
Mechanistic Insight into Apoptotic Induction in Human Rhabdomyosarcoma and Breast Adenocarcinoma Cells by Chnoospora minima: A Sri Lankan Brown Seaweed

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Yields of Crude Methanol Extract and Fractions of C. minima
2.2. Cytotoxic Effect of Crude Methanol Extracts and Fractions of C. minima on Human Rhabdomyosarcoma and MCF-7 Cells
2.2.1. MTT Assay
2.2.2. Neutral Red Assay
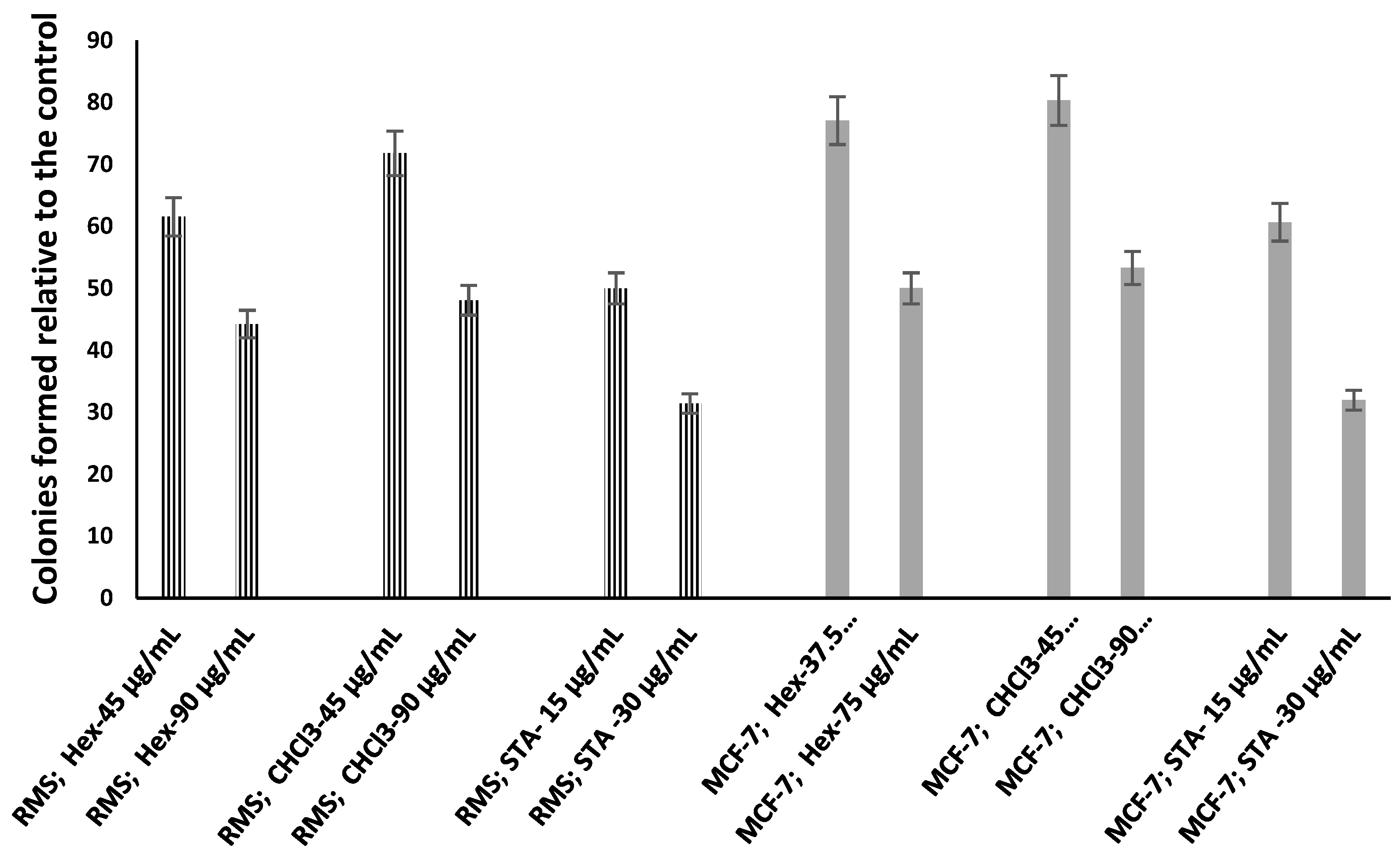
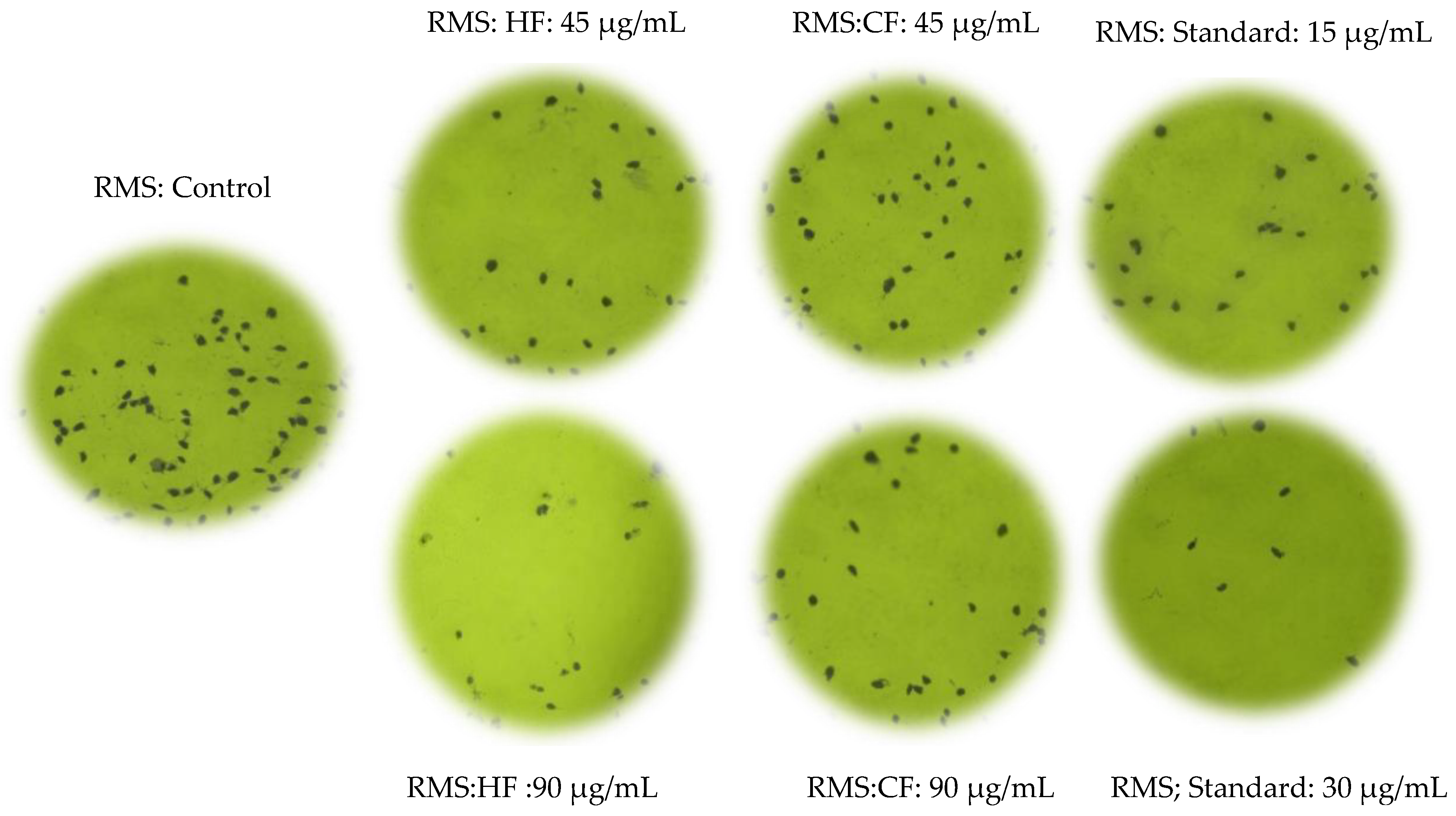
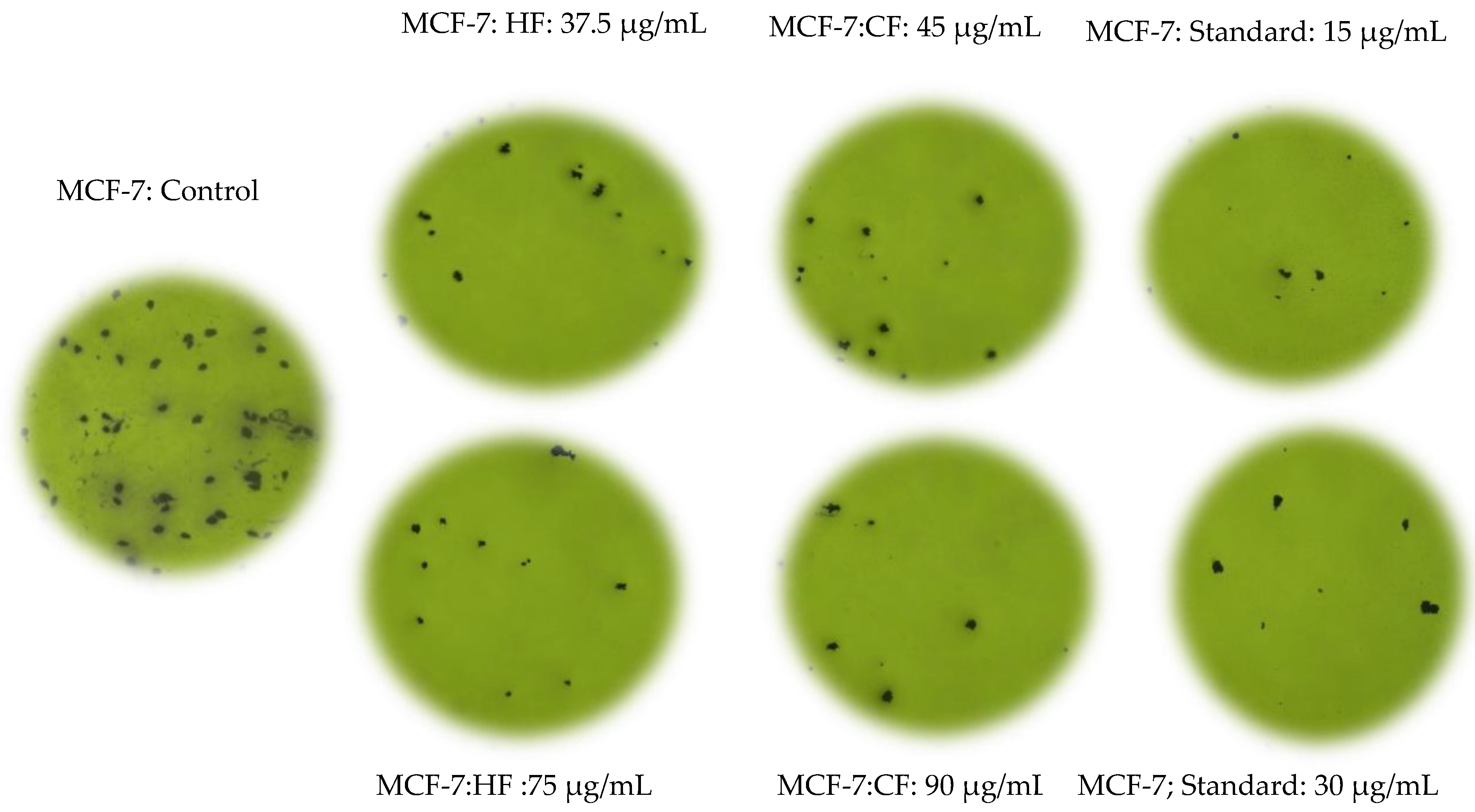
2.3. Hexane and Chloroform Fractions of C. minima-Inhibited Colony Formation
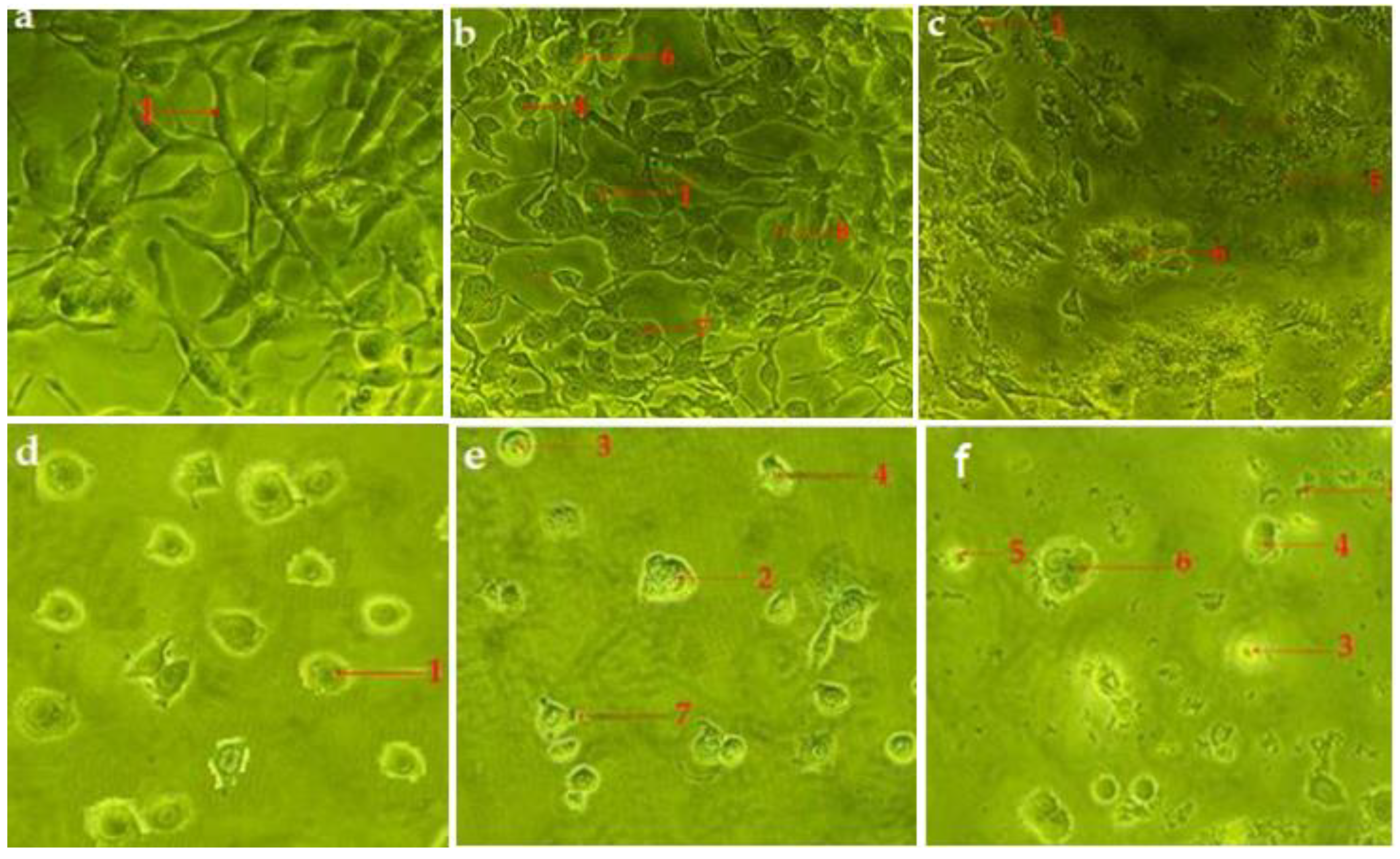
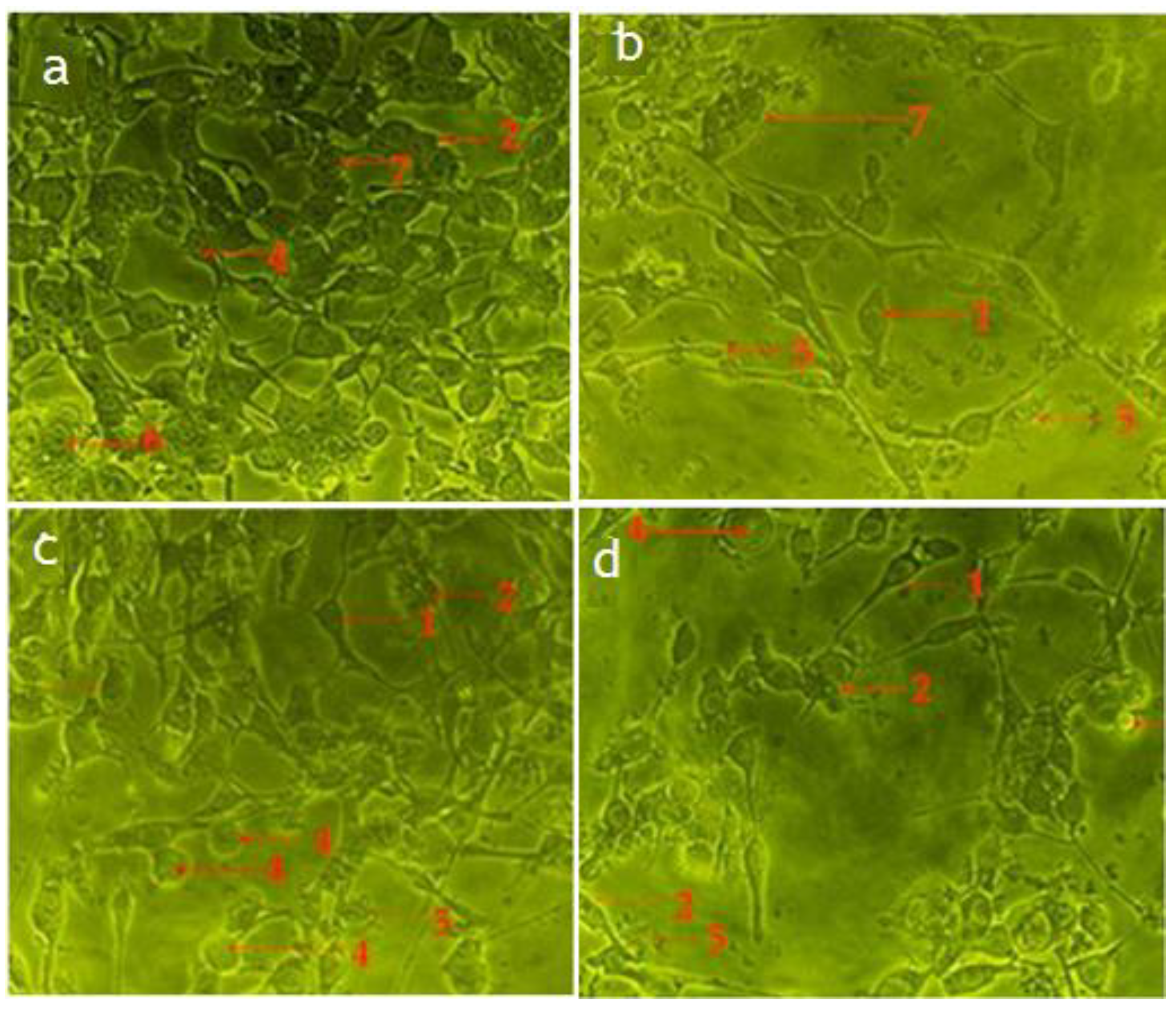
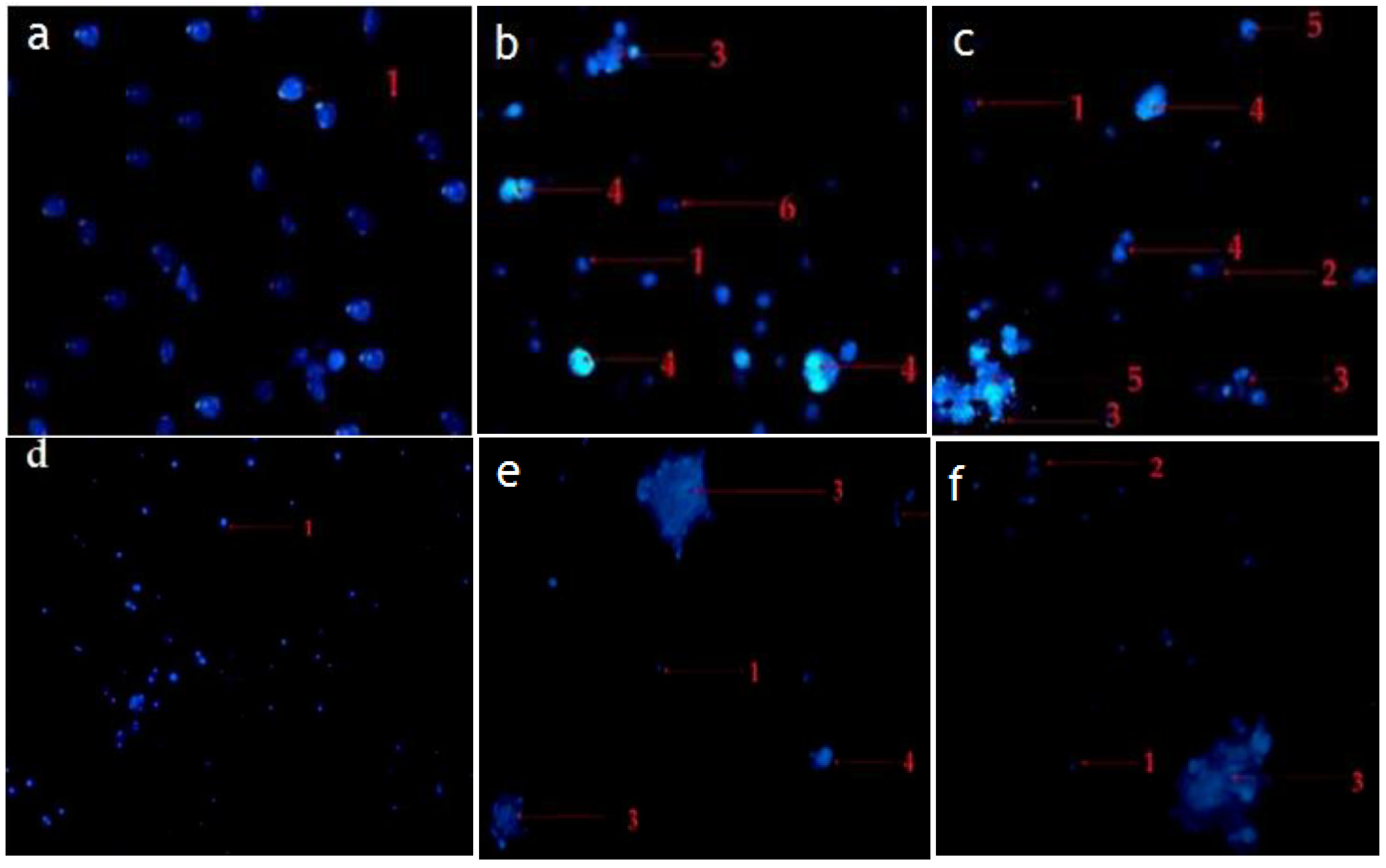
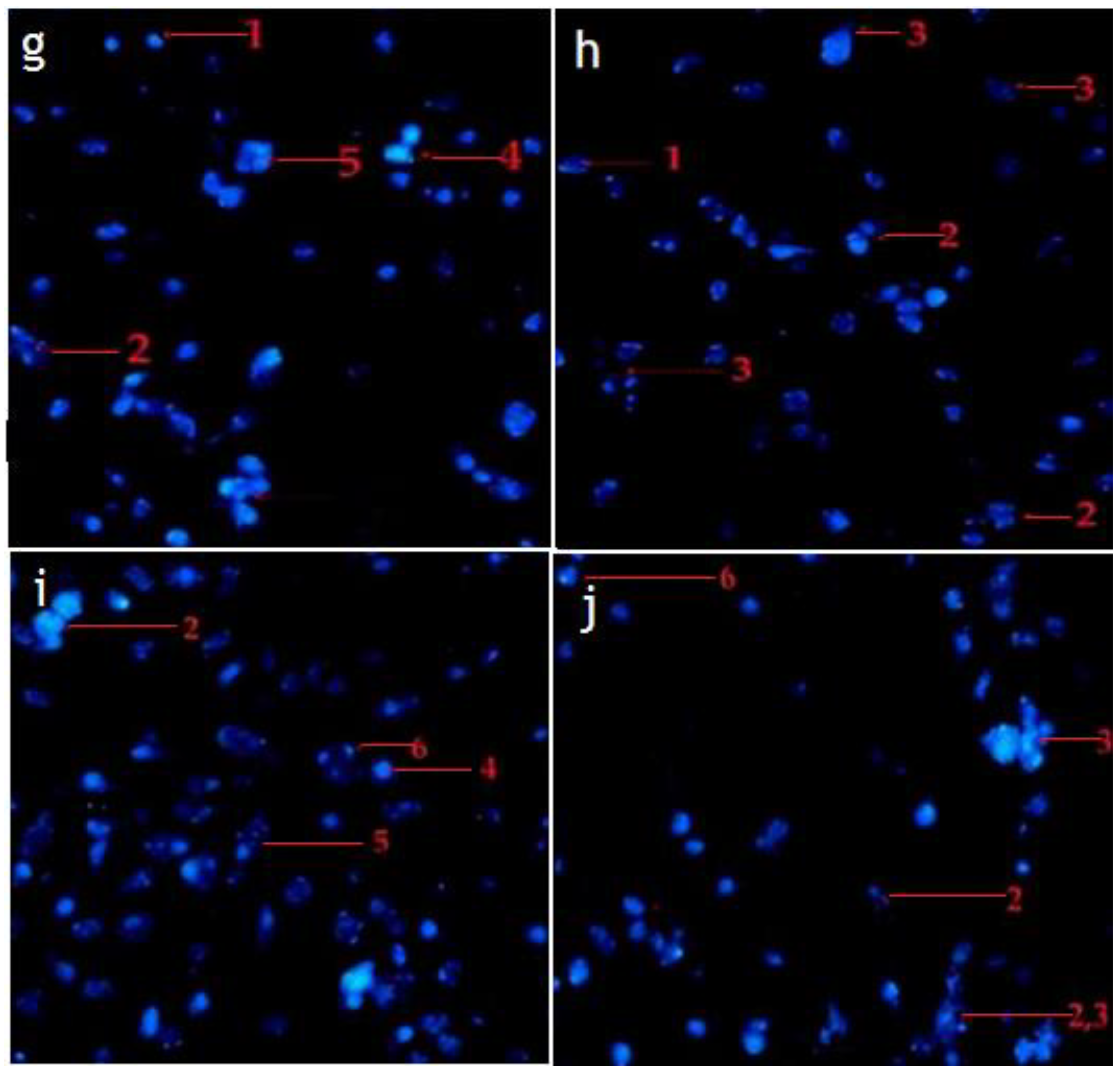
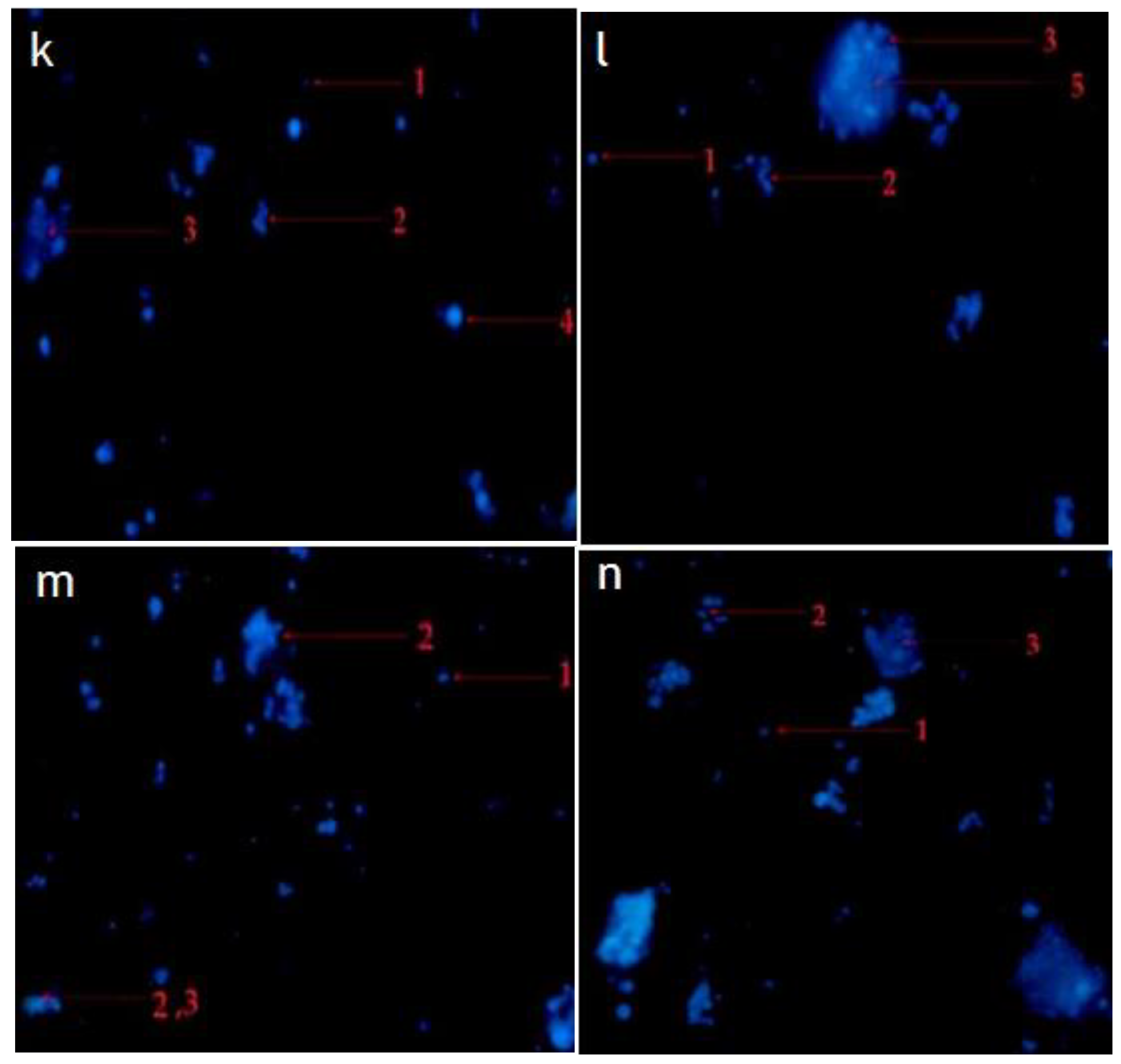
2.4. Hexane and Chloroform Fractions of C. minima Induced Morphological Alterations of Apoptosis in Human Rhabdomyosarcoma and MCF-7 Cells
2.4.1. Inverted Phase-Contrast Microscope
2.4.2. Fluorescence Staining (Hochest Staining)
2.5. Hexane and Chloroform Fractions of C. minima Induced Apoptosis and DNA Fragmentation by a Caspase-Dependent Pathway in Human Rhabdomyosarcoma Cells and by a Caspase-Independent Pathway in Human MCF-7 Cells
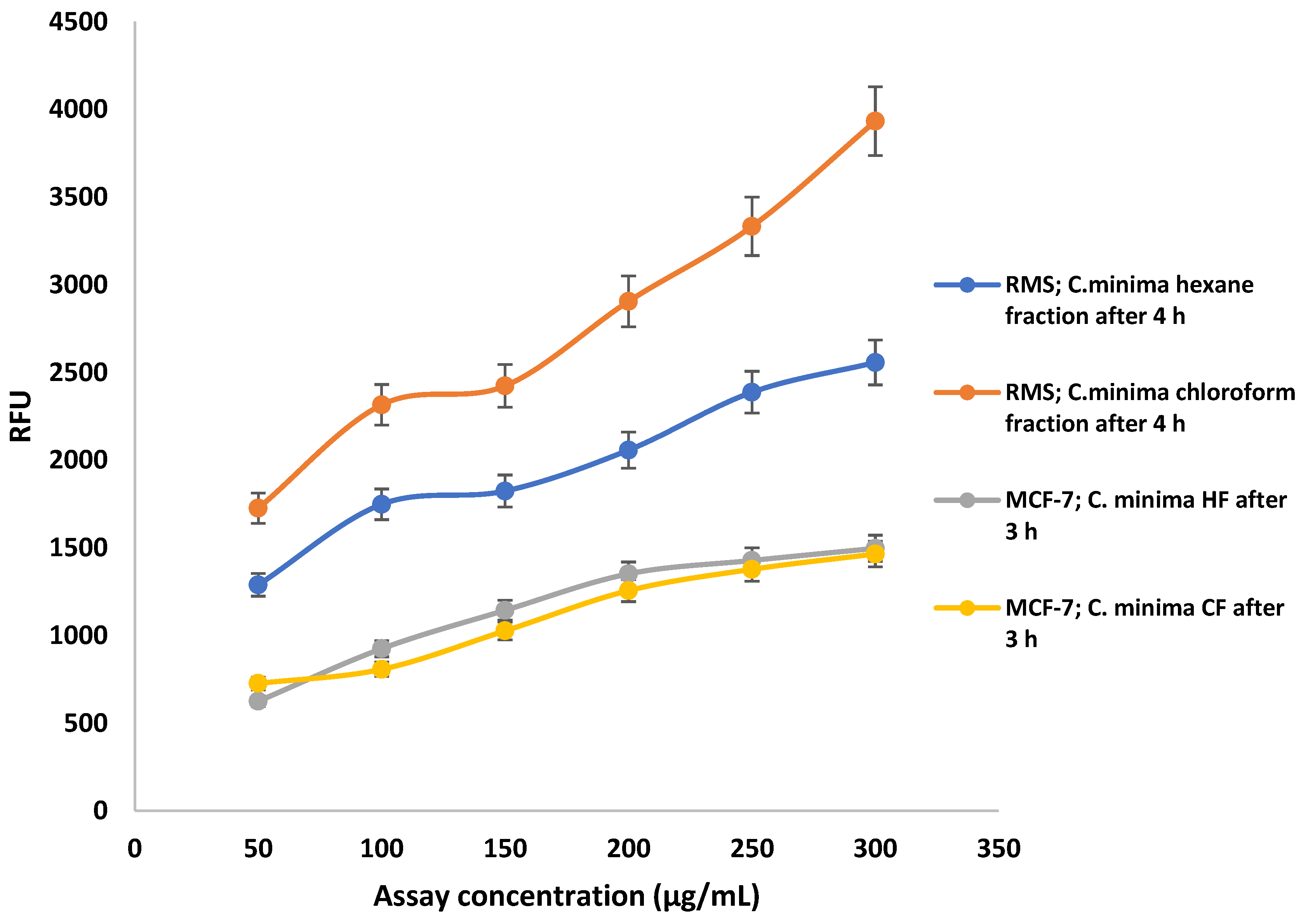
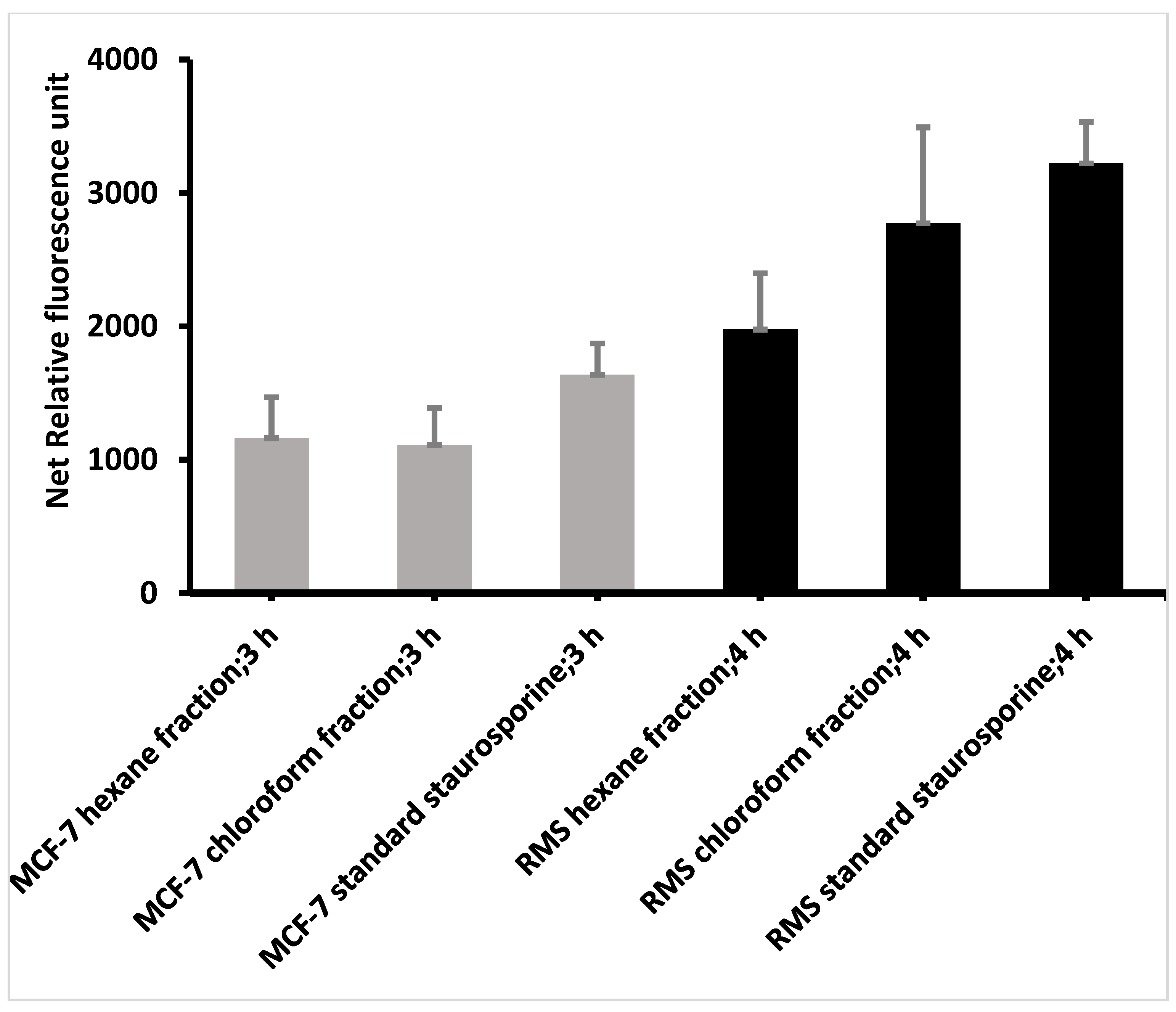
2.5.1. Caspase 3/7 Activity
2.5.2. DNA Fragmentation
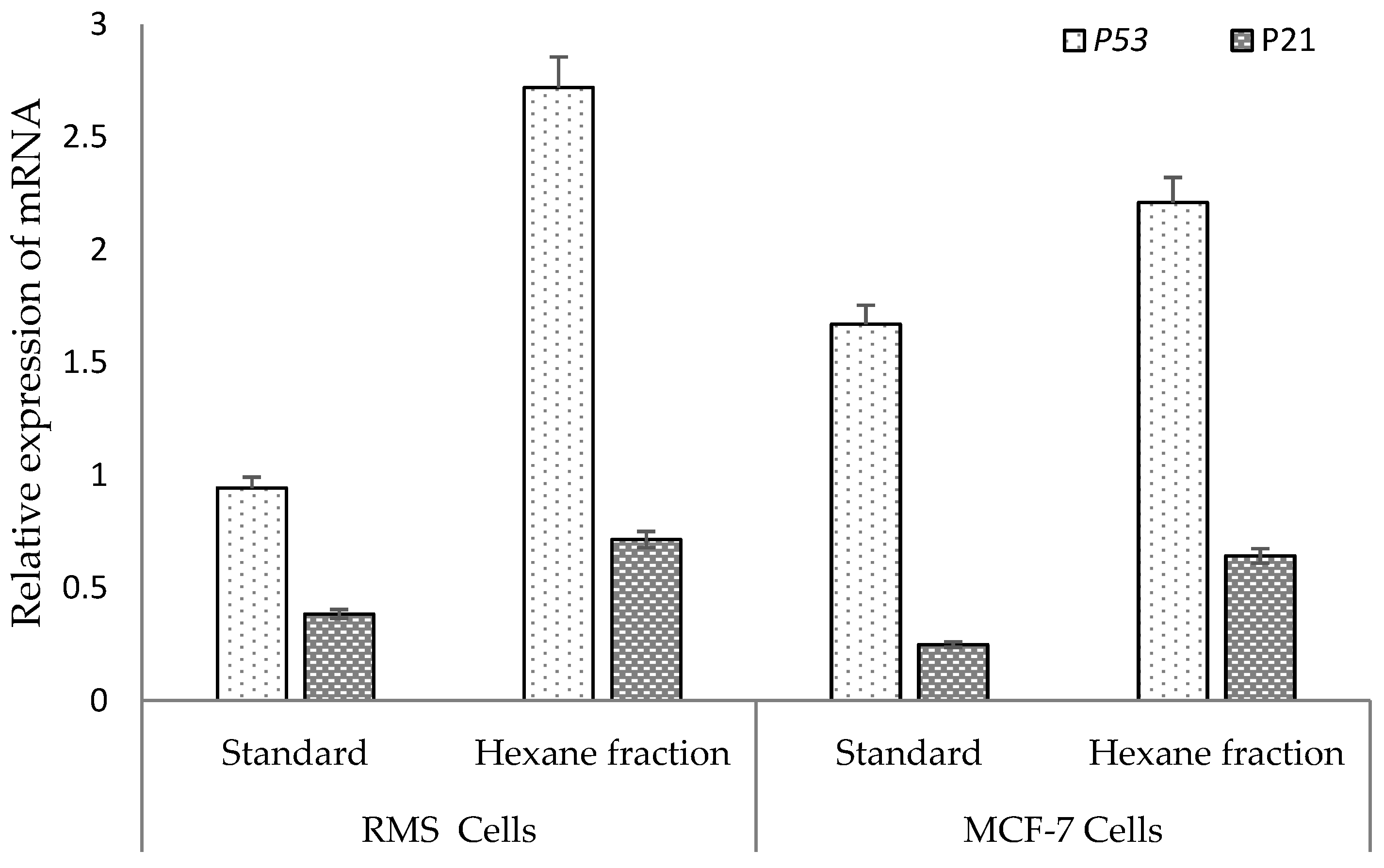
2.6. Hexane Fraction Induces Expression of the p53 Gene More Prominently Than the p21 Gene in RMS and MCF-7 Cells
2.7. Hexane and Chloroform Fractions Exhibited Moderate Levels of Total Phenolics, Flavonoids, Alkaloids, and Antioxidant Activity
2.7.1. Quantification of Total Phenols, Flavonoids, and Alkaloid Contents
2.7.2. In Vitro Antioxidant Activity
2.8. Identification of Volatile Components in Potent Anticancer Hexane Fraction by GC-MS
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Collection of Algae Sample
4.3. C. minima Extract and Solvent Fractions
4.4. Cell Culture Maintenance
4.5. Cell Survival Determination
4.5.1. 3-(4, 5-Dimethylthiazolyl-2)-2, 5-Diphenyltetrazolium Bromide (MTT Assay)
4.5.2. Neutral Red Assay
4.6. Clonogenic Assay
4.7. Morphological Alterations in Apoptosis
4.7.1. Inverted Phase-Contrast Microscope
4.7.2. Fluorescence Microscope
4.8. Caspase 3/7 Activity
4.9. DNA Fragmentation Assay
4.10. Gene Expression
4.11. Quantification of Phenolic, Flavonoid, and Alkaloid Contents
4.11.1. Estimation of Total Polyphenolic Content (TPC)
4.11.2. Estimation of Total Flavonoid Content (TFC)
4.11.3. Estimation of Total Alkaloid Content (TAC)
4.12. Antioxidant Capacity
4.12.1. DPPH Radical Scavenging Activity
4.12.2. ABTS+ Radical Scavenging Activity
4.12.3. Ferric Reducing Antioxidant Power (FRAP)
4.12.4. Ferrous Iron Chelating Capacity (FICC)
4.12.5. Oxygen Radical Absorbance Capacity (ORAC)
4.13. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RMS | Human rhabdomyosarcoma |
| MCF-7 | Human breast adenocarcinoma |
| MTT | 3-(4, 5-dimethyl thiazolyl-2)-2, 5-diphenyltetrazolium bromide |
| DNA | Deoxyribonucleic acid |
| FBS | Fetal bovine serum |
| DMEM | Dulbecco’s Modified Eagle’s medium |
| PBS | Phosphate buffered saline |
| GC-MS | Gas chromatography-mass spectrometry |
| TPC | Total phenolic content |
| TFC | Total flavonoid content |
| TAC | Total alkaloid content |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| ABTS | 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid |
| ORAC | Oxygen radical absorbance capacity |
| FRAP | ferric reducing antioxidant power |
| FICC | ferrous iron chelating capacity |
References
- Afonso, N.C.; Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Brown Macroalgae as Valuable Food Ingredients. Antioxidants 2019, 8, 365. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 252–263. [Google Scholar] [CrossRef]
- World Health Organization. Cancer. 2018. Available online: http://www.who.int/mediacentre/factsheets/fs297/en/ (accessed on 18 August 2020).
- National Breast Cancer Foundations. INC Breast Cancer Facts. Available online: https://www.nationalbreastcancer.org/breast-cancer-facts (accessed on 20 August 2020).
- BC Cancer Registry. BC Cancer Registry Annual Report 2019. Available online: http://www.bccancer.bc.ca/about-site/Documents/BCCancerResearch-AnnualReport2018.pdf (accessed on 15 September 2020).
- Balawardena, J.; Skandarajah, T.; Rathnayake, W.; Joseph, N. Breast Cancer Survival in Sri Lanka. JCO Glob. Oncol. 2020, 6, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Hettmer, S.; Li, Z.; Billin, A.N.; Barr, F.G.; Cornelison, D.D.W.; Ehrlich, A.R.; Guttridge, D.C.; Hayes-Jordan, A.; Helman, L.J.; Houghton, P.J.; et al. Rhabdomyosarcoma: Current Challenges and Their Implications for Developing Therapies. Cold Spring Harb. Perspect. Med. 2014, 4, a025650. [Google Scholar] [CrossRef] [PubMed]
- Fourquet, A.; Kirova, Y.; Bollet, M.-A.; Tournat, H.; Dendale, R.; Campana, F. Meta-analyses of the effects of radiotherapy in breast cancer: The ultimate evidence? Cancer Radiother. 2008, 12, 554–558. [Google Scholar] [CrossRef]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245–304. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.; Osborne, S.; Masci, P.; Gobe, G. Marine-Based Nutraceuticals: An Innovative Trend in the Food and Supplement Industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef] [Green Version]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cencic, A.; Chingwaru, W. The role of functional foods, nutraceuticals, and food supplements in intestinal health. Nutrients 2010, 2, 611–625. [Google Scholar] [CrossRef]
- Pradhan, B.; Nayak, R.; Patra, S.; Jit, B.P.; Ragusa, A.; Jena, M. Bioactive Metabolites from Marine Algae as Potent Pharmacophores against Oxidative stress-associated Human Diseases: A Comprehensive Review. Molecules 2020, 26, 37. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P.; Sulaiman Rahman, H. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front. Pharmacol. 2018, 9, 2–11. [Google Scholar] [CrossRef] [Green Version]
- An, W.; Lai, H.; Zhang, Y.; Liu, M.; Lin, X.; Cao, S. Apoptotic Pathway as the Therapeutic Target for Anticancer Traditional Chinese Medicines. Front. Pharmacol. 2019, 10, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Schwartsmann, G.; da Rocha, A.B.; Berlinck, R.G.; Jimeno, J. Marine organisms as a source of new anticancer agents. Lancet Oncol. 2001, 2, 221–225. [Google Scholar] [CrossRef]
- Lakmal, H.C.; Samarakoon, K.W.; Lee, W.; Lee, J.-H.; Abeytunga, D.; Lee, H.-S.; Jeon, Y.-J. Anticancer and antioxidant effects of selected Sri Lankan marine algae. J. Natl. Sci. Found. 2014, 42, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Franken, N.A.P.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Saraste, A. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef]
- Jassbi, A.R.; Mohabati, M.; Eslami, S.; Sohrabipour, J.; Miri, R. Biological activity and chemical constituents of red and brown algae from the persian gulf. Iran. J. Pharm. Res. 2013, 12, 339–348. [Google Scholar]
- Bhimba, V.; Meenupriya, J.; Joel, E.; Naveena, D.; Kumar, S.; Hangaraj, M. Antibacterial activity and characterization of secondary metabolites isolated from mangrove plant Avicennia officinalis. Asian Pac. J. Trop. Med. Trop. Med. 2010, 3, 544–546. [Google Scholar] [CrossRef] [Green Version]
- Okla, M.K.; Alatar, A.A.; Hegazy, A.K.; Ahmed Rady, A. Biological Activity of Calligonum comosum Extracts as Antibacterial and Antioxidant. J. Pure Appl. Microbiol. 2014, 8, 529–534. [Google Scholar]
- Elaiyaraja, A.; Chandramohan, G. Comparative phytochemical profile of crinum defixum ker-gawler leaves using GC-MS. J. Drug Deliv. Ther. 2018, 8, 365–380. [Google Scholar]
- Pal, A.; Kamthania, M.C.; Kumar, A. Bioactive Compounds and Properties of Seaweeds—A Review. OALib 2014, 1, 1–17. [Google Scholar] [CrossRef]
- Green, L.M.; Reade, J.L.; Ware, C.F. Rapid colormetric assay for cell viability: Application to quantitate cytotoxic and growth inhibitory lymphokines. J. Immunol. Methods 1984, 70, 257–268. [Google Scholar] [CrossRef]
- Parveen, S.; Nadumane, V.K. Anti-angiogenesis and apoptogenic potential of the brown marine alga, Chnoospora minima. Future J. Pharm. Sci. 2020, 6, 19. [Google Scholar] [CrossRef]
- Shyamala, V.; AnaghaK, T.N. In vitro antioxidant and antiproliferative activities of macro algae against MCF-7 cell line. J. Pharm. Biomed. Sci. 2010, 32, 1413–1424. [Google Scholar]
- Khanavi, M.; Nabavi, M.; Ardekani, S.M.; Sohrabipour, J.; Mohammed, S.; Nabavi, G.P.; Ostad, N. Cytotoxic activity of some marine brown algae against cancer cell lines. Biol. Res. 2010, 43, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Abd Aziz, N.A.; Hasham, R.; Sarmidi, M.R.; Suhaimi, S.H.; Idris, M.K.H. A review on extraction techniques and therapeutic value of polar bioactives from Asian medicinal herbs: Case study on Orthosiphon aristatus, Eurycoma longifolia and Andrographis paniculata. Saudi Pharm. J. 2021, 29, 143–165. [Google Scholar] [CrossRef]
- Pacheco, B.S.; dos Santos, M.A.Z.; Schultze, E.; Martins, R.M.; Lund, R.G.; Seixas, F.K.; Colepicolo, P.; Collares, T.; Paula, F.R.; De Pereira, C.M.P. Cytotoxic Activity of Fatty Acids from Antarctic Macroalgae on the Growth of Human Breast Cancer Cells. Front. Bioeng. Biotechnol. 2018, 6, 2–12. [Google Scholar] [CrossRef]
- Hussein, H.A.; Abdullah, M.A. Anticancer Compounds Derived from Marine Diatoms. Mar. Drugs 2020, 18, 356. [Google Scholar] [CrossRef]
- Houghton, P.J.; Raman, A. Methods for extraction and sample clean-up. In Laboratory Handbook for the Fractionation of Natural Extracts; Springer: Boston, MA, USA, 1998; pp. 22–53. [Google Scholar]
- Munshi, A.; Hobbs, M.; Meyn, R.E. Clonogenic Cell Survival Assay. In Chemosensitivity; Humana Press: Totowa, NJ, USA, 2005; pp. 21–28. [Google Scholar]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.C. Mechanisms of Apoptosis. Am. J. Pathol. 2000, 157, 1415–1430. [Google Scholar] [CrossRef]
- Jin, Z.; El-Deiry, W.S. Overview of cell death signaling pathways. Cancer Biol. Ther. 2005, 4, 147–171. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Plati, J.; Bucur, O.; Khosravi-Far, R. Apoptotic cell signaling in cancer progression and therapy. Integr. Biol. 2011, 3, 279–296. [Google Scholar] [CrossRef]
- Wolf, B.B.; Schuler, M.; Escheverri, F.; Green, D.R. Caspase-3 is the primary activator of apoptotic DNA fragmentation via DNA fragmentation factor-45/inhibitor of caspase-activated DNase inactivation. J. Biol. Chem. 1999, 274, 30651–30656. [Google Scholar] [CrossRef] [Green Version]
- Sivakumaran, N.; Samarakoon, S.R.; Adhikari, A.; Ediriweera, M.K.; Tennekoom, K.H.; Malavige, N.; Thabrew, I.; Shrestha, R.L. Cytotoxic and Apoptotic Effects of Govaniadine Isolated from Corydalis govaniana Wall. Roots on Human Breast Cancer (MCF-7) Cells. BioMed Res. Int. 2018, 2018, 3171348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuler, M.; Wetzel, B.E.; Goldstein, C.J.; Fitzgerald, P.; Green, R.D. p53 Induces apoptosis by caspase activation through mitochondrial cytochrome c release. Mol. Basis Cell Dev. Biol. 2000, 275, 7337–7342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marseglia, L.; Manti, S.; Angelo, G.D.; Nicotera, A.; Parisi, E.; Rosa, G.D.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity, A Critical Component in Human Diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, H.; Shad, M.A.; Rehman, N.; Andaleeb, H.; Ullah, N. Effect of solvent polarity on extraction yield and antioxidant properties of phytochemicals from bean (Phaseolus vulgaris) seeds. Braz. J. Pharm. Sci. 2020, 56, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Williams, W.B.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Sampath, S.G.; Dheeba, K.P.; Sivakumar, R. In vitro antidiabetic, antioxidant and anti- inflammatory activity of Clitoria Ternatea. Int. J. Pharm. Sci. 2014, 6, 342–347. [Google Scholar]
- Ranasinghe, P. Antioxidant Activity of Caryota urens L. (Kithul) Sap. Trop. Agric. Res. 2012, 23, 117–125. [Google Scholar] [CrossRef] [Green Version]
- RahbarSaadat, Y.; Saeidi, N.; Zununi Vahed, S.; Barzegari, A.; Barar, J. An update to DNA ladder assay for apoptosis detection. BioImpacts 2017, 5, 25–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, B.; Reddy, V.R.K.; Sulaiman, C.T. Total Phenolics and Flavonoids in Selected Justicia Species. J. Pharm. Phytochem. 2013, 2, 72–73. [Google Scholar]
- Gedara, S.R.; Abdel-Halim, O.B.; El-Sharkawy, S.H.; Salama, O.M.; Shier, T.W.; Halim, A.F. Cytotoxic hydroazulene diterpenes from the brown alga Dictyota dichotoma. Z. Naturforsch. C J. Biosci. 2003, 58, 17–22. [Google Scholar] [CrossRef]
- Miranda-Delgado, A.; Montoya, M.J.; Paz-Araos, M.; Mellado, M.; Villena, J.; Arancibia, P.; Madrid, A.; Jara-Gutiérrez, C. Antioxidant and anticancer activities of brown and red seaweed extracts from Chilean coasts. Lat. Am. J. Aquat. Res. 2008, 46, 301–313. [Google Scholar] [CrossRef]
- Xu, J.; Yue, R.Q.; Liu, J.; Ho, H.M.; Yi, T.; Chen, H.; Han, Q.B. Structural diversity requires individual optimization of ethanol concentration in polysaccharide precipitation. Int. J. Biol. Macromol. 2014, 67, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Syed Abdul Rahman, S.N.; Abdul Wahab, N.; Abd Malek, S.N. In Vitro Morphological Assessment of Apoptosis Induced by Antiproliferative Constituents from the Rhizomes of Curcuma zedoaria. Evid.-Based Complement. Altern. Med. 2013, 2013, 257108. [Google Scholar] [CrossRef] [Green Version]
- Crowley, L.C.; Marfell, B.J.; Waterhouse, N.J. Analyzing Cell Death by Nuclear Staining with Hoechst 33342. Cold Spring Harb. Protoc. 2016, 2016, 34–38. [Google Scholar] [CrossRef]
- Tor, Y.S.; Yazan, L.S.; Foo, J.B.; Wibowo, A.; Ismail, N.; Cheah, Y.K.; Abdullah, R.; Ismail, M.; Ismail, I.S.; Yeap, S.K. Induction of Apoptosis in MCF-7 Cells via Oxidative Stress Generation, Mitochondria-Dependent and Caspase-Independent Pathway by Ethyl Acetate Extract of Dillenia suffruticosa and Its Chemical Profile. PLoS ONE 2015, 10, e0127441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreevidya, N.; Mehrotra, S. Spectrometric method for estimation of precipitable with Dragendor’s reagent in plant material. J. Aoac. Int. 2003, 36, 1124–1127. [Google Scholar] [CrossRef] [Green Version]
- Blois, M.S. Antioxidant determination by use of stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization Assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Szeto, Y.T. Total antioxidant capacity of teas by the ferric reducing/antioxidant power assay. J. Agric. Food Chem. 1999, 47, 633–636. [Google Scholar] [CrossRef]
- Carter, P. Spectrophotometric determination of serum iron at the sub-microgram level with a new reagent ferrozine. Annu. Biochem. 1971, 40, 450–458. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Mallard, W.G.; Linstrom, P.J. (Eds.) NIST Standard Reference Database, National Institute of Standards and Technology; National Institute of Standards and Technology: Washington, DC, USA, 2008.
- Phang, C.W.; Malek, S.N.; Ibrahim, H. Antioxidant potential, cytotoxic activity and total phenolic content of Alpinia pahangensis rhizomes. BMC Complement. Altern. Med. 2013, 13, 243. [Google Scholar] [CrossRef] [Green Version]
- Abdelmageed, N.; Morad, S.A.F.; Elghoneimy, A.A.; Syrovets, T.; Simmet, T.; El-Zorba, H.; El-Banna, H.A.; Cabot, M.; Abdel-Aziz, M.I. Oleanolic acid methyl ester, a novel cytotoxic mitocan, induces cell cycle arrest and ROS-Mediated cell death in castration-resistant prostate cancer PC-3 cells. Biomed. Pharmacother. 2017, 96, 417–425. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts/Fractionated Extracts | % Yield |
|---|---|
| Crude methanol extract | 27.2% |
| Hexane fraction | 0.10% |
| Chloroform fraction | 0.47% |
| Ethyl acetate fraction | 0.19% |
| Aqueous fraction | 5.30% |
| Extract/Fraction | RMS | MCF-7 |
|---|---|---|
| Crude methanol extract | 197.23 ± 5.68 | 221.15 ± 4.90 |
| Hexane fraction | 93.98 ± 1.33 * | 90.58 ± 4.34 * |
| Chloroform fraction | 106.94 ± 1.68 * | 97.73 ± 2.92 * |
| Ethyl acetate fraction | 235.15 ± 5.45 | 140.39 ± 1.48 |
| Aqueous fraction | 292.65 ± 0.85 | 162.14 ± 4.44 |
| Standard cycloheximide | 36.17 ± 1.78 | 28.76 ± 0.55 |
| Extract/Fraction | Cytotoxicity on Vero Cells IC50 (μg/mL) |
|---|---|
| Hexane fraction | 109.23 ± 1.05 * |
| Chloroform fraction | 62.54 ± 0.38 * |
| Standard cycloheximide | 29.55 ± 1.56 |
| Extract/Fraction | Neutral Red; Cytotoxicity IC50 (μg/mL) | |
|---|---|---|
| RMS | MCF-7 | |
| Hexane fraction | 133.13 ± 4.50 * | 119.46 ± 1.32 * |
| Chloroform fraction | 142.09 ± 0.64 | 135.89 ± 1.16 |
| Standard cycloheximide | 32.78 ± 0.91 | 27.84 ± 0.33 |
| Sample (μg/mL) | Plating Efficiency (PE) | Surviving Fraction (SF) | Survival Rate (%) | |
|---|---|---|---|---|
| MCF-7 cells | C.H1: 45 | 0.096 | 0.009 | 61.54 ± 1.23 a |
| C.H2: 90 | 0.069 | 0.005 | 44.23 ± 1.08 | |
| C.C1: 45 | 0.112 | 0.013 | 71.79 ± 2.13 a | |
| C.C2: 90 | 0.075 | 0.006 | 48.08 ± 1.56 | |
| S1: 15 | 0.078 | 0.006 | 50.00 ± 0.98 | |
| S2: 30 | 0.049 | 0.002 | 31.41 ± 1.42 | |
| RMS cells | C.H1: 37.5 | 0.094 | 0.009 | 77.05 ± 2.54 |
| C.H2: 75 | 0.061 | 0.004 | 50.00 ± 1.34 | |
| C.C1: 45 | 0.098 | 0.010 | 80.33 ± 3.45 | |
| C.C2: 90 | 0.065 | 0.004 | 53.28 ± 1.45 | |
| S1: 15 | 0.074 | 0.005 | 60.66 ± 1.89 | |
| S2: 30 | 0.039 | 0.002 | 31.97 ± 3.45 |
| Extract/Fraction | TPC (mg GAE/g) | TFC (mg QE/g) | Total Alkaloids (mg of PE/g) |
|---|---|---|---|
| Hexane fraction | 2.86 ± 0.24 | 0.23 ± 0.03 | 1.36 ± 0.69 |
| Chloroform fraction | 38.42 ± 3.21 * | 3.11 ± 0.51 * | 2.79 ± 0.31 * |
| Extract/ Fraction | IC50 (mg/mL) | Activity Equivalent to Standard (mg TE/g) | |||
|---|---|---|---|---|---|
| DPPH | ABTS | FICA | FRAP | ORAC | |
| Hexane fraction | 1.88 ± 0.02 | 4.71 ± 0.31 | 3.67 ± 0.02 | 8.57 ± 1.13 ** | 4.08 ± 1.44 ** |
| Chloroform fraction | 0.75 ± 0.002 * | 0.12 ± 0.009 * | 0.93 ± 0.002 * | 13.95 ± 1.55 | 19.72 ± 2.92 |
| Trolox (standard) | 0.011 ± 0.00 | 0.008 ± 0.00 | N/A | N/A | N/A |
| EDTA (standard) | N/A | N/A | 0.019 ± 00 | N/A | N/A |
| Retention Time & % | Chemical Name | Nature of the Compound | Molecular Formula | Peak Area (%) | Reported Activity |
|---|---|---|---|---|---|
| 24.318 | Dodecanoic acid methyl ester | Fatty acid methyl esters | C16H26O2 | 6.873% | Antimicrobial Antioxidant [21] |
| 27.090 | Diethyl phthalate | Phthalate ester | C12H14O4 | 6.309% | Anticancer Anti-bacterial [22] |
| 31.623 | Methyl tetradecanoate | Fatty acid methyl esters | C15H30O2 | 38.314% | Antioxidant Anticancer [23] |
| 38.439 | Pentadecanoic acid, 14-methyl-, methyl ester | Palmitic acid methyl ester | C17H34O2 | 15.799% | Antioxidant, antifungal and antimicrobial [24] |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| p21 | 5′-CTG-TCA-CAG-GCG-GTT-ATG-AA-3′ | 3′-TGT-GCT-CA C-TTC-AGG-GTC-AC-5′. |
| p53 | 5′-GCG-CAC-AGA-GGA-AGA-GA A-TC-3′ | 5′-CTC-TCG-GAA-CAT-CTC-GAA-GC-3′; |
| ß actin | 5′-GTGGGCCGCCCTAGGCACCAG-3′ | 5′-GGAGGAAGAGGATGCGGCAGT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunathilaka, T.L.; Dilrangi, K.H.; Ranasinghe, P.; Samarakoon, K.W.; Peiris, L.D.C. Mechanistic Insight into Apoptotic Induction in Human Rhabdomyosarcoma and Breast Adenocarcinoma Cells by Chnoospora minima: A Sri Lankan Brown Seaweed. Pharmaceuticals 2021, 14, 1154. https://doi.org/10.3390/ph14111154
Gunathilaka TL, Dilrangi KH, Ranasinghe P, Samarakoon KW, Peiris LDC. Mechanistic Insight into Apoptotic Induction in Human Rhabdomyosarcoma and Breast Adenocarcinoma Cells by Chnoospora minima: A Sri Lankan Brown Seaweed. Pharmaceuticals. 2021; 14(11):1154. https://doi.org/10.3390/ph14111154
Chicago/Turabian StyleGunathilaka, Thilina Lakmini, Kulathungage Hiranthi Dilrangi, Pathmasiri Ranasinghe, Kalpa W. Samarakoon, and L. Dinithi C. Peiris. 2021. "Mechanistic Insight into Apoptotic Induction in Human Rhabdomyosarcoma and Breast Adenocarcinoma Cells by Chnoospora minima: A Sri Lankan Brown Seaweed" Pharmaceuticals 14, no. 11: 1154. https://doi.org/10.3390/ph14111154
APA StyleGunathilaka, T. L., Dilrangi, K. H., Ranasinghe, P., Samarakoon, K. W., & Peiris, L. D. C. (2021). Mechanistic Insight into Apoptotic Induction in Human Rhabdomyosarcoma and Breast Adenocarcinoma Cells by Chnoospora minima: A Sri Lankan Brown Seaweed. Pharmaceuticals, 14(11), 1154. https://doi.org/10.3390/ph14111154