Development of Rapid Detection and Genetic Characterization of Salmonella in Poultry Breeder Feeds

Abstract
:1. Introduction
2. Salmonella in Broiler Breeders
3. Feed
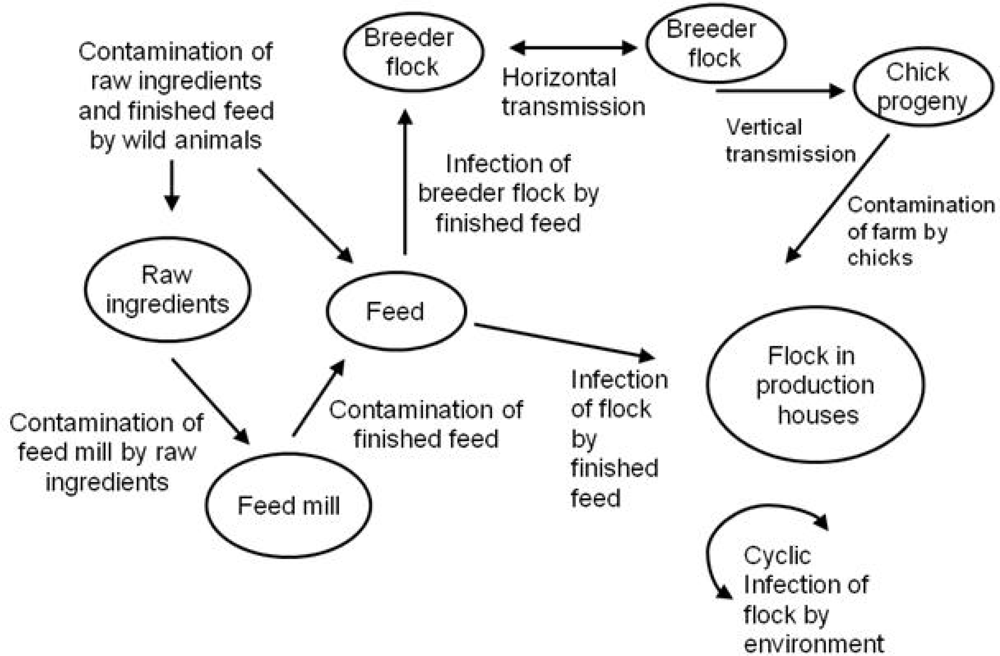
3.1. Sources of Salmonella to Feed
3.2. Processing of Feed
4. Detection
4.1. General Concepts
4.2. Culturing
4.3. Serology and Immunoassays
4.4. Nucleic Acid Detection
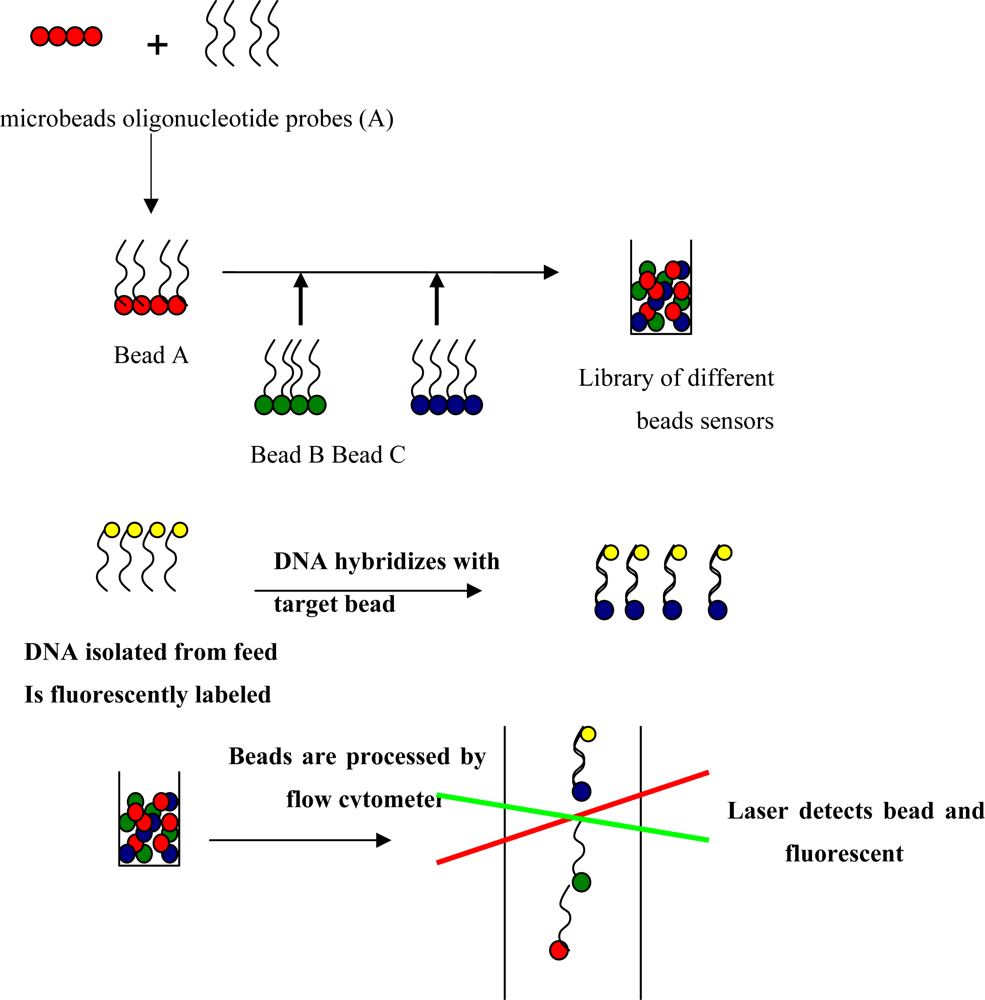
4.5. Biosensors
5. Future prospects
6. Conclusions
Acknowledgments
References
- Braden, C.R. Salmonella enterica serotype enteritidis and eggs: a national epidemic in the United States. Clin. Infect. Dis 2006, 43, 512–517. [Google Scholar]
- Linam, W.M.; Gerber, M.A. Changing epidemiology and prevention of Salmonella infections. Pediatr. Infect. Dis. J 2007, 26, 747–748. [Google Scholar]
- Bhatia, T.R.; McNabb, G.D. Dissemination of Salmonella in broiler-chicken operations. Avian Dis 1980, 24, 616–624. [Google Scholar]
- Cox, N.A.; Bailey, J.S.; Mauldin, J.M.; Blankenship, L.C.; Wilson, J.L. Extent of salmonellae contamination in breeder hatcheries. Poult. Sci 1991, 70, 416–418. [Google Scholar]
- Milner, K.C.; Shaffer, M.F. Bacteriologic studies of experimental Salmonella infections in chicks. J. Infect. Dis 1952, 90, 81–96. [Google Scholar]
- Bailey, J.S.; Cox, N.A.; Berrang, M.E. Hatchery-acquired salmonellae in broiler chicks. Poult. Sci 1994, 73, 1153–1157. [Google Scholar]
- Jones, F.T.; Axtell, R.C.; Rives, D.V.; Scheideler, S.E.; Tarver, F.R.; Walker, R.L.; Wineland, M.J. A survey of Salmonella contamination in modern broiler production. J. Food Prot 1991, 54, 502–507. [Google Scholar]
- Bailey, J.S.; Mitchell, B.W.; Buhr, R.J.; Berrang, M.E.; Cox, N.A. Reducing airborne pathogens, dust and Salmonella transmission in experimental hatching cabinets using an electrostatic space charge system. Poult. Sci 2002, 81, 49–55. [Google Scholar]
- Byrd, J.A.; DeLoach, J.R.; Corrier, D.E.; Nisbet, D.J.; Stanker, L.H. Evaluation of Salmonella serotype distributions from commercial broiler hatcheries and grower houses. Avian Dis 1999, 43, 39–47. [Google Scholar]
- Davies, R.H.; Wray, C. Persistence of Salmonella enteritidis in poultry units and poultry food. Br. Poult. Sci 1996, 37, 589–596. [Google Scholar]
- Davies, R.H.; Wray, C. Determination of an effective sampling regime to detect Salmonella enteritidis in the environment of poultry units. Vet. Microbiol 1996, 50, 117–127. [Google Scholar]
- Bailey, J.S.; Craven, S.E.; Stern, N.J.; Line, E.; Cox, N.A.; Fedorka-Cray, P. Determination of the incidence of Salmonella spp., Campylobacter jejuni, and Clostridium perfringens in wild birds near broiler chicken houses by sampling intestinal droppings. Avian Dis 2000, 44, 715–720. [Google Scholar]
- Blankenship, L.C.; Bailey, J.S.; Cox, N.A.; Stern, N.J.; Brewer, R.; Williams, O. Two-step mucosal competitive exclusion flora treatment to diminish salmonellae in commercial broiler chickens. Poult. Sci 1993, 72, 1667–1672. [Google Scholar]
- Lahellec, C.; Colin, P.; Bennejean, G.; Paquin, J.; Guillermo, A.; Debois, J.C. Influence of resident Salmonella on contamination of broiler flocks. Poult. Sci 1986, 65, 2034–2039. [Google Scholar]
- Olsen, A.R.; Hammack, T.S. Isolation of Salmonella spp. from the housefly, Musca domestica L., and the dump fly, Hydrotaea aenescens (Wiedemann) (Diptera: Muscidae), at caged-layer houses. J. Food Protect 2000, 63, 958–950. [Google Scholar]
- Maciorowski, K.G.; Herrera, P.; Kundinger, M.M.; Ricke, S.C. Animal production and contamination by foodborne Salmonella. J. Consumer Prot. Food Saf 2006, 1, 197–209. [Google Scholar]
- Cox, N.A.; Bailey, J.S.; Mauldin, J.M.; Blankenship, L.C. Presence and impact of Salmonella contamination in commercial broiler hatcheries. Poult. Sci 1990, 69, 1606–1609. [Google Scholar]
- Veldman, A.; Vahl, H.A.; Borggreve, G.J.; Fuller, D.C. A survey of the incidence of Salmonella species and Enterobacteriaceae in poultry feeds and feed components. Vet. Rec 1995, 136, 169–172. [Google Scholar]
- Jones, F.T.; Richardson, K.E. Salmonella in commercially manufactured feeds. Poult. Sci 2004, 83, 384–391. [Google Scholar]
- Hinton, M. Salmonella infection in chicks following the consumption of artificially contaminated feed. Epidemiol. Infect 1988, 100, 247–256. [Google Scholar]
- Alvarez, J.; Porwollik, S.; Laconcha, I.; Gisakis, V.; Vivanco, A.; Gonzalez, I.; Echenagusia, S.; Zabala, N.; Blackmer, F.; McClelland, M.; Rementeria, A.; Garaizar, J. Detection of a Salmonella enterica serovar California strain spreading in Spanish feed mills and genetic characterization with DNA microarrays. Appl. Environ. Microbiol 2003, 69, 7531–7354. [Google Scholar]
- Davies, R.; Wray, C. Distribution of Salmonella contamination in ten animal feedmills. Vet Microbiol 1997, 57, 159–169. [Google Scholar]
- Erwin, L.E. Examination of prepared poultry feeds for the presence Salmonella and other enteric organisms. Poult. Sci 1955, 34, 215–216. [Google Scholar]
- Cox, N.A.; Bailey, J.S.; Thomson, J.E.; Juven, B.J. Salmonella and other Enterobacteriaceae found in commercial poultry feed. Poult. Sci 1983, 62, 2169–2175. [Google Scholar]
- Morris, G.K.; McMurray, B.L.; Galton, M.M.; Wells, J.G. A study of the dissemination of salmonellosis in a commercial broiler chicken operation. Am. J. Vet. Res 1969, 30, 1413–1421. [Google Scholar]
- Hirsch, W.; Sapiro-Hirsch, R. The role of certain animal feeding stuffs especially bone meal, in the epidemiology of salmonellosis. Harefuah 1958, 54, 57–59. [Google Scholar]
- Semple, A.B.; Turner, G.C.; Lowry, D.M. Outbreak of food-poisoning caused by Salmonella Virchow in spit-roasted chicken. Br. Med. J 1968, 4, 801–803. [Google Scholar]
- Pennington, J.H.; Brooksbank, N.H.; Poole, P.M.; Seymour, F. Salmonella virchow in a chicken-packing station and associated rearing units. Br. Med. J 1968, 4, 804–806. [Google Scholar]
- Clark, G.M.; Kaufmann, A.F.; Gangarosa, E.J.; Thompson, M.A. Epidemiology of an international outbreak of Salmonella Agona. Lancet 1973, 2, 490–493. [Google Scholar]
- Shirota, K.; Katoh, H.; Murase, T.; Ito, T.; Otsuki, K. Monitoring of layer feed and eggs for Salmonella in eastern Japan between 1993 and 1998. J. Food Prot 2001, 64, 734–737. [Google Scholar]
- Wasyl, D.; Sandvang, D.; Skov, M.N.; Baggesen, D.L. Epidemiological characteristics of Salmonella typhimurium isolated from animals and feed in Poland. Epidemiol. Infect 2006, 134, 179–185. [Google Scholar]
- Bucher, O.; Holley, R.A.; Ahmed, R.; Tabor, H.; Nadon, C.; Ng, L.K.; D’Aoust, J.Y. Occurrence and characterization of Salmonella from chicken nuggets, strips, and pelleted broiler feed. J. Food Prot 2007, 70, 2251–2258. [Google Scholar]
- Leeson, S. Nutrition & Health: Poultry. Available online: http://fdsmagissues.feedstuffs.com/fds/Reference_issue_2009/Section6_2008.pdf/.
- Himanthonkham, S.; das Gracas Periera, M.; Riemann, H. Heat destruction of Salmonella in poultry feeds. Avian Dis 1996, 40, 72–77. [Google Scholar]
- Bastianelli, D.; Lebas, C. Evaluating the Role of Animal Feed in Food Safety: Perspectives for Action. 2002. Proceedings of the International Workshop of Food Safety Management in Developing Countries, Montpellier, France, 11–13 December, 2000.
- Riley, P.B. Salmonella Infection: The Position of Animal Food and Its Manufacturing Process; Taylor, J., Ed.; Royal Society of Health: London, UK, 1969; pp. 101–115. [Google Scholar]
- Zdragas, A.; Tsakos, P.; Mavrogeni, P. Evaluation of two assays, MSRV and RV, for the isolation of Salmonella spp. from wastewater samples and broiler chickens. Lett. Appl. Microbiol 2000, 31, 328–331. [Google Scholar]
- Carrique-Mas, J.J.; Davies, R.H. Sampling and bacteriological detection of Salmonella in poultry and poultry premises: A review. Rev. Sci. Tech 2008, 27, 665–677. [Google Scholar]
- Williams, J.E. Salmonellas in poultry feeds-A worldwide review. Part III. Methods in control and elimination. World’s Poult. Sci. J 1981, 37, 97–105. [Google Scholar]
- Maciorowski, K.G.; Herrera, P.; Jones, F.T.; Pillai, S.D.; Ricke, S.C. Cultural and immunological detection methods for Salmonella spp. in animal feeds- A review. Vet. Res. Comm 2006, 30, 127–137. [Google Scholar]
- Nye, K.J.; Fallon, D.; Frodsham, D.; Gee, B.; Graham, C.; Howe, S.; Messer, S.; Turner, T.; Warren, R.E. An evaluation of the performance of XLD, DCA, MLCB, and ABC agars as direct plating media for the isolation of Salmonella enterica from faeces. J. Clin. Pathol 2002, 55, 286–288. [Google Scholar]
- Perez, J.M.; Cavalli, P.; Roure, C.; Renac, R.; Gille, Y; Freydiere, A.M. Comparison of four chromogenic media and Hektoen agar for detection and presumptive identification of Salmonella strains in human stools. J. Clin. Microbiol 2003, 41, 1130–1134. [Google Scholar]
- Holt, P.S.; Mitchell, B.W.; Gast, R.K. Airborne horizontal transmission of Salmonella enteritidis in molted laying chickens. Avian Dis 1998, 42, 45–52. [Google Scholar]
- Reeves, M.W.; Evins, G.M.; Heiba, A.A.; Plikaytis, B.D.; Farmer, J.J. Clonal nature of Salmonella typhi and its genetic relatedness to other salmonellae as shown by multilocus enzyme electrophoresis, and proposal of Salmonella bongori comb. J. Clin. Microbiol 1989, 27, 313–320. [Google Scholar]
- Grimont, P.; Weill, F. Antigenic formulas of the Salmonella serovars, 9th ed.; (2007 with supplements). WHO Collaborating Centre for Reference and Research on Salmonella: Paris, France, 2007. Available online: http://www.pasteur.fr/sante/clre/cadrecnr/salmoms/WKLM_2007.pdf/.
- Schwan, W.R.; Huang, X.Z.; Hu, L.; Kopecko, D.J. Differential bacterial survival, replication, and apoptosis-inducing ability of Salmonella serovars within human and murine macrophages. Infect. Immun 2000, 68, 1005–1013. [Google Scholar]
- Okamura, M.; Kamijima, Y.; Miyamoto, T.; Tani, H.; Sasai, K.; Baba, E. Differences among six Salmonella serovars in abilities to colonize reproductive organs and to contaminate eggs in laying hens. Avian Dis 2001, 45, 61–69. [Google Scholar]
- Okamura, M.; Miyamoto, T.; Kamijima, Y.; Tani, H.; Sasai, K.; Baba, E. Differences in abilities to colonize reproductive organs and to contaminate eggs in intravaginally inoculated hens and in vitro adherences to vaginal explants between Salmonella Entertidis and other serovars. Avian Dis 2001, 45, 962–971. [Google Scholar]
- Pedersen, T.B.; Olsen, J.E.; Bisgaard, M. Persistence of Salmonella Senftenberg in poultry production environments and investigation of its resistance to desiccation. Avian Path 2008, 37, 421–427. [Google Scholar]
- Smith, B.P.; Dilling, G.W.; House, J.K.; Konrad, H.; Moore, N. Enzyme-linked immunosorbent assay for Salmonella serology using lipopolysaccharide antigen. J. Vet. Diagnos. Invest 1995, 7, 481–487. [Google Scholar]
- Luk, J.M.; Lindberg, A.A. Rapid and sensitive detection of Salmonella (O:6,7) by immunomagnetic monoclonal antibody-based assays. J. Immunol. Meth 1991, 137, 1–8. [Google Scholar]
- Gehring, A.G.; Crawford, C.G.; Mazenko, R.S.; van Houten, L.J.; Brewster, J.D. Enzyme-linked immunomagnetic electrochemical detection of Salmonella typhimurium. J. Immunol. Meth 1996, 195, 15–25. [Google Scholar]
- Hill, W.E. The polymerase chain reaction: applications for the detection of foodborne pathogens. Crit. Rev. Food Sci. Nutrit 1996, 36, 123–173. [Google Scholar]
- Lampel, K.A.; Orlandi, P.A.; Kornegay, L. Improved template preparation for PCR-based assays for detection of food-borne bacterial pathogens. Appl. Environ. Microbiol 2000, 66, 4539–4542. [Google Scholar]
- Wells, J.G.; Wachsmuth, I.K.; Kiehlbauch, J.A.; Bopp, C.A.; Cameron, D.N.; Strockbine, N.A.; Blake, P.A. The use of plasmid profiles and nucleic acid probes in epidemiologic investigations of foodborne, diarrheal diseases. Int. J. Food Microbiol 1991, 12, 77–89. [Google Scholar]
- Wang, L.; Reeves, P.R.; Jensen, S.; Hallman, R. Expression of the O antigen gene cluster is regulated by RfaH through the JUMPstart sequence. FEMS Microbiol. Lett 1998, 165, 201–206. [Google Scholar]
- Leader, B.T.; Frye, J.G.; Hu, J.; Fedorka-Cray, P.J.; Boyle, D.S. High-throughput molecular determination of Salmonella enterica serovars by use of multiplex PCR and capillary electrophoresis analysis. J. Clin. Microbiol 2009, 47, 1290–1299. [Google Scholar]
- USDA-FSIS. Serotypes Profile of Salmonella Isolates from Meat and Poultry Products, January 1998 through December 2007; United States Department of Agriculture: Washington, DC, USA, 7 August 2008. Available online: http://www.fsis.usda.gov/Science/Serotypes_Profile_Salmonella_Isolates/index.asp/.
- Lazcka, O.; Del Campob, F.J.; Munoz, F.X. Pathogen detection: A perspective of traditional methods and biosensors. Biosens. Bioelectron 2007, 22, 1205–1217. [Google Scholar]
- Olsen, E.V.; Sorokulova, I.B.; Petrenko, V.A.; Chen, I.H.; Barbaree, J.M.; Vodyanoy, V.J. Affinity-selected filamentous bacteriophage as a probe for acoustic wave biodetectors of Salmonella typhimurium. Biosens. Bioelectron 2006, 21, 1434–1442. [Google Scholar]
- Su, X.L.; Li, Y. A QCM immunosensor for Salmonella detection with simultaneous measurements of resonant frequency and motional resistance. Biosens. Bioelectron 2005, 21, 840–848. [Google Scholar]
- Pathirana, S.T.; Barbaree, J.; Chin, B.A.; Hartell, M.G.; Neely, W.C.; Vodyanoy, V. Rapid and sensitive biosensor for Salmonella. Biosens. Bioelectron 2000, 15, 135–141. [Google Scholar]
- Kim, G.H.; Rand, A.G.; Letcher, S.V. Impedance characterization of a piezoelectric immunosensor part II: Salmonella typhimurium detection using magnetic enhancement. Biosens. Bioelectron 2003, 18, 91–99. [Google Scholar]
- Liu, Y.; Fratamico, P. Escherichia coli O antigen typing using DNA microarrays. Mol. Cell. Probes 2006, 20, 239–244. [Google Scholar]
- Malorny, B.; Bunge, C.; Guerra, B.; Prietz, S.; Helmuth, R. Molecular characterization of Salmonella strains by an oligonucleotide multiprobe microarray. Mol. Cell. Probes 2007, 21, 56–65. [Google Scholar]
- Call, D.R.; Borucki, M.K.; Loge, F.J. Detection of bacterial pathogens in environmental samples using DNA microarrays. J. Microbiol. Methods 2003, 53, 235–243. [Google Scholar]
- Abee, T.; van Schaik, W.; Siezen, R.J. Impact of genomics on microbial food safety. Trends Biotech 2004, 22, 653–660. [Google Scholar]
- Call, D.R. Challenges and opportunities for pathogen detection using DNA microarrays. Crit. Rev. Microbiol 2005, 31, 91–99. [Google Scholar]
- Joos, T.O.; Stoll, D.; Templin, M.F. Miniaturised multiplexed immunoassays. Curr. Opin. Chem. Biol 2002, 6, 76–80. [Google Scholar]
- Gonzalez-Buitrago, J.M. Multiplexed testing in the autoimmunity laboratory. Clin. Chem. Lab. Med 2006, 44, 1169–1174. [Google Scholar]
- Dunbar, S.A.; Vander Zee, C.A.; Oliver, K.G.; Karem, K.L.; Jacobson, J.W. Quantitative, multiplexed detection of bacterial pathogens: DNA and protein applications of the Luminex LabMAP system. J. Microbiol. Meth 2003, 53, 245–252. [Google Scholar]
- Fulton, R.J.; McDade, R.L.; Smith, P.L.; Kienker, L.J.; Kettman, J.R. Advanced multiplexed analysis with the FlowMetrix system. Clin. Chem 1997, 43, 1749–1756. [Google Scholar]
- Carson, R.T.; Vignali, D.A. Simultaneous quantitation of 15 cytokines using a multiplexed flow cytometric assay. J. Immunol. Meth 1999, 227, 41–52. [Google Scholar]
- Chen, R.; Lowe, L.; Wilson, J.D.; Crowther, E.; Tzeggai, K.; Bishop, J.E.; Varro, R. Simultaneous quantification of six human cytokines in a single sample using microparticle-based flow cytometric technology. Clin. Chem 1999, 45, 1693–1694. [Google Scholar]
- Bellisario, R.; Colinas, R.J.; Pass, K.A. Simultaneous measurement of antibodies to three HIV-1 antigens in newborn dried blood-spot specimens using a multiplexed microsphere-based immunoassay. Early Hum. Dev 2001, 64, 21–25. [Google Scholar]
- Dunkley, K.D.; Dunkley, C.S.; McReynolds, J.L.; Hume, M.E.; Callaway, T.R.; Kubena, L.F.; Nisbet, D.J.; Ricke, S.C. Molting in Salmonella enteritidis-challenged laying hens fed alfalfa crumbles. II. Fermentation and microbial ecology response. Poul. Sci 2007, 86, 2101–2109. [Google Scholar]
- Swedish Zoonosis Center. Trends and sources of zoonotic infections recorded in Sweden during 2000; Swedish National Veterinary Institute: Uppsala, Sweden, 2001. [Google Scholar]
- Hopp, P.; Wahlstrom, H.; Hirn, J. A common Salmonella control programme in Finland, Norway, and Sweden. Acta. Vet. Scand 1999, 91, 45–49. [Google Scholar]
- Jones, F.T.; Ricke, S.C. Researchers propose tentative HACCP plan for feed manufacturers. Feedstuffs 1994, 66, 35–42. [Google Scholar]
- Hamilton, P.B. Proof of mycotoxicoses being a filed problem and a simple method for their control. Poult. Sci 1975, 54, 1706–1708. [Google Scholar]
- Hess, J. Feed system sanitation said worth the effort. Poul. Dig 1994, 53, 46–47. [Google Scholar]
- Best, P. The clean feed challenge. Feed Mngmt 1994, 45, 27–28. [Google Scholar]
- Kwon, Y.M.; Woodward, C.L.; Pillai, S.D.; Peña, J.; Corrier, D.E.; Byrd, J.A.; Ricke, S.C. Litter and aerosol sampling of chicken houses for rapid detection of Salmonella typhimurium using gene amplification. J. Ind. Microbiol. Biotech 2000, 24, 379–382. [Google Scholar]



{kind=link}
{kind=link}
|
© 2009 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jarquin, R.; Hanning, I.; Ahn, S.; Ricke, S.C. Development of Rapid Detection and Genetic Characterization of Salmonella in Poultry Breeder Feeds. Sensors 2009, 9, 5308-5323. https://doi.org/10.3390/s90705308
Jarquin R, Hanning I, Ahn S, Ricke SC. Development of Rapid Detection and Genetic Characterization of Salmonella in Poultry Breeder Feeds. Sensors. 2009; 9(7):5308-5323. https://doi.org/10.3390/s90705308
Chicago/Turabian StyleJarquin, Robin, Irene Hanning, Soohyoun Ahn, and Steven C. Ricke. 2009. "Development of Rapid Detection and Genetic Characterization of Salmonella in Poultry Breeder Feeds" Sensors 9, no. 7: 5308-5323. https://doi.org/10.3390/s90705308
APA StyleJarquin, R., Hanning, I., Ahn, S., & Ricke, S. C. (2009). Development of Rapid Detection and Genetic Characterization of Salmonella in Poultry Breeder Feeds. Sensors, 9(7), 5308-5323. https://doi.org/10.3390/s90705308