
Calcium Imaging Characterize the Neurobiological Effect of Terahertz Radiation in Zebrafish Larvae


Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry and Embryo Collection
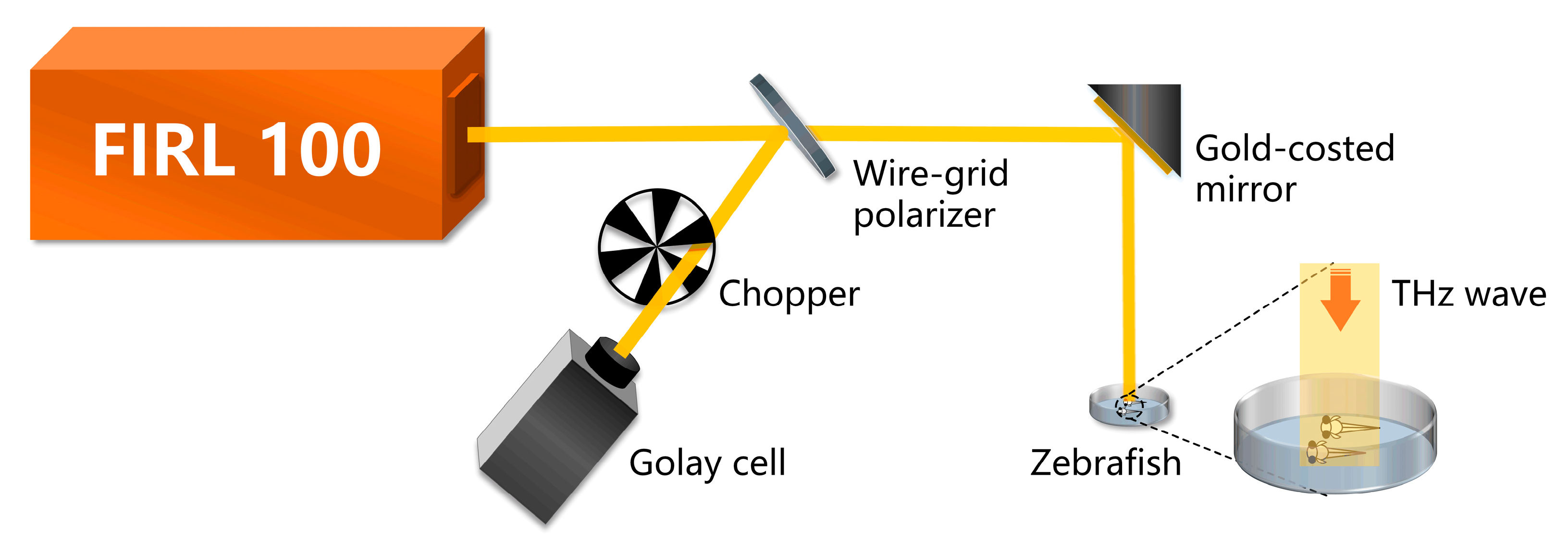
2.2. THz Radiation and Laser Confocal Imaging
2.3. Temperature Measurements
2.4. Zebrafish Behavior Experiment
2.5. qPCR Experiment
2.6. Statistical Analysis
3. Results
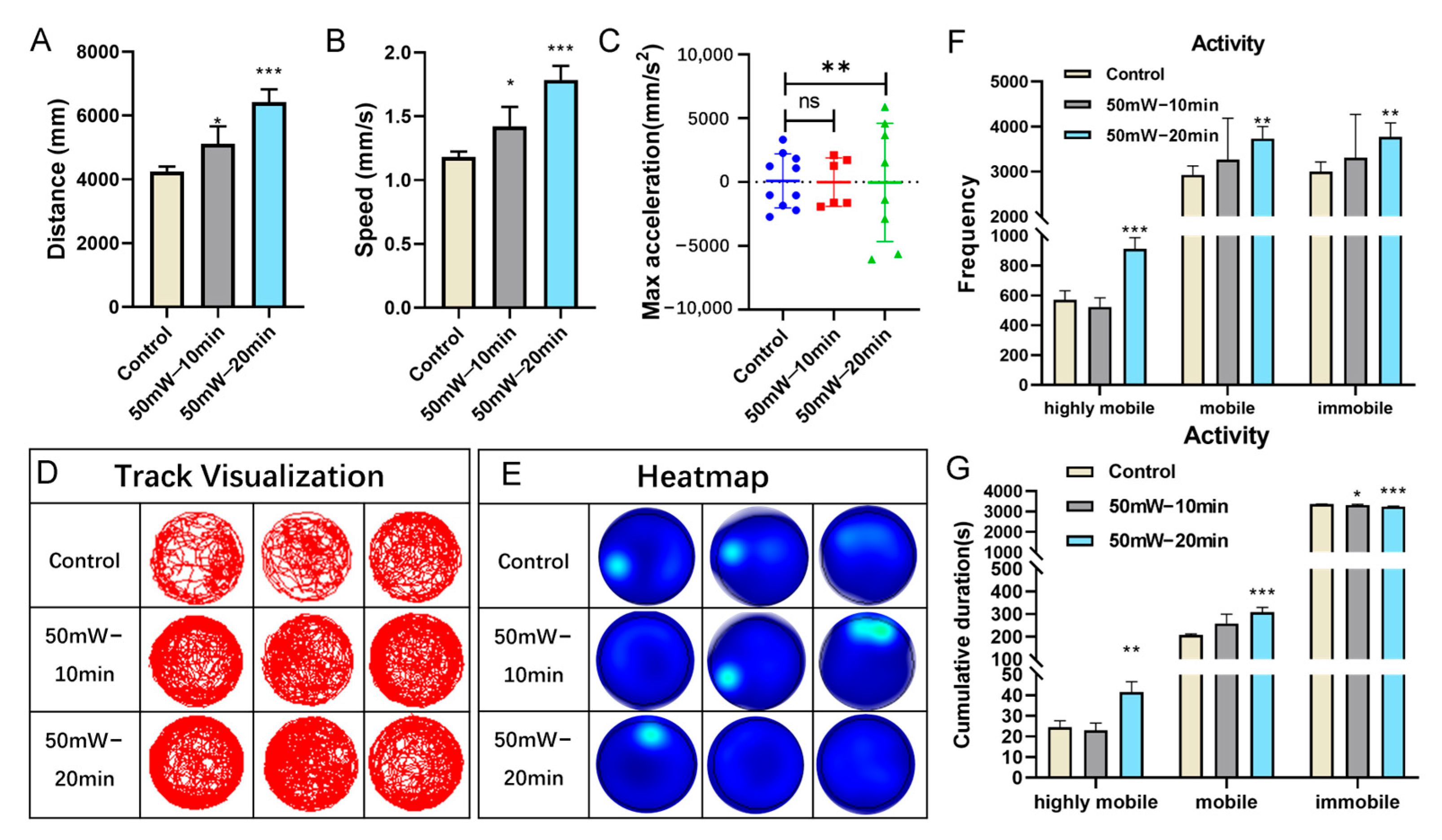
3.1. THz Radiation Promotes the Behavior of Zebrafish Larvae
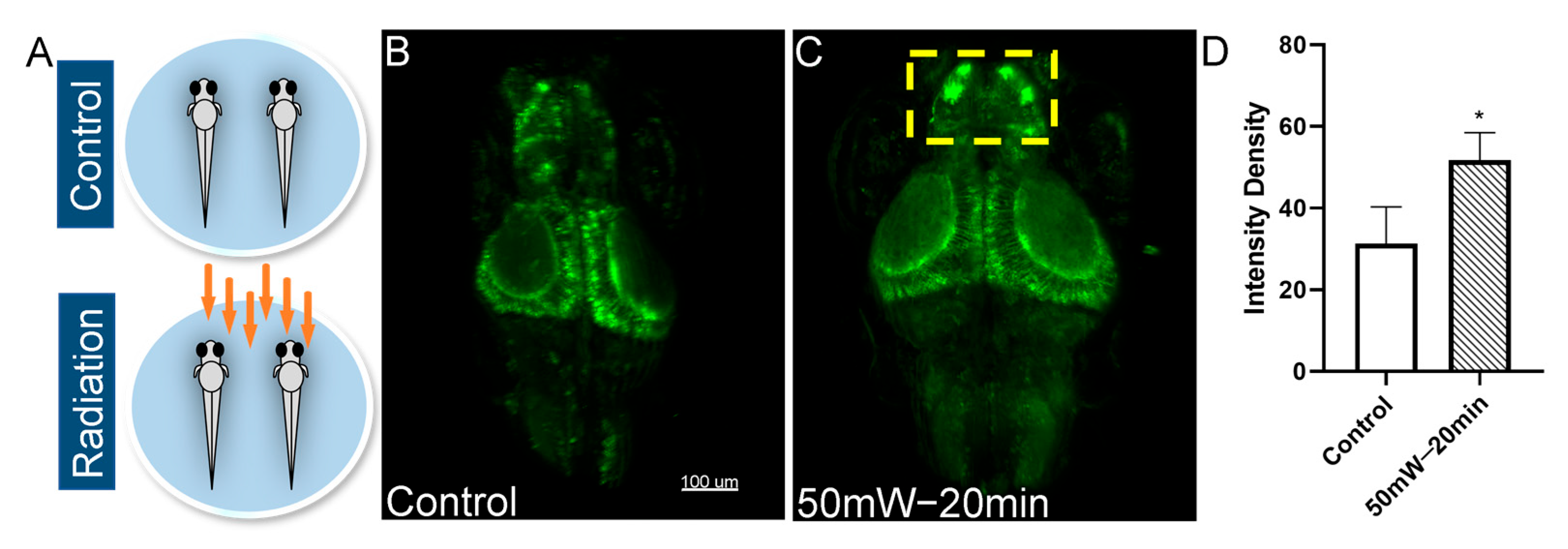
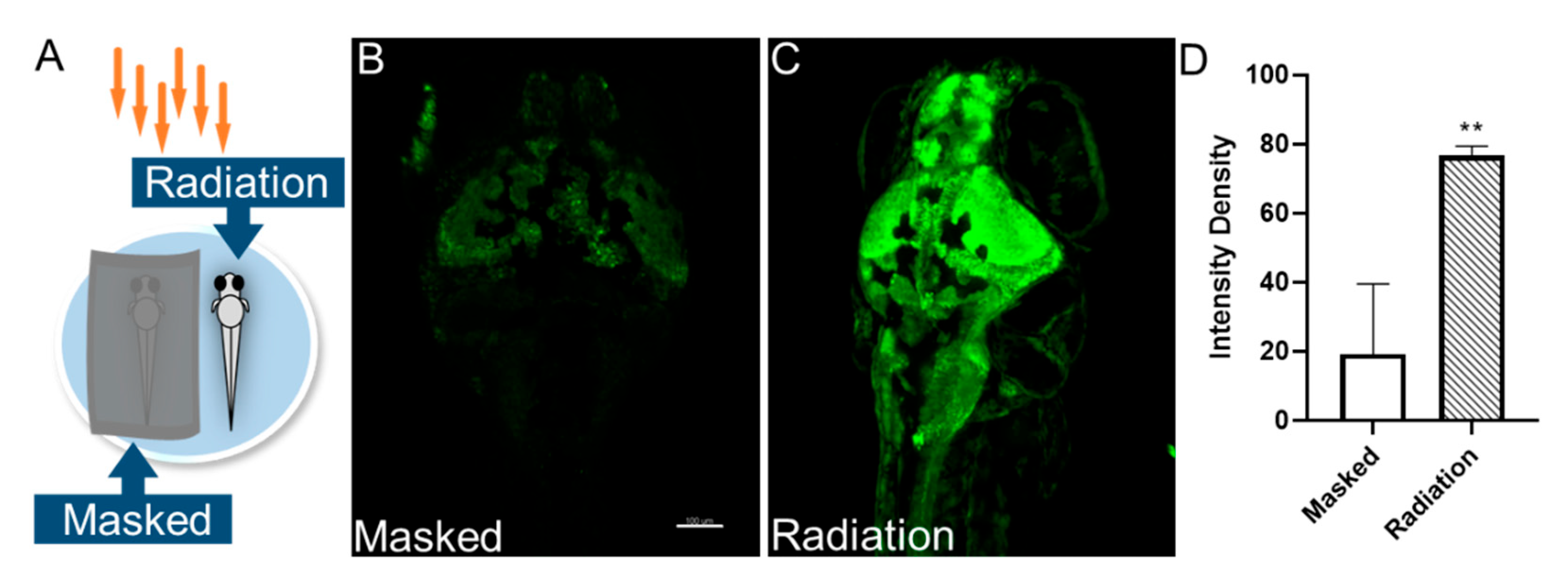
3.2. THz Radiation Promotes the Neural Activities of Zebrafish Larvae
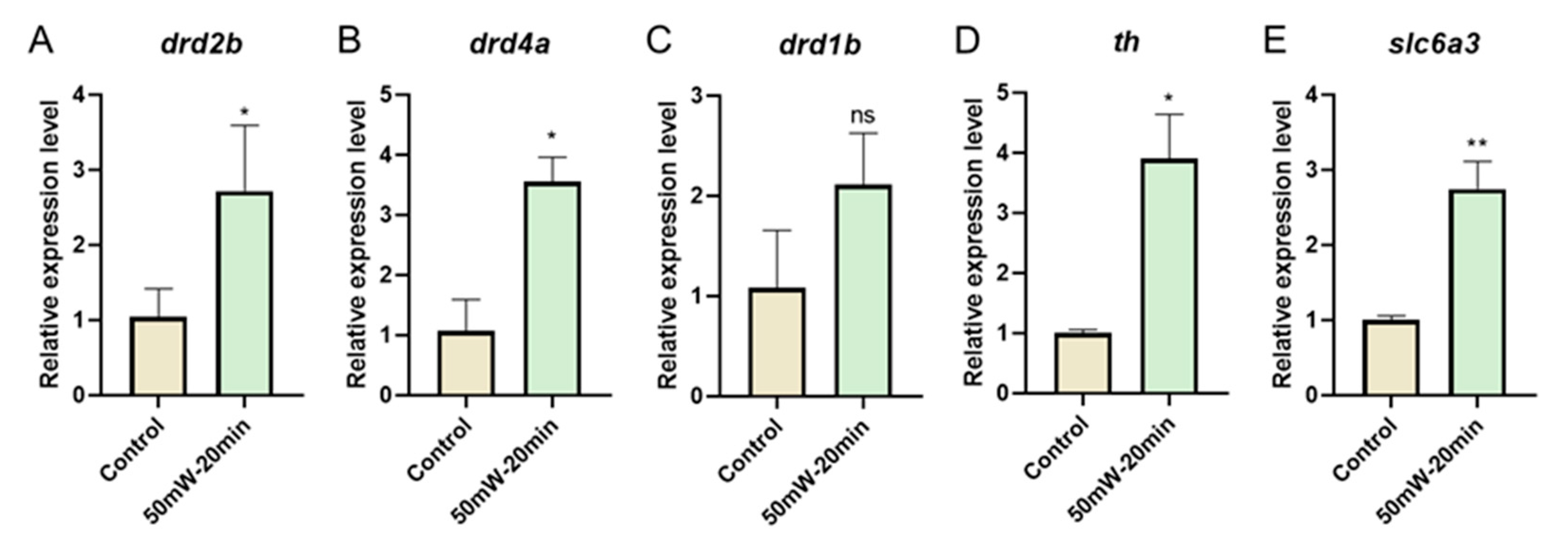
3.3. THz Radiation Promotes the Dopamine-Related Gene Expression of Zebrafish Larvae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Approval
References
- Andersen, J.; Heimdal, J.; Mahler, D.W.; Nelander, B.; Larsen, R.W. Communication: THz absorption spectrum of the CO2-H2O complex: Observation and assignment of intermolecular van der Waals vibrations. J. Chem. Phys. 2014, 140, 091103. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, N.S.; Nazli, R.; Zafar, H.; Fatima, S. Effects of lipid based Multiple Micronutrients Supplement on the birth outcome of underweight pre-eclamptic women: A randomized clinical trial. Pak. J. Med. Sci. 2022, 38, 219–226. [Google Scholar] [PubMed]
- Jamshed, M.A.; Héliot, F.; Brown, T.W.C. A Survey on Electromagnetic Risk Assessment and Evaluation Mechanism for Future Wireless Communication Systems. IEEE J. Electromagn. RF Microw. Med. Biol. 2020, 4, 24–36. [Google Scholar] [CrossRef]
- Cherkasova, O.P.; Serdyukov, D.S.; Ratushnyak, A.S.; Nemova, E.F.; Kozlov, E.N.; Shidlovskii, Y.V.; Zaytsev, K.I.; Tuchin, V.V. Effects of Terahertz Radiation on Living Cells: A Review. Opt. Spectrosc. 2020, 128, 855–866. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, M.; Liu, Y.; Liu, H.; Ren, K.; Xue, Q.; Zhang, H.; Zhi, N.; Wang, W.; Wu, S. Terahertz exposure enhances neuronal synaptic transmission and oligodendrocyte differentiation in vitro. iScience 2021, 24, 103485. [Google Scholar] [CrossRef]
- Kirichuk, V.F.; Antipova, O.N.; Krylova, Y.A. Effect of continuous irradiation with terahertz electromagnetic waves of the NO frequency range on behavioral reactions of male albino rats under stress conditions. Bull. Exp. Biol. Med. 2014, 157, 184–189. [Google Scholar] [CrossRef]
- Kirichuk, V.F.; Efimova, N.V.; Andronov, E.V. Effect of high power terahertz irradiation on platelet aggregation and behavioral reactions of albino rats. Bull. Exp. Biol. Med. 2009, 148, 746–749. [Google Scholar] [CrossRef]
- Bondar, N.P.; Kovalenko, I.L.; Avgustinovich, D.F.; Khamoyan, A.G.; Kudryavtseva, N.N. Behavioral effect of terahertz waves in male mice. Bull. Exp. Biol. Med. 2008, 145, 401–405. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef]
- Gerlai, R. A small fish with a big future: Zebrafish in behavioral neuroscience. Rev. Neurosci. 2011, 22, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Song, X.; Miao, Y.; Wang, Y.; Zhou, C.; Geng, Z.; Du, J.; Moussian, B.; Su, Y.; Liu, X.; et al. Aconitum carmichaelii triggers neurotoxicity and Parkinson-like symptoms through initiation of ROS-mitochondrial apoptosis and the Nox5/DJ-1 signaling pathway. BMEMat 2023, 1, e12019. [Google Scholar] [CrossRef]
- Portugues, R.; Feierstein, C.E.; Engert, F.; Orger, M.B. Whole-brain activity maps reveal stereotyped, distributed networks for visuomotor behavior. Neuron 2014, 81, 1328–1343. [Google Scholar] [CrossRef]
- Ahrens, M.B.; Li, J.M.; Orger, M.B.; Robson, D.N.; Schier, A.F.; Engert, F.; Portugues, R. Brain-wide neuronal dynamics during motor adaptation in zebrafish. Nature 2012, 485, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Wang, Y.; Yue, F.; Cheng, X.; Dang, R.; Qiao, Q.; Sun, X.; Li, X.; Jiang, Q.; Yao, J.; et al. The paraventricular thalamus is a critical thalamic area for wakefulness. Science 2018, 362, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Deng, J.; Liu, K.F.; Wu, Z.Y.; Shi, Y.F.; Guo, W.M.; Mao, Q.Q.; Liu, X.J.; Li, H.; Sun, Y.G. A central neural circuit for itch sensation. Science 2017, 357, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zhang, Q.; Liao, X.; Li, Q.; Liang, S.; Li, X.; Zhang, Y.; Li, X.; Wang, H.; Qin, H.; et al. A corticopontine circuit for initiation of urination. Nat. Neurosci. 2018, 21, 1541–1550. [Google Scholar] [CrossRef]
- Qin, H.; Fu, L.; Hu, B.; Liao, X.; Lu, J.; He, W.; Liang, S.; Zhang, K.; Li, R.; Yao, J.; et al. A Visual-Cue-Dependent Memory Circuit for Place Navigation. Neuron 2018, 99, 47–55.e44. [Google Scholar] [CrossRef]
- Vladimirov, N.; Mu, Y.; Kawashima, T.; Bennett, D.V.; Yang, C.T.; Looger, L.L.; Keller, P.J.; Freeman, J.; Ahrens, M.B. Light-sheet functional imaging in fictively behaving zebrafish. Nat. Methods 2014, 11, 883–884. [Google Scholar] [CrossRef]
- Wilmink, G.J.; Rivest, B.D.; Roth, C.C.; Ibey, B.L.; Payne, J.A.; Cundin, L.X.; Grundt, J.E.; Peralta, X.; Mixon, D.G.; Roach, W.P. In vitro investigation of the biological effects associated with human dermal fibroblasts exposed to 2.52 THz radiation. Lasers Surg. Med. 2011, 43, 152–163. [Google Scholar] [CrossRef]
- Cabib, S.; Puglisi-Allegra, S. The mesoaccumbens dopamine in coping with stress. Neurosci. Biobehav. Rev. 2012, 36, 79–89. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Le, W. Advances of terahertz technology in neuroscience: Current status and a future perspective. iScience 2021, 24, 103548. [Google Scholar] [CrossRef]
- Vernier, P.T.; Levine, Z.A.; Ho, M.C.; Xiao, S.; Semenov, I.; Pakhomov, A.G. Picosecond and Terahertz Perturbation of Interfacial Water and Electropermeabilization of Biological Membranes. J. Membr. Biol. 2015, 248, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Borovkova, M.; Serebriakova, M.; Fedorov, V.; Sedykh, E.; Vaks, V.; Lichutin, A.; Salnikova, A.; Khodzitsky, M. Investigation of terahertz radiation influence on rat glial cells. Biomed. Opt. Express 2017, 8, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, B.S.; Gelev, V.; Bishop, A.R.; Usheva, A.; Rasmussen, K.Ø. DNA Breathing Dynamics in the Presence of a Terahertz Field. Phys. Lett. A 2010, 374, 1214. [Google Scholar] [CrossRef]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Kettunen, P. Calcium Imaging in the Zebrafish. Adv. Exp. Med. Biol. 2020, 1131, 901–942. [Google Scholar]
- Peterka, D.S.; Takahashi, H.; Yuste, R. Imaging voltage in neurons. Neuron 2011, 69, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, R.; Shang, C.; Zhang, Y.; Feng, Y.; Pan, L.; Xu, B.; Hyeon, T.; Bu, W.; Shi, J.; et al. Near-Infrared Voltage Nanosensors Enable Real-Time Imaging of Neuronal Activities in Mice and Zebrafish. J. Am. Chem. Soc. 2020, 142, 7858–7867. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef]
- Ren, J.; Qin, C.; Hu, F.; Tan, J.; Qiu, L.; Zhao, S.; Feng, G.; Luo, M. Habenula “cholinergic” neurons co-release glutamate and acetylcholine and activate postsynaptic neurons via distinct transmission modes. Neuron 2011, 69, 445–452. [Google Scholar] [CrossRef]
- Berridge, C.W.; Schmeichel, B.E.; España, R.A. Noradrenergic modulation of wakefulness/arousal. Sleep Med. Rev. 2012, 16, 187–197. [Google Scholar] [CrossRef]
- Holroyd, C.B.; Coles, M.G.H. The neural basis of human error processing: Reinforcement learning, dopamine, and the error-related negativity. Psychol. Rev. 2002, 109, 679–709. [Google Scholar] [CrossRef]
- Sun, F.; Zeng, J.; Jing, M.; Zhou, J.; Feng, J.; Owen, S.F.; Luo, Y.; Li, F.; Wang, H.; Yamaguchi, T.; et al. A Genetically Encoded Fluorescent Sensor Enables Rapid and Specific Detection of Dopamine in Flies, Fish, and Mice. Cell 2018, 174, 481–496.e419. [Google Scholar] [CrossRef]
- Jing, M.; Zhang, P.; Wang, G.; Feng, J.; Mesik, L.; Zeng, J.; Jiang, H.; Wang, S.; Looby, J.C.; Guagliardo, N.A.; et al. A genetically encoded fluorescent acetylcholine indicator for in vitro and in vivo studies. Nat. Biotechnol. 2018, 36, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, C.; Lischinsky, J.E.; Jing, M.; Zhou, J.; Wang, H.; Zhang, Y.; Dong, A.; Wu, Z.; Wu, H.; et al. A Genetically Encoded Fluorescent Sensor for Rapid and Specific In Vivo Detection of Norepinephrine. Neuron 2019, 102, 745–761.e748. [Google Scholar] [CrossRef]
- Pinoli, M.; Marino, F.; Cosentino, M. Dopaminergic Regulation of Innate Immunity: A Review. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2017, 12, 602–623. [Google Scholar] [CrossRef] [PubMed]
- Marzagalli, R.; Leggio, G.M.; Bucolo, C.; Pricoco, E.; Keay, K.A.; Cardile, V.; Castorina, S.; Salomone, S.; Drago, F.; Castorina, A. Genetic blockade of the dopamine D3 receptor enhances hippocampal expression of PACAP and receptors and alters their cortical distribution. Neuroscience 2016, 316, 279–295. [Google Scholar] [CrossRef]
- Castorina, A.; D’Amico, A.G.; Scuderi, S.; Leggio, G.M.; Drago, F.; D’Agata, V. Dopamine D3 receptor deletion increases tissue plasminogen activator (tPA) activity in prefrontal cortex and hippocampus. Neuroscience 2013, 250, 546–556. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, Z.; Sun, L.; Xiang, Z.; Chang, C.; Fan, C. Physicochemical Insights on Terahertz Wave Diminished Side Effects of Drugs from Slow Dissociation. ACS Nano 2022, 16, 8419–8426. [Google Scholar] [CrossRef]
- Liu, X.; Qiao, Z.; Chai, Y.; Zhu, Z.; Wu, K.; Ji, W.; Li, D.; Xiao, Y.; Mao, L.; Chang, C.; et al. Nonthermal and reversible control of neuronal signaling and behavior by midinfrared stimulation. Proc. Natl. Acad. Sci. USA 2021, 118, e2015685118. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Wu, K.; Wang, P.; Ding, Y.; Yang, X.; Chang, C.; Yang, Y. Sensory input-dependent gain modulation of the optokinetic nystagmus by mid-infrared stimulation in pigeons. eLife 2023, 12, e78729. [Google Scholar] [CrossRef]
- Zhang, J.; He, Y.; Liang, S.; Liao, X.; Li, T.; Qiao, Z.; Chang, C.; Jia, H.; Chen, X. Non-invasive, opsin-free mid-infrared modulation activates cortical neurons and accelerates associative learning. Nat. Commun. 2021, 12, 2730. [Google Scholar] [CrossRef]
- Hao, Y.; Li, W.; Wang, H.; Zhang, J.; Wang, H.; Dong, J.; Yao, B.; Xu, X.; Zhao, L.; Peng, R. Microwave radiation induces neuronal autophagy through miR-30a-5p/AMPKα2 signal pathway. Biosci. Rep. 2022, 42, BSR20212584. [Google Scholar] [CrossRef]
- Shahin, S.; Banerjee, S.; Singh, S.P.; Chaturvedi, C.M. 2.45 GHz Microwave Radiation Impairs Learning and Spatial Memory via Oxidative/Nitrosative Stress Induced p53-Dependent/Independent Hippocampal Apoptosis: Molecular Basis and Underlying Mechanism. Toxicol. Sci. Off. J. Soc. Toxicol. 2015, 148, 380–399. [Google Scholar] [CrossRef]
- Zhi, W.J.; Peng, R.Y.; Li, H.J.; Zou, Y.; Yao, B.W.; Wang, C.Z.; Liu, Z.H.; Gao, X.H.; Xu, X.P.; Dong, J.; et al. Microwave radiation leading to shrinkage of dendritic spines in hippocampal neurons mediated by SNK-SPAR pathway. Brain Res. 2018, 1679, 134–143. [Google Scholar] [CrossRef]
- Sharma, A.; Kesari, K.K.; Saxena, V.K.; Sisodia, R. Ten gigahertz microwave radiation impairs spatial memory, enzymes activity, and histopathology of developing mice brain. Mol. Cell. Biochem. 2017, 435, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Z.F.; Shang, C.F.; Wang, Y.F.; Yang, Z.; Yang, C.; Li, F.N.; Xie, J.Z.; Pan, J.W.; Fu, L.; Du, J.L. All-optical imaging and manipulation of whole-brain neuronal activities in behaving larval zebrafish. Biomed. Opt. Express 2018, 9, 6154–6169. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Xu, X.; Wang, J.; Wang, J.; He, Y.; Mu, Y.; Shi, G. Polarization effects on the fluorescence emission of zebrafish neurons using light-sheet microscopy. Biomed. Opt. Express 2022, 13, 6733–6744. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chen, H.; Pang, M.; Liu, X.; Wang, J.I.; Zhang, X.D.; Ming, D. Airy-like beam-based light-sheet microscopy with improved FOV for zebrafish intracerebral hemorrhage. Opt. Express 2022, 30, 14709–14722. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Temp before Irradiation (°C) | Temp after Irradiation (°C) | Difference Value |
|---|---|---|---|
| 1 | 19.7 | 19.8 | 0.1 |
| 2 | 19.7 | 19.9 | 0.2 |
| 3 | 19.7 | 19.9 | 0.2 |
| 4 | 19.7 | 19.8 | 0.1 |
| 5 | 19.7 | 19.8 | 0.1 |
| 6 | 19.7 | 19.7 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Li, H.; Liu, X.; Pang, M.; Wang, Y. Calcium Imaging Characterize the Neurobiological Effect of Terahertz Radiation in Zebrafish Larvae. Sensors 2023, 23, 7689. https://doi.org/10.3390/s23187689
Song X, Li H, Liu X, Pang M, Wang Y. Calcium Imaging Characterize the Neurobiological Effect of Terahertz Radiation in Zebrafish Larvae. Sensors. 2023; 23(18):7689. https://doi.org/10.3390/s23187689
Chicago/Turabian StyleSong, Xin, Haibin Li, Xiuyun Liu, Meijun Pang, and Yuye Wang. 2023. "Calcium Imaging Characterize the Neurobiological Effect of Terahertz Radiation in Zebrafish Larvae" Sensors 23, no. 18: 7689. https://doi.org/10.3390/s23187689
APA StyleSong, X., Li, H., Liu, X., Pang, M., & Wang, Y. (2023). Calcium Imaging Characterize the Neurobiological Effect of Terahertz Radiation in Zebrafish Larvae. Sensors, 23(18), 7689. https://doi.org/10.3390/s23187689