
Effects of Lake Productivity on Density and Size Structure of Pelagic Fish Estimated by Means of Echosounding in 17 Lakes in Southeast Norway


Abstract
1. Introduction
2. Materials and Methods
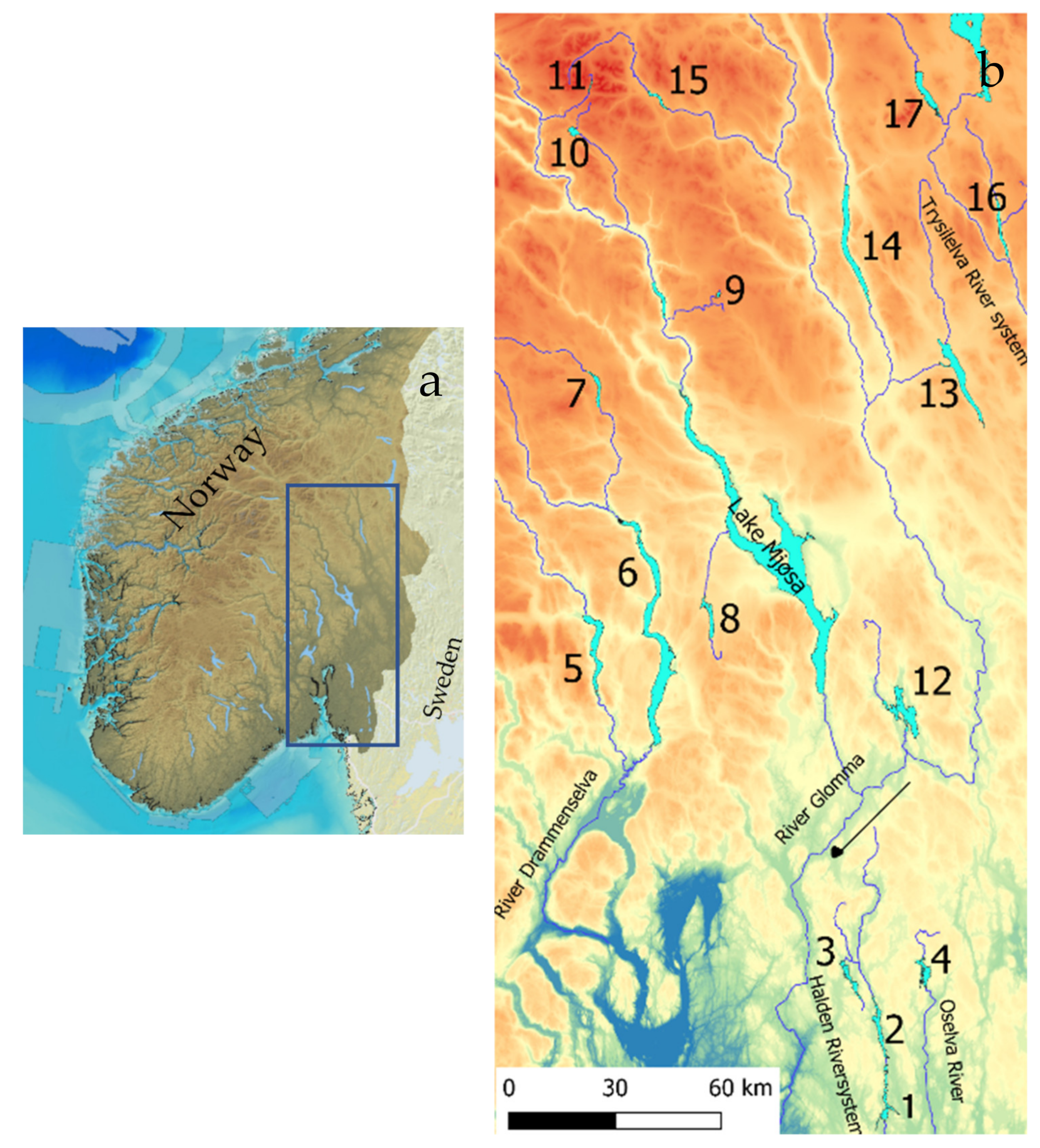
2.1. Study Area
2.2. Sampling and Analysis
2.3. Data Treatment and Statistical Analysis
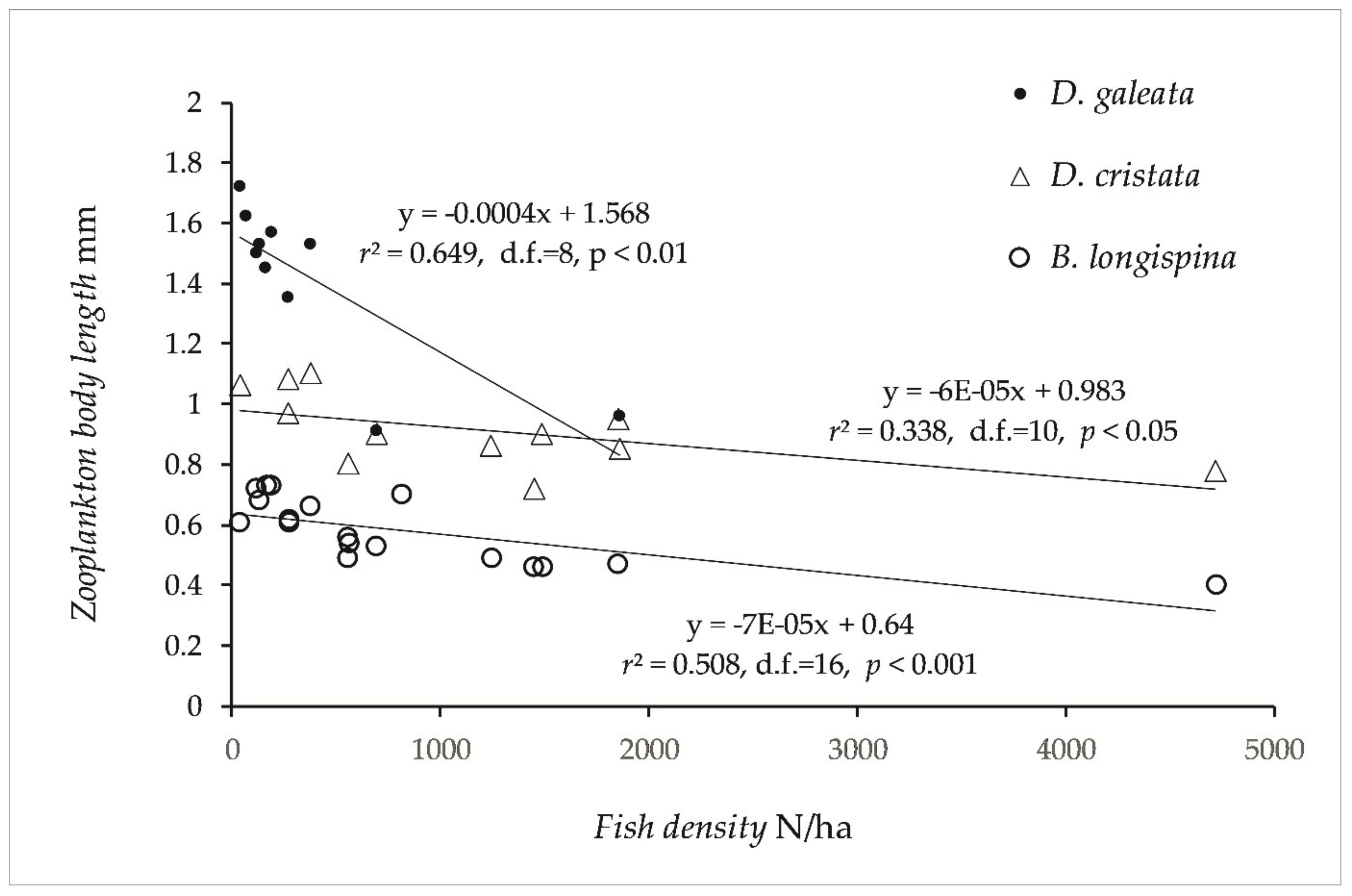
3. Results
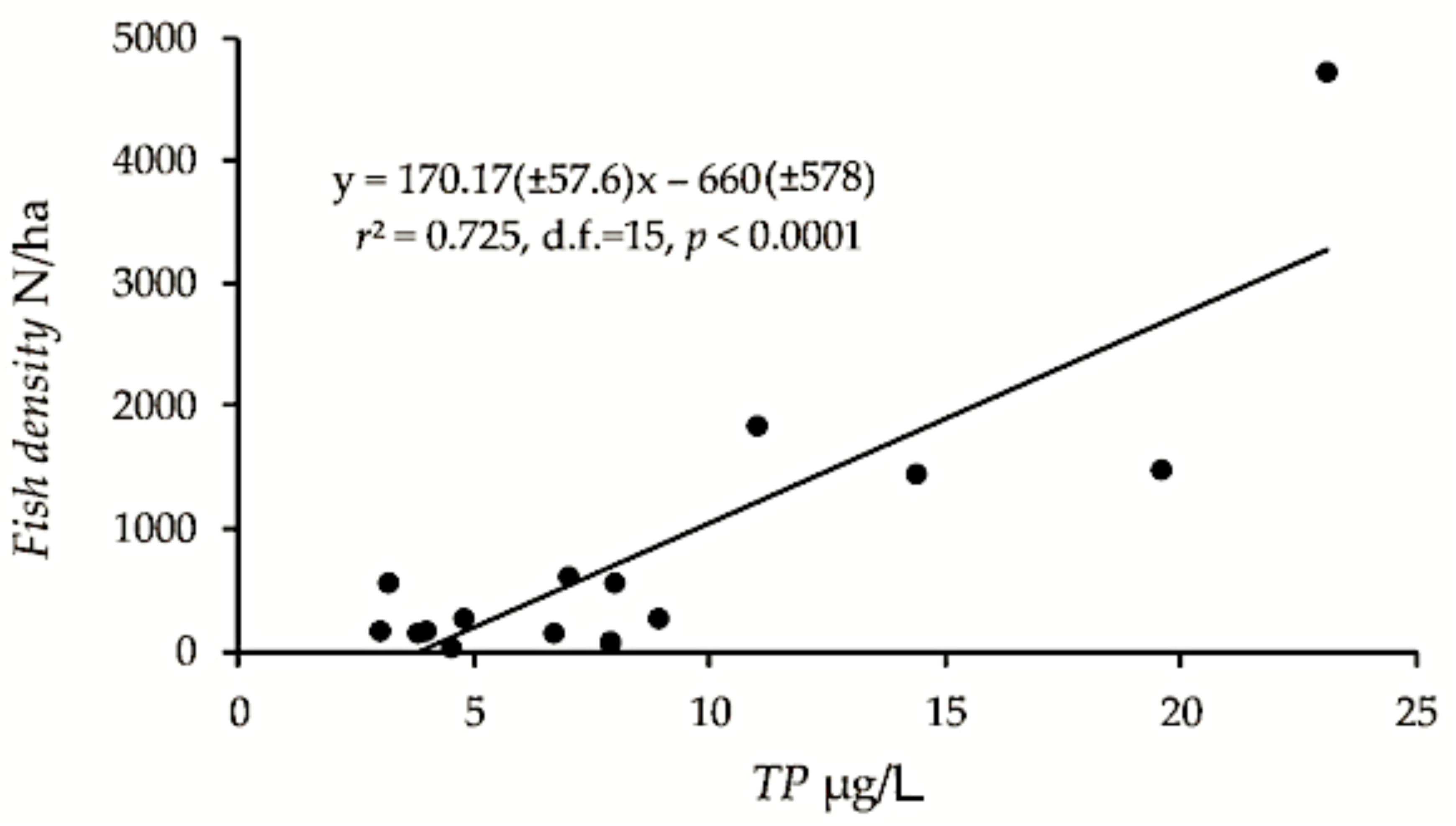
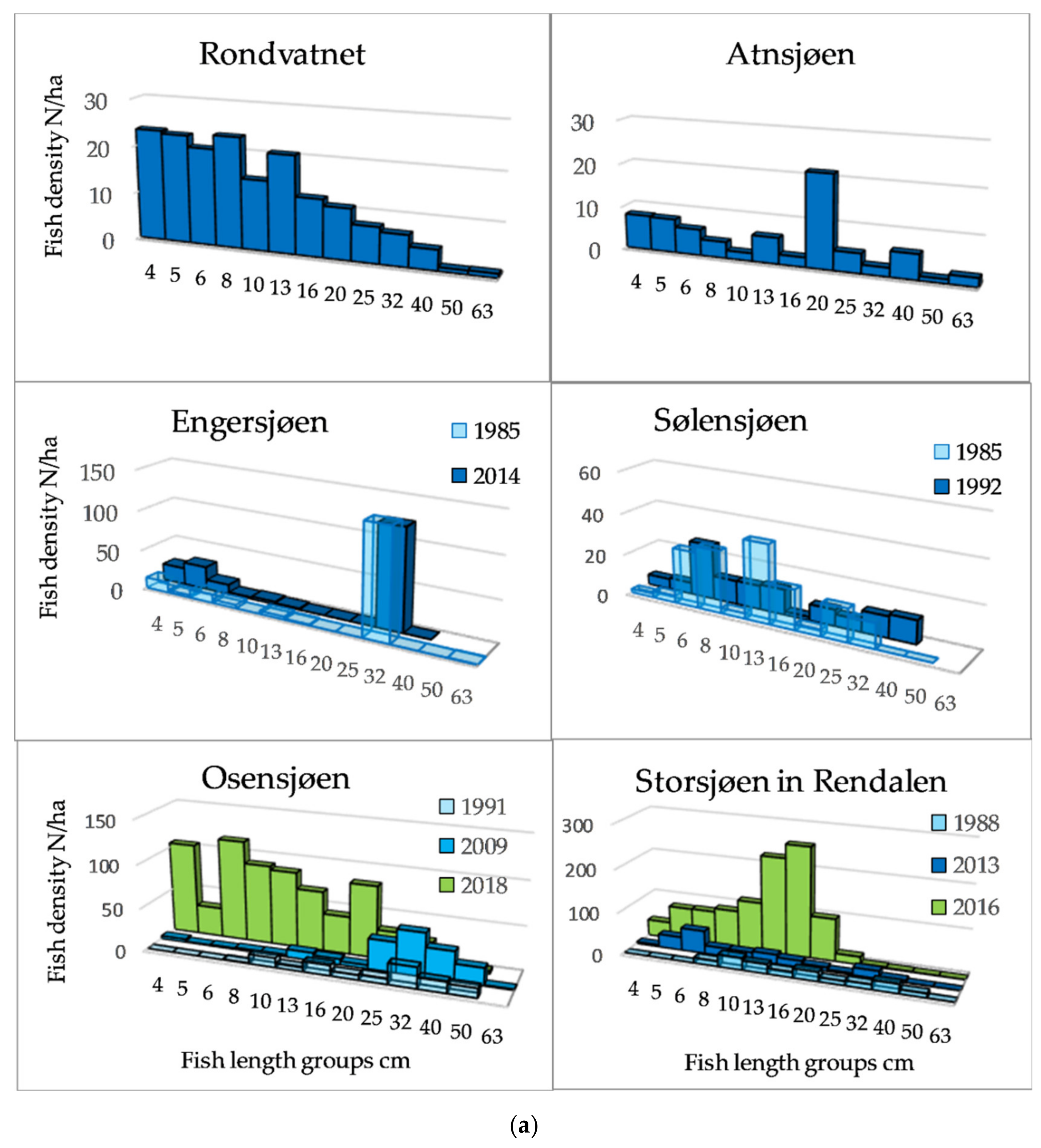
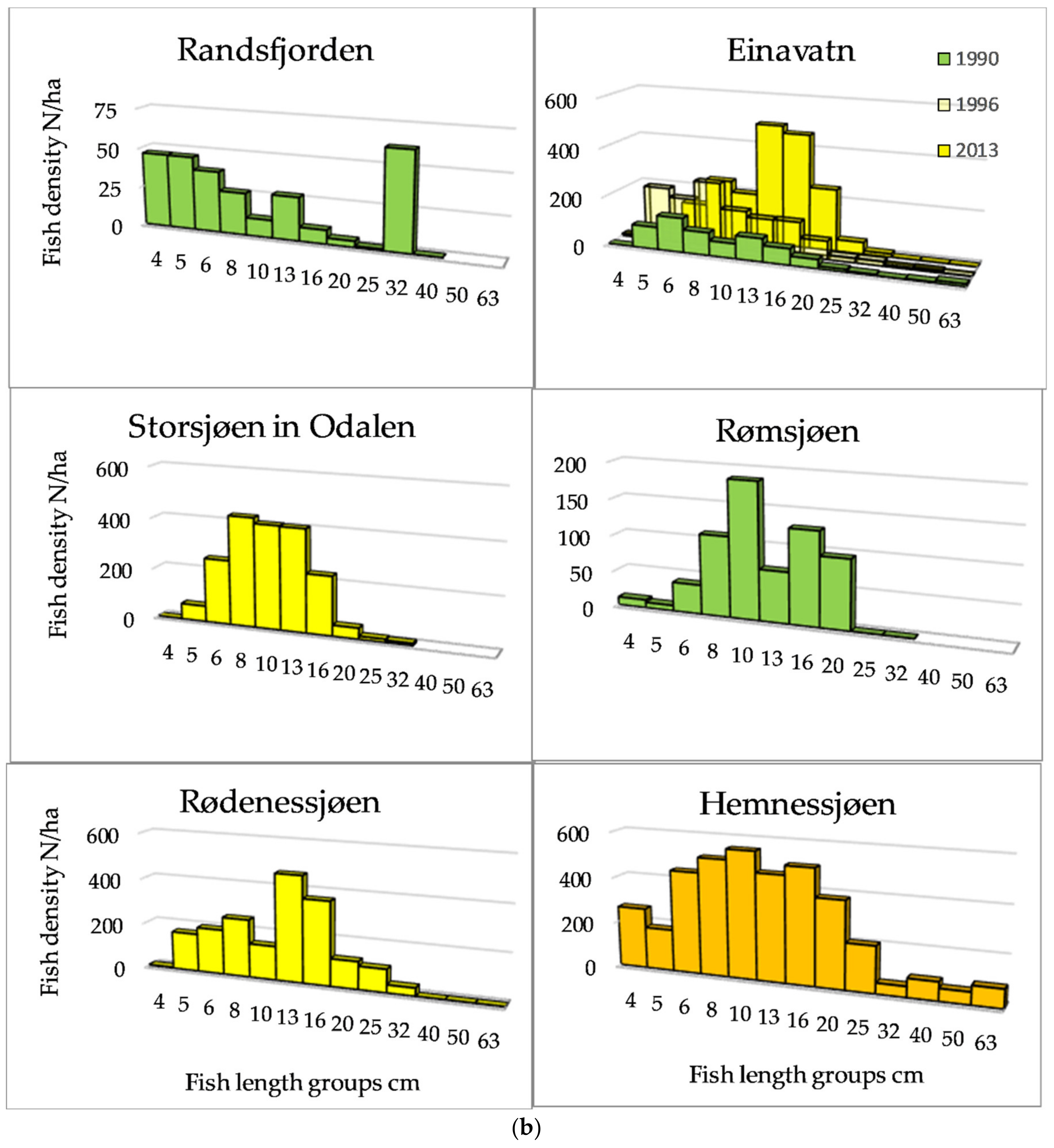
3.1. Fish Density
3.2. Zooplankton
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of the World Fisheries and Aquaculture; Mathiesen, A.M., Ed.; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2010; p. 197. [Google Scholar]
- Sarvala, J.; Helminen, H.; Auvinen, H. Portrait of a flourishing freshwater fishery: Pyhäjärvi, a lake in SW-Finland. Boreal Environ. Res. 1998, 3, 329–345. [Google Scholar]
- Anonymous. Innlandsfiskeforvaltning 2010–2015. In DN-rapport 6-2010; Svarte, Y., Ed.; The Directorate of Nature Management: Trondheim, Norway, 2010; p. 42. [Google Scholar]
- Fluet-Chouinard, E.; Funge-Smith, S.; McIntyre, P.B. Global hidden harvest of freshwater fish revealed by household surveys. Proc. Natl. Acad. Sci. USA 2018, 115, 7623–7628. [Google Scholar] [CrossRef]
- FAO. EU Intervention in Inland Fisheries; Mathiesen, A.M., Ed.; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2007; p. 134. [Google Scholar]
- Museth, J.; Sandlund, O.; Borgstrøm, R. Coexistence between introduced whitefish (Coregonus lavaretus) and native Arctic charr (Salvelinus alpinus) depends on heavy whitefish exploitation. Adv. Limnol. 2005, 60, 343–350. [Google Scholar]
- Van Gemert, R.; Andersen, K.H. Implications of late-in-life density-dependent growth for fishery size-at-entry leading to maximum sustainable yield. J. Mar. Sci. 2018, 75, 1295–1305. [Google Scholar] [CrossRef]
- Linløkken, A.N.; Sandlund, O.T. Recruitment of sympatric vendace (Coregonus albula) and whitefish (C. lavaretus) is affected by different environmental factors. Ecol. Freshw. Fish 2015, 25, 652–663. [Google Scholar] [CrossRef]
- Tolonen, A.; Lappalainen, J.; Pulliainen, E. Seasonal growth and year class strength variations of perch near the northern limits of its distribution range. J. Fish Biol. 2003, 63, 176–186. [Google Scholar] [CrossRef]
- Böhling, P.; Hudd, R.; Lehtonen, H.; Karås, P.; Neuman, E.; Thoresson, G. Variations in Year-Class Strength of Different Perch (Perca fluviatilis) Populations in the Baltic Sea with Special Reference to Temperature and Pollution. Can. J. Fish. Aquat. Sci. 1991, 48, 1181–1187. [Google Scholar] [CrossRef]
- Neuman, E. The growth and year-class strength of perch in some Baltic archipelagos, with special reference to temperature. Report from Institute of Freshwater Research. Drottningholm 1976, 55, 51–70. [Google Scholar]
- Brown, R.W.; Taylor, W.W.; Assel, R.A. Factors Affecting the Recruitment of Lake Whitefish in Two Areas of Northern Lake Michigan. J. Great Lakes Res. 1993, 19, 418–428. [Google Scholar] [CrossRef]
- Tõnno, I.; Agasild, H.; Kõiv, T.; Freiberg, R.; Nõges, P.; Nõges, T. Algal Diet of Small-Bodied Crustacean Zooplankton in a Cyanobacteria-Dominated Eutrophic Lake. PLoS ONE 2016, 11, e0154526. [Google Scholar] [CrossRef]
- Vakkilainen, K.; Kairesalo, T.; Hietala, J.; Balayla, D.M.; Becares, E.; Van De Bund, W.J.; Van Donk, E.; Fernandez-Alaez, M.; Gyllstrom, M.; Hansson, L.-A.; et al. Response of zooplankton to nutrient enrichment and fish in shallow lakes: A pan-European mesocosm experiment. Freshw. Biol. 2004, 49, 1619–1632. [Google Scholar] [CrossRef]
- Hansson, L.-A.; Nicolle, A.; Brodersen, J.; Romare, P.; Nilsson, P.A.; Brönmark, C.; Skov, C. Consequences of fish predation, migration, and juvenile ontogeny on zooplankton spring dynamics. Limnol. Oceanogr. 2007, 52, 696–706. [Google Scholar] [CrossRef]
- Brucet, S.; Boix, D.; Quintana, X.D.; Jensen, E.; Nathansen, L.W.; Trochine, C.; Meerhoff, M.; Gascon, S.; Jeppesena, E. Factors influencing zooplankton size structure at contrasting temperatures in coastal shallow lakes: Implications for effects of climate change. Limnol. Oceanogr. 2010, 55, 1697–1711. [Google Scholar] [CrossRef]
- Kagami, M.; Yoshida, T.; Gurung, T.B.; Urabe, J. Direct and indirect effects of zooplankton on algal composition in in situ grazing experiments. Oecologia 2002, 133, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Ginter, K.; Blank, K.; Haberman, J.; Kangur, A.; Kangur, K. Fish predation pressure on zooplankton in a large northern temperate lake: Impact of adult predators versus juvenile predators. Proc. Est. Acad. Sci. 2018, 67, 356–367. [Google Scholar] [CrossRef]
- Urrutia-Cordero, P.; Ekvall, M.K.; Hansson, L.-A. Controlling Harmful Cyanobacteria: Taxa-Specific Responses of Cyanobacteria to Grazing by Large-Bodied Daphnia in a Biomanipulation Scenario. PLoS ONE 2016, 11, e0153032. [Google Scholar] [CrossRef]
- Sanni, S.; Wærvågen, S.B. Oligotrophication as a result of planktivorous fish removal with rotenone in the small, eutrophic lake Mosvatn, Norway. Hydrobiologia 1990, 200, 263–274. [Google Scholar] [CrossRef]
- Cornett, R.J.; Rigler, F.H. The areal hypolimnetic oxygen deficit: An empirical test of the model1. Limnol. Oceanogr. 1980, 25, 672–679. [Google Scholar] [CrossRef]
- Cornett, R.J.; Rigler, F.H. Hypolinimetic oxygen deficits: Their prediction and interpretation. Science 1979, 205, 580–581. [Google Scholar] [CrossRef]
- Rhodes, J.; Hetzenauer, H.; Frassl, M.A.; Rothhaupt, K.O.; Rinke, K. Long-term development of hypolimnetic oxygen depletion rates in the large Lake Constance. Ambio 2017, 46, 554–565. [Google Scholar] [CrossRef]
- Breen, B.; Curtis, J.; Hynes, S. Water quality and recreational use of public waterways. J. Environ. Econ. Policy 2018, 7, 1–15. [Google Scholar] [CrossRef]
- Li, P.; Wu, J. Drinking Water Quality and Public Health. Expo. Health 2019, 11, 73–79. [Google Scholar] [CrossRef]
- Persson, L.; Byström, P.; Wahlström, E.; Westman, E. Trophic dynamics in a whole lake experiment: Size-structured interactions and recruitment variation. Oikos 2004, 106, 263–274. [Google Scholar] [CrossRef]
- Ersoy, Z.; Brucet, S.; Bartrons, M.; Mehner, T. Short-term fish predation destroys resilience of zooplankton communities and prevents recovery of phytoplankton control by zooplankton grazing. PLoS ONE 2019, 14, e0212351. [Google Scholar] [CrossRef]
- Pollom, R.; Rose, G. A Global Review of the Spatial, Taxonomic, and Temporal Scope of Freshwater Fisheries Hydroacoustics Research. Environ. Rev. 2016, 24, 333–347. [Google Scholar] [CrossRef]
- Kim, K.; Kim, M.H.; Shevlyakov, G.; Shin, S. Editorial for Special Issue: Fishery Acoustics, Applied Sciences, and Practical Applications. Appl. Sci. 2019, 9, 4774. [Google Scholar] [CrossRef]
- Linløkken, A.N.; Næstad, F.; Langdal, K.; Østbye, K. Comparing Fish Density and Echo Strength Distribution Recorded by Two Generations of Single Beam Echo Sounders. Appl. Sci. 2019, 9, 2041. [Google Scholar] [CrossRef]
- Huitfeldt-Kaas, H. Ferskvandsfiskenes Utbredelse og Indvandring i Norge: Med et Tillæg om Krebsen; Centraltrykkeriet: Kristiania, Norway, 1918. [Google Scholar]
- Walseng, B. Verneplan I og II, Østfold.—Krepsdyrundersøkelser. In Oppdragsmelding 304; Norwegian Institute of Nature Research: Oslo, Norway, 1994; p. 31. [Google Scholar]
- Vøllestad, A. Resultat av Prøvefiske i Bjørkelangen, Øgdern og Rødenessjøen nr. 2. In Fiskeribiologiske Undersøkelser i Haldenvassdraget; Fylkesmannen i Østfold: Moss, Norway, 1983; p. 57. [Google Scholar]
- Strand, D.A.S.; Hostyeva, V. Overvåking av Haldenvassdraget 2016. In Note; Norwegian Institute of Watre Research: Oslo, Norway, 2017; p. 25. [Google Scholar]
- Vannmiljø. Vannmiljø, Milkødirektoratet. 2021. Available online: https://vannmiljo.miljodirektoratet.no/ (accessed on 7 February 2021).
- Rognerud, S.; Romstad, R.; Mjelde, M. Undersøkelser av Begna 1984–1986; Norwegian Institute of Water Research: Oslo, Norway, 1986; p. 57. [Google Scholar]
- Kjellberg, G. Tiltaksorientert Overvåking av Mjøsa Med Tilløpselver. Samlerapport for 2001 og 2002; Norwegian Institute of Water Research: Oslo, Norway, 2004; p. 166. [Google Scholar]
- Løvik, J.E.; Kjellberg, G.; Brettum, P. Overvåking av Vannkvalitet og Biologiske Forhold i Randsfjorden Med Tilløpselver. Samlerapport for 2001–2004; Norwegian Institute of Water Reserach: Oslo, Norway, 2005; p. 54. [Google Scholar]
- Løvik, J.E.; Rognerud, S. Vannkvaliteten i Randsfjorden og Dokkfløymagasinet i Perioden 1988–98; Norwegian Institute of Water Research: Oslo, Norway, 1999; p. 50. [Google Scholar]
- Kjellberg, G.; Bækken, T.; Lindstrøm, E.-A. Utslipp av Prosessvann fra Skjelbreia Vannverk til Hunnselva. Virkninger på Vannkvalitet og Biota. Undersøkelser i 1997–2000; Norwegian Institute of Water Research: Oslo, Norway, 2001; p. 45. [Google Scholar]
- Løvik, J.E.; Skjelbred, B. Vurdering av Miljøtilstanden i Seks Fjellsjøer i Oppland i 2014; Norwegian Institute of Water Research: Oslo, Norway, 2015; p. 32. [Google Scholar]
- Schartau, A.K. Dyreplankton i Rondvatn og øvre deler av Atnavassdraget. In Program for Naturforvaltning 1987; Vassdragsforsk: Oslo, Norway, 1987. [Google Scholar]
- Kjellberg, G. Tiltaksorientert Overvåking av Vann og Vassdrag i Nord-Odal Kommune; Norwegian Institute of Water Research: Oslo, Norway, 2003; p. 59. [Google Scholar]
- Løvik, J.E.; Jensen, T.C.; Bongard, T.; Magerøy, J.H.; Bækkelie KA, E.; Edvardsen, H.; Kile, M.R.; Skjelbreg, B. Overvåking av Vassdrag i Hedmark 2016; Norwegian Institute of Water Reserach: Oslo, Norway, 2017; p. 80. [Google Scholar]
- Løvik, J.E.; Skjelbred, B. Overvåking av Vassdrag i Hedmark i 2011; Norwegian Institute of Water Reserach: Oslo, Norway, 2012; p. 57. [Google Scholar]
- Løvik, J.E.; Rognerud, S. Overvåking av Vannkvalitet i Regionfelt Østlandet og Rødsmoen Øvingsområde Årsrapport for 2005; Norwegian Institute of Water Research: Oslo, Norway, 2006; p. 59. [Google Scholar]
- Løvik, J.E.; Kjellberg, G. Glåma i Hedmark. Delrapport om Dyreplankton. Undersøkelser i Tidsrommet 1978–80; Norwegian Institute of Water Research: Oslo, Norway, 1982; p. 58. [Google Scholar]
- Kjellberg, G. Undersøkelse av Rena Med Storsjøen 1983–1986. Sluttrapport; Norwgian Institute of Water Research: Oslo, Norway, 1987; p. 89. [Google Scholar]
- Schartau, A.K.; Mjelde, M.; Bækkelie KA, E.; Dokk, J.G.; Moe, T.F.; Jensen, T.C.; Persson, J.; Pettersen, O.; Saksgård, R.; Sandlund, O.T.; et al. ØKOFERSK—Delprogram ØST: Basisovervåking av Utvalgte Innsjøer i 2018. 2019. Available online: https://www.miljodirektoratet.no/globalassets/publikasjoner/m1400/m1400.pdf (accessed on 10 May 2021).
- Lindstrøm, E.-A.; Bongard, T.; Brettum, P.; Bønsnes, T.; Fjellheim, A.; Halvorsen, G.; Hesthagen, T.; Johansen, S.W.; Kvambekk, Å.S.; Raddum, G.; et al. FORSKREF—Forsknings- og referansevassdrag. Årsrapporter Atna og Vikedal 1997–1999. In Utredning 2002–7; Directorate of Nature Management: Oslo, Norway, 2002; p. 189. [Google Scholar]
- Jensen, T.C. Winter decrease of zooplankton abundance and biomass in subalpine oligotrophic Lake Atnsjøen (SE Norway). J. Limnol. 2019, 78. [Google Scholar] [CrossRef]
- Rognerud, S. Basisundersøkelse i Engern 1983; Norsk institutt for vannforskning: Oslo, Norway, 1984; p. 37. [Google Scholar]
- Langeland, A.; Rognerud, S. Vern av Naturlige Næringsrike Innsjøer i Norge. Økologiske Undersøkelsere av Innsjøer og Dammer i Femundområdet 1972–1973; Norwegian Institute of Water Research: Oslo, Norway, 1973; p. 27. [Google Scholar]
- Linløkken, A.; Qvenild, T. Beskatning av Sik og Røye i Sølensjøen; Fylkesmannen i Hedmark, Miljøvernavdelingenf: Hamar, Norway, 1987. [Google Scholar]
- Museth, J.; Sandlund, O.T.; Borgstrøm, R. Coexistence between introduced whitefish (Coregonus lavaretus) and native Arctic charr (Salvelinus alpinus) depends on heavy whitefish exploitation. In Biology and Management of Coregonid Fishes: Proceedings of the Ninth International Symposium on the Biology and Management of Coregonid Fishes held in Olsztyn, Poland, 21 27 August 2005; Jankun, M., Ed.; Schweizerbart Science Publishers: Stuttgart, Germany, 2005; pp. 343–350. [Google Scholar]
- Brettum, P. Alger Som Indikator på Vannkvaliteten i Norske Innsjøer. Planteplankton; Norwegian Institute of Water Research: Oslo, Norway, 1989; p. 114. [Google Scholar]
- Lindem, T. Successes with conventional in situ determinations of fish target strength. In Symposium on Fisheries Acoustics No. 53; International Council for the Exploration of the Sea (ICES): Bergen, Norway, 1983. [Google Scholar]
- Kongsberg Maritime, A.S. SIMRAD EK15. Reference Manual; Kongsberg Maritime AS: Kongsberg, Norway, 2014; p. 309. [Google Scholar]
- Dervo, B.K.; Hegge, O.; Hessen, D.O.; Skurdal, J. Diel food selection of pelagic Arctic charr, Salvelinus alpinus (L), and brown trout, Salmo trutta L., in Lake Atnsjø, SE Norway. J. Fish Biol. 1991, 38, 199–209. [Google Scholar] [CrossRef]
- Hagenlund, M.; Østbye, K.; Langdal, K.; Hassve, M.; Pettersen, R.A.; Anderson, E.; Gregersen, F.; Præbel, K. Fauna crime: Elucidating the potential source and introduction history of European smelt (Osmerus eperlanus L.) into Lake Storsjøen, Norway. Conserv. Genet. 2015, 16, 1085–1098. [Google Scholar] [CrossRef]
- Aglen, A. Random errors of acoustic abundance estiamtes in relation to the survey grid density applied. In Symposium of Fisheries Acoustics; Nakken, O., Venenm, S.C., Eds.; Selected Papers of the ICES/FAO Symposium of Fisheries Acoustics; FAO: Bergen, Norway, 1993; pp. 293–298. [Google Scholar]
- Bagenal, T.B. Methods for assessment of fish production in fresh waters. In IBP Handbook; Blackwell: London, UK, 1978; Volume 3, p. 365. [Google Scholar]
- Kjellberg, G.; Hegge, O.; Lindstrøm, E.A.; Løvik, J. Tiltaksorientert Overvåkning av Mjøsa med Tilløpselver. Årsrapport for 1999; Norwegian Institute for Water Research: Oslo, Norway, 2000; p. 127. [Google Scholar]
- Lindem, T. Hydro Acoustic Data Acquisition System Instruction Manual ver. 3.9; Lindem Data Acquisition: Oslo, Norway, 1990; p. 25. [Google Scholar]
- Balk, H.; Lindem, T. Sonar4 and Sonar5-Pro. Post Processing Systems Operator Manual Version 6.0.3. 2015. Available online: http://folk.uio.no/hbalk/sonar4_5/Downloads.htm (accessed on 7 February 2021).
- Sawada, K.; Takahashi, H.; Abe, K.; Ichii, T.; Watanabe, K.; Takao, Y. Target-strength, length, and tilt-angle measurements of Pacific saury (Cololabis saira) and Japanese anchovy (Engraulis japonicus) using an acoustic-optical system. ICES J. Mar. Sci. 2009, 66, 1212–1218. [Google Scholar] [CrossRef]
- Elliot, J.M. Some Methods for the Statistical Analysis of Samples of Benthic Invertabrates, 2nd ed.; Titus Wilson & Son LTD: Kendal, UK, 1979. [Google Scholar]
- R-Core-Team. R-A language and Environment for Statistical Computing. 2020. Available online: https://www.r-project.org/ (accessed on 10 May 2021).
- Museth, J.; Dervo, B.; Brabrand, Å.; Heggenes, J.; Karlsson, S.; Kraabøl, M. Storørret i Norge. Definisjon, Status, Påvirkningsfaktorer og Kunnskapsbehov; Norwegian Institute of Nature Research: Lillehammer, Norway, 2018; p. 102. [Google Scholar]
- Eloranta, A.P.; Johnsen, S.I.; Power, M.; Bærum, K.M.; Sandlund, O.T.; Finstad, A.G.; Rognerud, S.; Museth, J. Introduced European smelt (Osmerus eperlanus) affects food web and fish community in a large Norwegian lake. Biol. Invasions 2018, 21, 85–98. [Google Scholar] [CrossRef]
- Hessen, D.O.; Faafeng, B.A.; Andersen, T. Replacement of herbovore zooplankton species along gradients of ecosystem productivity and fish predation pressure. Can. J. Fish. Aquat. Sci. 1995, 52, 733–742. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Kristensen, P.; Søndergaard, M.; Mortensen, E.; Sortkjær, O.; Olrik, K. Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes 2: Threshold levels, long-term stability and conclusions. Hydrobiologia 1990, 200, 219–227. [Google Scholar] [CrossRef]
- Horppila, J.; Peltonen, H.; Malinen, T.; Luokkanen, E.; Kairesalo, T. Top-down or Bottom-up Effects by Fish: Issues of Concern in Biomanipulation of Lakes. Restor. Ecol. 1998, 6, 20–28. [Google Scholar] [CrossRef]
- Løvik, J.; Kjellberg, G. Long-term changes of the crustacean zooplankton community in Lake Mjøsa, the largest lake in Norway. J. Limnol. 2003, 62, 143–150. [Google Scholar] [CrossRef]
- Rognerud, S.; Kjellberg, G. Long-term dynamics of the zooplankton community in Lake Mjøsa, the largest lake in Norway. Verh. Int. Ver. Limnol. 1990, 24, 580–585. [Google Scholar] [CrossRef]
- Holtan, H. Eutrophication of Lake Mjøsa in relation to the pollutional load. SIL Proc. 1978, 20, 734–742. [Google Scholar] [CrossRef]
- Ruttner, F. Fundamentals of Limnology, 3rd ed.; University of Toronto Press: Toronto, ON, Canada, 2018. [Google Scholar]
- Sandlund, O.T.; Næsje, T.F.; Kjellberg, G. The size selection of Bosmina longispina and Daphnia galeata by co-occuring cisco (Coregonus albula), whitefish (C. lavaretus) and smelt (Osmerus eperlanus). Arch. Hydrobiol. 1987, 110, 357–363. [Google Scholar]
- Bøhn, T.; Amundsen, P.-A. Effects of invading vendace (Coregonus albula L.) on species composition and body size in two zooplankton communities of the Pasvik River System, northern Norway. J. Plankton Res. 1998, 20, 243–256. [Google Scholar] [CrossRef][Green Version]
- Amundsen, P.-A.; Siwertsson, A.; Primicerio, R.; Bøhn, T. Long-term responses of zooplankton to invasion by a planktivorous fish in a subarctic watercourse. Freshw. Biol. 2009, 54, 24–34. [Google Scholar] [CrossRef]
- Kjellberg, G.; Hegge, O.; Lindstrøm, E.-A.; Løvik, J. Tiltaksorientert Overvåking av Mjøsa Med Tilløpselver. Årsrapport for 1998; Norwegian Institute of Water Research: Oslo, Norway, 1999; p. 96. [Google Scholar]
- Sandlund, O.T.; Linløkken, A.; Gjelland, K.Ø.; Johnsen, S.I.; Rognerud, S.; Museth, J.; Dokk, J.G.; Garmo, Ø.A.; Walseng, B. Fiskesamfunnet i Osensjøen, Trysil og Åmot Kommuner, Hedmark. Status og Endringer Siden 1970-åra; Research, N.I.F.N., Ed.; Norwegian Institute for Nature Research: Trondheim, Norway, 2014; p. 54. [Google Scholar]
- Museth, J.; Johnsen, S.I.; Eloranta, A.; Sandlund, O.T.; Linløkken, A.; Bærum, K.M.; Dokk, J.G. Fiskesamfunnet i Storsjøen i 2016. Effekter av Reguleringsinngrep, Fiske og Introdusert Krøkle; Norwegian Institute of Nature Research: Lillehammer, Norway, 2017; p. 43. [Google Scholar]
- Museth, J.; Sandlund, O.T.; Johnsen, S.I.; Rognerud, S.; Saksgård, R.J. Fiskesamfunnet i Storsjøen i Åmot og Rendalen; Norwegian Institute of Nature Research: Lillehammer, Norway, 2008; p. 63. [Google Scholar]
- Skulberg, O.; Kotai, J. Haldenvassdraget—Vannkvalitet og Forurensningsvirkninger. Resultater av Vassdragsundersøkelser for Haldenvassdragets Vassdragsforbund 1975–1981; Norwegian Institute of Water Research: Oslo, Norway, 1982; p. 179. [Google Scholar]
- Haande, S.; Hagman, C.H.C. Overvåking av Haldensvassdraget 2012; Norsk Institutt for Vannforskning: Oslo, Norway, 2013; p. 25. [Google Scholar]
- Haande, S.; Rohrlack, T.; Kyle, M. Utvikling av Vannkvalitet i Haldenvassdraget. Sammenstilling av Lange Tidsserier (1968–2013). Paleolimnologiske Undersøkelser i Bjørkelangen og Hemnessjøen; Norwegian Institute of Water Research: Oslo, Norway, 2014; p. 6652-2014. [Google Scholar]
- Brabrand, Å. Tetthet, dybdefordeling og Biomasse av Fisk i Bjørklangen og Hemnessjøen, Haldenvassdraget; Laboratorium for ferskvannsøkologi og innlandsfiske (LFI), Zoologisk Museum; University of Oslo: Oslo, Norway, 1993; p. 19. [Google Scholar]
- Brabrand, Å.; Enderlein, O.; Aass, P. Praktiske Vurderinger i for Bindelse Med Kvalitetsforbedring av Lagesildbestander; Laboratorium for ferskvannfisk og innlandsfiske (LFI), Zoologisk museum; Universitetet i Oslo: Oslo, Norway, 2000; p. 15. [Google Scholar]
- Vøllestad, A. Resultat av Prøvefiske i Bjørkelangen, Øgdern og Rødenessjøen nr. 1. In Fiskeribiologiske Undersøkelser i Haldenvassdraget; Fylkesmannen i Østfold: Moss, Norway, 1983; p. 60. [Google Scholar]
- Vašek, M.; Kubečka, J.; Sed’a, J. Cyprinid predation on zooplankton along the longitudinal profile of a canyon-shaped reservoir. Arch. Hydrobiol. 2003, 156, 535–550. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | M o.h. | Surface | Regulated | DepthMax | DepthMean | Sechi d. | Cond. | TP | Tot N | |
|---|---|---|---|---|---|---|---|---|---|---|
| m | Km2 | m | m | m | m | mS/m | µg/L | µg/L | ||
| 1 | Øymarksjøen | 107 | 13.6 | 1.0 | 35 | 16 | 1.6 | 5.4 | 14.4 | 867 |
| 2 | Rødenessjøen | 118 | 15.5 | 0.9 | 47 | 20 | 2.1 | 5.4 | 19.6 | 874 |
| 3 | Hemnessjøen | 133 | 12.7 | 1.4 | 35 | 10 | 1.6 | 7.1 | 23.1 | 494 |
| 4 | Rømsjøen | 138 | 13.7 | 0 | 100 | 24 | 4 | 3.3 | 8 | 415 |
| 5 | Sperillen | 150 | 37.3 | 2.3 | 129 | 43 | 5.0 | 2.2 | 4.8 | 314 |
| 6 | Randsfjorden | 135 | 140.7 | 3.2 | 131 | 52 | 7.0 | 3.9 | 8.8 | 389 |
| 7 | Dokkfløy | 735 | 9.5 | 65 | 53 1 | - | 5.0 | 1.6 | 4.5 | 266 |
| 8 | Einavatnet | 398 | 13.7 | 2.3 | 56 | - | 5.5 | 9.0 | 7 | 1200 |
| 9 | Gopollvatnet | 982 | 1.47 | 2.2 | 20 1 | - | 4 | 1.7 | 3.2 | - |
| 10 | Furusjøen | 852 | 5.3 | 0 | 23 1 | - | 7 | 1.6 | 3.8 | 117 |
| 11 | Rondvatnet | 1167 | 0.96 | 0 | 55 | - | 6 | 0.42 | 3 | 149 |
| 12 | Storsjøen Odal | 130 | 44.3 | 0 | 17 | 7 | 3.5 | 2.6 | 11 | 390 |
| 13 | Osensjøen | 439 | 45.1 | 6.6 | 117 | 37 | 4 | 1.8 | 7.9 | 265 |
| 14 | Storsjøen Rendal | 251 | 48.1 | 3.6 | 309 | 145 | 5 | 4.1 | 6.7 | 390 |
| 15 | Atnsjøen | 701 | 4.8 | 0 | 80 | 35 | 7.5 | 0.78 | 7 | 177 |
| 16 | Engeren | 472 | 14.5 | 0 | 80 | - | 5 | 3.7 | 5.3 | 168 |
| 17 | Sølensjøen | 688 | 22.6 | 0 | 58 | - | 5 | 1.3 | 4 | 165 |
| Algae | Zooplankton | Approximately Biomass Distribution of Herbivorous Plankton Groups | ||||
|---|---|---|---|---|---|---|
| Lake | References | mg Ww/m3 | g Dw/m2 | 20–50% | 10–20% | <10% |
| Øymarksjøen | [32] | - | - | B. coregoni, B. longirostris | B. coregoni, B. longispina | |
| Rødenessjøen | [32,33,34] | 281–1050 | - | B. coregoni, B. longirostris | B. longispina | |
| Hemnessjøen | [33,34] | 1200–1818 | - | D. cristata | B. coregoni, B. lilljeborgii | B. lingispina |
| Rømsjøen 1988 | [35] | 68–478 | 0.06–0.13 | Bosmina spp. | ||
| Sperillen | [36,37] | 103–349 | 0.3–0.5 | D. galeata, B. longispina | H. gibberum | E. gracilis, D. cristata |
| Randsfjorden 88 | [38,39] | 47–158 | 0.4–1.5 | E. gracilis | D. galeata, B. longispina | H. gibberum. D. cristata |
| Dokkfløy 1998/2004 | [38,39] | 64–164 | 0.2–2.8 | D. galeata, H. gibberum | D. cristata, B. longispina | D. longispina |
| Einavatnet 1988–2000 | [35,40] | 207–518 | 1.6–2.5 | D. galeata, B. longispina | D. longispina, H. gibberum | |
| Einavatnet 2013 | Unpubl. | - | - | D. cristata | D. galeata | D. longispina |
| Gopollen | Unpubl. | - | - | D. cristata, | B. longispina | H. gibberum |
| Furusjøen | [41] | - | - | D. lacustris, B. longispina | H. gibberum | |
| Rondvatnet | [42] | - | - | B. longispina | ||
| Storsjøen Odalen 1988–2013 | [43,44,45] | 634–734 | 0.8 | B. longispina | D. cristata, H. gibberum, B. coregoni | |
| Osensjøen 1988 | [35] | 88 | 1.0 | H. gibberum | D. galeata | |
| Osensjøen 2005 | [46] | - | 0.7–1.7 | D. cristata, B. longispina | D. longiremis | D. galeata, H. gibberum |
| Osensjøen 2011 | [35,45] | 133–238 | D. cristata, B. longispina | D. galeata | D. longiremis | |
| Storsjøen Rendalen 1985 | [47,48] | 38–247 | 0.6–0.9 | B. longispina | D. galeata | D. cristata, H. gibberum |
| Storsjøen Rendalen 2011 | [45] | 466–1034 | - | B. longispina | D. galeata | H. gibberum |
| Storsjøen Rendalen 2016/17 | [44] | 178–370 | - | B. longispina | D. galeata | D. longiremis |
| Atnsjøen | [49,50,51] | 143–180 | 1.7 | B. longispina | D. longispina, H. gibberum | |
| Engern 1983 | [52] | 103–286 | 0.1–0.4 | B. longispina, D. galeata | D. cristata | |
| Engern 2007/2011 | [45] | 143–335 | 0.7 | D. galeata | B. longispina | |
| Sølensjøen | [49,53] | 116–247 | - | Eubosmina longispina | D. galeata, H. gibberum | |
| Lake | References | Daphnia spp. | D. Galeata | D. Cristata | B. Longispina | Bosmina spp. | Pelagic Fish Species |
|---|---|---|---|---|---|---|---|
| Øymarksjøen | Unpubl. | - | 0.46 | Vendace, smelt, bleak, roach, bream, white bream roach, bleak | |||
| Rødenessjøen | Ref. [33] | - | 0.90 | 0.46 | Vendace, smelt, bleak, bream, white bream, roach | ||
| Hemnessjøen | Ref. [33] | - | 0.78 | 0.48 | Smelt, bleak, bream, white bream, roach | ||
| Rømsjøen | Unpubl. | - | 0.50 | Smelt, whitefish, vendace, roach, bleak | |||
| Sperillen | Unpul. | - | Whitefish, smelt, perch | ||||
| Randsfjorden | Ref. [38] | - | 1.35 | 1.08 | 0.61 | Whitefish, smelt, perch | |
| Dokkfløy | Ref. [38] | - | 1.72 | 1.06 | 0.61 | Whitefish | |
| Einavatnet 2000 | Ref. [40] | - | 0.85 | 0.52 | Whitefish, smelt, perch, roach | ||
| Einavatnet 2015 | Unpubl. | - | 0.96 | 0.87 | 0.49 | Whitefish, smelt, perch, roach | |
| Gopollen | Unpubl. | - | 0.80 | 0.54 | Whitefish | ||
| Furusjøen | Ref. [41] | 1.81 1 | 0.73 | Arctic charr | |||
| Rondvatn | - | - | - | - | - | Arctic charr | |
| Storsjøen Odalen | Ref. [43] | - | 0.96 | 0.47 | Whitefish, smelt, roach, bleak | ||
| Osensjøen 2005 | Ref. [46] | - | 1.13 | 0.63 | Vendace, whitefish | ||
| Osensjøen 2011 | Ref. [45] | - | 1.53 | 1.10 | 0.66 | Vendace, whitefish | |
| Osensjøen 2018 | Unpubl. | - | 0.91 | 0.90 | 0.47 | Vendace, whitefish | |
| Storsjøen Rendalen 2011 | Ref. [45] | - | 1.50 | 0.72 | (4 Smelt, sparse), whitefish, arctic charr, | ||
| Storsjøen Rendalen 2016 | Ref. [44] | 1.15 2 | 0.66 | 4 Smelt, whitefish, arctic charr, | |||
| Storsjøen Rendalen 2017 | Unpubl. | - | 0.98 | 0.61 | 4 Smelt, whitefish, arctic charr, | ||
| Atnsjøen | Ref. [59] | 1.62 3 | 0.60 | Arctic charr | |||
| Engern 2011 | Ref. [45] | - | 1.53 | 0.47 | Whitefish, arctic charr | ||
| Sølensjøen | Ref. [45] | - | 1.45 | - | 0.73 | Whitefish, arctic charr |
| Lake | Year | N/ha | 95% CI | Lm > 4 cm | Lm > 8 cm | B/ha |
|---|---|---|---|---|---|---|
| Øymarksjøen | 2014 10D | 1445 | 1116–1842 | 13.0 | 16 | 31.3 |
| Rødenessjøen | 2013 10D | 1486 | 1286–1670 | 13.0 | 13 | 41.8 |
| Hemnessjøen | 2018 10D | 4720 | 3956–5484 | 7.0 | 14 | 232 |
| Rømsjøen | 2014 10D | 553 | 433–638 | 12.6 | 13 | 17.4 |
| Sperillen | 2016 9D | 269 | 211–337 | 14.1 | 22 | 16.1 |
| Randsfjorden | 20159N | 270 | 162–970 | 8.0 | 12 | 21.3 |
| Dokkfløy | 2018 8N | 34 | 13–95 | 14.1 | 28 | 10.6 |
| Einavatn | 1990 5D | 610 | 494–726 | 7.9 | 13 | 41.8 |
| 1996 5D | 1240 | 908–1573 | 10.0 | 13 | 44.7 | |
| 2013 5D | 1855 | 1193–2884 | 12.6 | 16 | 62.9 | |
| Gopollvatnet | 2018 8N | 557 | 166–1617 | 12.6 | 20 | 54.5 |
| Furusjøen | 2017 8D | 153 | 58–396 | 5.0 | 11.5 | 1.0 |
| Rondvatn | 2017 8D | 169 | 42–662 | 8.0 | 16.0 | 9.5 |
| Storsjøen Odal | 2013 5D | 1841 | 1769–1914 | 10.0 | 13 | 28.4 |
| Osensjøen mean | 1986–19985D | 79 | 49–109 | 25.0 | 25 | 2.7 |
| 2011 5D | 373 | 743–403 | 12.6 | 28.5 | 37.0 | |
| 2018 9N | 689 | 613–765 | 10.0 | 16 | 24.1 | |
| Storsjøen Rendalen | 1985 6N | 26 | 22–29 | 5 | 25 | 3.6 |
| 1985 8N | 79 | 62–96 | 5 | 25 | 6.0 | |
| 1986 5D | 116 | 88–145 | 25.1 | 32 | 28.2 | |
| 1988 5D | 151 | 131–171 | 16.0 | 20 | 27.3 | |
| 2013 5D | 109 | 92–126 | 8.0 | 16 | 9.4 | |
| 2016 5D | 809 | 749–870 | 13.0 | 16 | 21.2 | |
| Atnsjøen | 2018 9N | 65 | 26–159 | 6.3 | 20 | 12.2 |
| Engeren | 1985 10D | 156 | 147–165 | 32.0 | 32 | 34.5 |
| 2014 5D | 189 | 172–206 | 31.6 | 32 | 31.7 | |
| Sølensjøen | 1985 7N | 107 | 94–120 | 10.0 | 20.0 | 7.1 |
| 1992 6N | 464 | 341–587 | 4.0 | 20.0 | 3.8 | |
| 1992 9N | 159 | 124–194 | 13.0 | 16.0 | 6.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linløkken, A.N. Effects of Lake Productivity on Density and Size Structure of Pelagic Fish Estimated by Means of Echosounding in 17 Lakes in Southeast Norway. Sensors 2021, 21, 3391. https://doi.org/10.3390/s21103391
Linløkken AN. Effects of Lake Productivity on Density and Size Structure of Pelagic Fish Estimated by Means of Echosounding in 17 Lakes in Southeast Norway. Sensors. 2021; 21(10):3391. https://doi.org/10.3390/s21103391
Chicago/Turabian StyleLinløkken, Arne N. 2021. "Effects of Lake Productivity on Density and Size Structure of Pelagic Fish Estimated by Means of Echosounding in 17 Lakes in Southeast Norway" Sensors 21, no. 10: 3391. https://doi.org/10.3390/s21103391
APA StyleLinløkken, A. N. (2021). Effects of Lake Productivity on Density and Size Structure of Pelagic Fish Estimated by Means of Echosounding in 17 Lakes in Southeast Norway. Sensors, 21(10), 3391. https://doi.org/10.3390/s21103391