
An Algae-Made RBD from SARS-CoV-2 Is Immunogenic in Mice

 ,
,  and
and 
Abstract
1. Introduction
2. Results
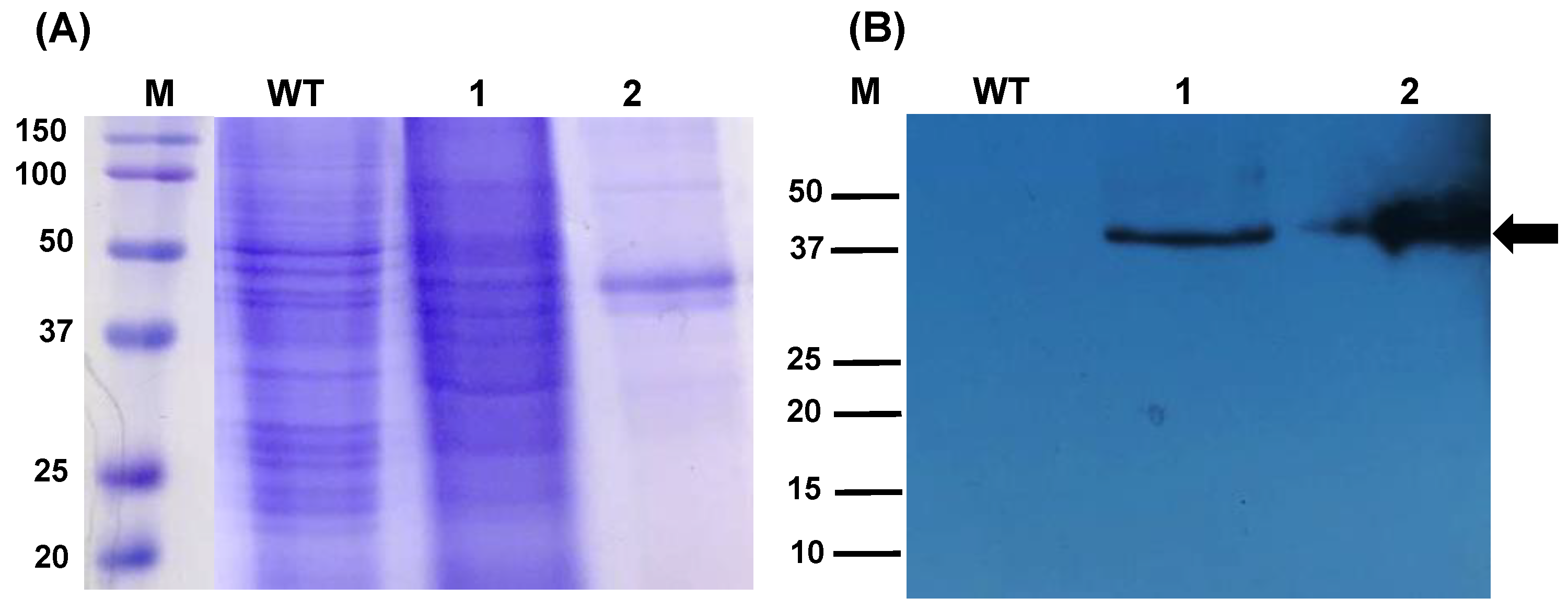
2.1. The Algae-Made RBD Is Highly Stable in the Freeze-Dried Biomass
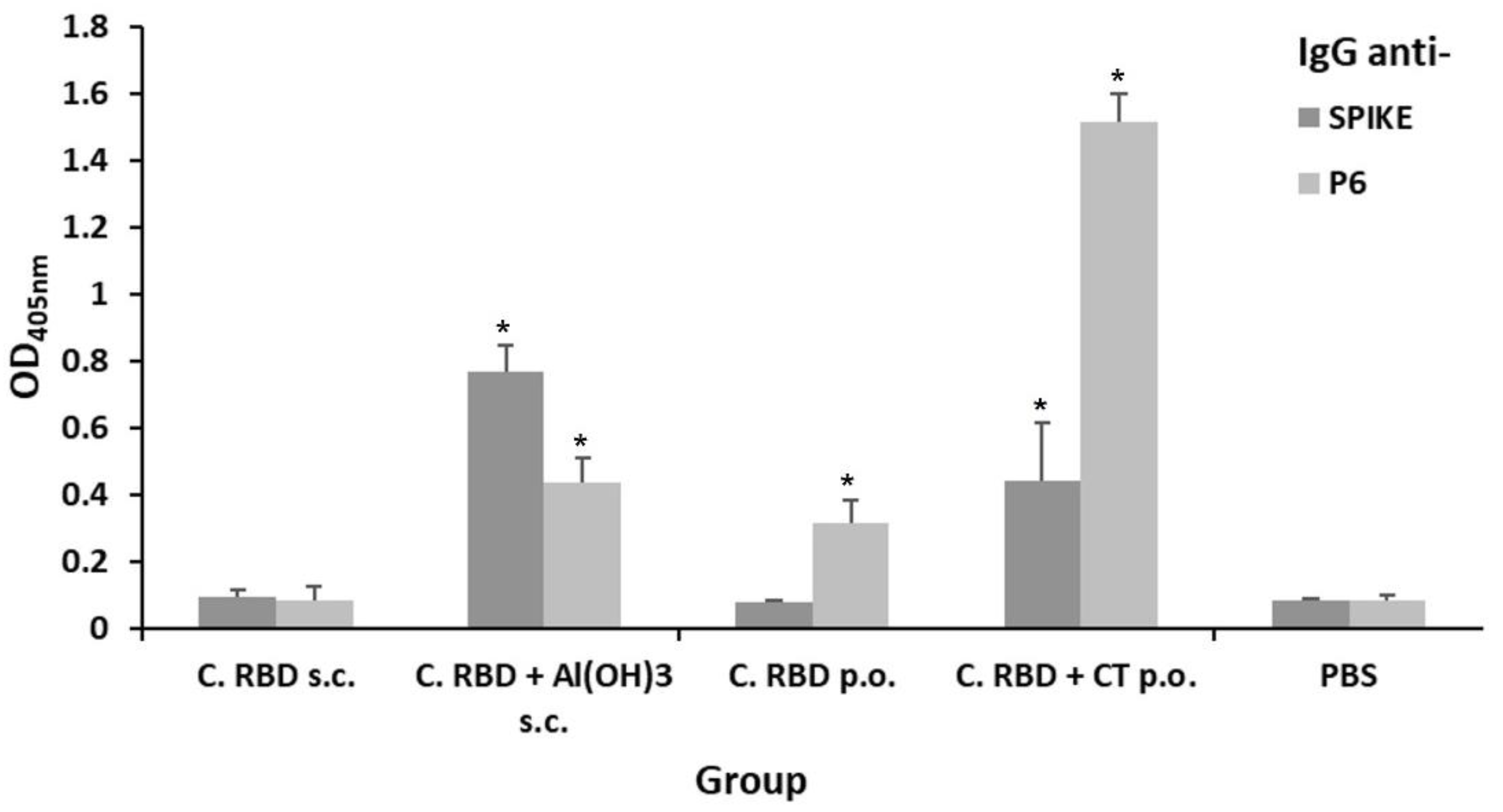
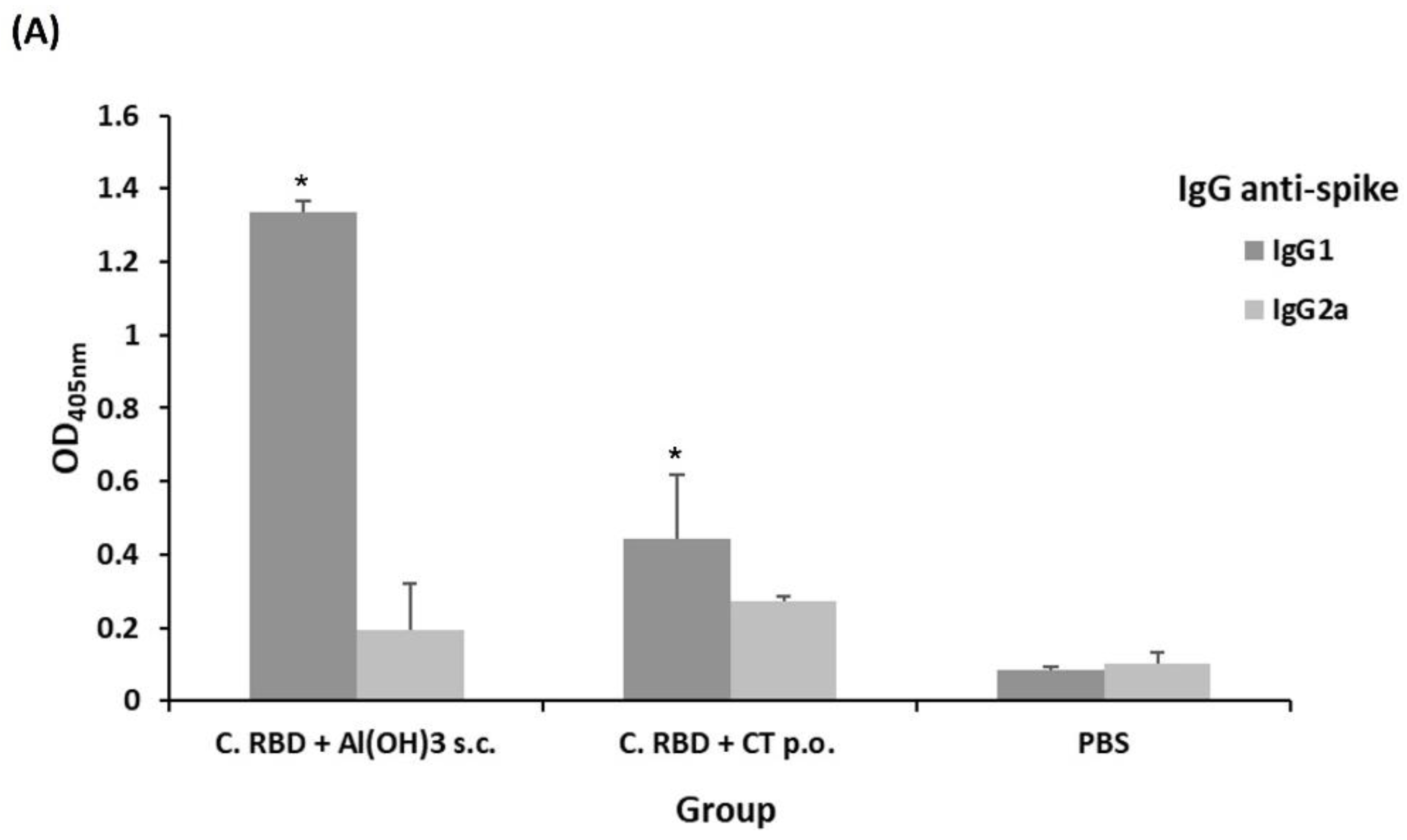
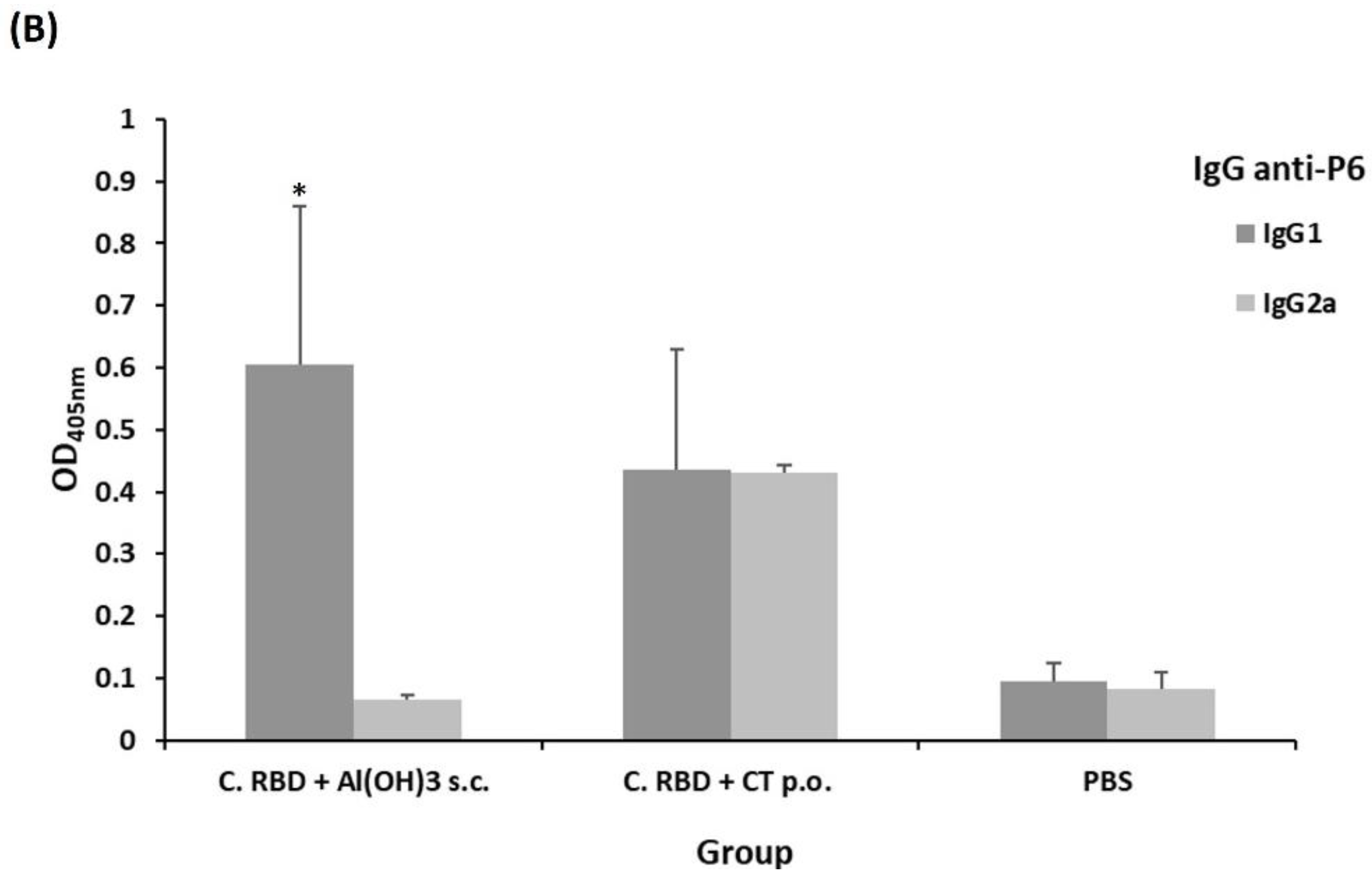
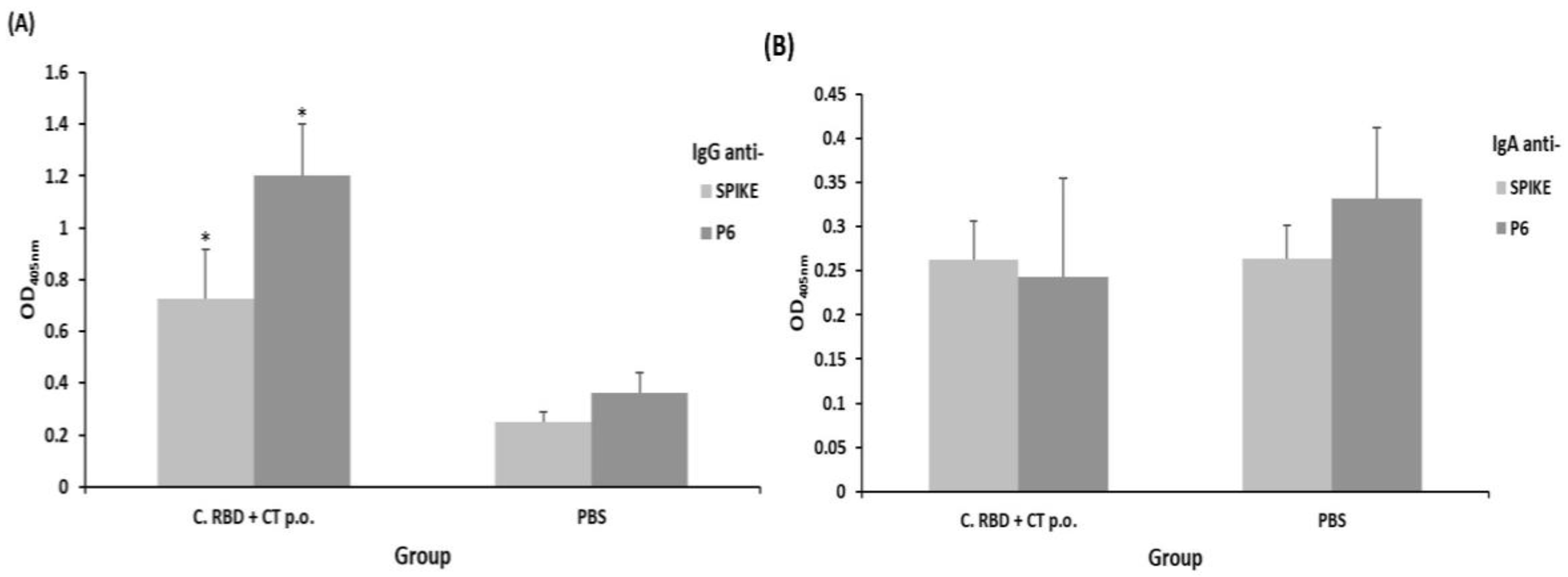
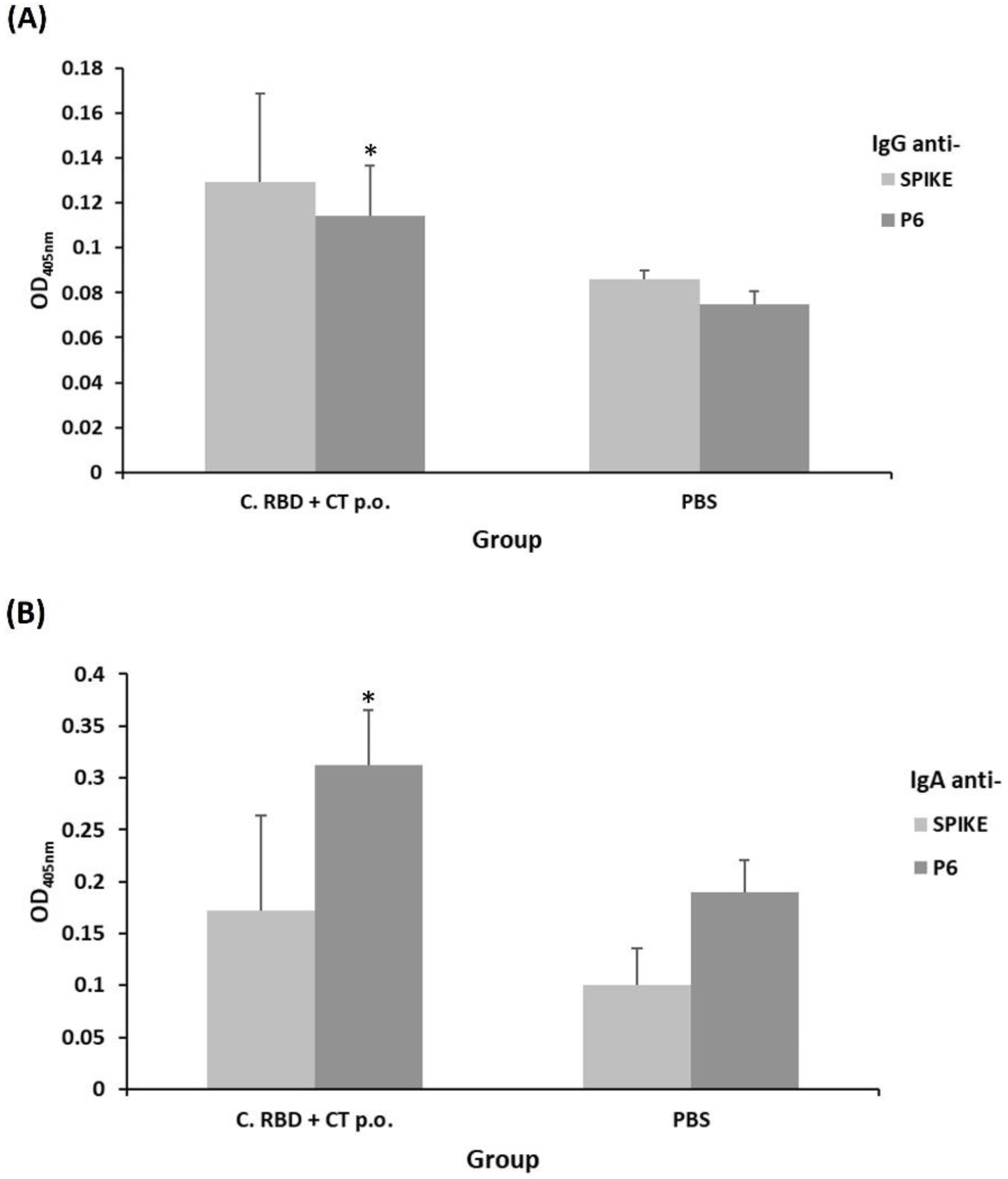
2.2. Algae-Made RBD Induced Both Systemic and Mucosal Responses
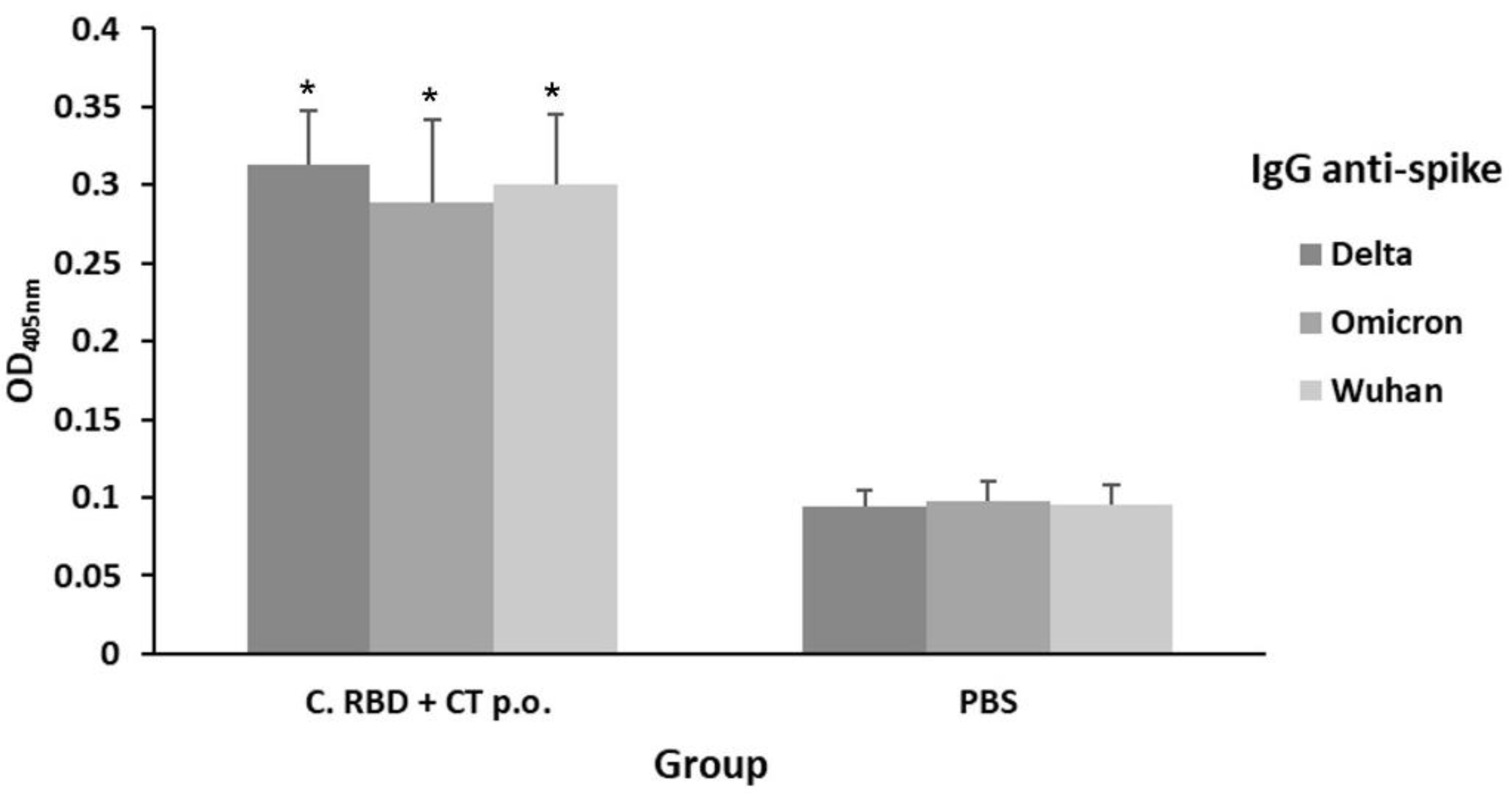
2.3. Algae-Made RBD Induced a Long-Lasting Response of Antibodies Recognizing Delta and Omicron Spike Variants
3. Discussion
4. Materials and Methods
4.1. Algae Transformation
4.2. Transient Transformation in Chlamydomonas reinhardtii
4.3. RBD Detection and Stability Assessment
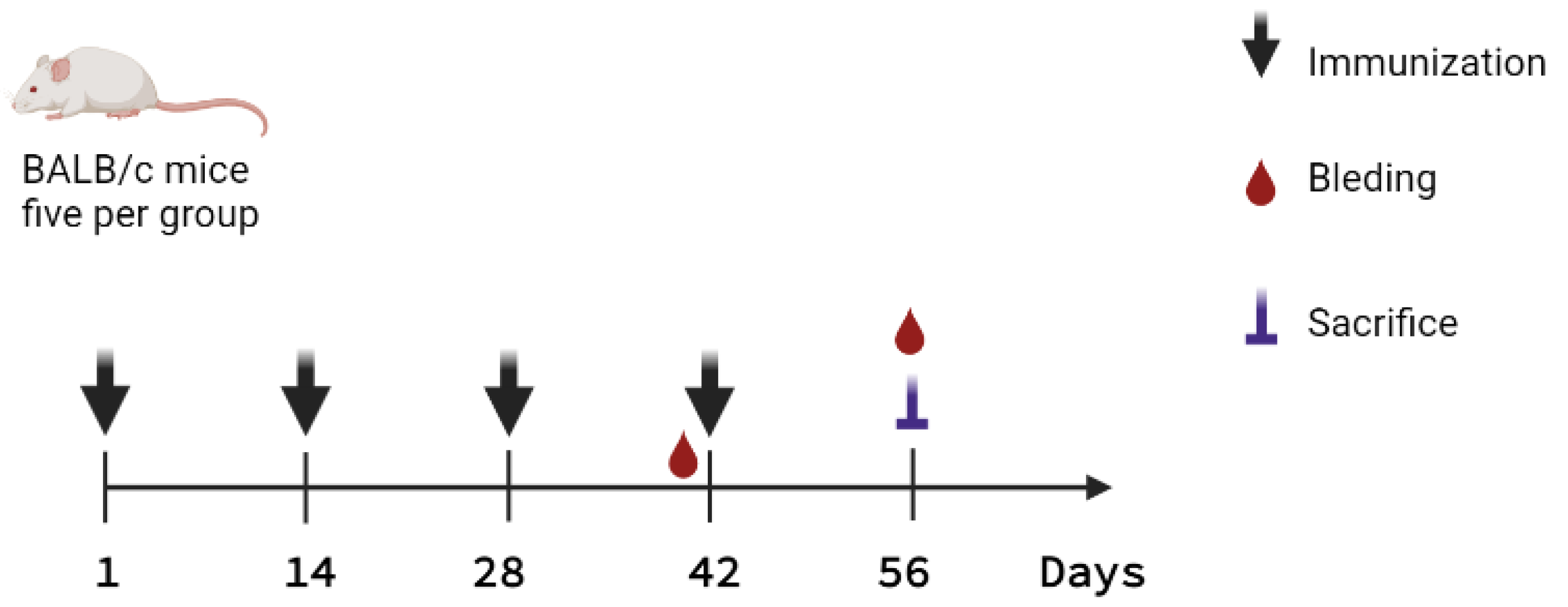
4.4. Immunization Schemes
4.5. Serum, Saliva, and Feces Sample Preparation
4.6. Antibody Measurement
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 10 August 2022).
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Flight, K.E.; Higham, S.L.; Wang, Z.; Pierce, B.F. Progress of the COVID-19 vaccine effort: Viruses, vaccines and variants versus efficacy, effectiveness and escape. Nat. Rev. Immunol. 2021, 21, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Sharif, N.; Alzahrani, K.J.; Ahmed, S.N.; Dey, S.K. Efficacy, immunogenicity and safety of COVID-19 vaccines: A systematic review and meta-analysis. Front. Immunol. 2021, 12, 714170. [Google Scholar] [CrossRef] [PubMed]
- Kashte, S.; Gulbake, A.; El-Amin Iii, S.F.; Gupta, A. COVID-19 vaccines: Rapid development, implications, challenges and future prospects. Hum. Cell 2021, 34, 711–733. [Google Scholar] [CrossRef]
- Koirala, A.; Joo, Y.J.; Khatami, A.; Chiu, C.; Britton, P.N. Vaccines for COVID-19: The current state of play. Paediatr. Respir. Rev. 2020, 35, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Ibarrondo, F.J.; Hofmann, C.; Fulcher, J.A.; Goodman-Meza, D.; Mu, W.; Hausner, M.A.; Ali, A.; Balamurugan, A.; Taus, E.; Elliott, J.; et al. Primary, recall, and decay kinetics of SARS-CoV-2 vaccine antibody responses. ACS Nano 2021, 15, 11180–11191. [Google Scholar] [CrossRef]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in mRNA vaccines for infectious diseases. Front. Immunol. 2019, 10, 594. [Google Scholar] [CrossRef]
- Soleimanpour, S.; Yaghoubi, A. COVID-19 vaccine: Where are we now and where should we go? Expert Rev. Vaccines 2021, 20, 23–44. [Google Scholar] [CrossRef]
- Alturaiki, W. Considerations for novel COVID-19 mucosal vaccine development. Vaccines 2022, 10, 1173. [Google Scholar] [CrossRef]
- Karczmarzyk, K.; Kęsik-Brodacka, M. Attacking the intruder at the gate: Prospects of mucosal anti SARS-CoV-2 vaccines. Pathogens 2022, 11, 117. [Google Scholar] [CrossRef]
- van Doremalen, N.; Purushotham, J.N.; Schulz, J.E.; Holbrook, M.G.; Bushmaker, T.; Carmody, A.; Port, J.R.; Yinda, C.K.; Okumura, A.; Saturday, G.; et al. Intranasal ChAdOx1 nCoV-19/AZD1222 vaccination reduces viral shedding after SARS-CoV-2 D614G challenge in preclinical models. Sci. Transl. Med. 2021, 13, eabh0755. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Li, K.; Rowe, D.K.; Diaz, M.; Griffin, E.F.; Beavis, A.C.; Johnson, S.K.; Padykula, I.; Jones, C.A.; Briggs, K. Protection of K18-hACE2 mice and ferrets against SARS-CoV-2 challenge by a single-dose mucosal immunization with a parainfluenza virus 5-based COVID-19 vaccine. Sci. Adv. 2021, 7, eabi5246. [Google Scholar] [CrossRef]
- Park, J.G.; Oladunni, F.S.; Rohaim, M.A.; Whittingham-Dowd, J.; Tollitt, J.; Hodges, M.; Fathallah, N.; Assas, M.B.; Alhazmi, W.; Almilaibary, A.; et al. Immunogenicity and protective efficacy of an intranasal live-attenuated vaccine against SARS-CoV-2. iScience 2021, 24, 102941. [Google Scholar] [CrossRef] [PubMed]
- Kumar, U.S.; Afjei, R.; Ferrara, K.; Massoud, T.F.; Paulmurugan, R. Gold-Nanostar-Chitosan-Mediated Delivery of SARS-CoV-2 DNA vaccine for respiratory mucosal immunization: Development and Proof-of-Principle. ACS Nano 2021, 15, 17582–17601. [Google Scholar] [CrossRef]
- Jearanaiwitayakul, T.; Seesen, M.; Chawengkirttikul, R.; Limthongkul, J.; Apichirapokey, S.; Sapsutthipas, S.; Phumiamorn, S.; Sunintaboon, P.; Ubol, S. Intranasal administration of RBD nanoparticles confers induction of mucosal and systemic immunity against SARS-CoV-2. Vaccines 2021, 9, 768. [Google Scholar] [CrossRef]
- Sung, J.C.; Liu, Y.; Wu, K.C.; Choi, M.C.; Ma, C.H.; Lin, J.; He, E.; Leung, D.Y.; Sze, E.T.; Hamied, Y.K.; et al. Expression of SARS-CoV-2 Spike protein receptor binding domain on recombinant B. subtilis on spore surface: A potential COVID-19 oral vaccine candidate. Vaccines 2021, 10, 2. [Google Scholar] [CrossRef]
- Peng, K.W.; Carey, T.; Lech, P.; Vandergaast, R.; Muñoz-Alía, M.Á.; Packiriswamy, N.; Gnanadurai, C.; Krotova, K.; Tesfay, M.; Ziegler, C.; et al. Boosting of SARS-CoV-2 immunity in nonhuman primates using an oral rhabdoviral vaccine. Vaccine 2022, 40, 2342–2351. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Martinez, C.I.; Tedjakusuma, S.N.; Peinovich, N.; Dora, E.G.; Birch, S.M.; Kajon, A.E.; Werts, A.D.; Tucker, S.N. Oral vaccination protects against severe acute respiratory syndrome coronavirus 2 in a Syrian hamster challenge model. J. Infect. Dis. 2022, 225, 34–41. [Google Scholar] [CrossRef]
- Sui, Y.; Li, J.; Zhang, R.; Prabhu, S.K.; Andersen, H.; Venzon, D.; Cook, A.; Brown, R.; Teow, E.; Velasco, J.; et al. Protection against SARS-CoV-2 infection by a mucosal vaccine in rhesus macaques. JCI Insight 2021, 6, e148494. [Google Scholar] [CrossRef]
- Pitcovski, J.; Gruzdev, N.; Abzach, A.; Katz, C.; Ben-Adiva, R.; Brand-Shwartz, M.; Yadid, I.; Ratzon-Ashkenazi, E.; Emquies, K.; Israeli, H.; et al. Oral subunit SARS-CoV-2 vaccine induces systemic neutralizing IgG, IgA and cellular immune responses and can boost neutralizing antibody responses primed by an injected vaccine. Vaccine 2022, 40, 1098–1107. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; García-Silva, I.; González-Ortega, O.; Sandoval-Vargas, J.M.; Malla, A.; Vimolmangkang, S. The potential of algal biotechnology to produce antiviral compounds and biopharmaceuticals. Molecules 2020, 25, 4049. [Google Scholar] [CrossRef] [PubMed]
- Garduño-González, K.A.; Peña-Benavides, S.A.; Araújo, R.G.; Castillo-Zacarías, C.; Melchor-Martínez, E.M.; Oyervides-Muñoz, M.A.; Sosa-Hernández, J.E.; Purton, S.; Iqbal, H.M.N.; Parra-Saldívar, R. Current challenges for modern vaccines and perspectives for novel treatment alternatives. J. Drug. Deliv. Sci. Technol. 2022, 70, 103222. [Google Scholar] [CrossRef]
- Specht, E.A.; Mayfield, S.P. Algae-based oral recombinant vaccines. Front. Microbiol. 2014, 5, 60. [Google Scholar] [CrossRef]
- Bañuelos-Hernández, B.; Monreal-Escalante, E.; González-Ortega, O.; Angulo, C.; Rosales-Mendoza, S. Algevir: An expression system for microalgae based on viral vectors. Front. Microbiol. 2017, 8, 1100. [Google Scholar] [CrossRef] [PubMed]
- Bolaños-Martínez, O.C.; Mahendran, G.; Rosales-Mendoza, S.; Vimolmangkang, S. Current Status and Perspective on the Use of Viral-Based Vectors in Eukaryotic Microalgae. Mar. Drugs 2022, 20, 434. [Google Scholar] [CrossRef] [PubMed]
- Diamos, A.G.; Hunter, J.G.L.; Pardhe, M.D.; Rosenthal, S.H.; Sun, H.; Foster, B.C.; DiPalma, M.P.; Chen, Q.; Mason, H.S. High level production of monoclonal antibodies using an optimized plant expression system. Front. Bioeng. Biotechnol. 2020, 7, 472. [Google Scholar] [CrossRef]
- Malla, A.; Rosales-Mendoza, S.; Phoolcharoen, W.; Vimolmangkang, S. Efficient transient expression of recombinant proteins using DNA viral vectors in freshwater microalgal species. Front. Plant Sci. 2021, 12, 650820. [Google Scholar] [CrossRef]
- Beltrán-López, J.I.; Romero-Maldonado, A.; Monreal-Escalante, E.; Bañuelos-Hernández, B.; Paz-Maldonado, L.M.; Rosales-Mendoza, S. Chlamydomonas reinhardtii chloroplasts express an orally immunogenic protein targeting the p210 epitope implicated in atherosclerosis immunotherapies. Plant Cell Rep. 2016, 35, 1133–1141. [Google Scholar] [CrossRef]
- Pniewski, T.; Kapusta, J.; Bociąg, P.; Wojciechowicz, J.; Kostrzak, A.; Gdula, M.; Fedorowicz-Strońska, O.; Wójcik, P.; Otta, H.; Samardakiewicz, S.; et al. Low-dose oral immunization with lyophilized tissue of herbicide-resistant lettuce expressing hepatitis B surface antigen for prototype plant-derived vaccine tablet formulation. J. Appl. Genet. 2011, 52, 125–136. [Google Scholar] [CrossRef]
- Sui, Y.; Bekele, Y.; Berzofsky, J.A. Potential SARS-CoV-2 immune correlates of protection in infection and vaccine immunization. Pathogens 2021, 10, 138. [Google Scholar] [CrossRef]
- Norton, E.B.; Lawson, L.B.; Freytag, L.C.; Clements, J.D. Characterization of a mutant Escherichia coli heat-labile toxin, LT(R192G/L211A), as a safe and effective oral adjuvant. Clin. Vaccine Immunol. 2011, 18, 546–551. [Google Scholar] [CrossRef]
- Fausther-Bovendo, H.; Kobinger, G. Plant-made vaccines and therapeutics. Science 2021, 373, 740–741. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Arunachalam, P.S.; Andreani, G.; Golden, N.; Fontenot, J.; Aye, P.P.; Röltgen, K.; Lehmicke, G.; Gobeil, P.; Dubé, C.; et al. Safety, immunogenicity, and protection provided by unadjuvanted and adjuvanted formulations of a recombinant plant-derived virus-like particle vaccine candidate for COVID-19 in nonhuman primates. Cell Mol. Immunol. 2022, 19, 222–233. [Google Scholar] [CrossRef]
- Gonzalez-Cruz, P.; Gill, H.S. Demystifying particle-based oral vaccines. Expert Opin. Drug Deliv. 2021, 18, 1455–1472. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Kulis, M.; Herzog, R.W. Plant cell-made protein antigens for induction of Oral tolerance. Biotechnol. Adv. 2019, 37, 107413. [Google Scholar] [CrossRef] [PubMed]
- Berndt, A.J.; Smalley, T.N.; Ren, B.; Simkovsky, R.; Badary, A.; Sproles, A.E.; Fields, F.J.; Torres-Tiji, Y.; Heredia, V.; Mayfield, S.P. Recombinant production of a functional SARS-CoV-2 spike receptor binding domain in the green algae Chlamydomonas reinhardtii. PLoS ONE 2021, 16, e0257089. [Google Scholar] [CrossRef]
- Larrea-Alvarez, M.; Young, R.; Purton, S. A simple technology for generating marker-free chloroplast transformants of the green alga Chlamydomonas reinhardtii. Methods Mol. Biol. 2021, 2317, 293–304. [Google Scholar] [CrossRef]
- Rattanapisit, K.; Shanmugaraj, B.; Manopwisedjaroen, S.; Purwono, P.B.; Siriwattananon, K.; Khorattanakulchai, N.; Hanittinan, O.; Boonyayothin, W.; Thitithanyanont, A.; Smith, D.R.; et al. Rapid production of SARS-CoV-2 receptor binding domain (RBD) and spike specific monoclonal antibody CR3022 in Nicotiana benthamiana. Sci. Rep. 2020, 10, 17698. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Route | RBD Dose | Adjuvant | Vehicle PBS |
|---|---|---|---|---|
| C. RBD s.c. | s.c. | 0.2 µg * | - | ✓ |
| C. RBD Al(OH)3 s.c. | s.c. | 0.2 µg * | Al(OH)3 1:5 (v/v) | ✓ |
| C. RBD p.o. | p.o. | 0.2 µg + | - | ✓ |
| C. RBD CT p.o. | p.o. | 0.2 µg + | CT 1:1 (v/v) | ✓ |
| PBS | s.c. | - | - | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Govea-Alonso, D.O.; Malla, A.; Bolaños-Martínez, O.C.; Vimolmangkang, S.; Rosales-Mendoza, S. An Algae-Made RBD from SARS-CoV-2 Is Immunogenic in Mice. Pharmaceuticals 2022, 15, 1298. https://doi.org/10.3390/ph15101298
Govea-Alonso DO, Malla A, Bolaños-Martínez OC, Vimolmangkang S, Rosales-Mendoza S. An Algae-Made RBD from SARS-CoV-2 Is Immunogenic in Mice. Pharmaceuticals. 2022; 15(10):1298. https://doi.org/10.3390/ph15101298
Chicago/Turabian StyleGovea-Alonso, Dania O., Ashwini Malla, Omayra C. Bolaños-Martínez, Sornkanok Vimolmangkang, and Sergio Rosales-Mendoza. 2022. "An Algae-Made RBD from SARS-CoV-2 Is Immunogenic in Mice" Pharmaceuticals 15, no. 10: 1298. https://doi.org/10.3390/ph15101298
APA StyleGovea-Alonso, D. O., Malla, A., Bolaños-Martínez, O. C., Vimolmangkang, S., & Rosales-Mendoza, S. (2022). An Algae-Made RBD from SARS-CoV-2 Is Immunogenic in Mice. Pharmaceuticals, 15(10), 1298. https://doi.org/10.3390/ph15101298