
The Real Cytotoxic Effect of Artemisinins on Colon Cancer Cells in a Physiological Cell Culture Setting. How Composition of the Culture Medium Biases Experimental Findings

Abstract

1. Introduction
2. Results
2.1. Cytotoxic Activity
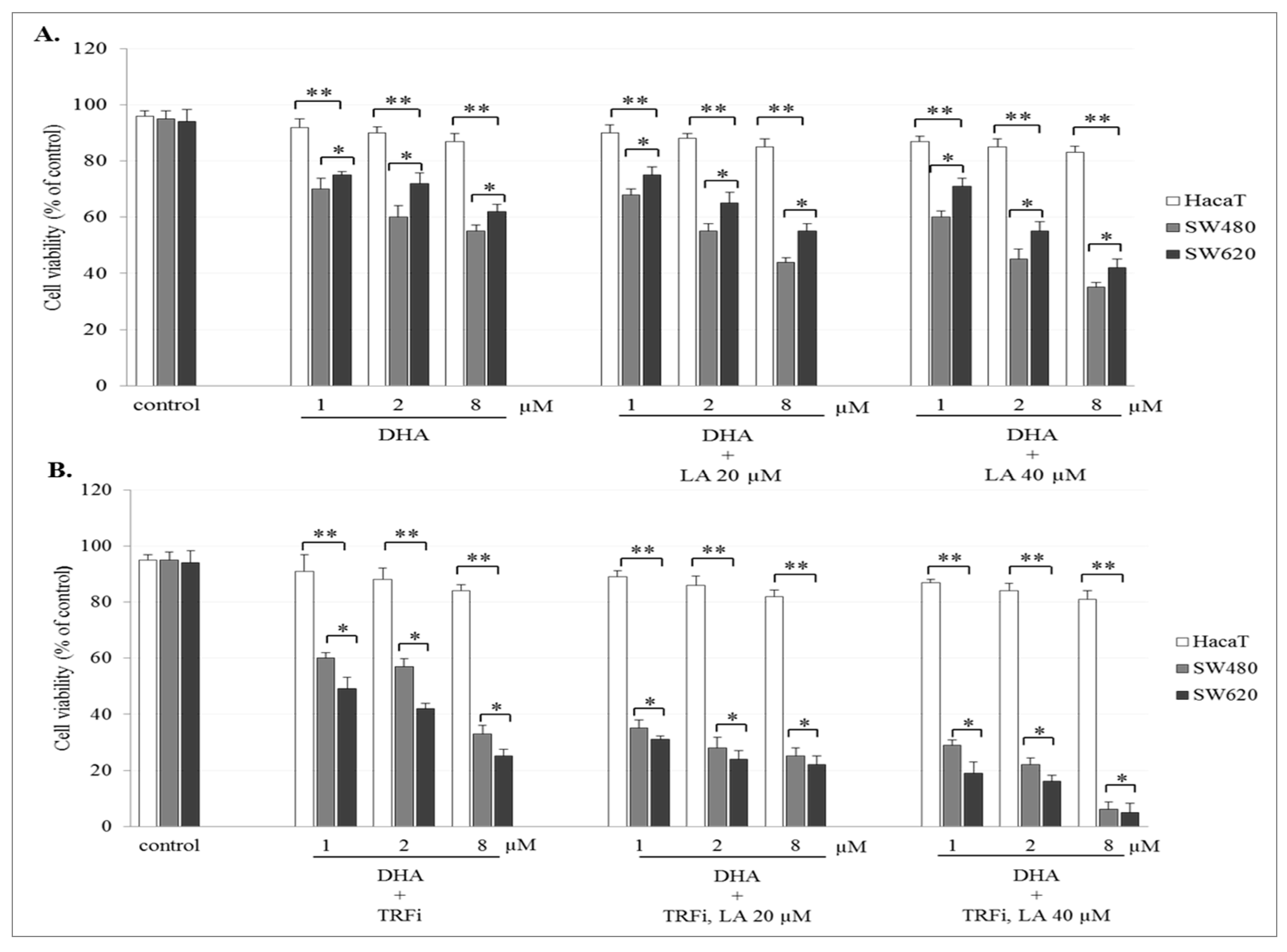
2.2. The Effect of Artemisinin and Dihydroartemisinin on Viability of Human Cancer and HaCaT Cells
2.3. Apoptotic Activity of Artemisinin and Dihydroartemisinin
2.4. IL-6 Level Studies
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Drug Preparation
4.3. Cell Culture and Treatment
4.4. MTT Assay
4.5. Trypan Blue Exclusion Assay
4.6. FITC Annexin V Binding Assay
4.7. Interleukin-6 Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| ART | Artemisinin |
| CRC | Colorectal cancer |
| DHA | Dihydroartemisinin |
| LA | Linoleic acid |
| PUFAs | Polyunsaturated fatty acids |
| ROS | Reactive oxygen species |
| TRFi | Holo-transferrin |
References
- Neubauer, K.; Misa, I.B.; Diakowska, D.; Kapturkiewicz, B.; Gamian, A.; Krzystek-Korpacka, M. Nampt/PBEF/Visfatin upregulation in colorectal tumors, mirrored in normal tissue and whole blood of colorectal cancer patients, is associated with metastasis, hypoxia, IL1 beta, and anemia. BioMed Res. Int. 2015, 523930. [Google Scholar] [CrossRef]
- Sadahiro, S.; Morita, S.; Sasaki, K.; Sakamoto, K.; Ohge, H.; Takahashi, T.; Tsuchiya, T.; Sato, T.; Kondo, K.; Ogata, Y.; et al. Treatment rationale and study design for clinical trial on the efficacy of UFT/LV for Stage II colorectal cancer with risk factors for recurrence (JFMC46-1201). Clin. Color. Cancer 2015, 14, 277–280. [Google Scholar] [CrossRef]
- Ulivi, P.; Scarpi, E.; Passardi, A.; Marisi, G.; Calistri, D.; Zoli, W.; Del Re, M.; Frassineti, G.L.; Tassinari, D.; Tamberi, S.; et al. eNOS polymorphisms as predictors of efficacy of bevacizumab-based chemotherapy in metastatic colorectal cancer: Data from a randomized clinical trial. J. Transl. Med. 2015, 13, 258. [Google Scholar] [CrossRef]
- Iwasa, S.; Nagashima, K.; Yamaguchi, T.; Matsumoto, H.; Ichikawa, Y.; Goto, A.; Yasui, H.; Kato, K.; Okita, N.T.; Shimada, Y.; et al. S-1 and irinotecan with or without bevacizumab versus 5-fluorouracil and leucovorin plus oxaliplatin with or without bevacizumab in metastatic colorectal cancer: A pooled analysis of four phase II studies. Cancer Chemother. Pharmacol. 2015, 76, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, W.L.W.; Liu, L. The Role of Traditional Chinese Herbal Medicines in Cancer Therapy—From TCM Theory to Mechanistic insights. Planta Med. 2010, 76, 1118–1131. [Google Scholar] [CrossRef]
- Keating, G.M. Dihydroartemisinin/Piperaquine: A review of its use in the treatment of uncomplicated plasmodium falciparum malaria. Drugs 2012, 72, 937–961. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Qinghaosu (artemisinin): Chemistry and pharmacology. Acta Pharmacol. Sin. 2012, 33, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Dunstan, H.; Sauerbrey, A.; Miyachi, H.; Chitambar, C. The anti-malarial artesunate is also active against cancer. Int. J. Oncol. 2001, 18, 767–773. [Google Scholar] [CrossRef]
- Handrick, R.; Ontikatze, T.; Bauer, K.-D.; Freier, F.; Rübel, A.; Dürig, J.; Belka, C.; Jendrossek, V. Dihydroartemisinin induces apoptosis by a Bak-dependent intrinsic pathway. Mol. Cancer Ther. 2010, 9, 2497–2510. [Google Scholar] [CrossRef]
- Michaelis, M.; Kleinschmidt, M.C.; Barth, S.; Rothweiler, F.; Geiler, J.; Breitling, R.; Mayer, B.; Deubzer, H.; Witt, O.; Kreuter, J.; et al. Anti-cancer effects of artesunate in a panel of chemoresistant neuroblastoma cell lines. Biochem. Pharmacol. 2010, 79, 130–136. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, P.M.; Barton, V.E.; Ward, S.A. The molecular mechanism of action of artemisinin—The debate continues. Molecules 2010, 15, 1705–1721. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yu, Q.; Huo, C.; Li, Y.; He, L.; Ran, B.; Chen, J.; Li, Y.; Liu, W. Ferroptosis: A Novel Mechanism of Artemisinin and its Derivatives in Cancer Therapy. Curr. Med. Chem. 2020, 28, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Ooko, E.; Saeed, M.E.; Kadioglu, O.; Sarvi, S.; Colak, M.; Elmasaoudi, K.; Janah, R.; Greten, J.; Efferth, T. Artemisinin derivatives induce iron-dependent cell death (ferroptosis) in tumor cells. Phytomedicine 2015, 22, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q.; Benthani, F.A.; Wu, J.; Liang, D.; Bian, Z.-X.; Jiang, X. Artemisinin compounds sensitize cancer cells to ferroptosis by regulating iron homeostasis. Cell Death Differ. 2020, 27, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Li, C.; Li, C.; Wei, L.; Li, L.; Zhang, Y.; Yao, Y.; Gu, X.; Cai, W.; Yang, Z.; et al. The artemisinin derivative artesunate inhibits corneal neovascularization by inducing ROS-dependent apoptosis in vascular endothelial cells. Investig. Opthalmol. Vis. Sci. 2013, 54, 3400–3409. [Google Scholar] [CrossRef]
- Zhang, S.; Gerhard, G.S. Heme activates artemisinin more efficiently than hemin, inorganic iron, or hemoglobin. Bioorg. Med. Chem. 2008, 16, 7853–7861. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Gallis, B.; Taya, M.; Wang, S.; Ho, R.J.Y.; Sasaki, T. pH-Responsive artemisinin derivatives and lipid nanoparticle formulations inhibit growth of breast cancer cells in vitro and induce down-regulation of HER family members. PLoS ONE 2013, 8, e59086. [Google Scholar] [CrossRef]
- Efferth, T.; Benakis, A.; Romero, M.R.; Tomicic, M.; Rauh, R.; Steinbach, D.; Häfer, R.; Stamminger, T.; Oesch, F.; Kaina, B.; et al. Enhancement of cytotoxicity of artemisinins toward cancer cells by ferrous iron. Free Radic. Biol. Med. 2004, 37, 998–1009. [Google Scholar] [CrossRef]
- Xiao, F.; Gao, W.; Wang, X.; Chen, T. Amplification activation loop between caspase-8 and -9 dominates artemisinin-induced apoptosis of ASTC-a-1 cells. Apoptosis 2012, 17, 600–611. [Google Scholar] [CrossRef]
- Mercer, A.E.; Copple, I.; Maggs, J.L.; O’Neill, P.M.; Park, B.K. The role of heme and the mitochondrion in the chemical and molecular mechanisms of mammalian cell death induced by the artemisinin antimalarials. J. Biol. Chem. 2011, 286, 987–996. [Google Scholar] [CrossRef]
- Kwok, J.C.; Richardson, D.R. The iron metabolism of neoplastic cells: Alterations that facilitate proliferation? Crit. Rev. Oncol. Hematol. 2002, 42, 65–78. [Google Scholar] [CrossRef]
- Crespo-Ortiz, M.P.; Wei, M.Q. Antitumor activity of artemisinin and its derivatives: From a well-known antimalarial agent to a potential anticancer drug. J. Biomed. Biotechnol. 2011, 2012, 247597. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.; Sasaki, T.; Singh, N.P. Targeted treatment of cancer with artemisinin and artemisinin-tagged iron-carrying compounds. Expert Opin. Ther. Targets 2005, 9, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.; Liu, R.; Cheng, Y. Artesunate alleviates liver fibrosis by regulating ferroptosis signaling pathway. Biomed. Pharmacother. 2019, 109, 2043–2053. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Monian, P.; Pan, Q.; Zhang, W.; Xiang, J.; Jiang, X. Ferroptosis Is an Autophagic Cell Death Process. Cell Res. 2016, 26, 1021–1032. [Google Scholar] [CrossRef]
- Kagan, V.E.; Mao, G.; Qu, F.; Angeli, J.P.F.; Doll, S.; Croix, C.S.; Dar, H.H.; Liu, B.; Tyurin, V.A.; Ritov, V.B.; et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat. Chem. Biol. 2017, 13, 81–90. [Google Scholar] [CrossRef]
- Yang, W.S.; Kim, K.J.; Gaschler, M.M.; Patel, M.; Shchepinov, M.S.; Stockwell, B.R. Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc. Natl. Acad. Sci. USA 2016, 113, E4966–E4975. [Google Scholar] [CrossRef] [PubMed]
- Sapandowski, A.; Stope, M.B.; Evert, K.; Evert, M.; Zimmermann, U.; Peter, D.; Päge, I.; Burchardt, M.; Schild, L. Cardiolipin composition correlates with prostate cancer cell proliferation. Mol. Cell Biochem. 2015, 410, 175–185. [Google Scholar] [CrossRef]
- Hall, A.J.; Chappell, M.J.; Aston, J.A.; Ward, S.A. Pharmacokinetic modelling of the anti-malarial drug artesunate and its active metabolite dihydroartemisinin. Comput. Methods Programs Biomed. 2013, 112, 1–15. [Google Scholar] [CrossRef][Green Version]
- Mcgready, R.; Phyo, A.P.; Rijken, M.; Tarning, J.; Lindegardh, N.; Hanpithakpon, W.; Than, H.H.; Hlaing, N.; Zin, N.T.; Singhasivanon, P.; et al. Artesunate/dihydroartemisinin pharmacokinetics in acute falciparum malaria in pregnancy: Absorption, bioavailability, disposition and disease effects. Br. J. Clin. Pharmacol. 2012, 73, 467–477. [Google Scholar] [CrossRef]
- Konstat-Korzenny, E.; Ascencio-Aragón, J.A.; Niezen-Lugo, S.; Vázquez-López, R. Artemisinin and its synthetic derivatives as a possible therapy for cancer. Med. Sci. 2018, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Du, M.; Schoen, R.E.; Hoffmeister, M.; Newcomb, P.A.; Berndt, S.I.; Caan, B.; Campbell, P.T.; Chan, A.T.; Chang-Claude, J.; et al. Determining risk of colorectal cancer and starting age of screening based on lifestyle, environmental, and genetic factors. Gastroenterology 2018, 154, 2152–2164.e19. [Google Scholar] [CrossRef] [PubMed]
- Slezakova, S.; Ruda-Kucerova, J. Anticancer activity of artemisinin and its derivatives. Anticancer. Res. 2017, 37, 5995–6003. [Google Scholar] [CrossRef]
- Wang, D.; Zhong, B.; Li, Y.; Liu, X. Dihydroartemisinin increases apoptosis of colon cancer cells through targeting Janus kinase 2/signal transducer and activator of transcription 3 signaling. Oncol. Lett. 2017, 15, 1949–1954. [Google Scholar] [CrossRef]
- Kumari, K.; Keshari, S.; Sengupta, D.; Sabat, S.C.; Mishra, S.K. Transcriptome analysis of genes associated with breast cancer cell motility in response to artemisinin treatment. BMC Cancer 2017, 17, 858. [Google Scholar] [CrossRef]
- Lu, Z.-H.; Peng, J.-H.; Zhang, R.-X.; Wang, F.; Sun, H.-P.; Fang, Y.-J.; Wan, D.-S.; Pan, Z.-Z. Dihydroartemisinin inhibits colon cancer cell viability by inducing apoptosis through up-regulation of PPARγ expression. Saudi J. Biol. Sci. 2018, 25, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Sun, L.; Zhou, J.; Yang, J. Dihydroartemisinin induces apoptosis in colorectal cancer cells through the mitochondria-dependent pathway. Tumor Biol. 2014, 35, 5307–5314. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Traer, E.; Zimmerman, G.A.; McIntyre, T.M.; Prescott, S.M. Cloning, expression, and chromosomal localization of human long-chain Fatty Acid-CoA Ligase 4 (FACL4). Genomics 1998, 49, 327–330. [Google Scholar] [CrossRef]
- Doll, S.; Proneth, B.; Tyurina, Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, M.I.J.; Aichler, M.; Walch, M.A.A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Magtanong, L.; Ko, P.-J.; To, M.; Cao, J.Y.; Forcina, G.C.; Tarangelo, A.; Ward, C.C.; Cho, K.; Patti, G.J.; Nomura, D.K.; et al. Exogenous monounsaturated fatty acids promote a ferroptosis-resistant cell state. Cell Chem. Biol. 2019, 26, 420–432. [Google Scholar] [CrossRef]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef]
- Schwab, U.S.; Callaway, J.C.; Erkkilä, A.T.; Gynther, J.; Uusitupa, M.I.J.; Järvinen, T. Effects of hempseed and flaxseed oils on the profile of serum lipids, serum total and lipoprotein lipid concentrations and haemostatic factors. Eur. J. Nutr. 2006, 45, 470–477. [Google Scholar] [CrossRef]
- Mielczarek-Puta, M.; Otto-Ślusarczyk, D.; Chrzanowska, A.; Filipek, A.; Graboń, W. Telmisartan influences the antiproliferative activity of linoleic acid in human colon cancer cells. Nutr. Cancer 2019, 72, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.R.; Liu, Z.X.; Liu, F.; Pan, L.; Yu, H.P.; Jiang, J.P.; Zhang, J.J.; Liu, L.; Yu, J. Holotransferrin enhances selective anticancer activity of artemisinin against human hepatocellular carcinoma cells. J. Huazhong Univ. Sciand. Technol. [Med. Sci.] 2013, 33, 862–865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ji, Y.; Chen, Q.; Jiao, X.; Hou, L.; Zhu, X.; Zhang, Z. Enhancement of cytotoxicity of artemisinin toward cancer cells by transferrin-mediated carbon nanotubes nanoparticles. J. Drug Target 2015, 23, 552–567. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Yang, P.; Zeng, X.; Wang, H.; Cai, H.; Cai, J. Visual characterization of targeted effect of holo-transferrin-tagged dihydroartemisinin on human breast cancer cells. Chin. Sci. Bull. 2010, 55, 2390–2395. [Google Scholar] [CrossRef]
- Jia, J.; Qin, Y.; Zhang, L.; Guo, C.; Wang, Y.; Yue, X.; Qian, J. Artemisinin inhibits gallbladder cancer cell lines through triggering cell cycle arrest and apoptosis. Mol. Med. Rep. 2016, 13, 4461–4468. [Google Scholar] [CrossRef]
- Kim, S.H.; Kang, S.H.; Kang, B.S. Therapeutic effects of dihydroartemisinin and transferrin against glioblastoma. Nutr. Res. Pr. 2016, 10, 393–397. [Google Scholar] [CrossRef]
- Culig, Z. Proinflammatory cytokine interleukin-6 in prostate carcinogenesis. Am. J. Clin. Exp. Urol. 2014, 2, 231–238. [Google Scholar]
- Culig, Z.; Puhr, M. Interleukin-6 and prostate cancer: Current developments and unsolved questions. Mol. Cell. Endocrinol. 2018, 462, 25–30. [Google Scholar] [CrossRef]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumor Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.C.; Hobisch, A.; Lin, D.-L.; Culig, Z.; Keller, E.T. Interleukin-6 and prostate cancer progression. Cytokine Growth Factor Rev. 2001, 12, 33–40. [Google Scholar] [CrossRef]
- Brookes, M.J.; Hughes, S.; Turner, F.E.; Reynolds, G.; Sharma, N.; Ismail, T.; Berx, G.; McKie, A.T.; Hotchin, N.; Anderson, G.J.; et al. Modulation of iron transport proteins in human colorectal carcinogenesis. Gut 2006, 55, 1449–1460. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Cancer Cells | Normal Cells | |||
|---|---|---|---|---|---|
| SW480 d | SW620 e | HaCaT f | |||
| IC50 b | SI c | IC50 | SI | IC50 | |
| ART | 39 ± 2.75 | 5.2 | 42.8 ± 4.6 | 4.7 | 203 ± 16.2 |
| ART-LA 20 | 17.5 ± 0.4 | 10.9 | 21.7 ± 7.3 | 8.8 | 191 ± 2.83 |
| ART-LA 40 | 8.9 ± 3.1 | 16.4 | 18.4 ± 2.6 | 7.9 | 146.5 ± 6.38 |
| ART-TRFi | 4.58 ± 0.2 | 40.8 | 16.3 ± 6.1 | 11.6 | 187.1 ± 5.56 |
| ART-LA 20-TRFi | 2.36 ± 0.1 | 82.6 | 7.4 ± 0.11 | 26.1 | 195 ± 7.2 |
| ART-LA 40-TRFi | 0.69 ± 0.01 | 202 | 0.6 ± 0.04 | 225 | 142 ± 3.5 |
| DHA | 11.4 ± 1.67 | 10.1 | 11.9 ± 2.3 | 9.8 | 116 ± 5.5 |
| DHA-LA 20 | 8.6 ± 0.9 | 16.1 | 9.8 ± 1.2 | 14.1 | 138 ± 3.1 |
| DHA-LA 40 | 3.6 ± 0.3 | 13.8 | 1.27 ± 0.25 | 39.8 | 50.4 ± 3.4 |
| DHA-TRFi | 1.64 ± 0.07 | 78.7 | 10.3 ± 6.2 | 12.4 | 129.2 ± 0.5 |
| DHA-LA 20-TRFi | 0.38 ± 0.09 | 273 | 8.4 ± 0.9 | 12.3 | 104.4 ± 9.8 |
| DHA-LA 40-TRFi | 0.24 ± 0.02 | 288 | 0.14 ± 0.01 | 480 | 72.2 ± 1.6 |
| DOX g | 0.75 ± 0.1 | 0.38 | 0.26 ± 0.1 | 1.11 | 0.29 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otto-Ślusarczyk, D.; Mielczarek-Puta, M.; Graboń, W. The Real Cytotoxic Effect of Artemisinins on Colon Cancer Cells in a Physiological Cell Culture Setting. How Composition of the Culture Medium Biases Experimental Findings. Pharmaceuticals 2021, 14, 976. https://doi.org/10.3390/ph14100976
Otto-Ślusarczyk D, Mielczarek-Puta M, Graboń W. The Real Cytotoxic Effect of Artemisinins on Colon Cancer Cells in a Physiological Cell Culture Setting. How Composition of the Culture Medium Biases Experimental Findings. Pharmaceuticals. 2021; 14(10):976. https://doi.org/10.3390/ph14100976
Chicago/Turabian StyleOtto-Ślusarczyk, Dagmara, Magdalena Mielczarek-Puta, and Wojciech Graboń. 2021. "The Real Cytotoxic Effect of Artemisinins on Colon Cancer Cells in a Physiological Cell Culture Setting. How Composition of the Culture Medium Biases Experimental Findings" Pharmaceuticals 14, no. 10: 976. https://doi.org/10.3390/ph14100976
APA StyleOtto-Ślusarczyk, D., Mielczarek-Puta, M., & Graboń, W. (2021). The Real Cytotoxic Effect of Artemisinins on Colon Cancer Cells in a Physiological Cell Culture Setting. How Composition of the Culture Medium Biases Experimental Findings. Pharmaceuticals, 14(10), 976. https://doi.org/10.3390/ph14100976