
Trypanosoma cruzi Antigenic Proteins Shared with Acute Lymphoblastic Leukemia and Neuroblastoma

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
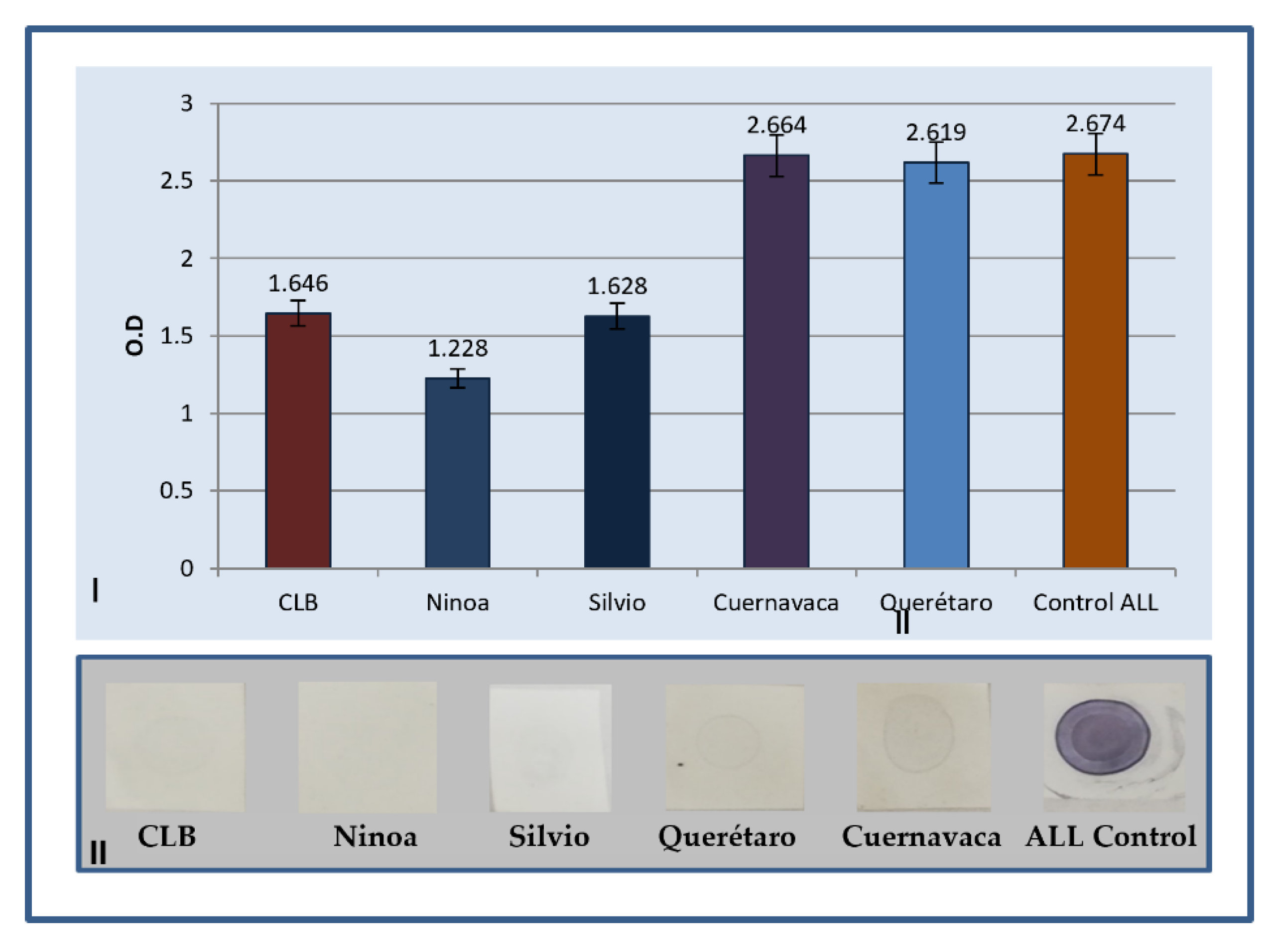
2.1. Determination of Antibodies by ELISA
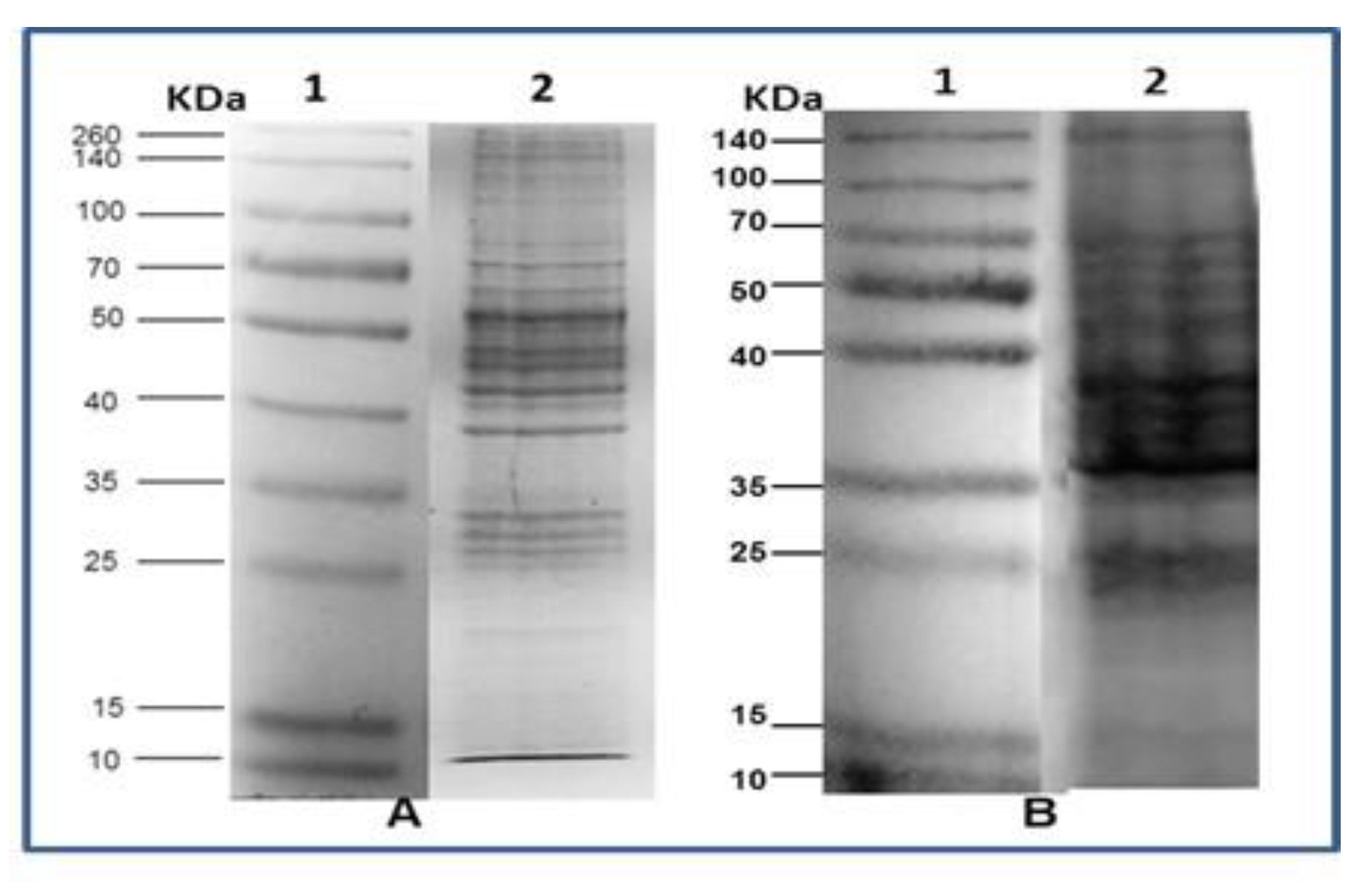
2.2. SDS Polyacrylamide Gel Electrophoresis
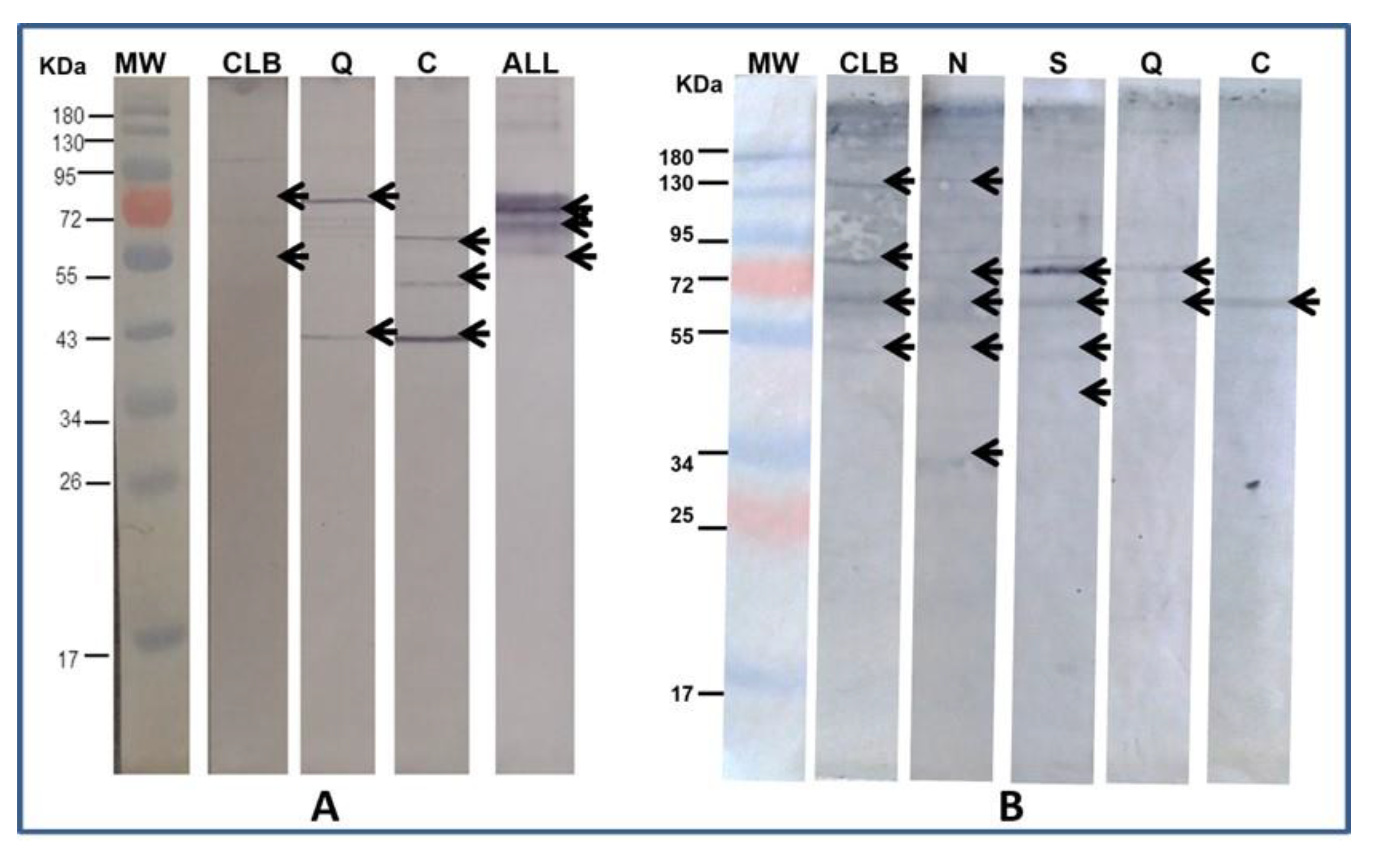
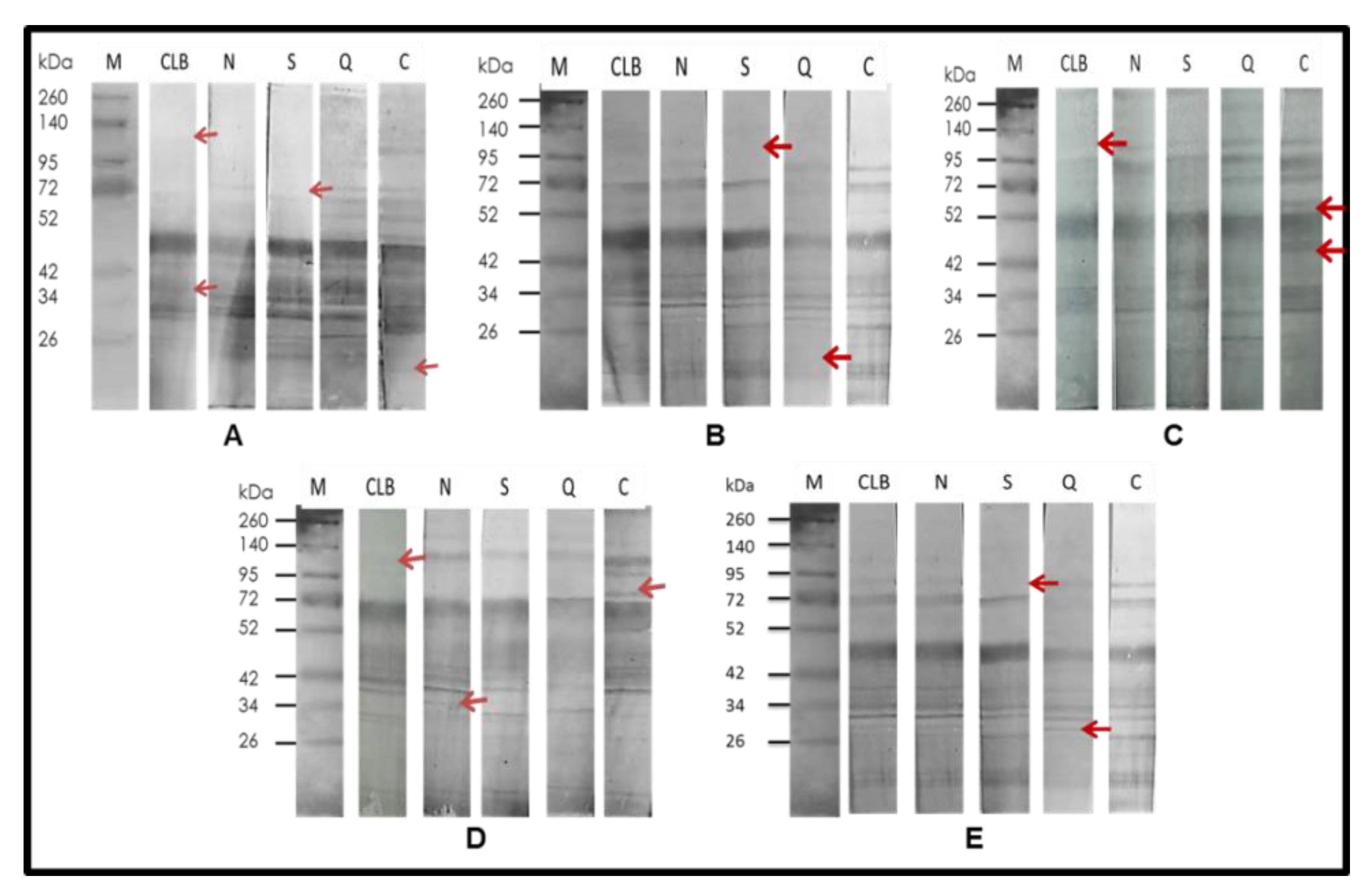
2.3. Immunodetection
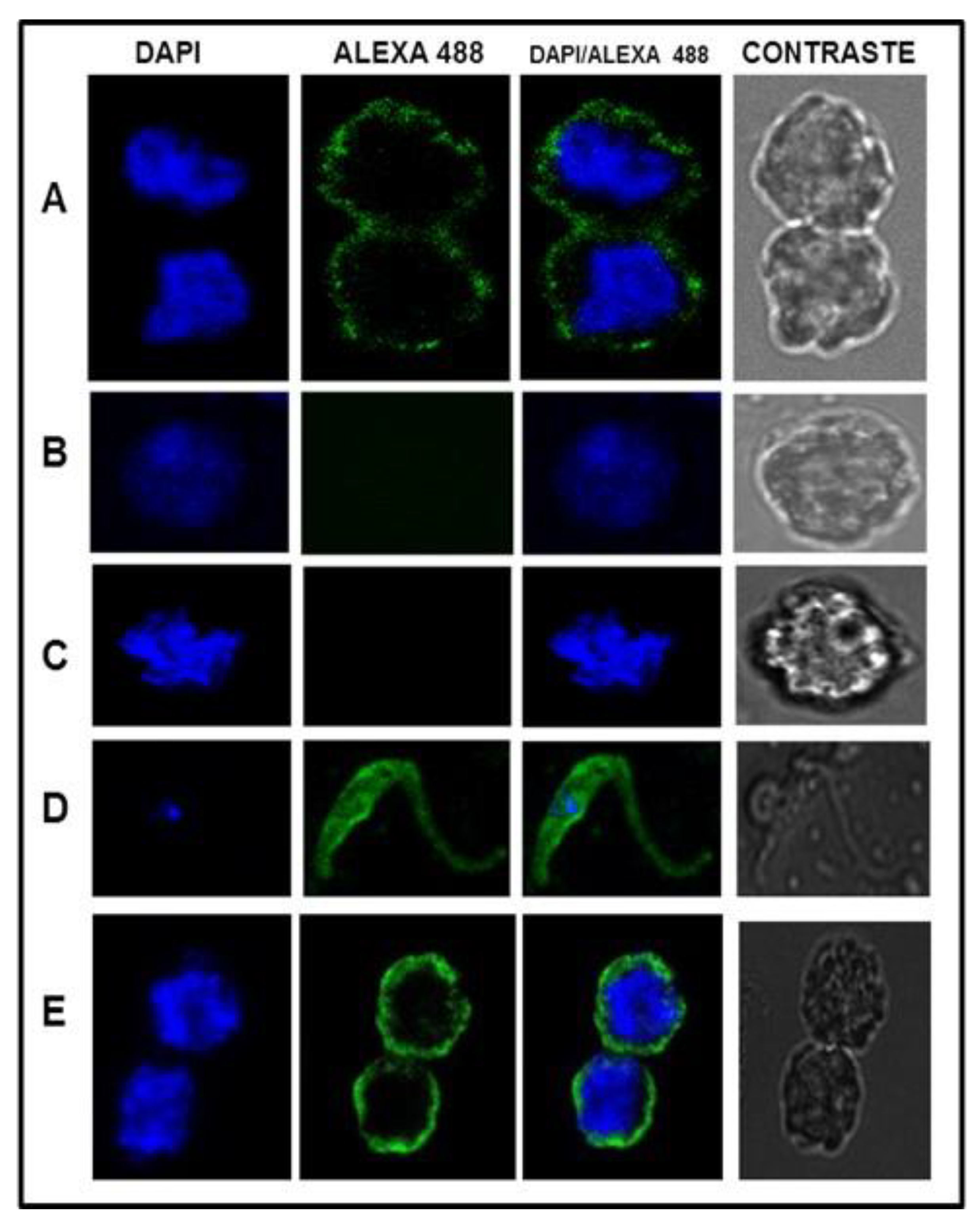
2.4. Confocal Microscopy
3. Materials and Methods
3.1. Production and Determination of Antibodies against T. cruzi in New Zealand Rabbits
3.1.1. Parasite Culture
3.1.2. Cellular Line
3.1.3. Antigen Preparation
3.1.4. Cell Extract of ALL
3.2. SDS Polyacrylamide Gel Electrophoresis
3.3. Immunodetection
3.4. Confocal Microscopy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sudhakar, A. History of Cancer, Ancient and Modern Treatment Methods. J. Cancer Sci. Ther. 2009, 1, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods, and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Secretaría de Salud. Comunicado en 08 de Agosto de 2017. Available online: www.gob.mx/salud (accessed on 8 August 2017).
- Paul, S.; Kantarjian, H.; Jabbour, E.J. Adult Acute Lymphoblastic Leukemia. Mayo. Clin. Proc. 2016, 91, 1645–1666. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, E.; O’Brien, S.; Konopleva, M.; Kantarjian, H. New insights into the pathophysiology and therapy of adult acute lymphoblastic leukemia. Cancer 2015, 121, 2517–2528. [Google Scholar] [CrossRef] [PubMed]
- Maris, J.M. Recent advances in neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef]
- Ishola, T.A.; Chung, D.H. Neuroblastoma. Surg. Oncol. 2007, 16, 149–156. [Google Scholar] [CrossRef]
- Berry, A.; Iriart, X.; Fillaux, J.; Magnaval, J.F. Urinary schistosomiasis and cancer. Bull. Soc. Pathol. Exot. 2017, 110, 68–75. [Google Scholar] [CrossRef]
- Mawson, A.R.; Majumdar, S. Malaria, Epstein-Barr virus infection and the pathogenesis of Burkitt’s lymphoma. Int. J. Cancer 2017, 141, 1849–1855. [Google Scholar] [CrossRef]
- Sutcliffe, S.; Alderete, J.F.; Till, C.; Goodman, P.J.; Hsing, A.W.; Zenilman, J.M.; De Marzo, A.M.; Platz, E.A. Trichomonosis and subsequent risk of prostate cancer in the Prostate Cancer Prevention Trial. Int. J. Cancer 2009, 124, 2082–2087. [Google Scholar] [CrossRef]
- Junqueira, C.; Guerrero, A.T.; Galvão-Filho, B.; Andrade, W.A.; Salgado, A.P.; Cunha, T.M.; Ropert, C.; Campos, M.A.; Penido, M.L.; Mendonça-Previato, L.; et al. Trypanosoma cruzi adjuvants potentiate T cell-mediated immunity induced by a NY-ESO-1 based antitumor vaccine. PLoS ONE 2012, 7, e36245. [Google Scholar] [CrossRef]
- Callejas, B.E.; Martínez-Saucedo, D.; Terrazas, L.I. Parasites as negative regulators of cancer. Biosci. Rep. 2018, 38, BSR20180935. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, Z.; Qin, L.; Li, Q.; Shi, X.; Zhao, S.; Chen, L.; Zhong, N.; Chen, X. Antitumor effect of malaria parasite infection in a murine Lewis lung cancer model through induction of innate and adaptive immunity. PLoS ONE 2011, 6, e24407. [Google Scholar] [CrossRef] [PubMed]
- Baird, J.R.; Byrne, K.T.; Lizotte, P.H.; Toraya-Brown, S.; Scarlett, U.K.; Alexander, M.P.; Sheen, M.R.; Fox, B.A.; Bzik, D.J.; Bosenberg, M.; et al. Immune-mediated regression of established B16F10 melanoma by intratumoral injection of attenuated Toxoplasma gondii protects against rechallenge. J. Immunol. 2013, 190, 469–478. [Google Scholar] [CrossRef]
- Noya, V.; Bay, S.; Festari, M.F.; García, E.P.; Rodriguez, E.; Chiale, C.; Garneau, C.; Baleux, F.; Astrada, S.; Bollati-Fogolín, M.; et al. Mucin-like peptides from Echinococcus granulosus induce antitumor activity. Int. J. Oncol. 2013, 43, 775–784. [Google Scholar] [CrossRef]
- Akgül, H.; Tez, M.; Unal, A.E.; Kekek, M.; Sayek, I.; Ozçelik, T. Echinococcus against cancer: Why not? Cancer 2003, 98, 1999–2000. [Google Scholar] [CrossRef] [PubMed]
- Van Knapen, F. Echinococcus granulosus infection and malignancy. Br. Med. J. 1980, 281, 195–196. [Google Scholar] [CrossRef] [PubMed]
- Berriel, E.; Russo, S.; Monin, L.; Festari, M.F.; Berois, N.; Fernández, G.; Freire, T.; Osinaga, E. Antitumor activity of human hydatid cyst fluid in a murine model of colon cancer. Sci. World J. 2013, 2013, 230176. [Google Scholar] [CrossRef]
- Abello-Cáceres, P.; Pizarro-Bauerle, J.; Rosas, C.; Maldonado, I.; Aguilar-Guzmán, L.; González, C.; Ramírez, G.; Ferreira, J.; Ferreira, A. Does native Trypanosoma cruzi calreticulin mediate growth inhibition of a mammary tumor during infection? BMC Cancer 2016, 16, 731. [Google Scholar] [CrossRef]
- van Tong, H.; Brindley, P.J.; Meyer, C.G.; Velavan, T.P. Parasite Infection, Carcinogenesis, and Human Malignancy. EBioMedicine 2017, 15, 12–23. [Google Scholar] [CrossRef]
- Peña Álvarez, J.; Teneb, J.; Maldonado, I.; Weinberger, K.; Rosas, C.; Lemus, D.; Valck, C.; Olivera-Nappa, Á.; Asenjo, J.A.; Ferreira, A. Structural bases that underline Trypanosoma cruzi calreticulin proinfective, antiangiogenic and antitumor properties. Immunobiology 2020, 225, 151863. [Google Scholar] [CrossRef]
- Kravtsov, E.G.; Karpenko, L.P.; Dalin, M.V. In vivo anticancer activity of lysates from Trypanosoma cruzi of different genetic groups. Bull. Exp. Biol. Med. 2006, 142, 470–473. [Google Scholar]
- Machicado, C.; Marcos, L.A. Carcinogenesis associated with parasites other than Schistosoma, Opisthorchis and Clonorchis: A systematic review. Int. J. Cancer 2016, 138, 2915–2921. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Xu, P.; Gallagher, S. Immunoblotting and Immunodetection. Curr. Protoc. Protein Sci. 2017, 88, 10.10.1–10.10.37. [Google Scholar] [CrossRef] [PubMed]
- Ubillos, L.; Freire, T.; Berriel, E.; Chiribao, M.L.; Chiale, C.; Festari, M.F.; Medeiros, A.; Mazal, D.; Rondán, M.; Bollati-Fogolín, M.; et al. Trypanosoma cruzi extracts elicit protective immune response against chemically induced colon and mammary cancers. Int. J. Cancer 2016, 138, 1719–1731. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, C.; Santos, L.I.; Galvão-Filho, B.; Teixeira, S.M.; Rodrigues, F.G.; DaRocha, W.D.; Chiari, E.; Jungbluth, A.A.; Ritter, G.; Gnjatic, S.; et al. Trypanosoma cruzi as an effective cancer antigen delivery vector. Proc. Natl. Acad. Sci. USA 2011, 108, 19695–19700. [Google Scholar] [CrossRef] [PubMed]
- Rondinelli, E.; Silva, R.; Carvalho, J.F.; de Almeida Soares, C.M.; de Carvalho, E.F.; de Castro, F.T. Trypanosoma cruzi: An in vitro cycle of cell differentiation in axenic culture. Exp. Parasitol. 1988, 66, 197–204. [Google Scholar] [CrossRef]
- Rellick, S.L.; Hu, G.; Piktel, D.; Martin, K.H.; Geldenhuys, W.J.; Nair, R.R.; Gibson, L.F. Co-culture model of B-cell acute lymphoblastic leukemia recapitulates a transcription signature of chemotherapy-refractory minimal residual disease. Sci. Rep. 2011, 11, 15840. [Google Scholar] [CrossRef]
- Luo, Z.; Zhao, Y.; Wang, Y.; Yang, X.; Zhao, B. Protective effect of theaflavins on neuron against 6-hydroxydopamine-induced apoptosis in SH-SY5Y cells. J. Clin. Biochem. Nutr. 2012, 50, 133–138. [Google Scholar] [CrossRef]
- Mejía, A.J.; Paláu, M.T.; Zúñiga, C.A. Protein profiles of Trypanosoma cruzi and Trypanosoma rangeli. Parasitol Lat. 2004, 59, 142–147. [Google Scholar] [CrossRef]
- Lee, C. Protein extraction from mammalian tissues. Methods Mol. Biol. 2007, 362, 385–389. [Google Scholar]
- Málaga-Machaca, E.S.; Romero-Ramirez, A.; Gilman, R.H.; Astupiña-Figueroa, S.; Angulo, N.; Florentini, A.; Lovon-Luque, C.J.; Gonza, R.A.; Del Carpio-Sanz, A.; Cabello, I.; et al. Polyclonal antibodies for the detection of Trypanosoma cruzi circulating antigens. PLoS Negl. Trop. Dis. 2017, 11, e0006069. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, S.; Chakavarti, D. Immunoblot analysis. J. Vis. Exp. 2008, 16, 759. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eligio García, L.; Crisóstomo Vázquez, M.d.P.; Maravelez Acosta, V.A.; Soria Guerrero, M.; Cortés Campos, A.; Jiménez Cardoso, E. Trypanosoma cruzi Antigenic Proteins Shared with Acute Lymphoblastic Leukemia and Neuroblastoma. Pharmaceuticals 2022, 15, 1421. https://doi.org/10.3390/ph15111421
Eligio García L, Crisóstomo Vázquez MdP, Maravelez Acosta VA, Soria Guerrero M, Cortés Campos A, Jiménez Cardoso E. Trypanosoma cruzi Antigenic Proteins Shared with Acute Lymphoblastic Leukemia and Neuroblastoma. Pharmaceuticals. 2022; 15(11):1421. https://doi.org/10.3390/ph15111421
Chicago/Turabian StyleEligio García, Leticia, María del Pilar Crisóstomo Vázquez, Víctor Alberto Maravelez Acosta, Mariana Soria Guerrero, Adrián Cortés Campos, and Enedina Jiménez Cardoso. 2022. "Trypanosoma cruzi Antigenic Proteins Shared with Acute Lymphoblastic Leukemia and Neuroblastoma" Pharmaceuticals 15, no. 11: 1421. https://doi.org/10.3390/ph15111421
APA StyleEligio García, L., Crisóstomo Vázquez, M. d. P., Maravelez Acosta, V. A., Soria Guerrero, M., Cortés Campos, A., & Jiménez Cardoso, E. (2022). Trypanosoma cruzi Antigenic Proteins Shared with Acute Lymphoblastic Leukemia and Neuroblastoma. Pharmaceuticals, 15(11), 1421. https://doi.org/10.3390/ph15111421