Using Eddy Covariance Sensors to Quantify Carbon Metabolism of Peatlands: A Case Study in Turkey

Abstract
: Net ecosystem exchange (NEE) of carbon dioxide (CO2) was measured in a cool temperate peatland in northwestern Turkey on a continuous basis using eddy covariance (EC) sensors and multiple (non-)linear regression-M(N)LR-models. Our results showed that hourly NEE varied between −1.26 and 1.06 mg CO2 m−2 s−1, with a mean value of 0.11 mg CO2 m−2 s−1. Nighttime ecosystem respiration (RE) was on average measured as 0.23 ± 0.09 mg CO2 m−2 s−1. Two best-fit M(N)LR models estimated daytime RE as 0.64 ± 0.31 and 0.24 ± 0.05 mg CO2 m−2 s−1. Total RE as the sum of nighttime and daytime RE ranged from 0.47 to 0.87 mg CO2 m−2 s−1, thus yielding estimates of gross primary productivity (GPP) at −0.35 ± 0.18 and −0.74 ± 0.43 mg CO2 m−2 s−1. Use of EC sensors and M(N)LR models is one of the most direct ways to quantify turbulent CO2 exchanges among the soil, vegetation and atmosphere within the atmospheric boundary layer, as well as source and sink behaviors of ecosystems.1. Introduction
Though spatially small (5% of the terrestrial biosphere) compared with most other ecosystems [1,2], peatlands play a significant role in carbon (C) and water metabolism of the World. Understanding and quantifying C dynamics of peatlands are crucial to prediction of responses to global climate change and rehabilitation of peatlands under the increasing magnitude and rate of human-induced disturbances. Eddy Covariance (EC) sensors are one of the most direct ways to measure and estimate turbulent carbon dioxide (CO2), water vapor and energy fluxes exchanged among soil, vegetation, and atmosphere within the atmospheric boundary layer. The use of the EC method and sensors on a long-term and continuous basis across the World has led to the establishment of an integrated global network for standardization of flux tower activities (called FLUXNET) and a network for standardization and development of spectral sensors toward bridging the gap between remote and proximal sensing (called SpecNET).
Our study area in Turkey, the Yenicaga peatland area that at one time occupied 240 km2, has diminished to less than 30 km2 due to drainage, cultivation, afforestation, and peat mining [3,4]. There is a lack of information about C metabolism of peatland ecosystems in Turkey, and this study is the first to comprehensively determine C dynamics and components in one of the remaining major peatlands. The objective of this study was to quantify the rate, magnitude, and timing of CO2 exchange between the atmosphere and Yenicaga peatland using the EC sensors and multiple (non-)linear regression-M(N)LR-models.
2. Study Site and Methodology
2.1. Description of Study Site
The Yenicaga peatland is located about 38 km east of the city of Bolu (40°47N′, 32°1′E) in the western Black Sea region of Turkey (Figure 1).
An EC flux tower site was installed about 1 km north of Lake Yenicaga (18 km2) at the elevation of 988 m above seal level on July 12, 2010. The climate in the Yenicaga region is classified as cool temperate, with a mean annual temperature and precipitation of 10.2 °C and 538 mm, respectively [5]. The Yenicaga peatland is reported to contain Devonian and cretaceous limestone, basaltic tuff, lava, and olistolites, with the uppermost layer consisting of tertiary and quaternary formations [5]. The natural vegetation types of the Yenicaga peatland consist of the following dominant communities: (1) Phragmites australis and Typha domingensis (41.2 ha); and (2) Ranunculus lingua, Acorus calamus, Najas marina, Pedicularis palustris, Senecio paludosus (107.8 ha) [6]. The mean vegetation height around the flux tower is about 0.5 m, and the terrain observed around the flux tower exhibits flat and uniform grasslands.
2.2. Eddy Covariance Flux and Ancillary Measurements
Net ecosystem exchange (NEE) rates of carbon dioxide (Fc, mg m−2 s−1) in Yenicaga peatland were estimated using an eddy covariance (EC) system consisting of an open-path CO2/H2O gas analyzer (LI-7500, Licor Inc., Lincoln, NB, USA), a 3-D sonic anemometer/thermometer (CSAT3, Campbell Scientific Inc., Logan, UT, USA), a data logger (CR3000, Campbell Scientific Inc.), and a 3-m tower on which EC flux sensors were mounted. The distance between the LI-7500 and CSAT3 sensors was 0.15 m, with CSAT3 oriented towards the prevailing wind direction (an azimuth angle of 30° from true north) and LI-7500 vertically rotated 15° towards the footprint.
Eddy fluxes and associated signals were recorded at 10 Hz, block averaged over one hour (h) and corrected for the effects of fluctuations in air density on CO2/H2O fluxes (Fc_wpl and LEwpl with WPL correction) through the online flux computation. EC data were collected swapping two 2-GB Compact Flash cards at 14-to-18-day intervals. The net radiation (Rn), downwelling and upwelling longwave (4 to 50 μm) and shortwave radiation (0.2 to 4 μm) (Rl_dn, Rl_up, Rs_dn, and Rs_up, W m−2) were measured using Kipp & Zonen CNR-4 net radiometers (Kipp & Zonen USA Inc., Bohemia, NY, USA). Air temperature (Ta, °C), and relative humidity (RH, %) were sampled using HMP45C probe (Vaisala, Finland). Precipitation (PPT, mm), evapotranspiration (ET, mm), soil water content (SWC, %), and mean, maximum and minimum soil temperature (ST, STmax, and STmin, °C) were measured on a hourly basis using ET107 weather monitoring station (Campbell Scientific Inc.). Photosynthetically active radiation (PAR) was estimated from net shortwave radiation (Rs_n) using a conversion factor of PAR: Rs_n = 0.5 [7–9]. The values of CO2 fluxes in unit of mg m−2 s−1 were also converted to unit of kg C ha−1day−1, based on the following conversion ratios of mg:kg = 106; s:day = 86,400; m2:ha = 10,000; and CO2:C = 44/12.
2.3. Data Processing and Analyses
As with the conventional meteorological sign notation, downward and upward fluxes are considered negative (−) and positive (±), respectively. The first step of data processing involved the removal of erroneous spikes and their associated CO2 fluxes when latent heat flux (LEwpl) < −100 or > 800 W m−2; sensible heat flux (Hs) < −150 or > 500 W m−2; and precipitation events occurred [10]. For both hourly daytime and nighttime CO2 fluxes, descriptive statistics were given, and best-fit cumulative distribution function (CDF) was selected. Tukey’s multiple comparison was performed after one-way analysis of variance (ANOVA) for the entire dataset in order to test significant differences in hourly, nighttime versus daytime and monthly means. All the statistical analyses were performed with Minitab 15.1 (Minitab Inc. 2006).
Second, EC data were separated into daytime (Rn and/or Rs_dn > 10 W m−2) and nighttime (Rn and/or Rs_dn ≤ 10 W m−2) periods since EC data are more reliable during daytime hours than during nighttime hours, and nighttime EC data ( ) can be used to estimate daytime as well as nighttime ecosystem respiration (both plant and soil respiration) ( ). Negative night CO2 flux data were deleted as no gross primary productivity (GPP = 0) occurs during the nighttime. The site-specific threshold value of friction velocity (u*) was determined as 0.03 m s−1 below which low vertical wind velocity led to underestimation of the nighttime CO2 fluxes [11]. Likewise, nighttime periods where horizontal wind velocity was less than 1 m s−1 were removed from the dataset. Finally, night fluxes where CO2 density data had the standard deviation of > 14 mg m−3 (0.6 μmole m−3) were eliminated from further analyses [12]. Multiple (non-)linear regression models were fitted to the resultant night CO2 dataset, and best-fit M(N)LR models with and without the inclusion of temporal variables (hour, and month) were chosen using best subsets procedure (low Mallows’ Cp, high adjusted R2, and low SE). The following soil respiration equations of RothC [13] and CENTURY [14] models were also used to model daytime RE:
Hourly GPP values were estimated as the sum of NEE and daytime RE. Daytime RE was obtained extrapolating nighttime RE fluxes to the remaining daytime fluxes, based on M(N)LR models.
3. Results and Discussion
3.1. Diurnal CO2 Fluxes
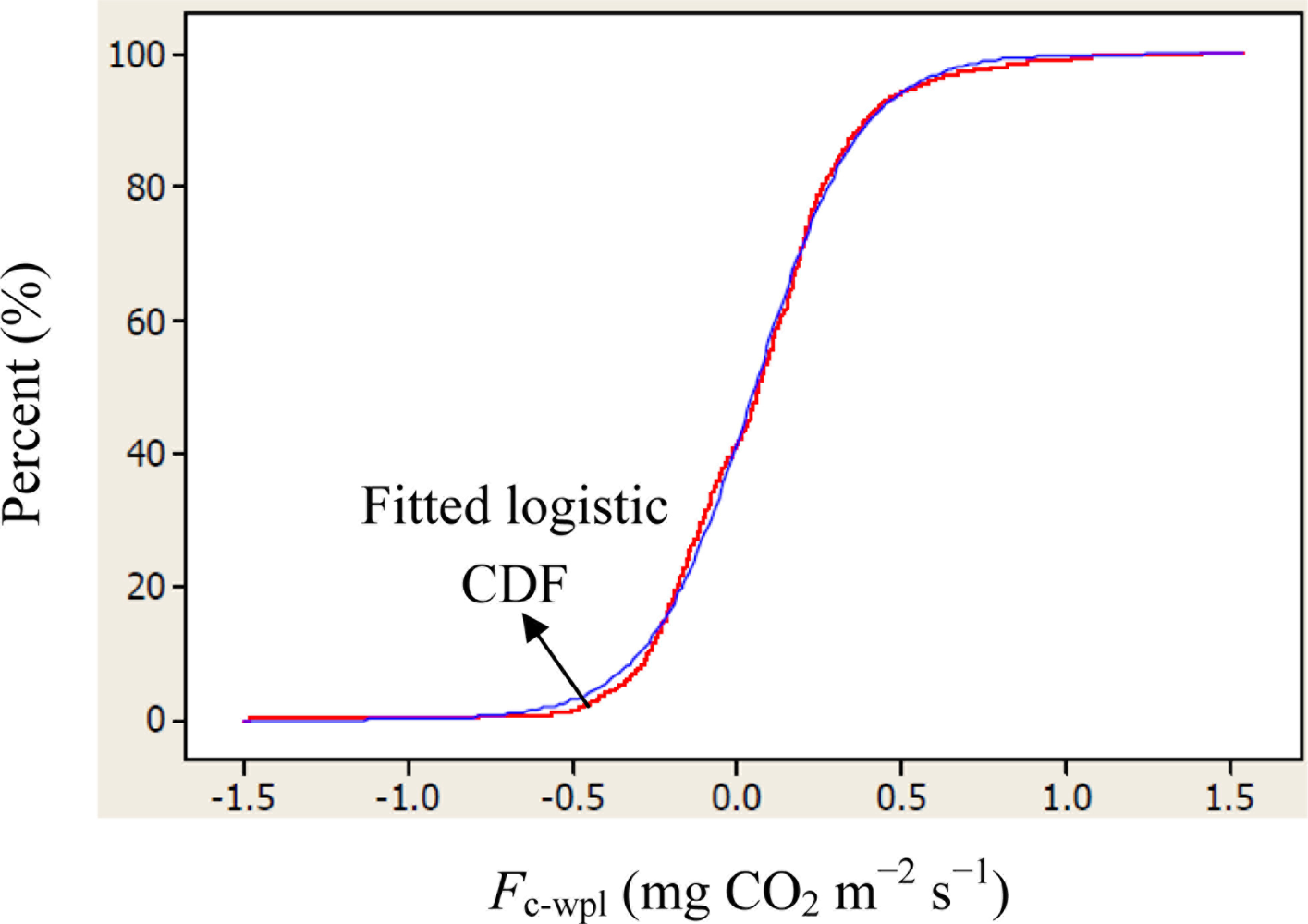
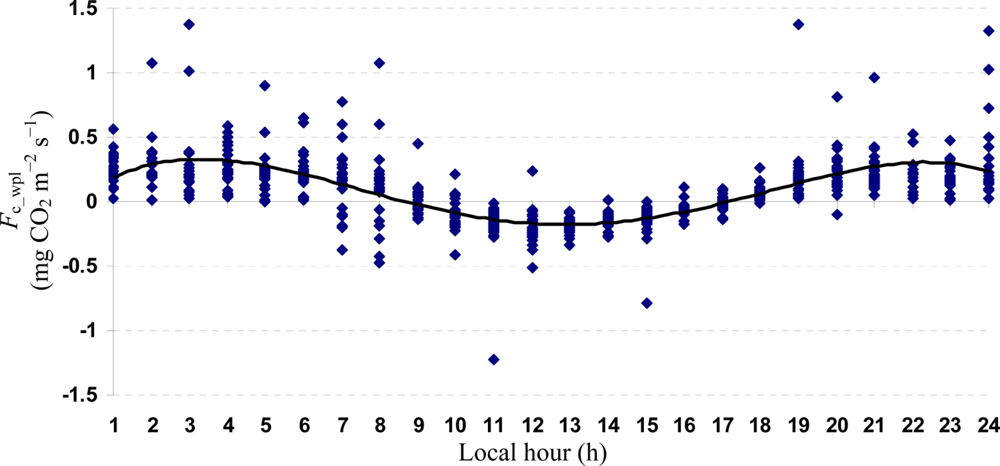
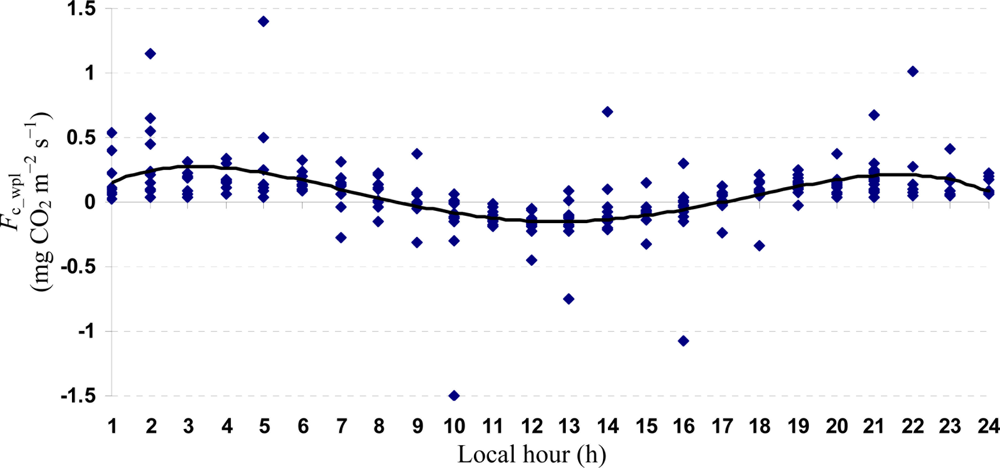
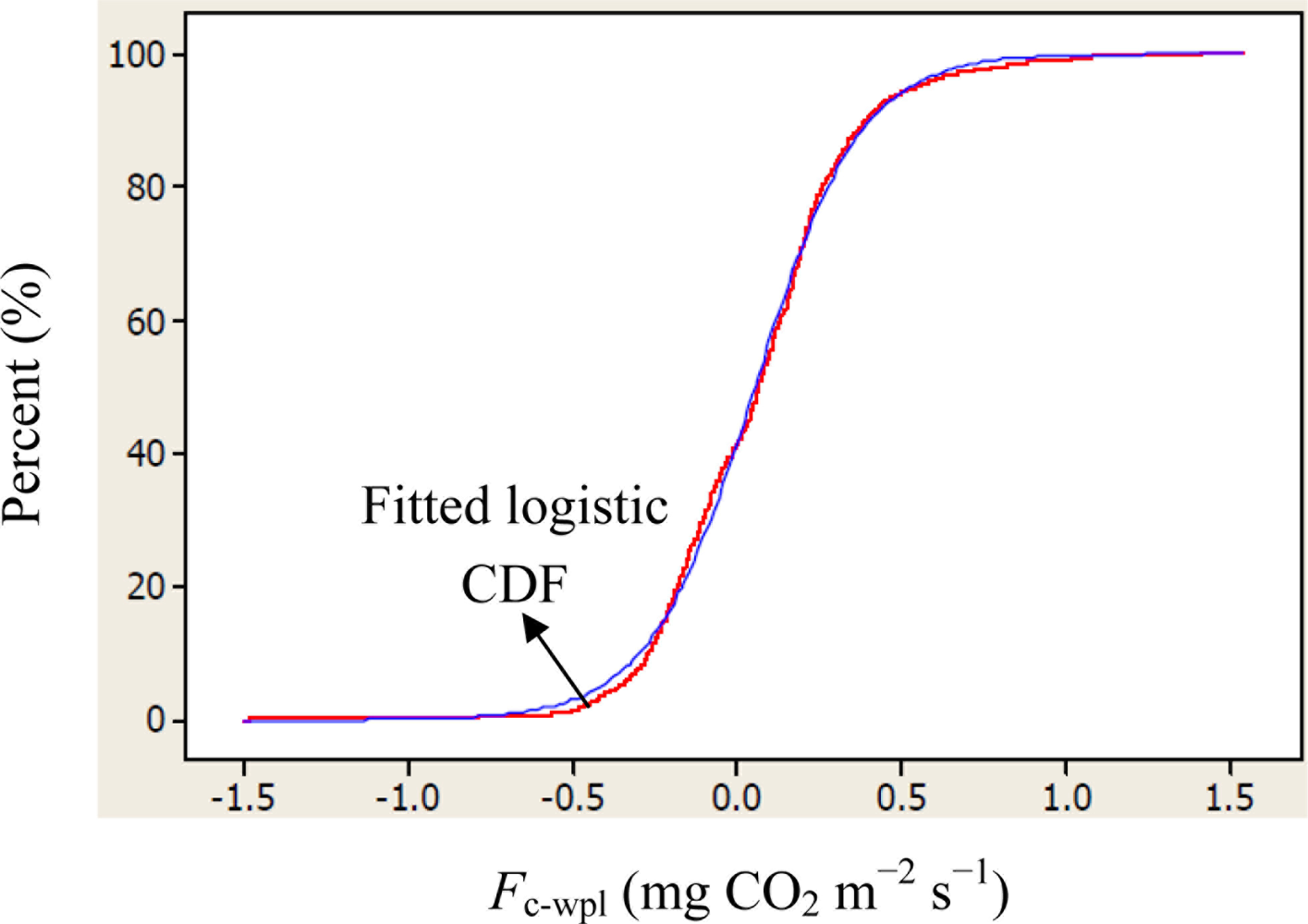
Descriptive statistics of despiked and averaged EC data for the period of July 12 to October 17, 2010 indicated that mean hourly CO2 fluxes above the canopy had a larger temporal variability (CV = 498) than the rest of the measured variables and varied between −1.5 and 1.5 mg m−2 s−1 (Table 1). During the study period, the study site cumulatively received 92 mm PPT and lost 305 mm water vapor via ET. The mean Bowen ratio of Hs:LEwpl quantifying the evaporative demand of an environment was estimated at 0.37, thus characterizing the Yenicaga peatland as a mesic environment. A logistic CDF with a location of 0.05197 and a scale of 0.1596 appeared to fit our daytime and nighttime Fc_wpl data fairly well as follows (r = 0.99; n = 1884; P < 0.001) (Figure 2):
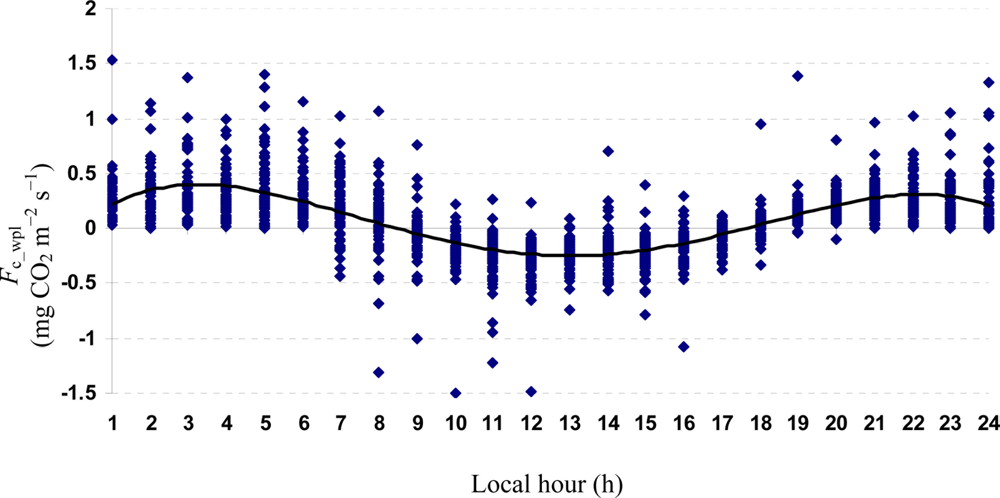
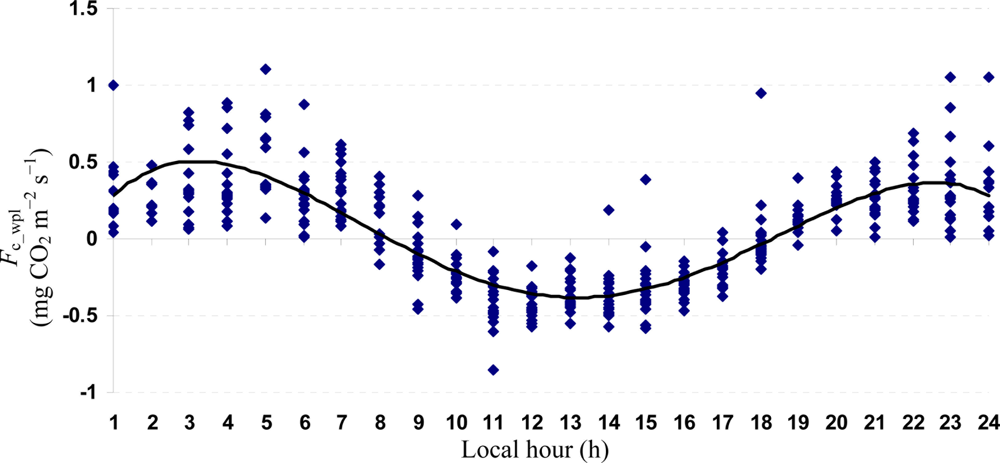
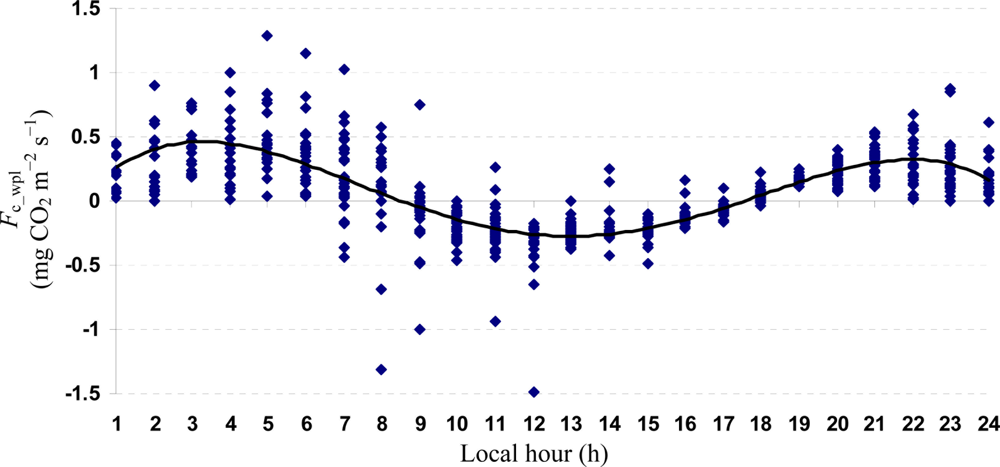
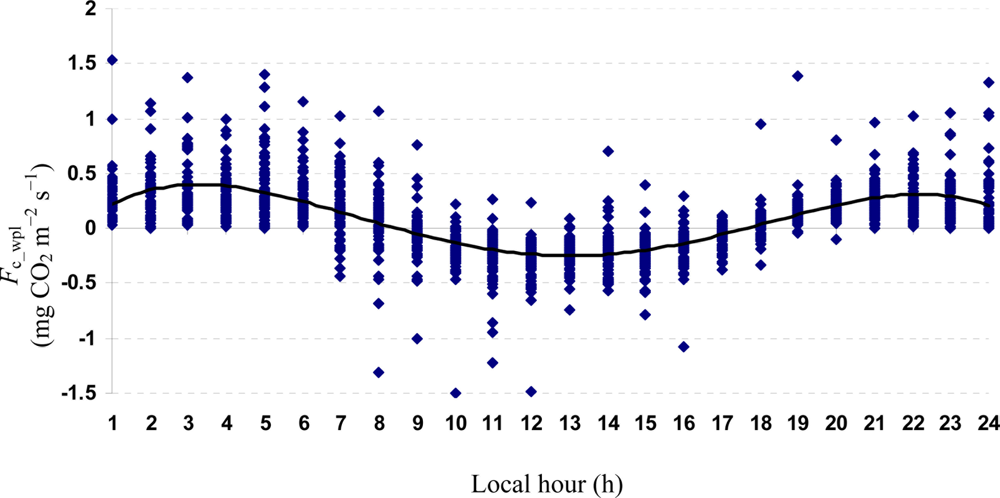
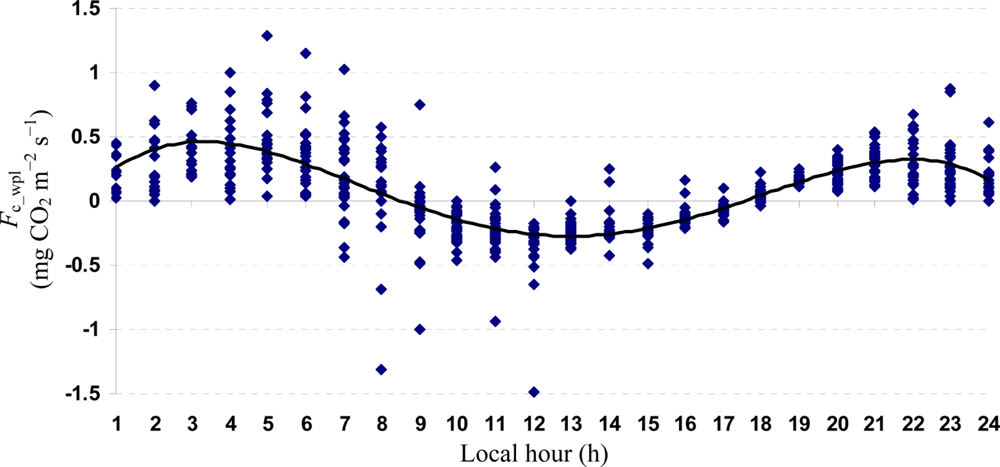
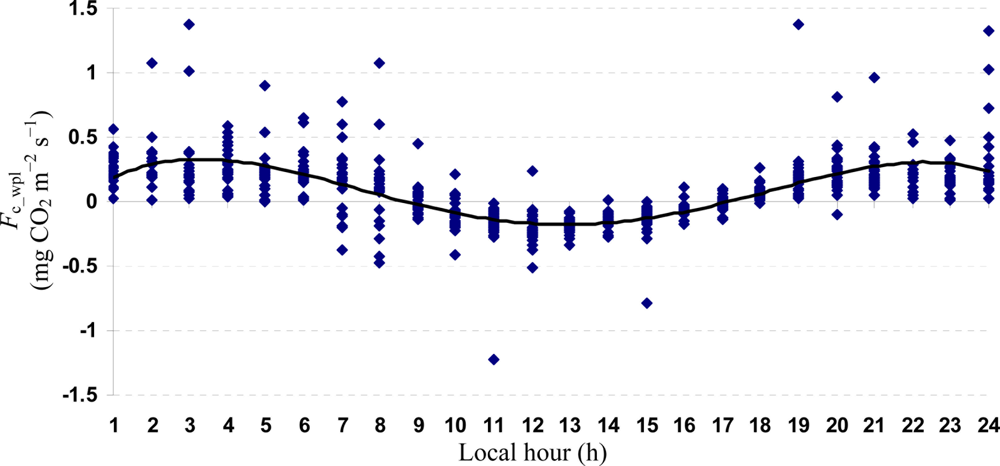
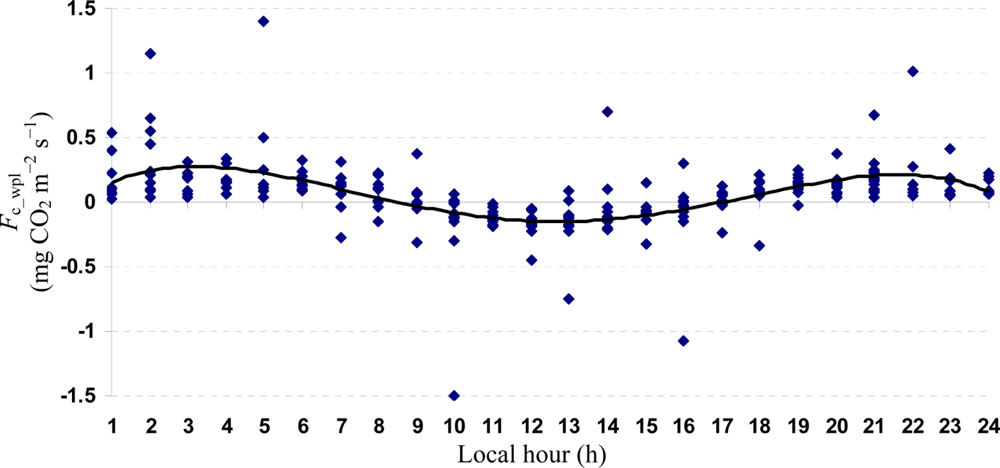
Quartic functions (an equation of fourth degree) fitted to both entire and monthly Fc_wpl datasets as a function of the explanatory temporal variable (h) provided a meaningful representation of CO2 fluxes, with a R2adj value range of 70.9% in July to 31.3% in October (Figures 3 to 7).
3.2. Multitemporal Comparisons of CO2 Fluxes
The entire dataset of daytime and nighttime CO2 fluxes (n = 1,884) was used for a multiple comparison according to hours, nighttime versus daytime, and months, based on Tukey’s test following one-way ANOVA (Table 2). A comparison of hourly mean Fc_wpl values showed that the Yenicaga peatland acted as a CO2 sink for the daytime periods between 9:00 AM and 17:00 PM and acted as a CO2 source for the periods between 18:00 PM and 8:00 AM. On average, the Yenicaga peatland had minimum and maximum CO2 fluxes ranging from −0.05 ± 0.21 mg CO2 m−2 s−1 at 9:00 AM to −0.28 ± 0.18 mg CO2 m−2 s−1 at 12:00 PM as a CO2 sink and from 0.05 ± 0.12 mg CO2 m−2 s−1 at 18:00 PM to 0.38 ± 0.29 mg CO2 m−2 s−1 at 5:00 AM as a CO2 source, respectively (Table 2).
Mean daytime CO2 flux (−0.11 ± 0.22 mg CO2 m−2 s−1) significantly differed from mean nighttime CO2 flux (0.26 ± 0.23 mg CO2 m−2 s−1) (P < 0.001). All the monthly mean CO2 fluxes were positive (upward into the atmosphere) and varied between 0.03 ± 0.35 mg CO2 m−2 s−1 in July and 0.07 ± 0.25 mg CO2 m−2 s−1 in September. However, the monthly mean CO2 fluxes did not appear to significantly differ from one another (Table 2). The best-fit MNLR model was derived from the entire dataset of nighttime and daytime Fc_wpl by exploring possible interaction effects among the EC and meteorological variables as follows:
3.3. Ecosystem Components of Carbon Metabolism
Nighttime RE was modeled using (1) best-fit M(N)LR models with/without the forced inclusion of temporal variables (hour and month), and (2) ST-dependent Rsoil equations [Equations (1) and (2)] of RothC and CENTURY models with/without the forced addition of SWC. All the RE models were built with the intercept forced through zero as follows:
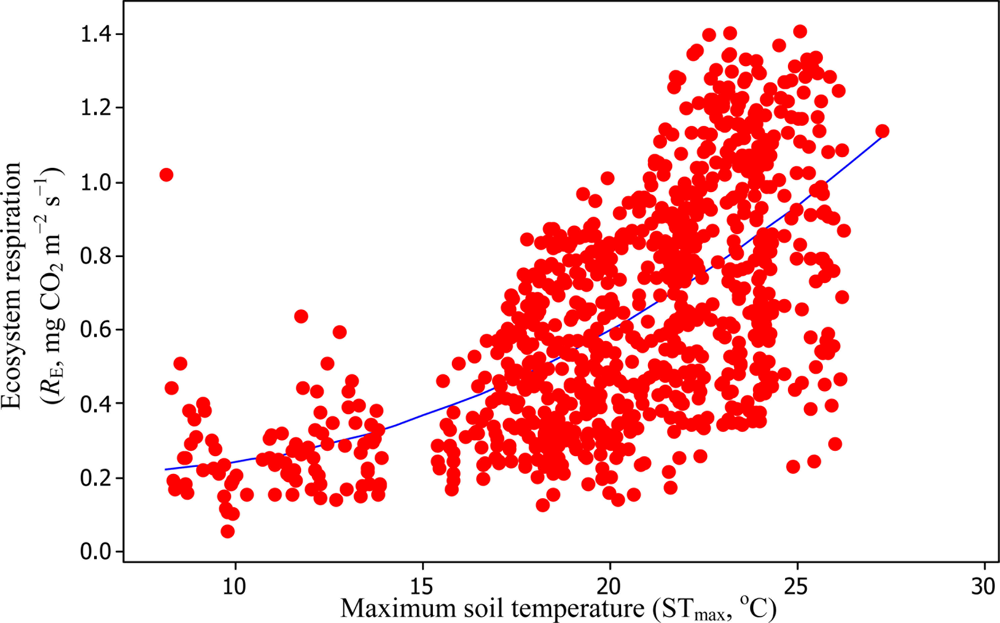
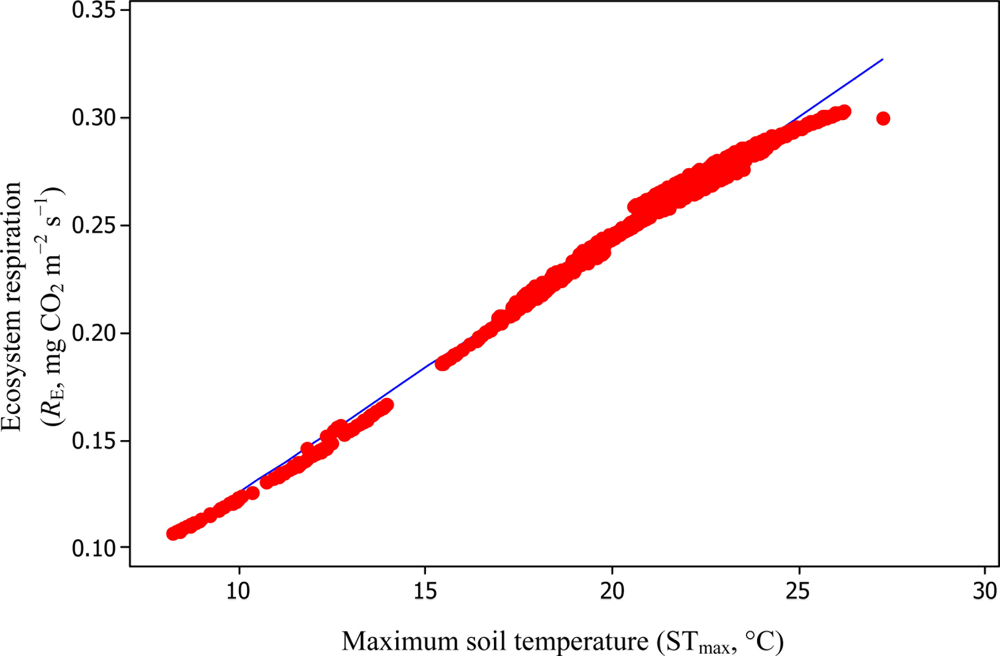
A comparison of daytime RE versus STmax resulted in R2adj values of 39.3% and 99.3% based on Equations (6) and (11), respectively (Figures 10 and 11).
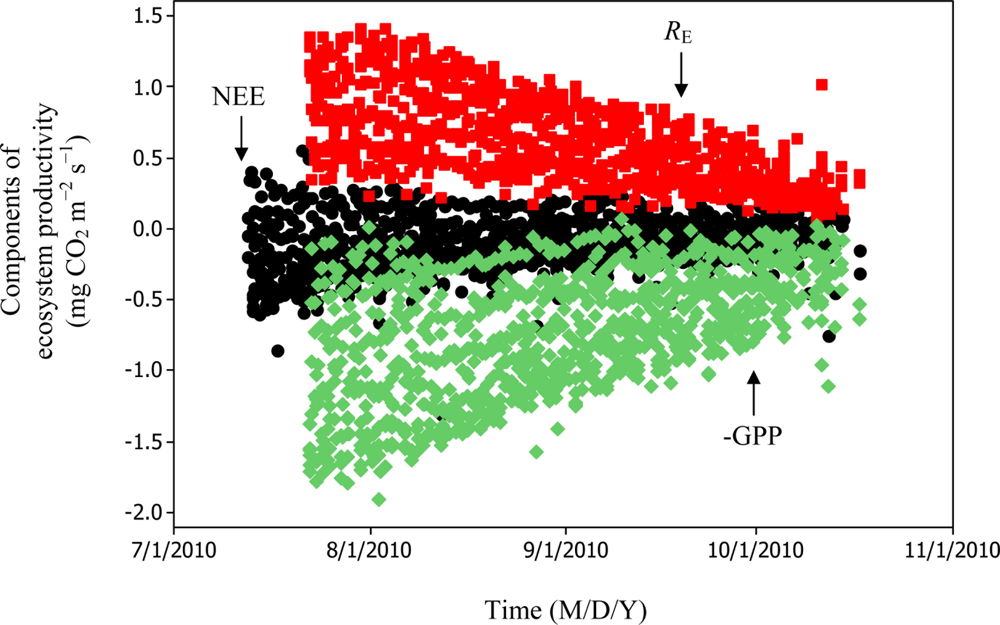
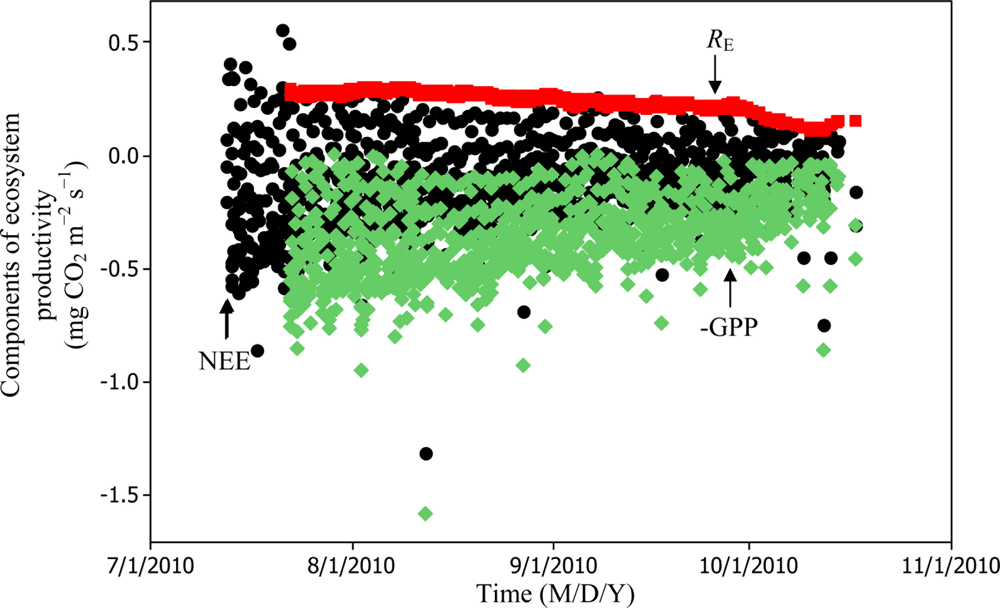
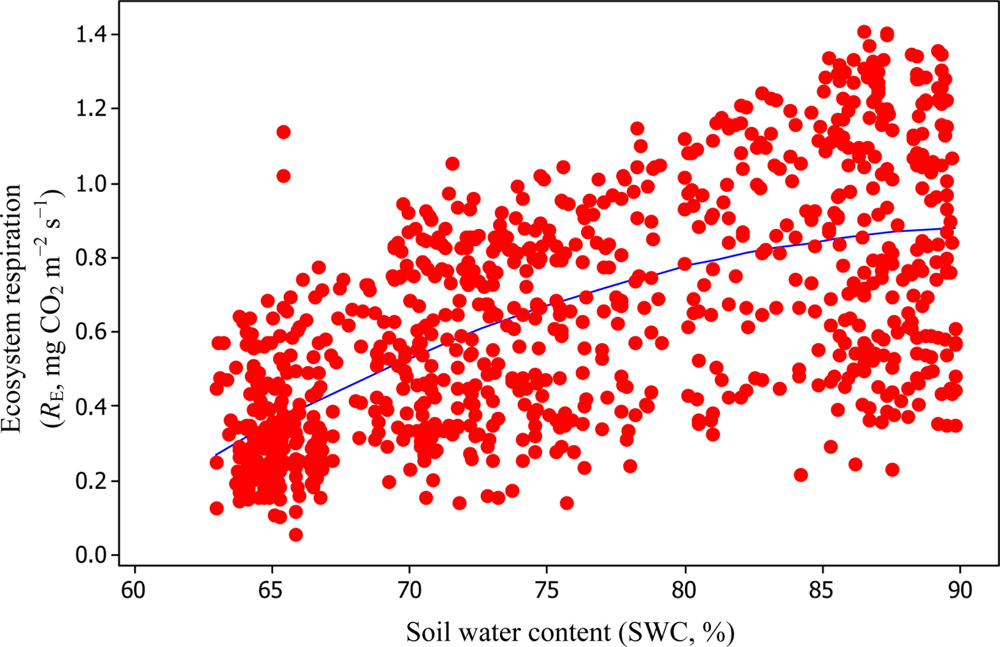
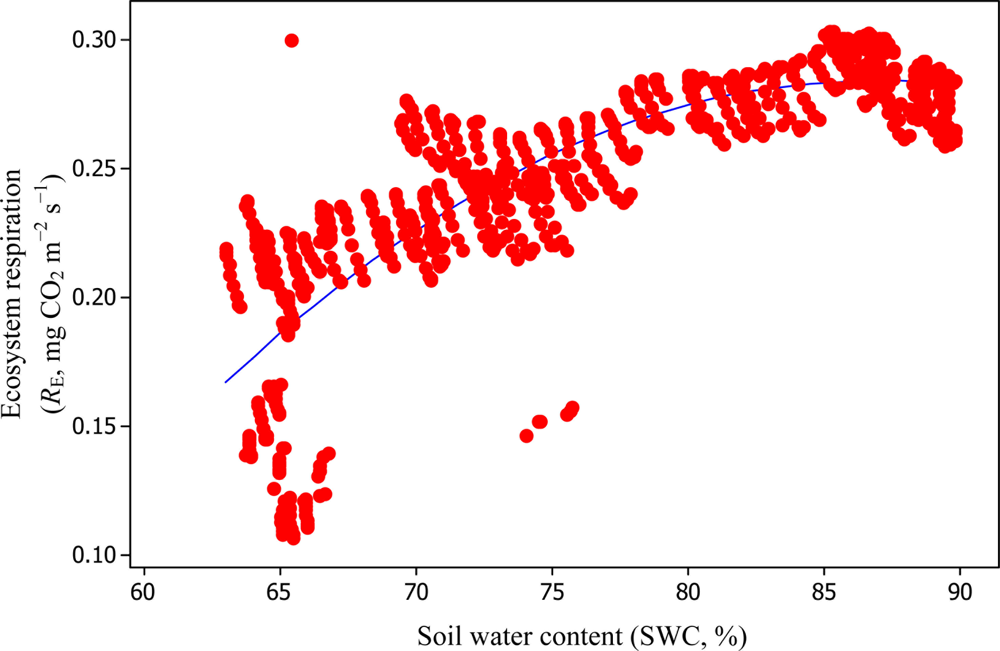
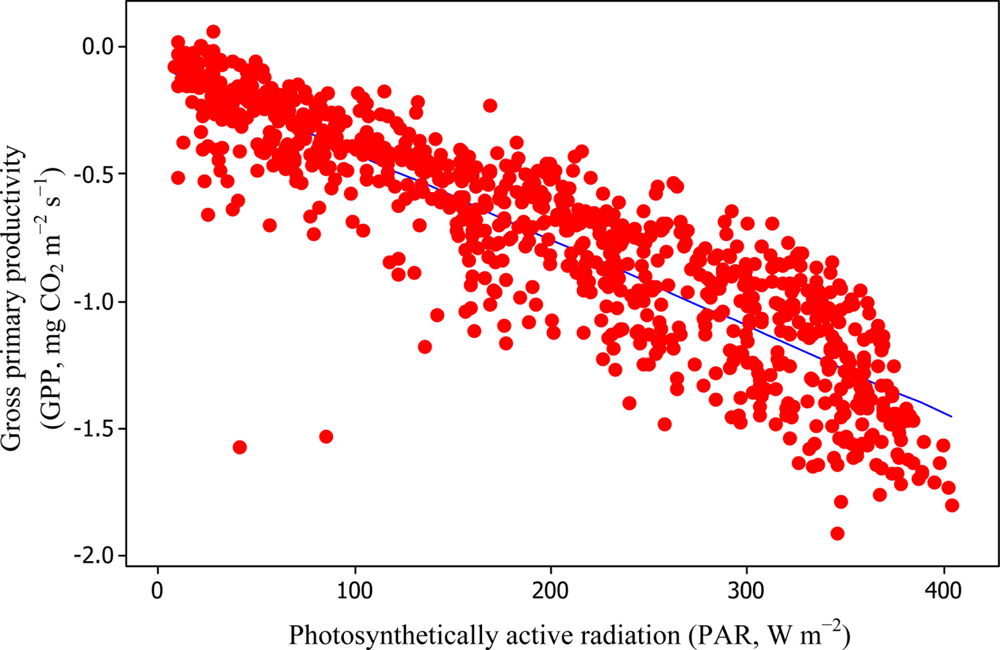
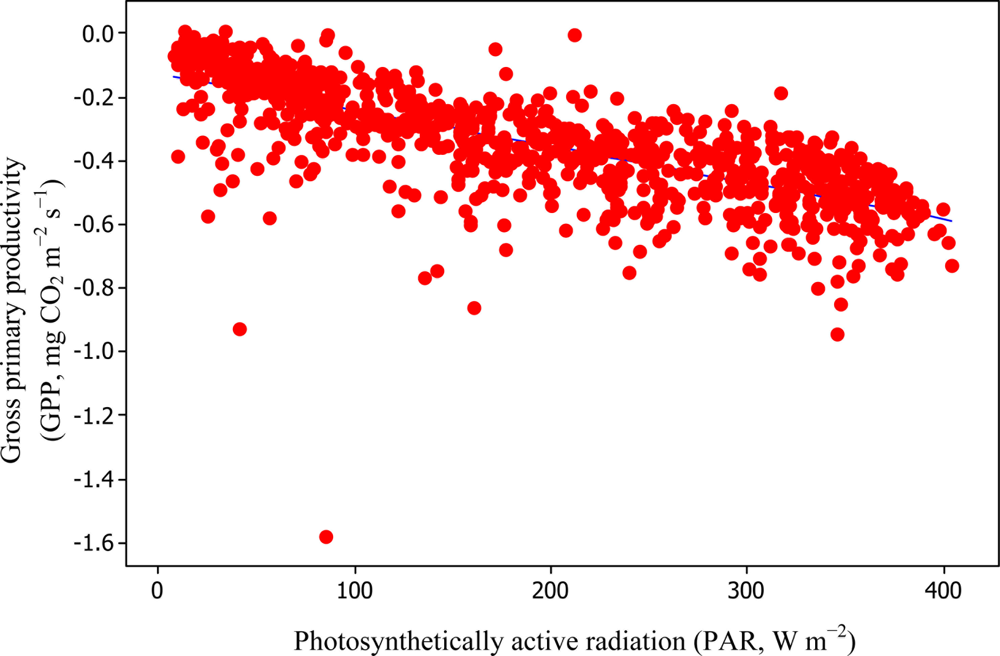
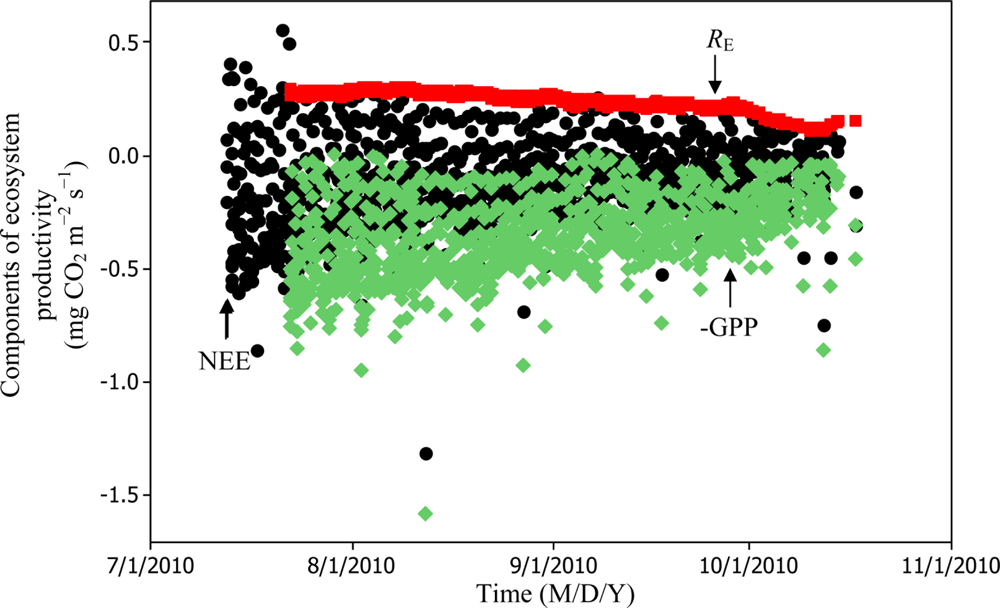
Soil water content was also determined to play a significant role in controlling daytime RE from the Yenicaga peatland. As a function of SWC, quadratic regression models elucidated 39.2% and 68.7% of variation in daytime RE according to Equations (6) and (11), respectively (Figures 12 and 13). A significant relationship between estimated GPP and PAR values was found, yielding R2adj values of 80.3% and 55% from Equations (6) and (11), respectively (Figures 14 and 15). The rate of NEE in the Yenicaga peatland for the study period was on average estimated at 0.11 mg CO2 m−2 s−1 (26.5 kg C ha−1 day−1) and ranged from −1.26 mg CO2 m−2 s−1 (−297 kg C ha−1 day−1) to 1.06 mg CO2 m−2 s−1 (249 kg C ha−1 day−1). This case points to the domination of total (positive daytime ± nighttime fluxes) RE over GPP (negative flux) throughout most of the study period, especially, towards the end of the period during which GPP decreased at a faster rate than RE (Figures 8 and 9). During the peak growing season in July, GPP peaked and decreased as the season progressed. Over the study period, GPP was on average −0.74 ± 0.43 mg CO2 m−2 s−1 (−175 ± 102 kg C ha−1 day−1) and −0.35 ± 0.18 mg CO2 m−2 s−1 (−2 ± 41 kg C ha−1 day−1) according to Equations (6) and (11), respectively. On average, nighttime RE was estimated at 0.23 ± 0.09 mg CO2 m−2 s−1 (53 ± 22 kg C ha−1 day−1) based on the EC sensors, while daytime RE values were 0.64 ± 0.31 mg CO2 m−2 s−1 (150 ± 74 kg C ha−1 day−1) and 0.24 ± 0.05 mg CO2 m−2 s−1 (57 ± 11 kg C ha−1 day−1) based on Equations (6) and (11), respectively.
Peatlands (bog hummock, bog hallow, and poor fen) in a cool temperate region of eastern Canada were reported to have NEE range from −0.18 to 0.13 mg CO2 m−2 s−1; nighttime RE range from 0.07 to 0.36 mg CO2 m−2 s−1; and GPP range from −0.20 to −0.33 mg CO2 m−2 s−1 [15,16]. The reported ranges of NEE, RE and GPP values are in a close agreement with our findings for the Yenicaga peatland [NEE = 0.11 mg CO2 m−2 s−1 based on the EC data; nighttime RE = 0.23 mg CO2 m−2 s−1 based on the EC data; daytime RE = 0.24 mg CO2 m−2 s−1 based on Equation (11); daytime RE = 0.64 mg CO2 m−2 s−1 based on Equation (6); GPP = −0.35 mg CO2 m−2 s−1 based on Equation (11); and GPP = −0.74 mg CO2 m−2 s−1 based on Equation (6)], in particular, based on Equation (11). Similarly, EC measurements from 12 wetlands ranging from ombrotrophic and minerotrophic peatlands to wet tundra ecosystems across Europe and North America under temperate-to-arctic climate regimes showed that CO2 fluxes in July varied considerably between −3 and 1 g C m−2 day−1 for NEE; 1 and 4 g C m−2 day−1 for RE; and −1 and −6 g C m−2 day−1 for GPP [17], which were close to our results.
4. Conclusions
Carbon metabolism components of the Yenicaga peatland as measured by EC sensors and quantified by M(N)LR models clearly revealed that diurnal and seasonal variations in exchange rates of CO2 between the atmosphere and the peatland ecosystem are dependent on the magnitude, rate and timing of GPP and total RE, which are in turn strongly controlled by the dynamics of soil moisture and temperature, and PAR. Peatlands experiencing drought conditions are reported to be able to act as a CO2 source. Given the total PPT: ET ratio of 0.3 during the study period, the Yenicaga peatland is considered to undergo a dry season and signals what future rate and amount of changes may be expected in the face of an increase in ET, air temperature and ecosystem disturbances as well as a decrease in PPT. The present study is the first one to quantify CO2 exchanges for a peatland in Turkey using real-time monitoring by the EC sensors. It is also the first time in Turkey that diurnal source and sink behaviors of a rarely occurring ecosystem like the Yenicaga peatland, which at the same time undergoes severe human-induced pressures such as conversions to rangeland and cropland, and peat mining, were quantitatively assessed using EC sensors. Further research is needed to explore an integration of remote and proximal sensors, and biogeochemical process-based models in improving our understanding and predicting impacts of human-induced disturbances on ecosystem metabolism.
Acknowledgments
We would like to thank TUBITAK and COST Action ES0903 for funding our project (CAYDAG109Y186). We are grateful to Mr. Aydın Gülmez and Staff of Yenicaga Municipality for their logistic support during our field studies. We acknowledge the financial and administrative contributions of Scientific Research Project offices of Abant Izzet Baysal and Akdeniz Universities for the project.
References
- Gorham, E. Northern peatlands: role in the carbon balance and probable responses to climatic warming. Ecol. Appl 1991, 1, 182–195. [Google Scholar]
- Schindler, D.W. The mysterious missing sink. Nature 1999, 398, 105–106. [Google Scholar]
- Cayci, G.; Ataman, Y.; Unver, I.; Munsuz, N. Distribution and horticultural values of the peats in Anatolia. Acta Hortic 1988, 238, 189–196. [Google Scholar]
- Malone, S.; O’Connell, C. Ireland’s Peatland Conservation Action Plan 2020: Halting the Loss of Peatland Biodiversity; Irish Peatland Conservation Council: Kildare, UK, 2009. [Google Scholar]
- Dengiz, O.; Ozaytekin, H.; Cayci, G.; Baran, A. Characteristics, genesis and classification of a basin peat soil under negative human impact in Turkey. Environ. Geol 2009, 56, 1057–1063. [Google Scholar]
- Sumer, N. Flora of Yeniçağa, M.Sc. Thesis,. Graduate School of Abant İzzet Baysal University, Department of Biology, Bolu, Turkey, 2002.
- Daughtry, C.S.T.; Gallo, K.P.; Goward, S.N.; Prince, S.D.; Kustas, W.P. Spectral estimates of absorbed radiation and phytomass production in corn and soybean canopies. Remote Sens. Environ 1992, 39, 141–152. [Google Scholar]
- Running, S.W.; Hunt, E.R. Generalization of a forest ecosystem model. In Scaling Physiological Processes: Leaf to Globe; Ehleringer, J.R., Fields, C.B., Eds.; Academic Press: New York, NY, USA, 1993; pp. 141–157. [Google Scholar]
- Melillo, J.M.; McGuire, A.D.; Kicklighter, D.W.; Moore, B., III; Vorosmarty, C.J.; Schloss, A.L. Global climate change. Nature 1993, 363, 234–240. [Google Scholar]
- Thomas, M.V.; Malhi, Y.; Fenn, K.M.; Fisher, J.B.; Morecroft, M.D.; Lloyd, C.R.; Taylor, M.E.; McNeil, D.D. Carbon dioxide fluxes over an ancient broadleaved deciduous woodland in southern England. Biogeosciences 2010, 7, 3765–3814. [Google Scholar]
- Arneth, A.; Veenendaal, E.M.; Best, C.; Timmermans, W.; Kolle, O.; Montagnani, L.; Shibistova, O. Water use strategies and ecosystem-atmosphere exchange of CO2 in two highly seasonal environments. Biogeosciences 2006, 3, 421–437. [Google Scholar]
- Billesbach, D.P.; Fischer, M.L.; Torn, M.S.; Berry, J.A. A portable eddy covariance system for the measurement of ecosystem—atmosphere exchange of CO2, water vapor, and energy. J. Atmos. Oceanic. Technol 2004, 21, 639–650. [Google Scholar]
- Coleman, K.; Jenkinson, D.S. ROTHC-26.3: A Model for the Turnover of Carbon in Soil. Model Description and Windows Users Guide; Rothamsted Research: Harpenden, UK, 2005; p. 45. [Google Scholar]
- Parton, B.; Ojima, D.; Del Grosso, S.; Keough, C. CENTURY Tutorial. Supplement to CENTURY User’s Manual; Great Plain System. Research Unit Technical Report No. 4 [R]; NREC Pub. Natural Resource Ecology Laboratory, Colorado State University: Fort Collins, CO, USA, 2001. [Google Scholar]
- Moore, T.R.; Bubier, J.L.; Frolking, S.E.; Lafleur, P.M.; Roulet, N.T. Plant biomass and production and CO2 exchange in an ombrotrophic bog. J. Ecol 2002, 90, 25–36. [Google Scholar]
- Lafleur, P.M.; Roulet, N.T.; Bubier, J.L.; Frolking, S.; Moore, T.R. Interannual variability in the peatland-atmosphere carbon dioxide exchange at an ombrotrophic bog. Global Biogeochem. Cy 2003, 17, 1036. [Google Scholar]
- Lund, M.; Lafleur, P.M.; Roulet, N.T.; Lindroth, A.; Christensen, T.R.; Aurela, M.; Chojnicki, B.H.; Flanagan, L.B.; Humphreys, E.R.; Laurila, T.; Oechel, W.C.; Olejnik, J.; Rinne, J.; Schubert, P.; Nilsson, M.B. Variability in exchange of CO2 across 12 northern peatland and tundra sites. Glob. Change Biol 2010, 16, 2436–2448. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | n | mean | ±SD | CV | min | median | max |
|---|---|---|---|---|---|---|---|
| Fc_wpl (mg CO2 m−2 s−1) | 1,884 | 0.06 | 0.30 | 498 | −1.50 | 0.06 | 1.53 |
| Hs(W m−2) | 1,884 | 38.40 | 58.44 | 152 | −78.86 | 3.01 | 238.80 |
| LEwpl (W m−2) | 1,884 | 102.69 | 130.79 | 127 | −92.89 | 41.40 | 517.29 |
| u* (m s−1) | 1,884 | 0.18 | 0.12 | 68 | 0.01 | 0.14 | 0.57 |
| CO2 density (mg m−3) | 1,884 | 633.34 | 118.19 | 19 | 243.45 | 598.57 | 1,577.23 |
| H2O density (g m−3) | 1,884 | 12.34 | 4.77 | 39 | −2.62 | 12.94 | 47.82 |
| Ta (°C) | 1,884 | 18.15 | 7.05 | 39 | −0.67 | 17.95 | 34.00 |
| RH (%) | 1,884 | 71.54 | 22.30 | 31 | 16.09 | 76.13 | 107.80 |
| WD (degree) | 1,884 | 219.91 | 127.29 | 58 | 0.04 | 265.93 | 359.89 |
| WS (m s−1) | 1,884 | 1.73 | 1.40 | 81 | 0.04 | 1.19 | 6.14 |
| Rs_up (W m−2) | 1,884 | 263.80 | 309.91 | 117 | −4.91 | 104.18 | 981.88 |
| Rs_dn (W m−2) | 1,884 | 44.14 | 48.88 | 111 | −0.19 | 18.63 | 153.55 |
| Rl_up (W m−2) | 1,884 | 386.85 | 34.95 | 9 | 294.45 | 386.94 | 474.95 |
| Rl_dn (W m−2) | 1,884 | 413.84 | 47.55 | 11 | 307.63 | 405.96 | 531.50 |
| Rn (W m−2) | 1,884 | 192.66 | 245.44 | 127 | −35.80 | 68.69 | 766.03 |
| ET (mm h−1) | 1,680 | 0.14 | 0.21 | 124 | 0.0 | 0.02 | 0.75 |
| PPT (mm h−1) | 1,680 | 0.04 | 0.36 | 825 | 0.0 | 0.0 | 7.8 |
| STmin (°C) | 1,680 | 19.66 | 3.92 | 20 | 8.11 | 20.03 | 26.18 |
| STmax (°C) | 1,680 | 19.84 | 3.96 | 20 | 8.19 | 20.24 | 27.24 |
| SWC (%) | 1,680 | 75.37 | 8.73 | 12 | 62.98 | 73.75 | 89.90 |
Fc_wpl = WPL-corrected CO2 flux; Hs = sensible heat flux; LEwpl = WPL-corrected latent heat flux; u* = friction velocity; Ta = air temperature; RH = relative humidity; WD = wind direction; WS = wind speed; Rs_up = upwelling shortwave radiation; Rs_dn = downwelling shortwave radiation; Rl_up = upwelling longwave radiation; Rl_dn = downwelling longwave radiation; Rn = net radiation; ET = evapotranspiration; STmin = minimum soil temperature; STmax = maximum soil temperature; SWC = soil water content; CV = coefficient of variation; and SD = standard deviation.
| Local hour | n | mean | SD | Comparison based on 95% confidence intervals |
|---|---|---|---|---|
| 1:00 | 57 | 0.254 | 0.248 | 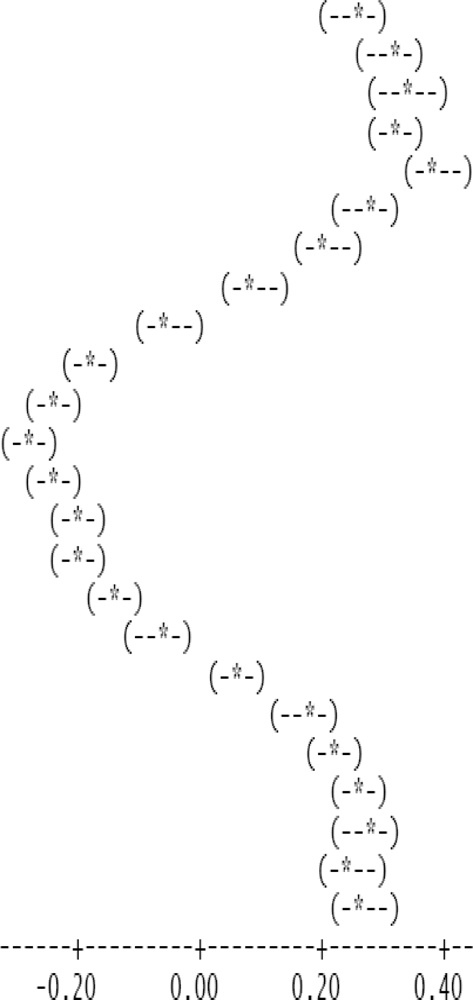 |
| 2:00 | 53 | 0.313 | 0.242 | |
| 3:00 | 55 | 0.337 | 0.277 | |
| 4:00 | 69 | 0.320 | 0.229 | |
| 5:00 | 65 | 0.382 | 0.296 | |
| 6:00 | 72 | 0.276 | 0.215 | |
| 7:00 | 85 | 0.209 | 0.256 | |
| 8:00 | 76 | 0.087 | 0.295 | |
| 9:00 | 82 | −0.053 | 0.210 | |
| 10:00 | 90 | −0.171 | 0.187 | |
| 11:00 | 89 | −0.242 | 0.203 | |
| 12:00 | 91 | −0.280 | 0.187 | |
| 13:00 | 91 | −0.237 | 0.122 | |
| 14:00 | 91 | −0.191 | 0.168 | |
| 15:00 | 89 | −0.200 | 0.156 | |
| 16:00 | 88 | −0.144 | 0.158 | |
| 17:00 | 90 | −0.069 | 0.101 | |
| 18:00 | 91 | 0.052 | 0.128 | |
| 19:00 | 89 | 0.171 | 0.145 | |
| 20:00 | 91 | 0.222 | 0.117 | |
| 21:00 | 85 | 0.262 | 0.149 | |
| 22:00 | 69 | 0.275 | 0.193 | |
| 23:00 | 72 | 0.248 | 0.212 | |
| 24:00 | 54 | 0.267 | 0.266 | |
| Day vs. night | n | mean | SD |  |
| Daytime | 1030 | −0.111 | 0.222 | |
| Nighttime | 852 | 0.266 | 0.235 | |
| Month | n | mean | SD |  |
| July | 412 | 0.039 | 0.351 | |
| August | 620 | 0.054 | 0.307 | |
| September | 603 | 0.077 | 0.257 | |
| October | 249 | 0.064 | 0.248 | |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Evrendilek, F.; Karakaya, N.; Aslan, G.; Ertekin, C. Using Eddy Covariance Sensors to Quantify Carbon Metabolism of Peatlands: A Case Study in Turkey. Sensors 2011, 11, 522-538. https://doi.org/10.3390/s110100522
Evrendilek F, Karakaya N, Aslan G, Ertekin C. Using Eddy Covariance Sensors to Quantify Carbon Metabolism of Peatlands: A Case Study in Turkey. Sensors. 2011; 11(1):522-538. https://doi.org/10.3390/s110100522
Chicago/Turabian StyleEvrendilek, Fatih, Nusret Karakaya, Guler Aslan, and Can Ertekin. 2011. "Using Eddy Covariance Sensors to Quantify Carbon Metabolism of Peatlands: A Case Study in Turkey" Sensors 11, no. 1: 522-538. https://doi.org/10.3390/s110100522
APA StyleEvrendilek, F., Karakaya, N., Aslan, G., & Ertekin, C. (2011). Using Eddy Covariance Sensors to Quantify Carbon Metabolism of Peatlands: A Case Study in Turkey. Sensors, 11(1), 522-538. https://doi.org/10.3390/s110100522