E.; Quinn, T.C.; Chadwick, K.; Margolick, J.; Brookmeyer, R.; et al. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 1997, 278, 1295–1300. [Google Scholar] [CrossRef] [PubMed]
- Van Lint, C.; Bouchat, S.; Marcello, A. HIV-1 transcription and latency: An update. Retrovirology 2013, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Barton, K.M.; Burch, B.D.; Soriano-Sarabia, N.; Margolis, D.M. Prospects for treatment of latent HIV. Clin. Pharmacol. Ther. 2013, 93, 46–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deeks, S.G. HIV: Shock and kill. Nature 2012, 487, 439–440. [Google Scholar] [CrossRef]
- Bjerg Christensen, A.; Dige, A.; Vad-Nielsen, J.; Brinkmann, C.R.; Bendix, M.; Ostergaard, L.; Tolstrup, M.; Sogaard, O.S.; Rasmussen, T.A.; Randel Nyengaard, J.; et al. Administration of panobinostat is associated with increased IL-17A mRNA in the intestinal epithelium of HIV-1 patients. Mediat. Inflamm. 2015, 2015, 120605. [Google Scholar] [CrossRef]
- Delagreverie, H.M.; Delaugerre, C.; Lewin, S.R.; Deeks, S.G.; Li, J.Z. Ongoing clinical trials of human immunodeficiency virus latency-reversing and immunomodulatory agents. Open Forum Infect. Dis. 2016, 3, ofw189. [Google Scholar] [CrossRef]
- Elliott, J.H.; McMahon, J.H.; Chang, C.C.; Lee, S.A.; Hartogensis, W.; Bumpus, N.; Savic, R.; Roney, J.; Hoh, R.; Solomon, A.; et al. Short-term administration of disulfiram for reversal of latent HIV infection: A phase 2 dose-escalation study. Lancet HIV 2015, 2, e520–e529. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, C.; Serrano-Villar, S.; Madrid-Elena, N.; Perez-Elias, M.J.; Martin, M.E.; Barbas, C.; Ruiperez, J.; Munoz, E.; Munoz-Fernandez, M.A.; Castor, T.; et al. Bryostatin-1 for latent virus reactivation in HIV-infected patients on antiretroviral therapy. AIDS 2016, 30, 1385–1392. [Google Scholar] [CrossRef]
- Leth, S.; Schleimann, M.H.; Nissen, S.K.; Hojen, J.F.; Olesen, R.; Graversen, M.E.; Jorgensen, S.; Kjaer, A.S.; Denton, P.W.; Mork, A.; et al. Combined effect of Vacc-4x, recombinant human granulocyte macrophage colony-stimulating factor vaccination, and romidepsin on the HIV-1 reservoir (REDUC): A single-arm, phase 1B/2A trial. Lancet HIV 2016, 3, e463–e472. [Google Scholar] [CrossRef]
- Jacobs, J.L.; Halvas, E.K.; Tosiano, M.A.; Mellors, J.W. Persistent HIV-1 Viremia on Antiretroviral Therapy: Measurement and Mechanisms. Front. Microbiol. 2019, 10, 2383. [Google Scholar] [CrossRef] [PubMed]
- Alleboina, S.; Aljouda, N.; Miller, M.; Freeman, K.W. Therapeutically targeting oncogenic CRCs facilitates induced differentiation of NB by RA and the BET bromodomain inhibitor. Mol. Ther. Oncolytics 2021, 23, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Filippakopoulos, P.; Qi, J.; Picaud, S.; Shen, Y.; Smith, W.B.; Fedorov, O.; Morse, E.M.; Keates, T.; Hickman, T.T.; Felletar, I.; et al. Selective inhibition of BET bromodomains. Nature 2010, 468, 1067–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, B.; Zhu, Z.; Li, H.; Hong, Q.; Wang, C.; Ma, Y.; Zheng, W.; Jiang, F.; Zhang, Z.; Ran, T.; et al. Discovery of 1-(5-(1H-benzo[d]imidazole-2-yl)-2,4-dimethyl-1H-pyrrol-3-yl)ethan-1-one derivatives as novel and potent bromodomain and extra-terminal (BET) inhibitors with anticancer efficacy. Eur. J. Med. Chem. 2021, 227, 113953. [Google Scholar] [CrossRef] [PubMed]
- Conrad, R.J.; Fozouni, P.; Thomas, S.; Sy, H.; Zhang, Q.; Zhou, M.M.; Ott, M. The short isoform of BRD4 promotes HIV-1 latency by engaging repressive SWI/SNF chromatin-remodeling complexes. Mol. Cell 2017, 67, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Yik, J.H.; Chen, R.; He, N.; Jang, M.K.; Ozato, K.; Zhou, Q. Recruitment of P-TEFb for stimulation of transcriptional elongation by the bromodomain protein Brd4. Mol. Cell 2005, 19, 535–545. [Google Scholar] [CrossRef]
- Zhu, J.; Gaiha, G.D.; John, S.P.; Pertel, T.; Chin, C.R.; Gao, G.; Qu, H.; Walker, B.D.; Elledge, S.J.; Brass, A.L. Reactivation of latent HIV-1 by inhibition of BRD4. Cell Rep. 2012, 2, 807–816. [Google Scholar] [CrossRef] [Green Version]
- Boehm, D.; Calvanese, V.; Dar, R.D.; Xing, S.; Schroeder, S.; Martins, L.; Aull, K.; Li, P.C.; Planelles, V.; Bradner, J.E.; et al. BET bromodomain-targeting compounds reactivate HIV from latency via a Tat-independent mechanism. Cell Cycle 2013, 12, 452–462. [Google Scholar] [CrossRef]
- Banerjee, C.; Archin, N.; Michaels, D.; Belkina, A.C.; Denis, G.V.; Bradner, J.; Sebastiani, P.; Margolis, D.M.; Montano, M. BET bromodomain inhibition as a novel strategy for reactivation of HIV-1. J. Leukoc. Biol. 2012, 92, 1147–1154. [Google Scholar] [CrossRef]
- Lu, P.; Qu, X.; Shen, Y.; Jiang, Z.; Wang, P.; Zeng, H.; Ji, H.; Deng, J.; Yang, X.; Li, X.; et al. The BET inhibitor OTX015 reactivates latent HIV-1 through P-TEFb. Sci. Rep. 2016, 6, 24100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Lin, J.; Liang, T.Z.; Duan, H.; Tan, X.H.; Xi, B.M.; Li, L.; Liu, S.W. The BET bromodomain inhibitor apabetalone induces apoptosis of latent HIV-1 reservoir cells following viral reactivation. Acta. Pharmacol. Sin. 2019, 40, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Han, J.; Wang, Z.; Li, X.; Sun, Y.; Hu, Z. Safety and efficacy of bromodomain and extra-terminal inhibitors for the treatment of hematological malignancies and solid tumors: A systematic study of clinical trials. Front. Pharmacol. 2020, 11, 621093. [Google Scholar] [CrossRef] [PubMed]
- Von Schaper, E. Roche bets on bromodomains. Nat. Biotechnol. 2016, 34, 361–362. [Google Scholar] [CrossRef]
- Hezareh, M.; Moukil, M.A.; Szanto, I.; Pondarzewski, M.; Mouche, S.; Cherix, N.; Brown, S.J.; Carpentier, J.L.; Foti, M. Mechanisms of HIV receptor and co-receptor down-regulation by prostratin: Role of conventional and novel PKC isoforms. Antivir. Chem. Chemother. 2004, 15, 207–222. [Google Scholar] [CrossRef]
- Egloff, S. CDK9 keeps RNA polymerase II on track. Cell Mol. Life Sci. 2021, 78, 5543–5567. [Google Scholar] [CrossRef] [PubMed]
- Asamitsu, K.; Fujinaga, K.; Okamoto, T. HIV Tat/P-TEFb interaction: A potential target for novel anti-HIV therapies. Molecules 2018, 23, 933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamer, E.; Zhong, C.; Hajnik, R.; Soong, L.; Hu, H. Modulation of BRD4 in HIV epigenetic regulation: Implications for finding an HIV cure. Retrovirology 2021, 18, 3. [Google Scholar] [CrossRef]
- Pearson, A.D.; DuBois, S.G.; Buenger, V.; Kieran, M.; Stegmaier, K.; Bandopadhayay, P.; Bennett, K.; Bourdeaut, F.; Brown, P.A.; Chesler, L.; et al. Bromodomain and extra-terminal inhibitors-A consensus prioritisation after the Paediatric Strategy Forum for medicinal product development of epigenetic modifiers in children-ACCELERATE. Eur. J. Cancer 2021, 146, 115–124. [Google Scholar] [CrossRef]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Ruelas, D.S.; Greene, W.C. An integrated overview of HIV-1 latency. Cell 2013, 155, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichterfeld, M. Reactivation of latent HIV moves shock-and-kill treatments forward. Nature 2020, 578, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Archin, N.M.; Kirchherr, J.L.; Sung, J.A.; Clutton, G.; Sholtis, K.; Xu, Y.; Allard, B.; Stuelke, E.; Kashuba, A.D.; Kuruc, J.D.; et al. Interval dosing with the HDAC inhibitor vorinostat effectively reverses HIV latency. J. Clin. Investig. 2017, 127, 3126–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.A.; Elliott, J.H.; McMahon, J.; Hartogenesis, W.; Bumpus, N.N.; Lifson, J.D.; Gorelick, R.J.; Bacchetti, P.; Deeks, S.G.; Lewin, S.R.; et al. Population pharmacokinetics and pharmacodynamics of disulfiram on inducing latent HIV-1 transcription in a phase IIb trial. Clin. Pharmacol. Ther. 2019, 105, 692–702. [Google Scholar] [CrossRef]
- Sogaard, O.S.; Graversen, M.E.; Leth, S.; Olesen, R.; Brinkmann, C.R.; Nissen, S.K.; Kjaer, A.S.; Schleimann, M.H.; Denton, P.W.; Hey-Cunningham, W.J.; et al. The depsipeptide romidepsin reverses HIV-1 latency in vivo. PLoS Pathog. 2015, 11, e1005142. [Google Scholar] [CrossRef] [Green Version]
- Tsai, P.; Wu, G.; Baker, C.E.; Thayer, W.O.; Spagnuolo, R.A.; Sanchez, R.; Barrett, S.; Howell, B.; Margolis, D.; Hazuda, D.J.; et al. In vivo analysis of the effect of panobinostat on cell-associated HIV RNA and DNA levels and latent HIV infection. Retrovirology 2016, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Shen, Y.; Yang, H.; Wang, Y.; Jiang, Z.; Yang, X.; Zhong, Y.; Pan, H.; Xu, J.; Lu, H.; et al. BET inhibitors RVX-208 and PFI-1 reactivate HIV-1 from latency. Sci. Rep. 2017, 7, 16646. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek Michaels, K.; Wolschendorf, F.; Schiralli Lester, G.M.; Natarajan, M.; Kutsch, O.; Henderson, A.J. RNAP II processivity is a limiting step for HIV-1 transcription independent of orientation to and activity of endogenous neighboring promoters. Virology 2015, 486, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Marcello, A.; Zoppé, M.; Giacca, M. Multiple modes of transcriptional regulation by the HIV-1 Tat transactivator. IUBMB Life 2001, 51, 175–181. [Google Scholar] [CrossRef]
- Ott, M.; Geyer, M.; Zhou, Q. The control of HIV transcription: Keeping RNA polymerase II on track. Cell Host Microbe 2011, 10, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Karn, J. The molecular biology of HIV latency: Breaking and restoring the Tat-dependent transcriptional circuit. Curr. Opin. HIV AIDS 2011, 6, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, J.; Wu, Y.; Zhou, Q. The BET bromodomain inhibitor JQ1 activates HIV latency through antagonizing Brd4 inhibition of Tat-transactivation. Nucleic Acids Res. 2013, 41, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Korin, Y.D.; Brooks, D.G.; Brown, S.; Korotzer, A.; Zack, J.A. Effects of prostratin on T-cell activation and human immunodeficiency virus latency. J. Virol. 2002, 76, 8118–8123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.A.; Chen, L.F.; Kwon, H.; Fenard, D.; Bisgrove, D.; Verdin, E.; Greene, W.C. Prostratin antagonizes HIV latency by activating NF-kappaB. J. Biol. Chem. 2004, 279, 42008–42017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenfield, G.M. Fixed combination drug therapy. Drugs 1982, 23, 462–480. [Google Scholar]
- Liu, G.J.; Wang, J.P.; Xiao, J.C.; Zhao, Z.W.; Zheng, Y.T. Preparation and characterization of three monoclonal antibodies against HIV-1 p24 capsid protein. Cell Mol. Immunol. 2007, 4, 203–208. [Google Scholar]
- Chen, H.; Zhang, R.; Luo, R.H.; Yang, L.M.; Wang, R.R.; Hao, X.J.; Zheng, Y.T. Anti-HIV Activities and Mechanism of 12-O-Tricosanoylphorbol-20-acetate, a Novel Phorbol Ester from Ostodes katharinae. Molecules 2017, 22, 1498. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C.; Talalay, P. Analysis of combined drug effects–A new look at a very old problem. Trends Pharmacol. Sci. 1983, 4, 450–454. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donors | CA HIV-1 RNA (Copies/μg) | ||||
|---|---|---|---|---|---|
| Mock | BMS-986158 | JQ1 | |||
| 1 | ND | 51.31 | ▲ | 626.68 | ▲ |
| 2 | ND | 324.1 | ▲ | 34,767.24 | ▲ |
| 3 | ND | 408.54 | ▲ | 43.64 | ▲ |
| 4 | ND | 1698.66 | ▲ | 157.18 | ▲ |
| 5 | ND | 191,317.33 | ▲ | 30.95 | ▲ |
| 6 | 18.24 | 256.25 | ▲ | 47.04 | ▲ |
| 7 | 29.03 | 54,035.73 | ▲ | ND | ▼ |
| 8 | 50.96 | 4378.61 | ▲ | 6947.05 | ▲ |
| 9 | 151.11 | 1005.2 | ▲ | 1676.74 | ▲ |
| 10 | 154.8 | ND | ▼ | 81.24 | ▼ |
| 11 | 268.85 | 5906.85 | ▲ | 1148.21 | ▲ |
| 12 | 672.81 | 972.46 | ▲ | 4582.25 | ▲ |
| 13 | 1275.58 | 211.36 | ▼ | 688.28 | ▼ |
| 14 | 1357.75 | 272.8 | ▼ | 302.68 | ▼ |
| 15 | 1528.55 | 2317.63 | ▲ | 813.12 | ▼ |
| 16 | 2384.24 | 287.39 | ▼ | 939.99 | ▼ |
| 17 | 4359.75 | 3.42 | ▼ | 5408.31 | ▲ |
| 18 | 6711.11 | 9.23 | ▼ | 2372.51 | ▼ |
| 19 | 9190.65 | 8532.24 | ▼ | 2857.87 | ▼ |
| 20 | 63,438.38 | 4198.45 | ▼ | 13,307.23 | ▼ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.-S.; Tian, R.-R.; Ma, M.-D.; Luo, R.-H.; Yang, L.-M.; Peng, G.-H.; Zhang, M.; Dong, X.-Q.; Zheng, Y.-T. Bromodomain and Extra-Terminal Inhibitor BMS-986158 Reverses Latent HIV-1 Infection In Vitro and Ex Vivo by Increasing CDK9 Phosphorylation and Recruitment. Pharmaceuticals 2022, 15, 338. https://doi.org/10.3390/ph15030338
Huang X-S, Tian R-R, Ma M-D, Luo R-H, Yang L-M, Peng G-H, Zhang M, Dong X-Q, Zheng Y-T. Bromodomain and Extra-Terminal Inhibitor BMS-986158 Reverses Latent HIV-1 Infection In Vitro and Ex Vivo by Increasing CDK9 Phosphorylation and Recruitment. Pharmaceuticals. 2022; 15(3):338. https://doi.org/10.3390/ph15030338
Chicago/Turabian StyleHuang, Xu-Sheng, Ren-Rong Tian, Meng-Di Ma, Rong-Hua Luo, Liu-Meng Yang, Guang-Hui Peng, Mi Zhang, Xing-Qi Dong, and Yong-Tang Zheng. 2022. "Bromodomain and Extra-Terminal Inhibitor BMS-986158 Reverses Latent HIV-1 Infection In Vitro and Ex Vivo by Increasing CDK9 Phosphorylation and Recruitment" Pharmaceuticals 15, no. 3: 338. https://doi.org/10.3390/ph15030338
APA StyleHuang, X.-S., Tian, R.-R., Ma, M.-D., Luo, R.-H., Yang, L.-M., Peng, G.-H., Zhang, M., Dong, X.-Q., & Zheng, Y.-T. (2022). Bromodomain and Extra-Terminal Inhibitor BMS-986158 Reverses Latent HIV-1 Infection In Vitro and Ex Vivo by Increasing CDK9 Phosphorylation and Recruitment. Pharmaceuticals, 15(3), 338. https://doi.org/10.3390/ph15030338