
Network Pharmacology Study and Experimental Confirmation Revealing the Ameliorative Effects of Decursin on Chemotherapy-Induced Alopecia



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Gene Comparison Indicates the Close Relationship between Decursin and Alopecia
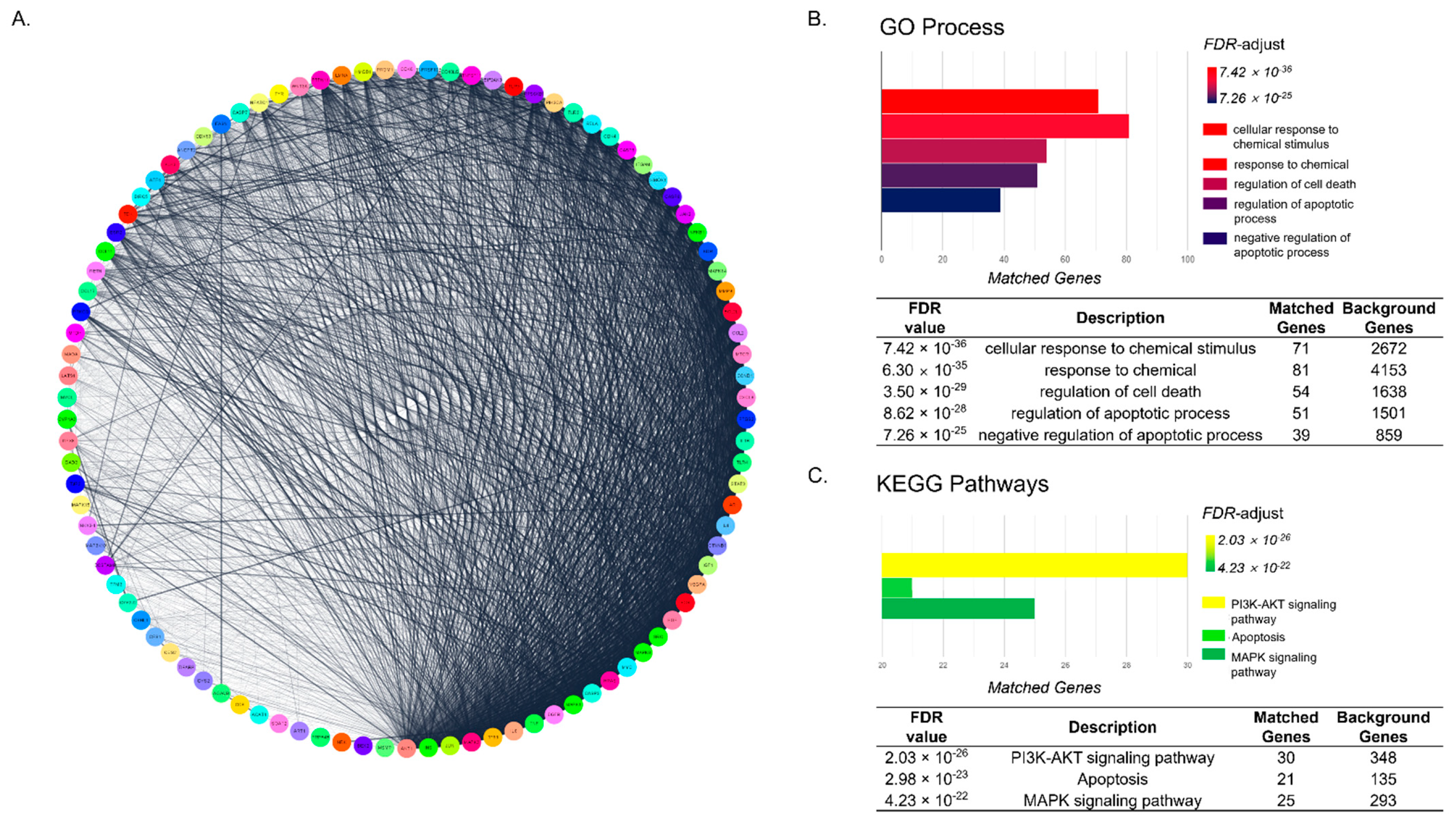
2.2. Functional Enrichment Analysis Shows That Underlying Mechanism of Decursin Is Predicted to Be Related to Apoptosis Pathway in CIA
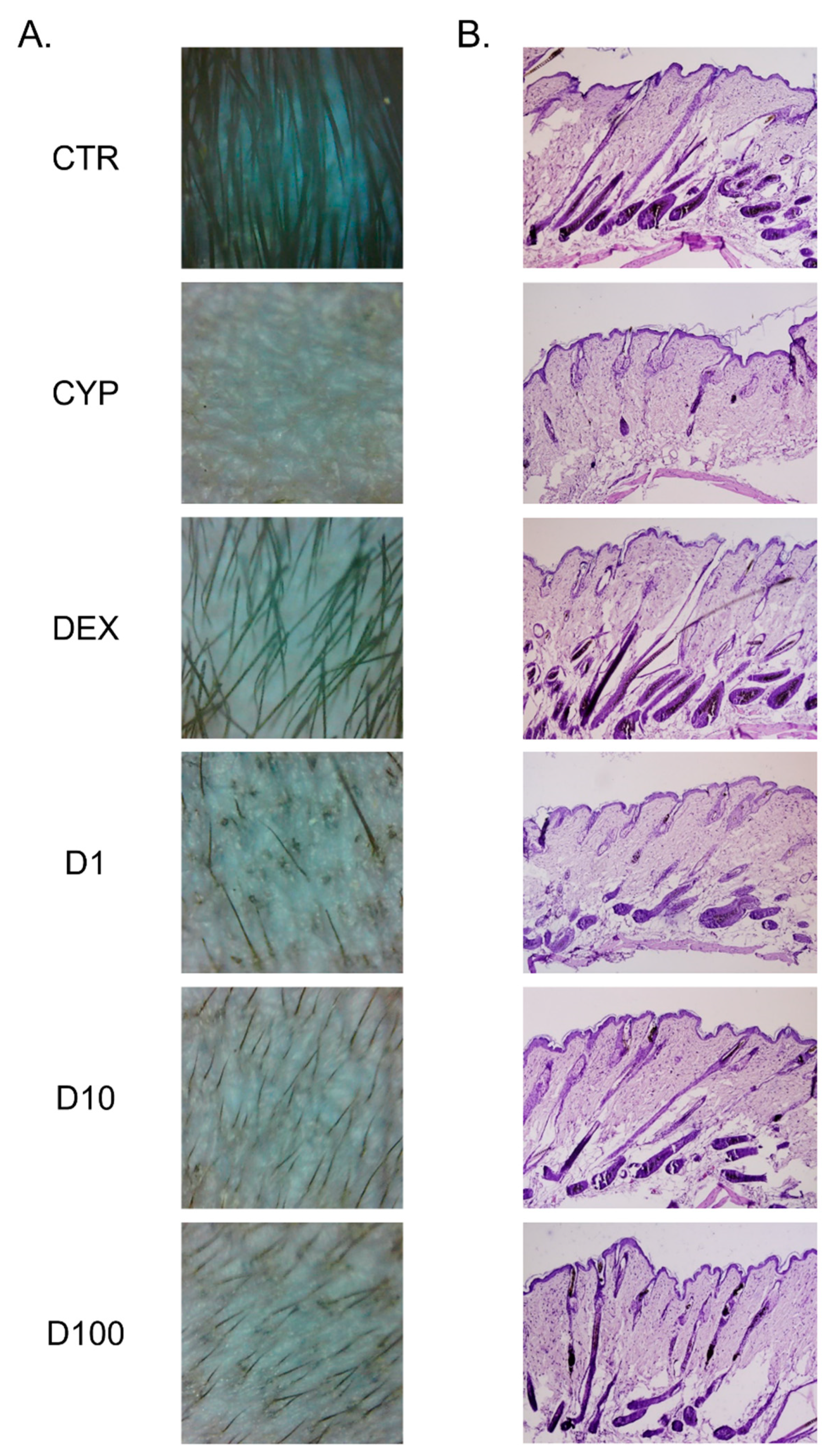
2.3. Decursin Promotes the Growth of Hair Fiber in CIA Mice
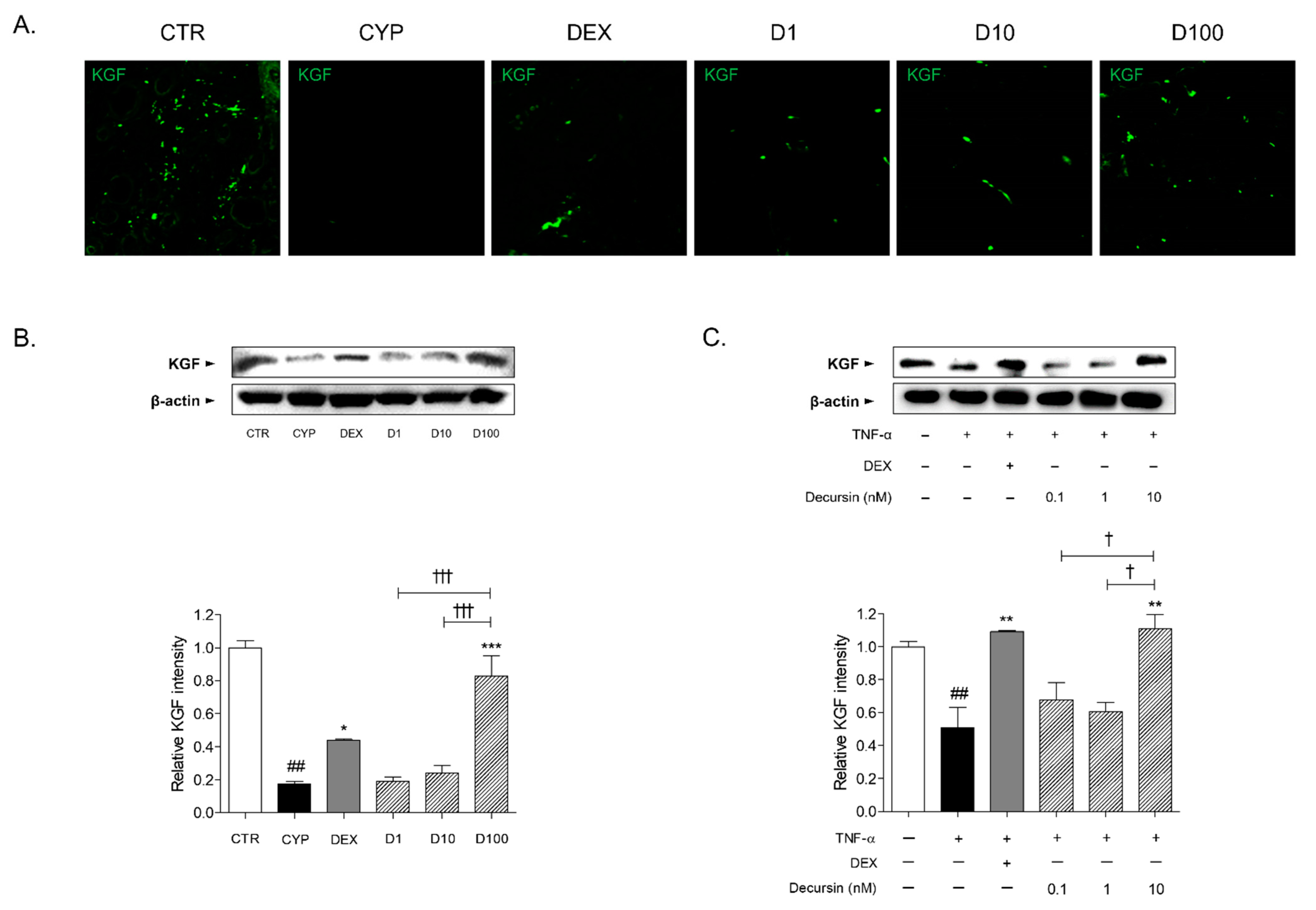
2.4. Decursin Increases KGF Expression in Skin Tissues of CIA Mice and TNF-α-Stimulated Keratinocytes
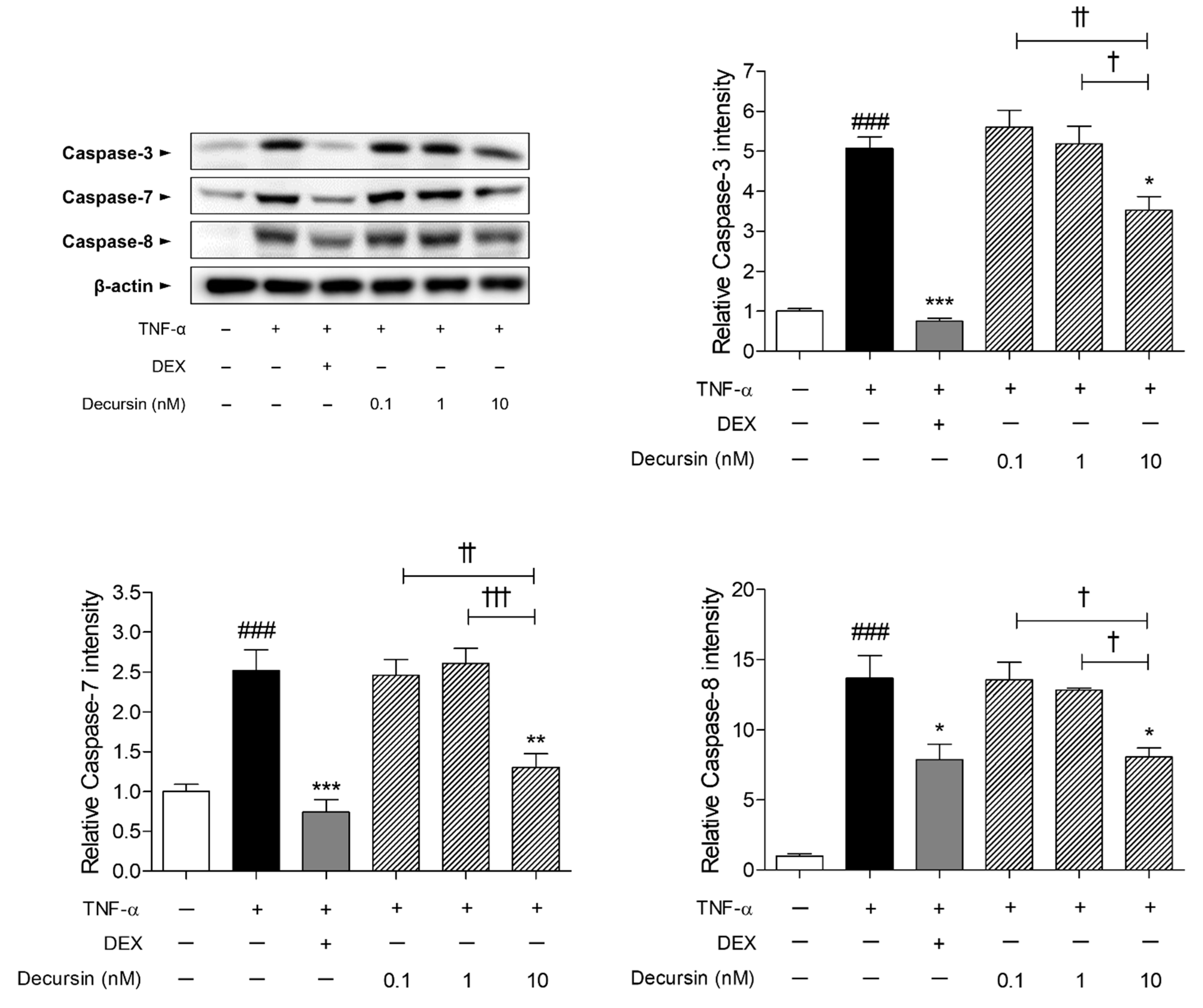
2.5. Decursin Decreased the Expressions of Apoptosis Factors including Caspase-3, -7, and -8 in TNF-α-Stimulated Keratinocytes
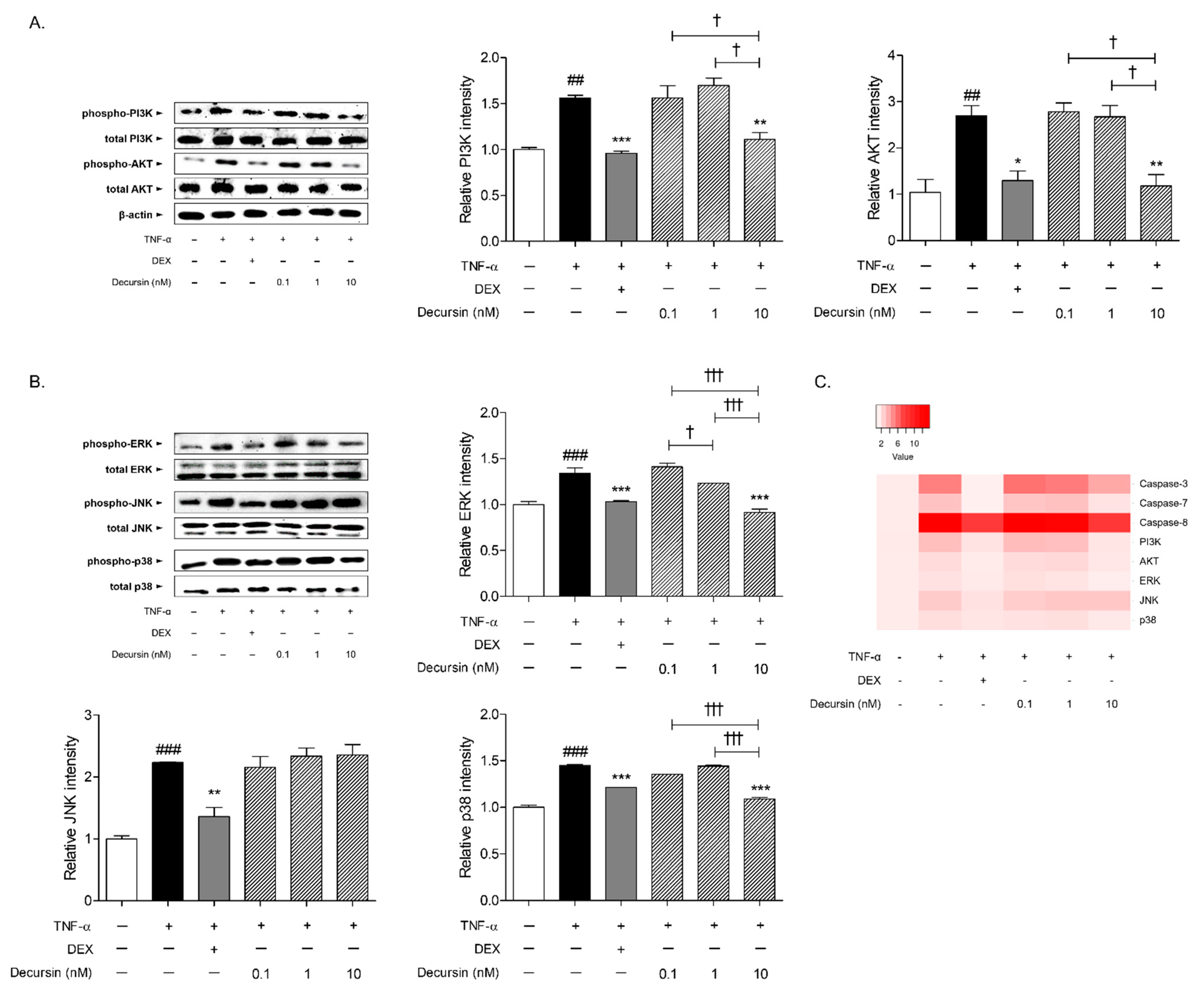
2.6. Decursin Decreased the Expressions of PI3K/AKT Signaling and MAPKs Signaling Pathway in TNF-α-Stimulated Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Network Construction and Comparison of Common Genes between Decursin and Alopecia-Targeted Genes
4.2. Functional Enrichment Analysis
4.3. Animal Experiments
4.4. Skin Monitoring by Digital Dermatoscope
4.5. Histology
4.6. Immunofluorescence
4.7. Cell Treatment
4.8. Immunoblotting Analysis and Heatmap
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, S.; Zhu, Z.; Sun, X.; Fu, X. Functional hair follicle regeneration: An updated review. Signal Transduct. Target. Ther. 2021, 6, 66. [Google Scholar] [CrossRef] [PubMed]
- Al Aboud, A.M.; Zito, P.M. Alopecia. In Treasure Island; StatPearls: Florida, FL, USA, 2021. [Google Scholar]
- Bodó, E.; Tobin, D.J.; Kamenisch, Y.; Bíró, T.; Berneburg, M.; Funk, W.; Paus, R. Dissecting the Impact of Chemotherapy on the Human Hair Follicle: A Pragmatic in Vitro Assay for Studying the Pathogenesis and Potential Management of Hair Follicle Dystrophy. Am. J. Pathol. 2007, 171, 1153–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botchkarev, V.A. Molecular Mechanisms of Chemotherapy-Induced Hair Loss. J. Investig. Dermatol. Symp. Proc. 2003, 8, 72–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehmke, M.M.; Dickerson, S.S. Symptom, Symptom Experiences, and Symptom Distress Encountered by Women With Breast Cancer Undergoing Current Treatment Modalities. Cancer Nurs. 2005, 28, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Mustoe, M.M.; Lee, C.M.; E Melisko, M.; Esserman, L.J.; Rugo, H.S. The DigniCap Scalp Cooling System and its use in the treatment of chemotherapy-induced alopecia. Futur. Oncol. 2018, 14, 2461–2469. [Google Scholar] [CrossRef]
- Dunnill, C.; Ibraheem, K.; Peake, M.; Ioannou, M.; Palmer, M.; Smith, A.; Collett, A.; Georgopoulos, N.T. Cooling-mediated protection from chemotherapy drug-induced cytotoxicity in human keratinocytes by inhibition of cellular drug uptake. PLoS ONE 2020, 15, e0240454. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Caro, G.; Fortuna, M.C.; Pigliacelli, F.; D’Arino, A.; Carlesimo, M. Prevention and Treatment of Chemotherapy-Induced Alopecia. Dermatol. Pr. Concept. 2020, 10, e2020074. [Google Scholar] [CrossRef]
- Rubio-Gonzalez, B.; Juhász, M.; Fortman, J.; Mesinkovska, N.A. Pathogenesis and treatment options for chemotherapy-induced alopecia: A systematic review. Int. J. Dermatol. 2018, 57, 1417–1424. [Google Scholar] [CrossRef]
- Park, B.-S.; Sim, J.-B.; Park, S.-O.; Noh, G.-Y. Effect of Natural Plant Extracts on Hair Loss Prevent in People with Alopecia. Asian J. Dermatol. 2016, 8, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.; Mehta, S.; Thakurdesai, P. Efficacy and safety of standardized cinnamon bark extract for the prevention of chemo-therapy-induced weight loss and alopecia in patients with breast cancer: A randomized, double-blind, and placebo-controlled study. Asian J. Pharm. Clin. Res. 2019, 12, 163–168. [Google Scholar] [CrossRef]
- He, L.; Pan, Y.; Yu, J.; Wang, B.; Dai, G.; Ying, X. Decursin alleviates the aggravation of osteoarthritis via inhibiting PI3K-Akt and NF-kB signal pathway. Int. Immunopharmacol. 2021, 97, 107657. [Google Scholar] [CrossRef]
- Kim, M.H.; Choi, Y.Y.; Cho, I.-H.; Hong, J.; Kim, S.; Yang, W.M. Angelica sinensis Induces Hair Regrowth via the Inhibition of Apoptosis Signaling. Am. J. Chin. Med. 2014, 42, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-K.; Kim, B.; Kim, D.W.; Ahn, J.H.; Sim, H.; Lee, J.-C.; Yang, G.E.; Her, Y.; Park, J.H.; Kim, H.S.; et al. Effects of Decursin and Angelica gigas Nakai Root Extract on Hair Growth in Mouse Dorsal Skin via Regulating Inflammatory Cytokines. Molecules 2020, 25, 3697. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Handjiski, B.; Eichmüller, S.; Czarnetzki, B.M. Chemotherapy-induced alopecia in mice. Induction by cyclophosphamide, inhibition by cyclosporine A, and modulation by dexamethasone. Am. J. Pathol. 1994, 144, 719–734. [Google Scholar] [PubMed]
- Paus, R.; Haslam, I.; A Sharov, A.; Botchkarev, V. Pathobiology of chemotherapy-induced hair loss. Lancet Oncol. 2013, 14, e50–e59. [Google Scholar] [CrossRef]
- Danilenko, D.; Ring, B.D.; Yanagihara, D.; Benson, W.; Wiemann, B.; Starnes, C.O.; Pierce, G.F. Keratinocyte growth factor is an important endogenous mediator of hair follicle growth, development, and differentiation. Normalization of the nu/nu follicular differentiation defect and amelioration of chemotherapy-induced alopecia. Am. J. Pathol. 1995, 147, 145–154. [Google Scholar] [PubMed]
- Katikaneni, R.; Ponnapakkam, T.; Seymour, A.; Sakon, J.; Gensure, R. Parathyroid hormone linked to a collagen binding domain promotes hair growth in a mouse model of chemotherapy-induced alopecia in a dose-dependent manner. Anti-Cancer Drugs 2014, 25, 819–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paik, S.H.; Yoon, J.-S.; Ryu, H.H.; Lee, J.Y.; Shin, C.Y.; Min, K.H.; Jo, S.J.; Kim, K.H.; Kwon, O. Pretreatment of epidermal growth factor promotes primary hair recovery via the dystrophic anagen pathway after chemotherapy-induced alopecia. Exp. Dermatol. 2013, 22, 496–499. [Google Scholar] [CrossRef]
- Yoneda, K.; Fujii, M.; Imaoka, A.; Kobayashi, R.; Hayashi, R.; Yoshida, Y.; Kohno, T.; Tsuji, T. Preventive effect of edaravone ointment on cyclophosphamide-chemotherapy induced alopecia. Support. Care Cancer 2021, 29, 6127–6134. [Google Scholar] [CrossRef]
- Botchkareva, N.V.; Ahluwalia, G.; Shander, D. Apoptosis in the Hair Follicle. J. Investig. Dermatol. 2006, 126, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Hendrix, S.; Handjiski, B.; Peters, E.M.; Paus, R. A Guide to Assessing Damage Response Pathways of the Hair Follicle: Lessons From Cyclophosphamide-Induced Alopecia in Mice. J. Investig. Dermatol. 2005, 125, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.H.; Kim, K.; Lee, H.; Yang, W.M. Human placenta induces hair regrowth in chemotherapy-induced alopecia via inhibition of apoptotic factors and proliferation of hair follicles. BMC Complement. Med. Ther. 2020, 20, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Khanna, N.; Sharma, H.; Kundu, S.; Azmi, S. Insights into the molecular mechanism of apoptosis induced by TNF-α in mouse epidermal JB6-derived RT-101 cells. Biochem. Biophys. Res. Commun. 2002, 295, 24–30. [Google Scholar] [CrossRef]
- Grosbois, J.; Demeestere, I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum. Reprod. 2018, 33, 1705–1714. [Google Scholar] [CrossRef]
- Chen, X.-Y.; Xia, H.-X.; Guan, H.-Y.; Li, B.; Zhang, W. Follicle Loss and Apoptosis in Cyclophosphamide-Treated Mice: What’s the Matter? Int. J. Mol. Sci. 2016, 17, 836. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, S.R.; Choi, Y.-H.; Shin, J.Y.; Kim, C.D.; Kang, N.-G.; Park, B.C.; Lee, S. Quercitrin Stimulates Hair Growth with Enhanced Expression of Growth Factors via Activation of MAPK/CREB Signaling Pathway. Molecules 2020, 25, 4004. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Wu, Y.; Wusiman, A.; Gu, P.; Mao, N.; Xu, S.; Zhu, T.; Feng, Z.; Liu, Z.; Wang, D. Alhagi honey polysaccharides attenuate intestinal injury and immune suppression in cyclophosphamide-induced mice. Food Funct. 2021, 12, 6863–6877. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.K.; Park, S.J.; Kim, M.H.; Choi, L.Y.; Yang, W.M. Pharmacopuncture of Taraxacum platycarpum extract reduces localized fat by regulating the lipolytic pathway. Biomed. Pharmacother. 2021, 141, 111905. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.H.; Park, S.J.; Yang, W.M. Network Pharmacology Study and Experimental Confirmation Revealing the Ameliorative Effects of Decursin on Chemotherapy-Induced Alopecia. Pharmaceuticals 2021, 14, 1150. https://doi.org/10.3390/ph14111150
Kim MH, Park SJ, Yang WM. Network Pharmacology Study and Experimental Confirmation Revealing the Ameliorative Effects of Decursin on Chemotherapy-Induced Alopecia. Pharmaceuticals. 2021; 14(11):1150. https://doi.org/10.3390/ph14111150
Chicago/Turabian StyleKim, Mi Hye, Sang Jun Park, and Woong Mo Yang. 2021. "Network Pharmacology Study and Experimental Confirmation Revealing the Ameliorative Effects of Decursin on Chemotherapy-Induced Alopecia" Pharmaceuticals 14, no. 11: 1150. https://doi.org/10.3390/ph14111150
APA StyleKim, M. H., Park, S. J., & Yang, W. M. (2021). Network Pharmacology Study and Experimental Confirmation Revealing the Ameliorative Effects of Decursin on Chemotherapy-Induced Alopecia. Pharmaceuticals, 14(11), 1150. https://doi.org/10.3390/ph14111150