Using Automated Point Dendrometers to Analyze Tropical Treeline Stem Growth at Nevado de Colima, Mexico



Abstract
:
1. Introduction
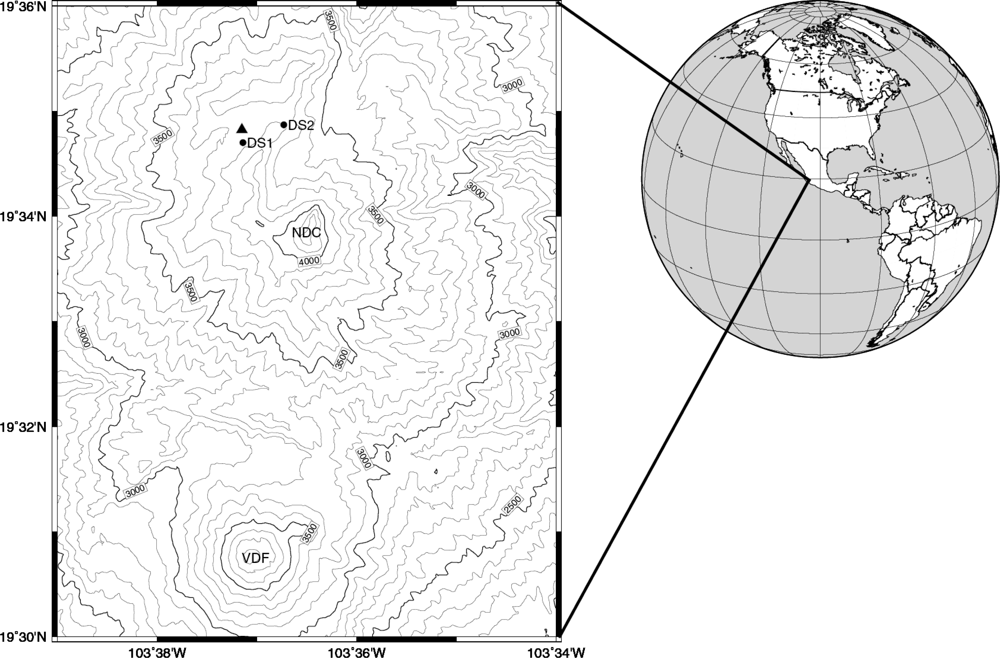
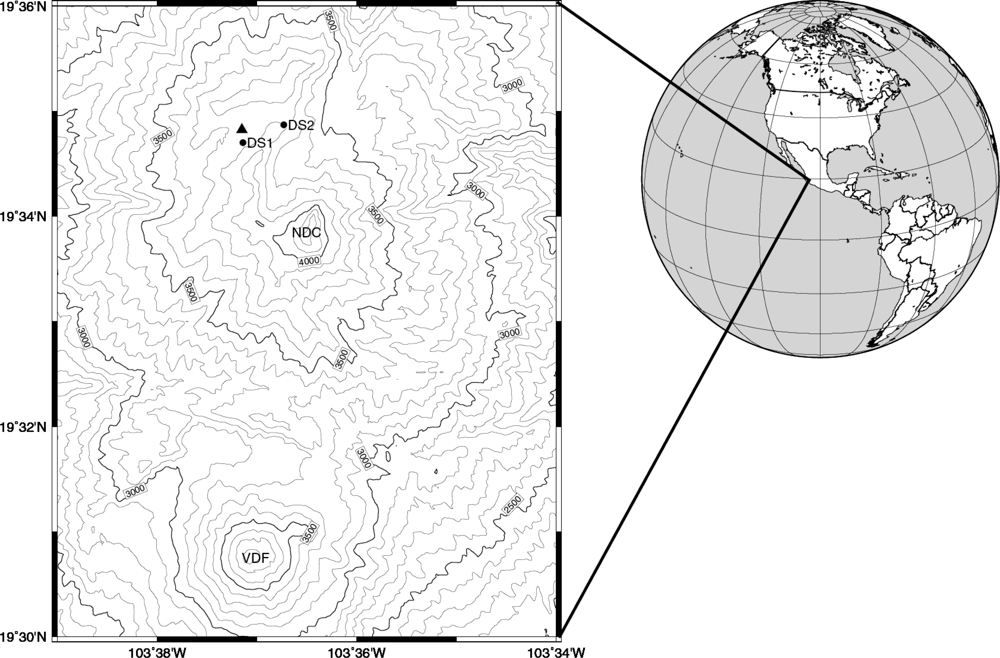
2. Experimental Section
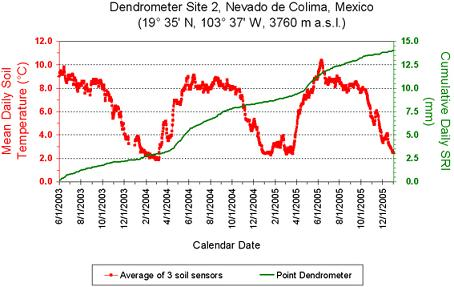
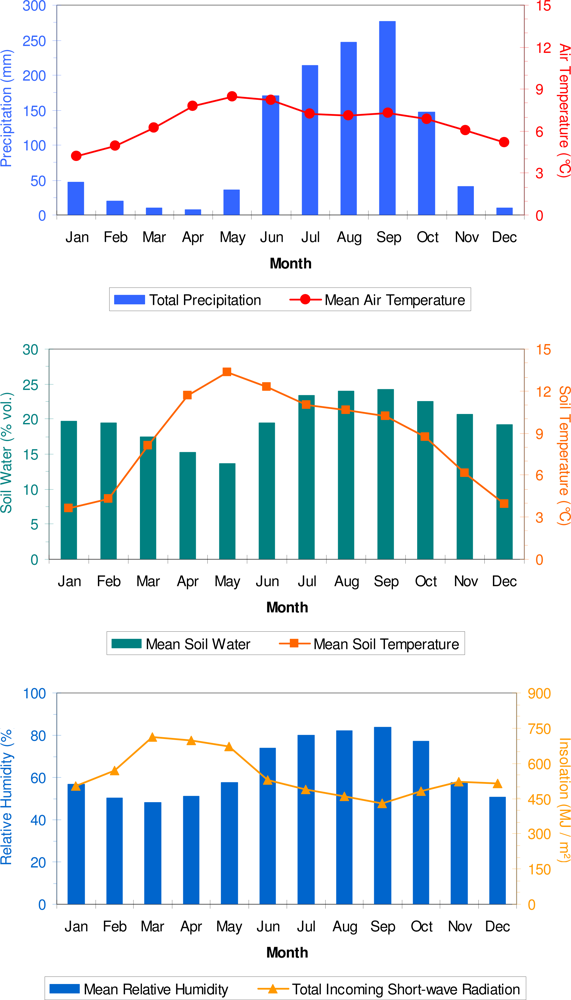
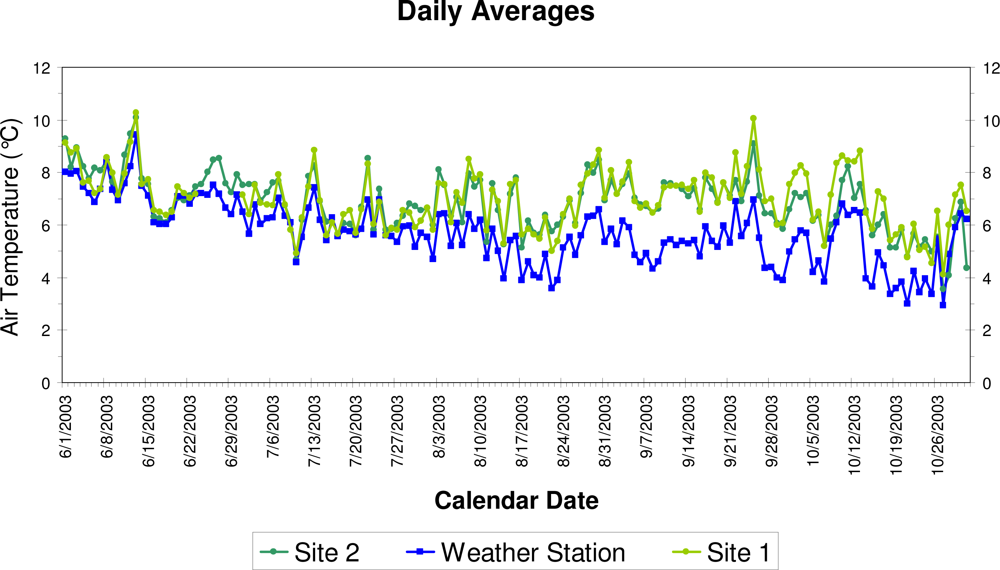
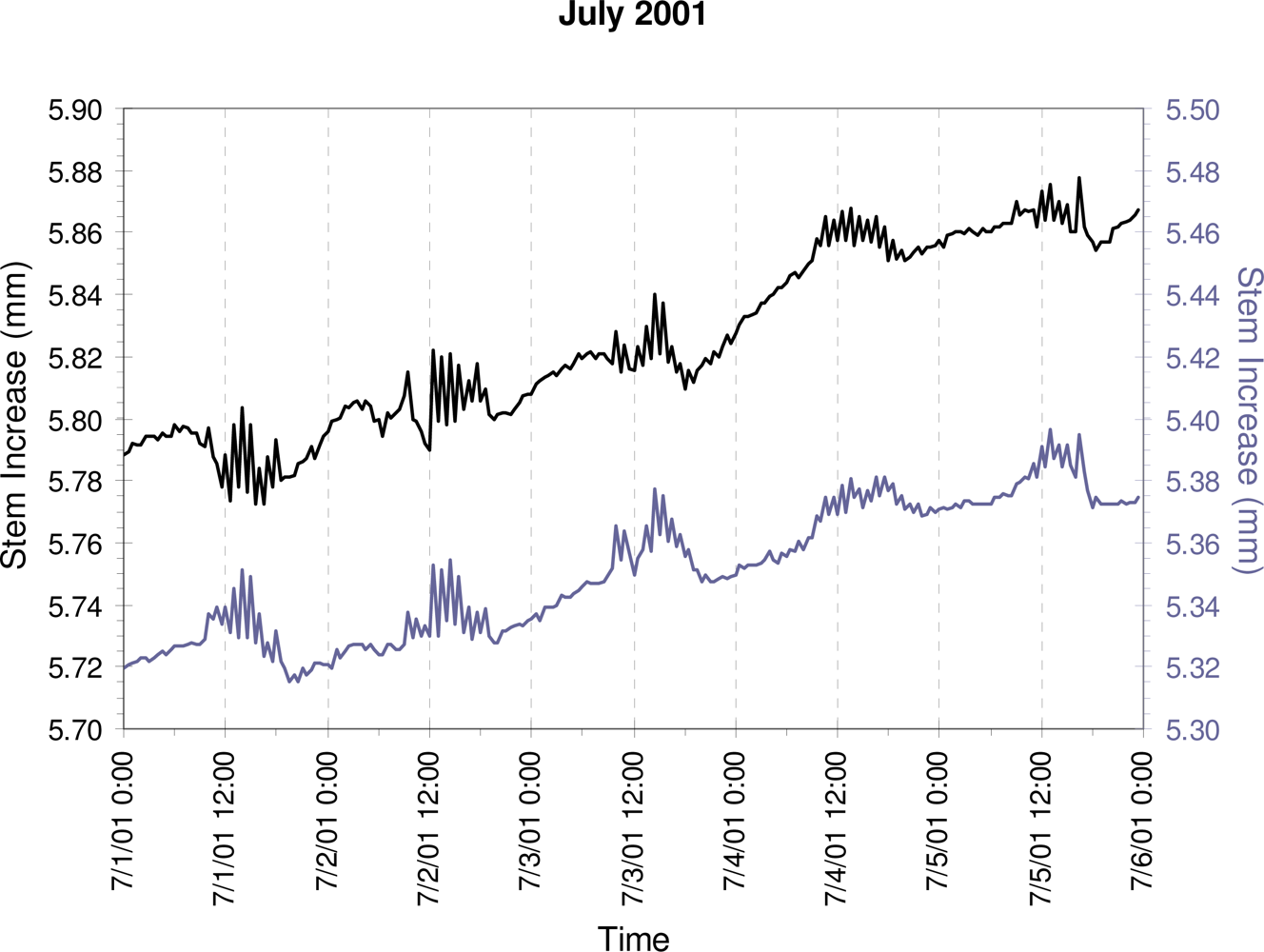
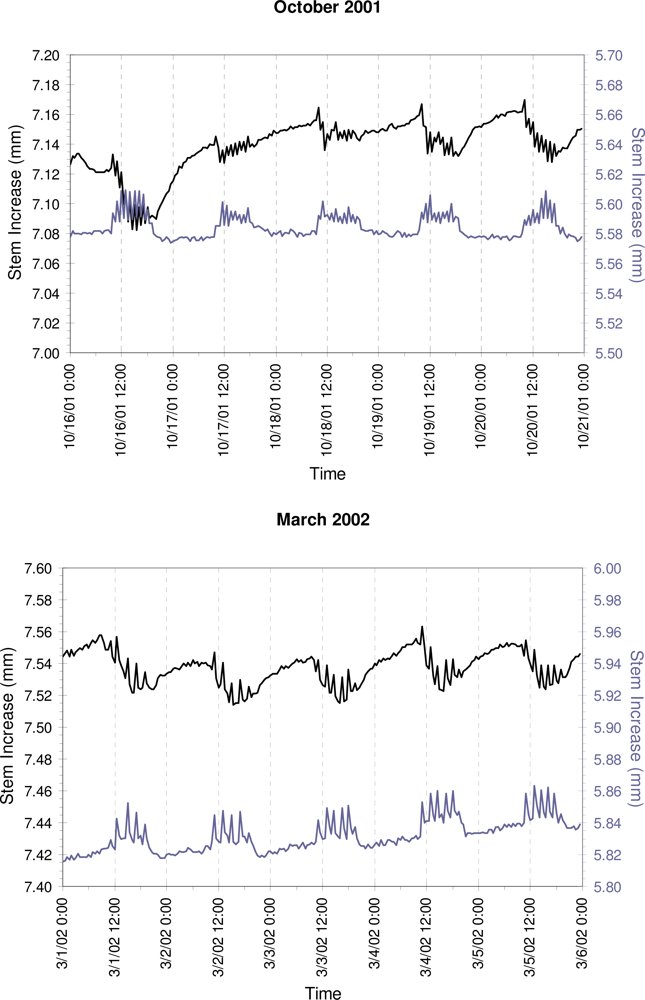
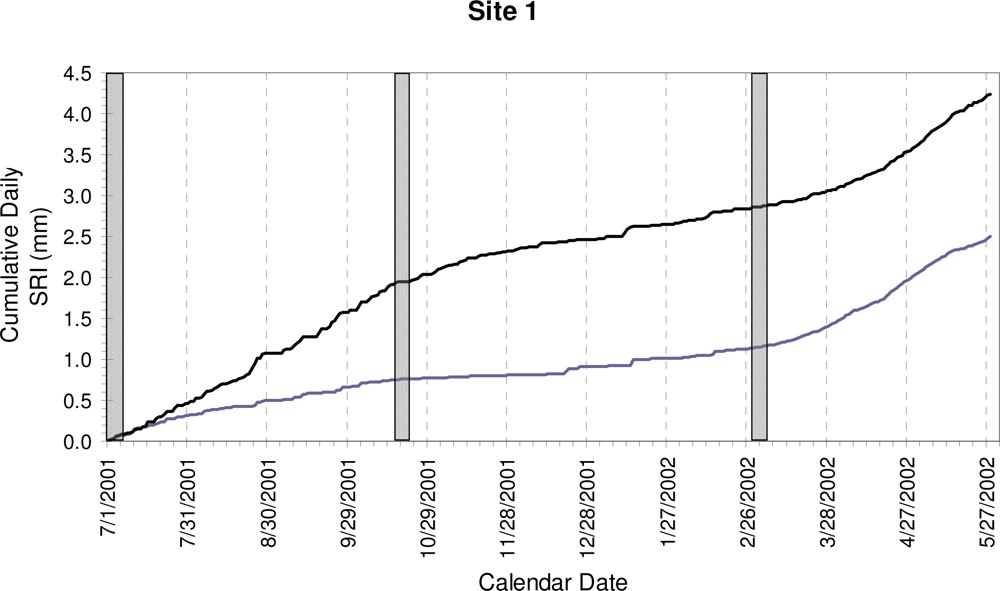
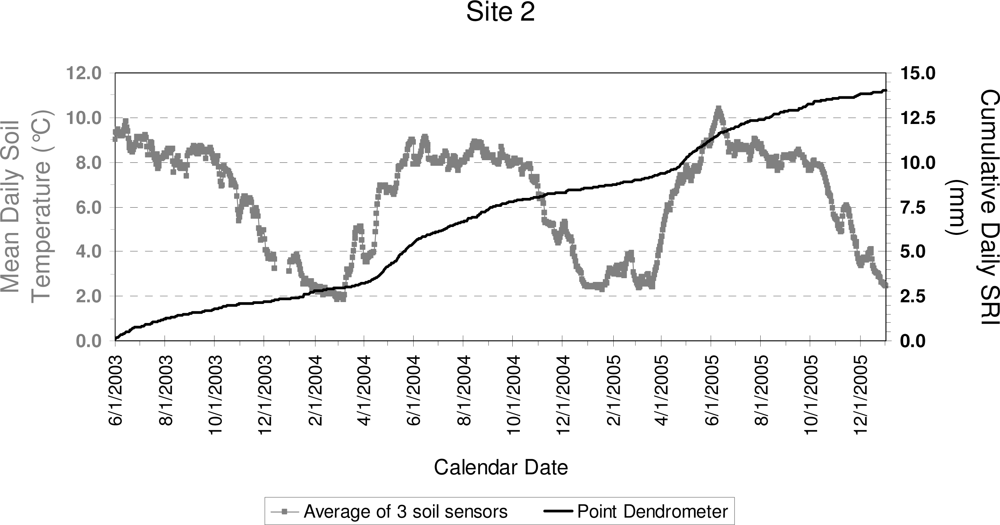
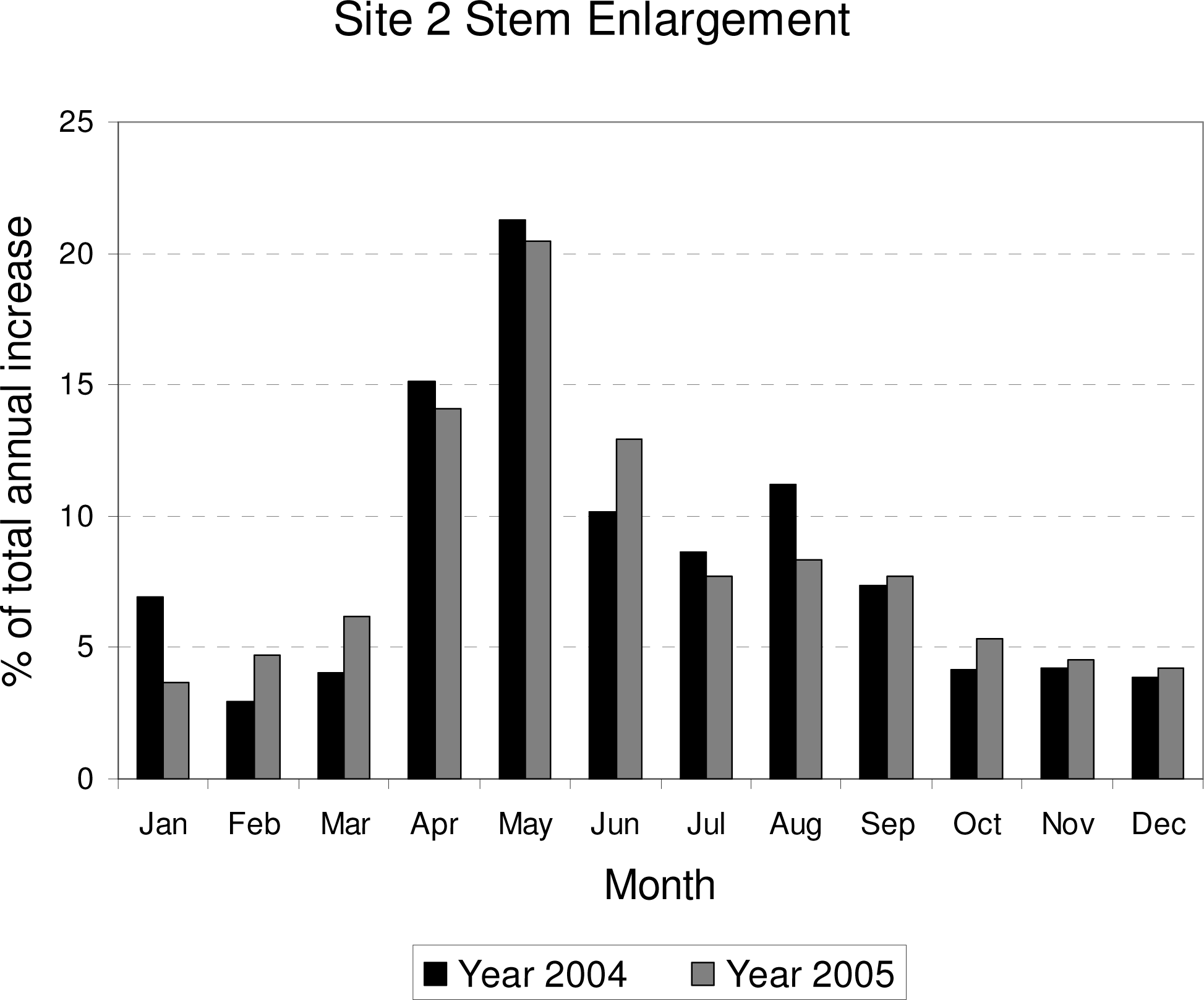
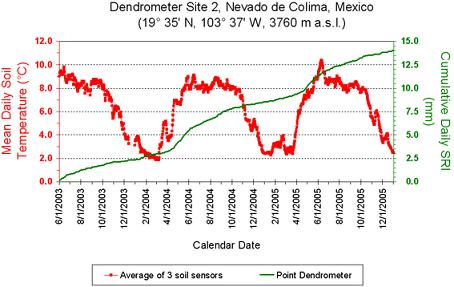
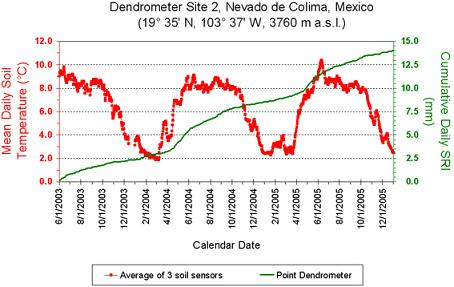
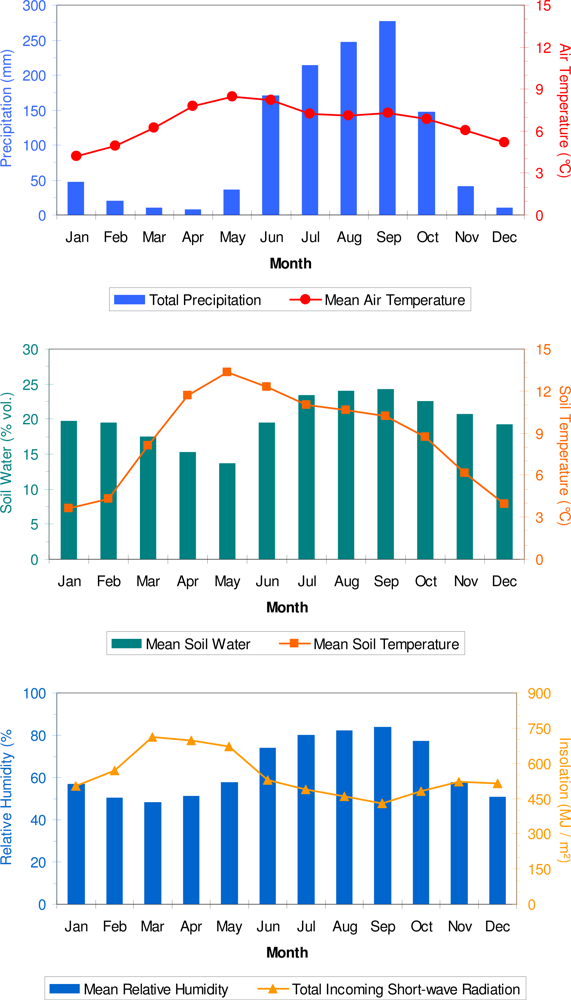
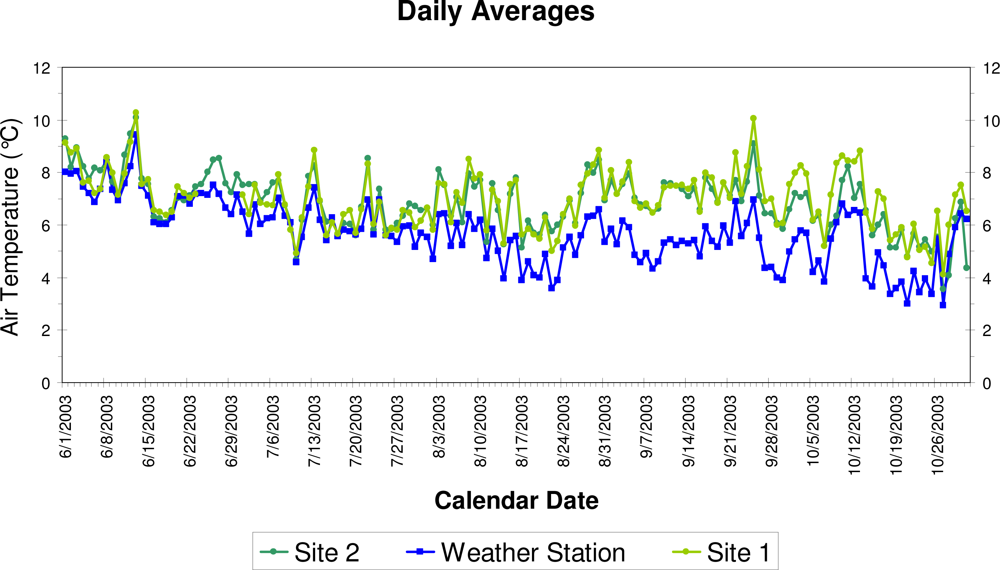
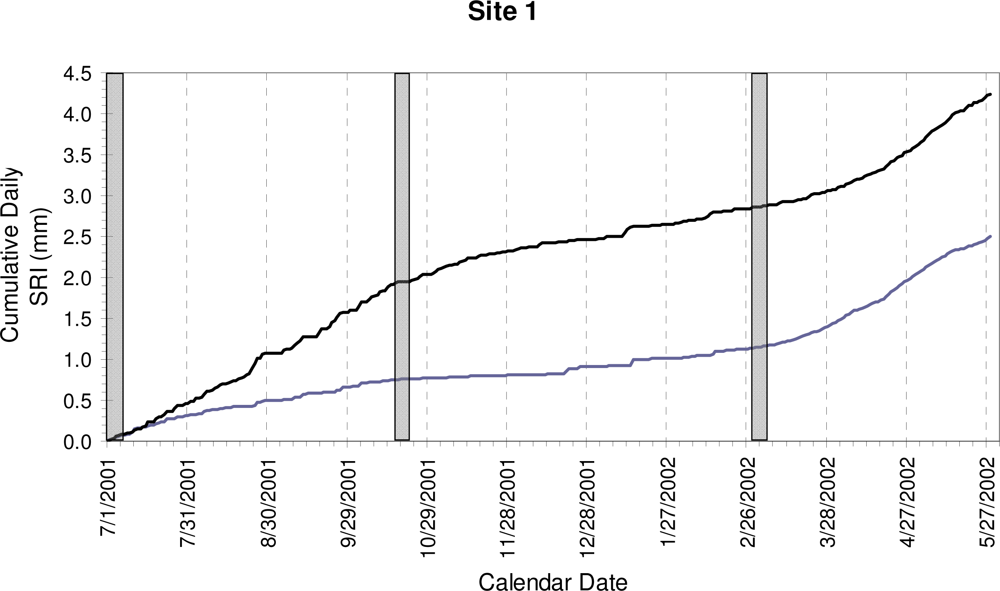
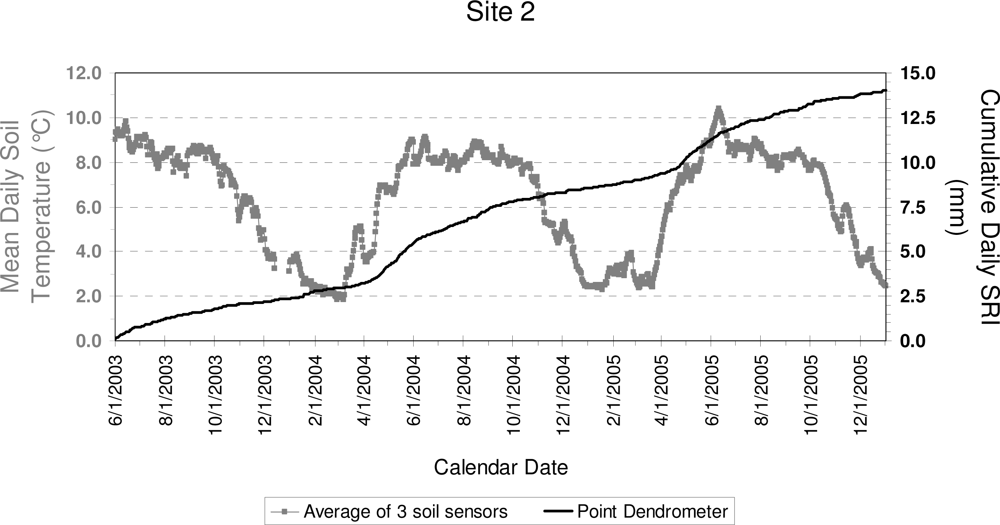
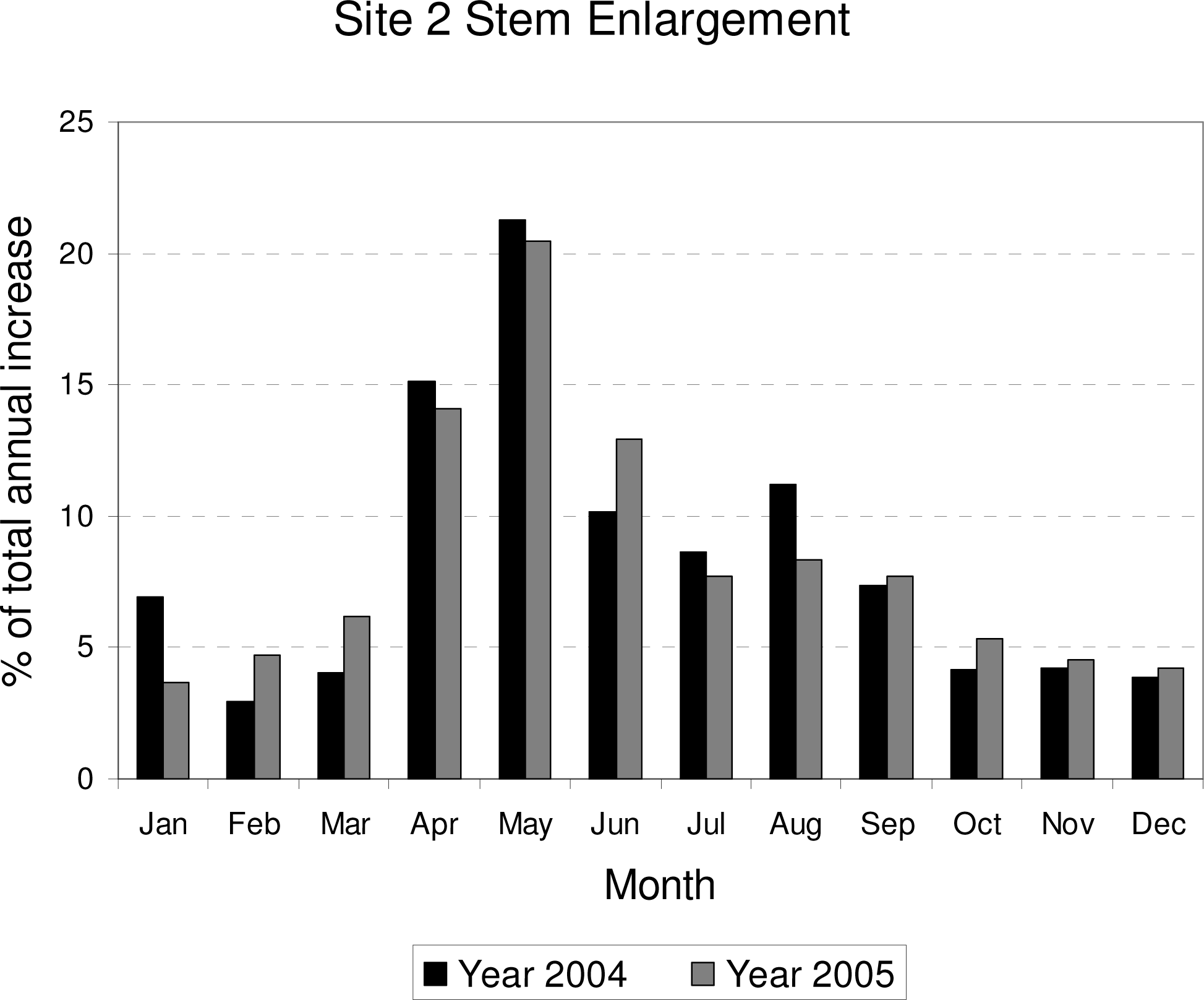
3. Results
4. Discussion
Acknowledgments
References
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Bradley, R.S. Paleoclimatology, 2nd ed; Academic Press: San Diego, CA, USA, 1999. [Google Scholar]
- Anderson, S.P.; Bales, R.C.; Duffy, C.J. Critical Zone Observatories: Building a Network to Advance Interdisciplinary Study Of Earth Surface Processes. Miner. Mag 2008, 72, 7–10. [Google Scholar]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer-Verlag: Berlin, Germany, 1999. [Google Scholar]
- National Research Council. Surface Temperature Reconstructions for the Last 2,000 Years. In Committee on Surface Temperature Reconstructions for the Last 2,000 Years; The National Academies Press: Washington, DC, USA, 2006; p. 196. [Google Scholar]
- Wieser, G.; Matyssek, R.; Luzian, R.; Zwerger, P.; Pindur, P.; Oberhuber, W.; Gruber, A. Effects of atmospheric and climate change at the timberline of the Central European Alps. Ann. Sci 2009, 66, 1–12. [Google Scholar]
- Salzer, M.W.; Hughes, M.K.; Bunn, A.G.; Kipfmueller, K.F. Recent unprecedented tree-ring growth in bristlecone pine at the highest elevations and possible causes. Proc. Nat. Acad. Sci. USA 2009, 106, 20348–20353. [Google Scholar]
- Daly, C.; Neilson, R.P.; Phillips, D.L. A statistical-topographic model for mapping climatological precipitation over mountainous terrain. J. Appl. Meteorol 1994, 33, 140–158. [Google Scholar]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar]
- Wilmking, M.; Juday, G.P. Longitudinal variation of radial growth at Alaska’s northern treeline—recent changes and possible scenarios for the 21st century. Global Planet. Change 2005, 47, 282–300. [Google Scholar]
- Biondi, F. A 400-year tree-ring chronology from the tropical treeline of North America. Ambio 2001, 30, 162–166. [Google Scholar]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr 2004, 31, 713–732. [Google Scholar]
- Phillips, O.L.; Malhi, Y.; Higuchi, N.; Laurance, W.F.; Nunez, P.V.; Vasquez, R.M.; Laurance, S.G.; Ferreira, L.V.; Stern, M.; Brown, S.; Grace, J. Changes in the carbon balance of tropical forests: Evidence from long-term plots. Science 1998, 282, 439–442. [Google Scholar]
- Worbes, M. How to measure growth dynamics in tropical trees: a review. IAWA J 1995, 16, 337–351. [Google Scholar]
- Baker, T.R.; Affum-Baffoe, K.; Burslem, D.F.R.P.; Swain, M.D. Phenological differences in tree water use and the timings of tropical forest inventories: conclusions from patterns of dry season diameter change. Forest Ecol. Manage 2002, 171, 261–274. [Google Scholar]
- Pelissier, R.; Pascal, J. Two-year tree growth patterns investigated from monthly girth records using dendrometer bands in a wet evergreen forest in India. J. Trop. Ecol 2000, 16, 429–446. [Google Scholar]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J.; Holland, E.A. Net primary production in tropical forests: An evaluation and synthesis of existing field data. Ecol. Appl 2001, 11, 371–384. [Google Scholar]
- Phillips, O.L.; Malhi, Y.; Vinceti, B.; Baker, T.; Lewis, S.L.; Higuchi, N.; Laurance, W.F.; Nunez, P.V.; Vasquez, R.M.; Laurance, S.G.; Ferreira, L.V.; Stern, M.; Brown, S.; Grace, J. Changes in growth of tropical forests: Evaluating potential biases. Ecol. Appl 2002, 12, 576–587. [Google Scholar]
- Sheil, D. Growth assessment in tropical tree: large daily diameter fluctuations and their concealment by dendrometer bands. Can. J. For. Res 2003, 33, 2027–2035. [Google Scholar]
- Breitsprecher, A.; Bethel, J.S. Stem-growth periodicity of trees in a tropical wet forest of Costa Rica. Ecology 1990, 7, 1156–1164. [Google Scholar]
- Worbes, M. Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela. J. Ecol 1999, 87, 391–403. [Google Scholar]
- Herzog, K.M.; Hasler, R.; Thum, R. Diurnal changes in the radius of a subalpine Norway spruce stem: their relation to the sap flow and their use to estimate transpiration. Trees 1995, 10, 94–101. [Google Scholar]
- Zweifel, R.; Item, H.; Hasler, R. Link between diurnal stem radius changes and tree water relations. Tree Physiol 2001, 21, 869–877. [Google Scholar]
- Perämäki, M.; Vesala, T.; Nikinmaa, E. Modeling the dynamics of pressure propagation and diameter variation in tree sapwood. Tree Physiol 2005, 25, 1091–1099. [Google Scholar]
- Irvine, J.; Grace, J. Continuous measurement of water tensions in the xylem of trees based on the elastic properties of wood. Planta 1997, 202, 455–461. [Google Scholar]
- Lovdahl, L.; Odin, H. Diurnal changes in the stem diameter of Norway spruce in relation to relative humidity and air temperature. Trees 1992, 6, 245–251. [Google Scholar]
- Deslauriers, A.; Morin, H.; Urbinati, C.; Carrer, M. Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees 2003, 17, 477–484. [Google Scholar]
- Downes, G.M.; Beadle, C.; Gensler, W.; Mummery, D.; Worledge, D. Diurnal variation and radial growth of stems in young plantation eucalyptus. In Tree-Ring Analysis: Biological, Methodological and Environmental Aspects; Wimmer, R., Vetter, R.E., Eds.; CABI Publishing: New York, NY, USA, 1999; pp. 83–103. [Google Scholar]
- McLaughlin, S.B.; Wullschlereger, S.D.; Nosal, M. Diurnal and seasonal changes in stem increment and water use by yellow poplar trees in response to environmental stress. Tree Physiol 2003, 23, 1125–1136. [Google Scholar]
- Bouriaud, O.; Leban, J.M.; Bert, D.; Deleuze, C. Intra-annual variations in climate influence growth and wood density of Norway spruce. Tree Physiol 2005, 25, 651–660. [Google Scholar]
- Adams, D.K.; Comrie, A.C. The North American Monsoon. Bull. Amer. Meteorol. Soc 1997, 78, 2197–2213. [Google Scholar]
- Dominguez, F.; Villegas, J.C.; Breshears, D.D. Spatial extent of the North American Monsoon: Increased crossregional linkages via atmospheric pathways. Geophys. Res. Let 2009, 36, L07401. [Google Scholar]
- Perry, J.P., Jr. The Pines of Mexico and Central America; Timber Press: Portland, OR, USA, 1991. [Google Scholar]
- Richardson, D.M. Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Gómez-Tuena, A.; Orozco-Esquivel, M.T.; Ferrari, L. Igneous Petrogenesis of the Trans-Mexican Volcanic Belt. In Geology of México; Alaniz-Álvarez, S.A., Nieto-Samaniego, A.F., Eds.; Geological Society of America: Boulder, CO, USA, 2007; Volume 422, pp. 129–182. [Google Scholar]
- Barlow, M.; Nigam, S.; Berbery, E.H. Evolution of the North American monsoon system. J. Climate 1998, 11, 2238–2257. [Google Scholar]
- Galindo Estrada, I.; Elizalde Torres, A.; Solano Barajas, R.; Cruz Calvario, M. Climatología del Volcán de Fuego de Colima; Universidad de Colima: Colima, Mexico, 1998; p. 73. [Google Scholar]
- Biondi, F.; Hartsough, P.C.; Galindo Estrada, I. Recent warming at the tropical treeline of North America. Front. Ecol. Environ 2009, 7, 463–464. [Google Scholar]
- Murray, F.W. On the computation of saturation vapor pressure. J. Appl. Meteorol 1967, 6, 203–204. [Google Scholar]
- Brutsaert, W. Evaporation into the Atmosphere; D. Reidel Publishing Company: Boston, MA, USA, 1982. [Google Scholar]
- Biondi, F.; Hartsough, P.C.; Galindo Estrada, I. Daily weather and tree growth at the tropical treeline of North America. Arctic Antarct. Alp. Res 2005, 37, 16–24. [Google Scholar]
- Dawdy, D.R.; Matalas, N.C. Statistical and probability analysis of hydrological data. Part III: Analysis of variance, covariance, and time series. In Handbook of Applied Hydrology; Chow, V.T., Ed.; McGraw-Hill: New York, NY, USA, 1969; pp. 68–90. [Google Scholar]
- Morrison, D.F. Applied Linear Statistical Methods; Prentice-Hall: Englewood Cliffs, NJ, USA, 1983; p. 562. [Google Scholar]
- Efron, B.; Tibshirani, R.J. Bootstrap methods for standard errors, confidence intervals, and other measures of statistical accuracy. Stat. Sci 1986, 1, 54–75. [Google Scholar]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Chapman & Hall/CRC: Boca Raton, FL, USA, 1993. [Google Scholar]
- Morzuch, B.J.; Ruark, G.A. Principal components regression to mitigate the effects of multicollinearity. Forest Sci 1991, 37, 191–199. [Google Scholar]
- Wilks, D.S. Statistical Methods in the Atmospheric Sciences; Academic Press: San Diego, CA, USA, 1995; Volume 59, p. 467. [Google Scholar]
- Guiot, J. Methods of calibration. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer: Dordrecht, The Netherlands, 1990; pp. 165–178. [Google Scholar]
- Dixon, P.M. Bootstrap resampling. In The Encyclopedia of Environmetrics; El-Shaarawi, A.H., Piegorsch, W.W., Eds.; Wiley: New York, NY, USA, 2001; Vol. Statistical and Numerical Computing,; p. 9. [Google Scholar]
- Gaylord, M.L.; Kolb, T.E.; Wallin, K.F.; Wagner, M.R. Seasonality and lure preference of bark beetles (Curculionidae: Scolytinae) and associates in a northern Arizona ponderosa pine forest. Environ. Entomol 2006, 35, 37–47. [Google Scholar]
- Negrón, J.F. Estimating probabilities of infestation and extent of damage by the roundheaded pine beetle in ponderosa pine in the Sacramento Mountains, New Mexico. Can. J. For. Res 1997, 27, 1936–1945. [Google Scholar]
- Negrón, J.F.; Wilson, J.L.; Anhold, J.A. Stand conditions associated with roundheaded pine beetle (Coleoptera: Scolytidae) infestations in Arizona and Utah. Environ. Entomol 2000, 29, 20–27. [Google Scholar]
- Biondi, F. Recent increase in maximum temperature at the tropical treeline of North America. EOS Trans. Amer. Geophys. Union 2009, 90, B33A-0361. [Google Scholar]
- Schutte, K.H.; Burger, C.P. Sensitive dendrometers for contemporary research: A critical evaluation of strain gauge dendrometers. J. S. Afr. Bot 1981, 47, 273–291. [Google Scholar]
- Biondi, F.; Galindo Estrada, I.; Elizalde Torres, A.; Gavilanes Ruiz, J.C. Tree growth response to the 1913 eruption of Volcán de Fuego de Colima, Mexico. Quaternary Res 2003, 59, 293–299. [Google Scholar]
- Downes, G.M.; Drew, D.; Battaglia, M.; Schulze, D. Measuring and modelling stem growth and wood formation: An overview. Dendrochronologia 2009, 27, 147–157. [Google Scholar]
- Deslauriers, A.; Rossi, S.; Anfodillo, T. Dendrometer and intra-annual tree growth: What kind of information can be inferred? Dendrochronologia 2007, 25, 113–124. [Google Scholar]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol 2006, 170, 301–310. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | SolRad | Prec | TSmax | TSmin | TAmax | TAmin | RH | SWV | AirPr | WG | WS | VPD | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | |
| Jul 2001 | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | 0.54 | / | ns | / | −0.42 | / | ns | / | ns | / | −0.50 | / |
| Aug 2001 | −0.41 | / | ns | / | ns | / | ns | / | −0.44 | / | 0.41 | / | 0.37 | / | ns | / | ns | / | 0.48 | / | ns | / | −0.36 | / |
| Sep 2001 | −0.48 | / | 0.45 | / | ns | / | ns | / | −0.59 | / | 0.47 | / | 0.51 | / | ns | / | −0.51 | / | 0.55 | / | 0.43 | / | −0.51 | / |
| Apr 2002 | ns | 0.39 | ns | Ns | 0.46 | 0.57 | 0.54 | 0.55 | 0.35 | 0.58 | 0.44 | 0.43 | ns | ns | −0.45 | −0.54 | / | / | ns | ns | ns | ns | / | / |
| May 2002 | ns | 0.64 | ns | −0.40 | 0.46 | 0.55 | 0.60 | 0.35 | 0.31 | 0.58 | 0.46 | 0.63 | ns | −0.52 | −0.29 | ns | / | / | ns | ns | ns | 0.45 | / | / |
| Jun 2002 | −0.54 | / | 0.54 | / | ns | / | ns | / | −0.47 | / | ns | / | 0.74 | / | ns | / | ns | / | ns | / | ns | / | −0.72 | / |
| Jul 2002 | −0.56 | / | 0.41 | / | ns | / | 0.49 | / | ns | / | 0.35 | / | ns | / | ns | / | ns | / | −0.31 | / | −0.37 | / | ns | / |
| Aug 2002 | −0.49 | / | ns | / | ns | / | 0.45 | / | −0.45 | / | 0.41 | / | ns | / | ns | / | −0.53 | / | 0.46 | / | 0.47 | / | ns | / |
| Sep 2002 | −0.47 | / | 0.48 | / | ns | / | ns | / | −0.61 | / | 0.38 | / | ns | / | ns | / | −0.46 | / | 0.57 | / | 0.52 | / | ns | / |
| Jun 2003 | −0.56 | −0.43 | 0.41 | Ns | ns | ns | ns | ns | ns | ns | 0.36 | ns | 0.43 | ns | / | / | ns | −0.38 | ns | ns | ns | ns | −0.42 | ns |
| Jul 2003 | −0.59 | −0.46 | 0.63 | 0.32 | ns | ns | ns | ns | −0.47 | ns | ns | ns | ns | ns | / | / | ns | −0.31 | ns | ns | ns | ns | ns | ns |
| Aug 2003 | −0.49 | −0.47 | ns | Ns | ns | ns | 0.38 | ns | −0.47 | −0.44 | 0.51 | 0.38 | ns | ns | / | / | ns | ns | 0.45 | 0.35 | 0.54 | 0.40 | ns | ns |
| Sep 2003 | −0.45 | −0.50 | ns | Ns | ns | −0.34 | ns | ns | −0.60 | −0.72 | 0.41 | 0.31 | 0.61 | 0.59 | / | / | ns | −0.36 | ns | ns | 0.37 | 0.46 | −0.57 | −0.56 |
| Apr 2004 | / | 0.60 | / | −0.29 | / | 0.72 | / | 0.70 | / | 0.54 | / | 0.67 | / | −0.53 | / | −0.68 | / | ns | / | ns | / | ns | / | 0.69 |
| May 2004 | / | 0.33 | / | Ns | / | 0.56 | / | 0.43 | / | 0.61 | / | 0.46 | / | −0.42 | / | ns | / | ns | / | ns | / | ns | / | 0.52 |
| Jun 2004 | / | −0.57 | / | 0.57 | / | ns | / | ns | / | −0.53 | / | 0.53 | / | 0.47 | / | ns | / | ns | / | 0.50 | / | 0.49 | / | −0.45 |
| Jul 2004 | / | Ns | / | 0.43 | / | ns | / | ns | / | −0.40 | / | ns | / | 0.33 | / | ns | / | ns | / | ns | / | ns | / | −0.32 |
| Aug 2004 | / | Ns | / | Ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns |
| Sep 2004 | / | Ns | / | Ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns |
| Apr 2005 | / | Ns | / | Ns | / | 0.58 | / | 0.52 | / | 0.43 | / | 0.35 | / | −0.38 | / | −0.64 | / | ns | / | ns | / | ns | / | 0.44 |
| May 2005 | / | Ns | / | Ns | / | ns | / | ns | / | ns | / | ns | / | −0.39 | / | ns | / | ns | / | ns | / | ns | / | 0.41 |
| Jun 2005 | / | Ns | / | Ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns | / | ns |
| Jul 2005 | / | −0.43 | / | 0.44 | / | ns | / | ns | / | −0.54 | / | ns | / | 0.54 | / | 0.51 | / | ns | / | ns | / | 0.41 | / | −0.53 |
| Aug 2005 | / | −0.49 | / | Ns | / | ns | / | ns | / | −0.58 | / | ns | / | 0.56 | / | 0.41 | / | ns | / | ns | / | 0.37 | / | −0.63 |
| Sep 2005 | / | −0.34 | / | 0.57 | / | ns | / | ns | / | −0.41 | / | ns | / | 0.45 | / | ns | / | ns | / | 0.35 | / | 0.43 | / | −0.43 |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Biondi, F.; Hartsough, P. Using Automated Point Dendrometers to Analyze Tropical Treeline Stem Growth at Nevado de Colima, Mexico. Sensors 2010, 10, 5827-5844. https://doi.org/10.3390/s100605827
Biondi F, Hartsough P. Using Automated Point Dendrometers to Analyze Tropical Treeline Stem Growth at Nevado de Colima, Mexico. Sensors. 2010; 10(6):5827-5844. https://doi.org/10.3390/s100605827
Chicago/Turabian StyleBiondi, Franco, and Peter Hartsough. 2010. "Using Automated Point Dendrometers to Analyze Tropical Treeline Stem Growth at Nevado de Colima, Mexico" Sensors 10, no. 6: 5827-5844. https://doi.org/10.3390/s100605827
APA StyleBiondi, F., & Hartsough, P. (2010). Using Automated Point Dendrometers to Analyze Tropical Treeline Stem Growth at Nevado de Colima, Mexico. Sensors, 10(6), 5827-5844. https://doi.org/10.3390/s100605827