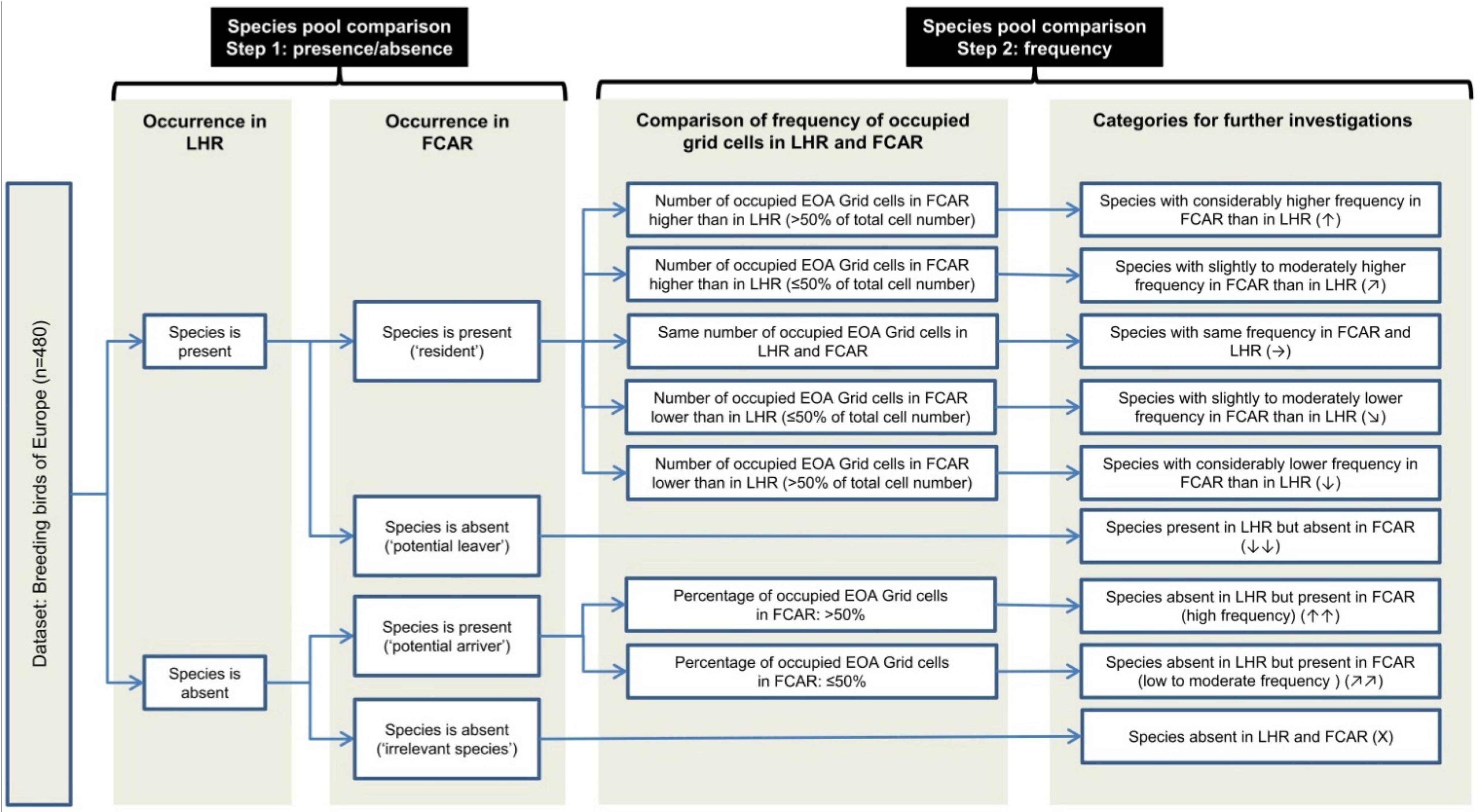
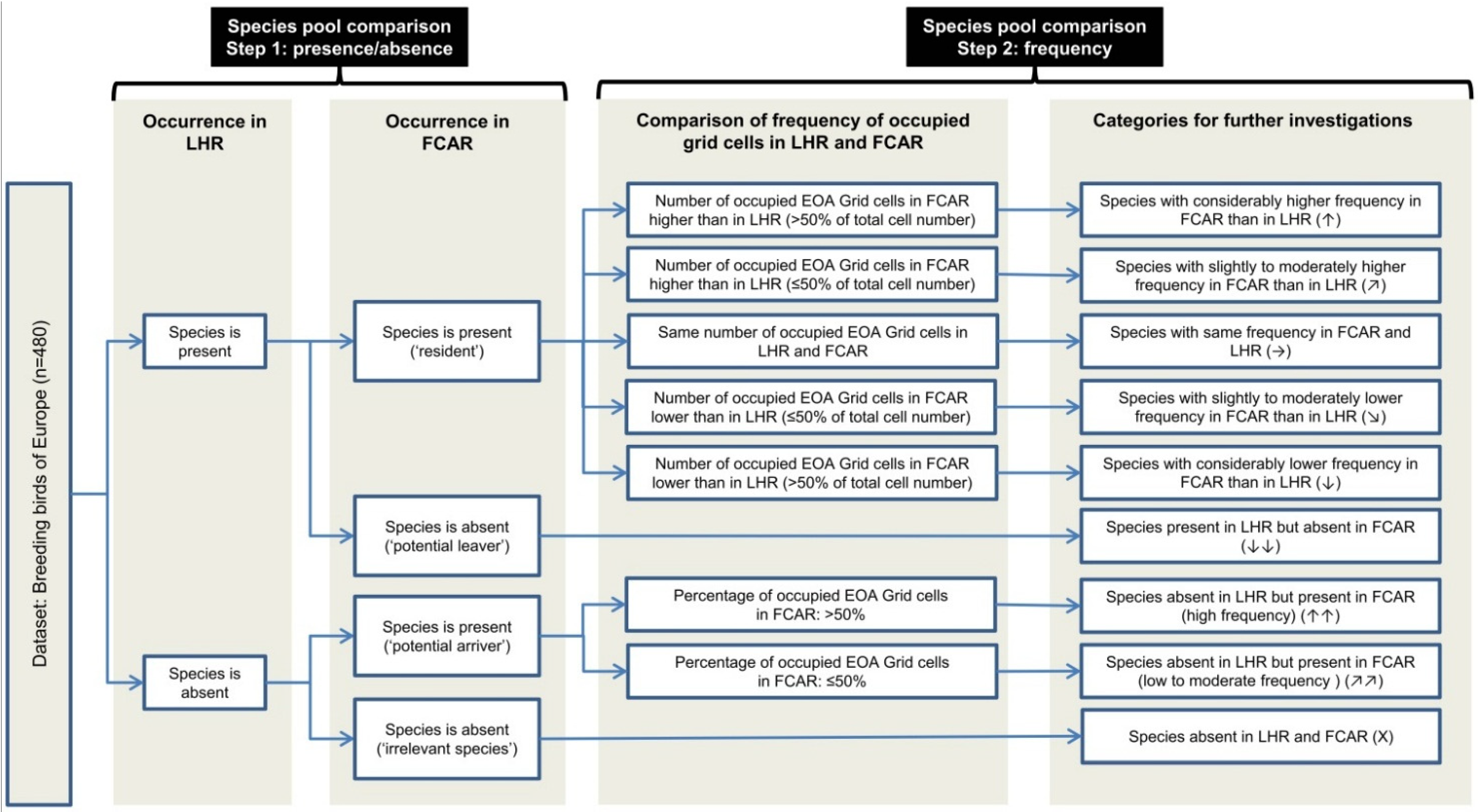
4.1. Species Pool Comparisons of Future Climatically Analogous Regions – A Useful Approach to Assess Climate Change Impacts on Regional Bird Communities
We developed an approach to examine possible turnovers in regional bird communities induced by climate change and applied this approach to assess to what extent climate change might lead to a turnover in the bird community of the Lüneburg Heath region. In addition, we sought to determine which species groups and corresponding habitat types might be most affected in this region. By using species pool comparisons of future climatically analogous regions, we developed an alternative to the widely used bioclimatic envelope models (e.g., [
18,
46,
47]) to assess climate change impacts within a regional context.
The FCAR approach has several advantages. One advantage is that it does not depend on the availability of distribution data for the whole range of the species. In bioclimatic envelope modeling the suitable climate space for species can be underestimated if the complete distribution range of a species cannot be taken into account due to incomplete data or if the range limits of a species are determined by geographical instead of climatic factors. This can lead to an overestimation of local extinctions [
48]. Further advantages of the FCAR method are that we are able to include more accurate regional climate projections and that we can take into account the landscape context when assessing climate change impacts.
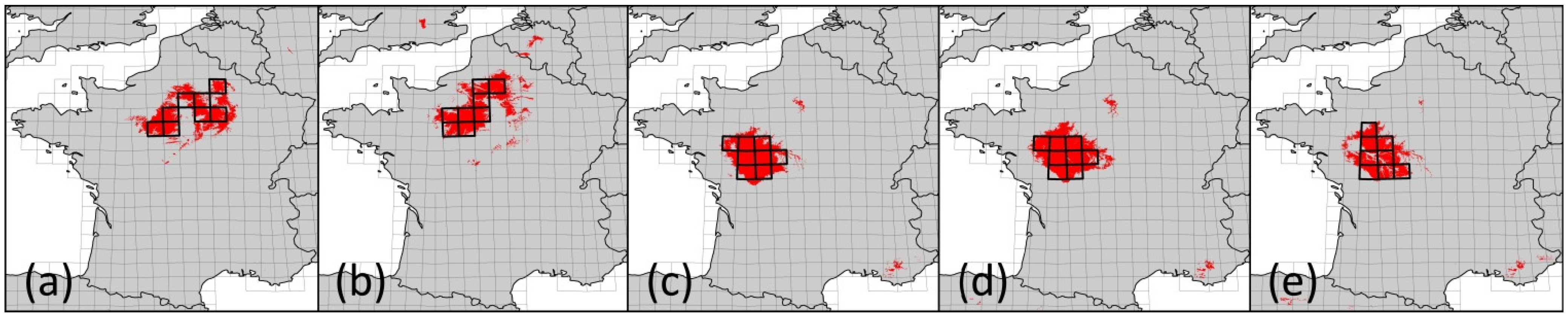
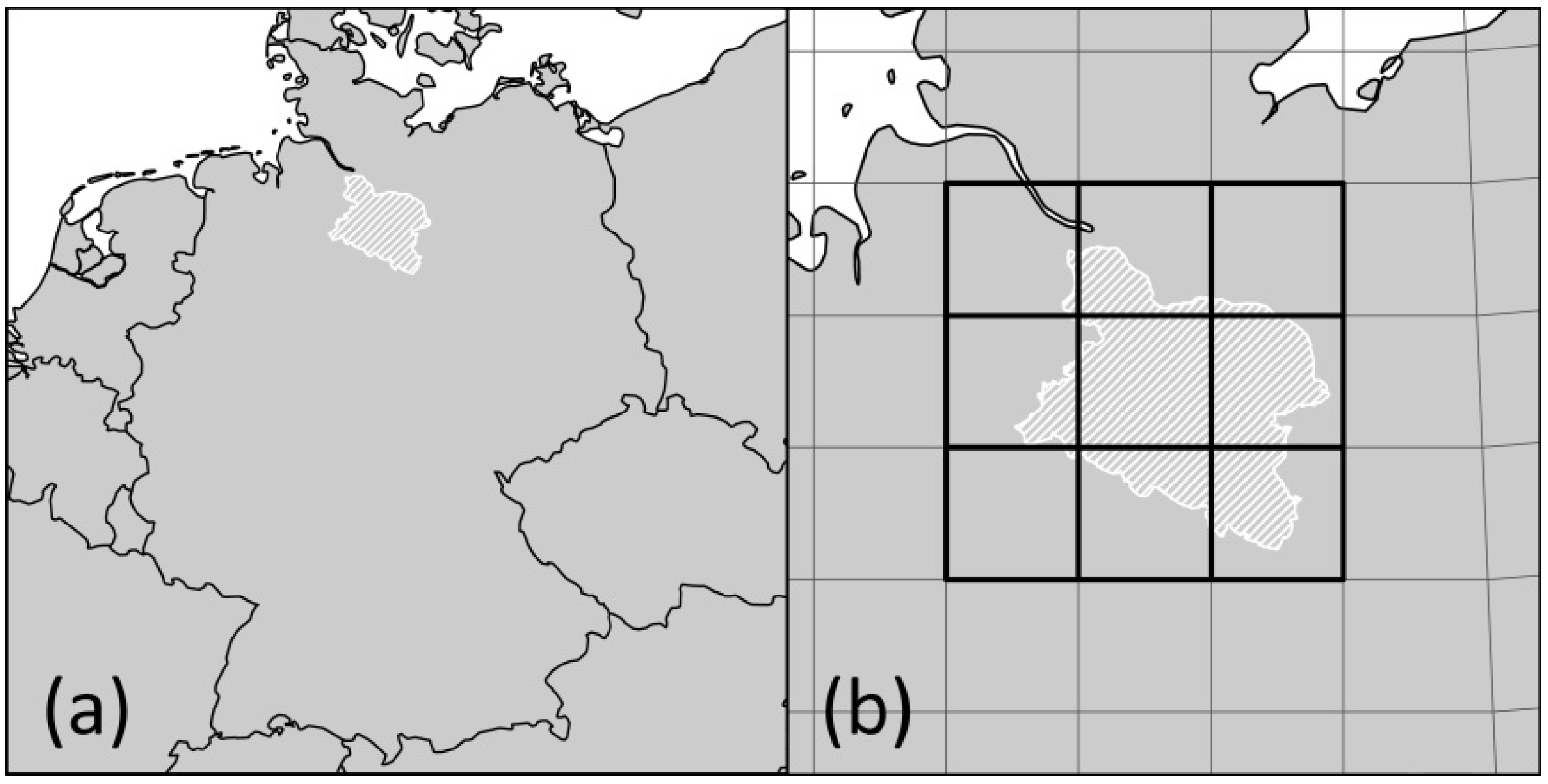
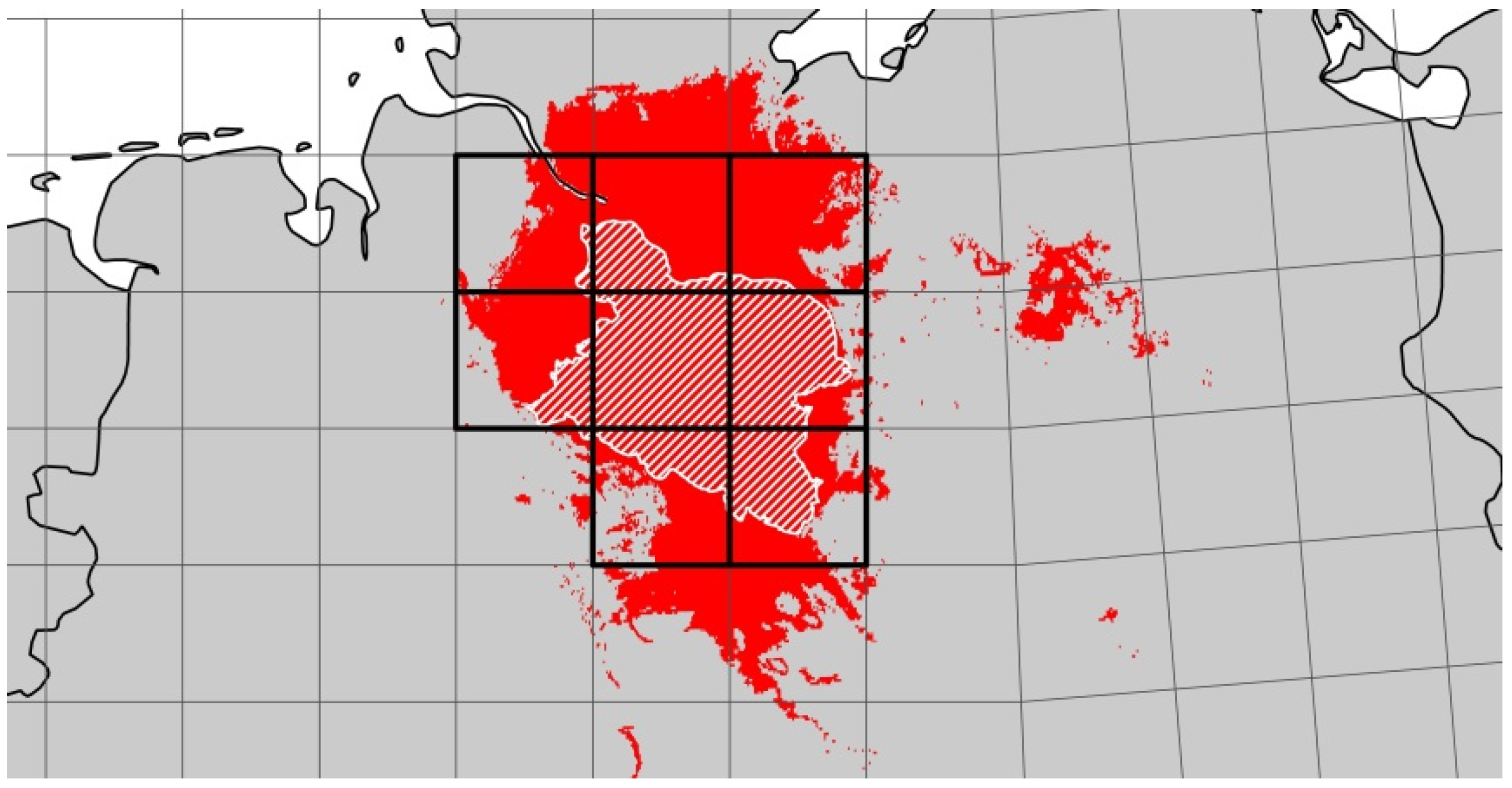
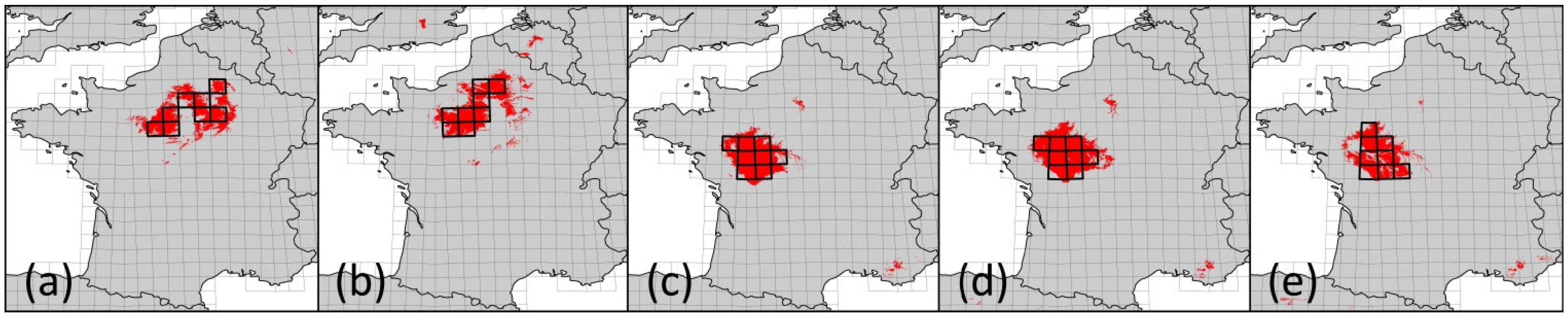
For the LHR we were able to identify four different FCARs representing five different climate projections. However, for two of the altogether seven assessed climate projections, the identified climatically analogous areas were too small to justify species pool comparisons. Thus, when working with an FCAR approach, it is recommendable to use an ensemble of several climate projections because the successful identification of FCARs might not be possible in every case. Further, altitude and land cover have to be sufficiently comparable—as in our study case. However, further research should apply this approach to other regions to test whether it can be transferred to a wide range of regions.
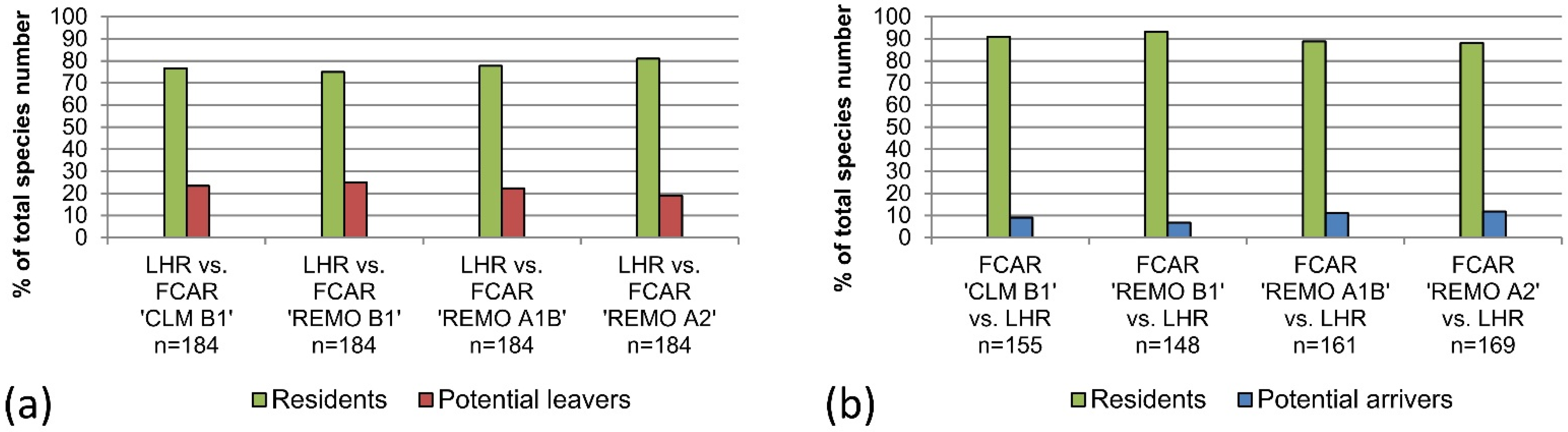
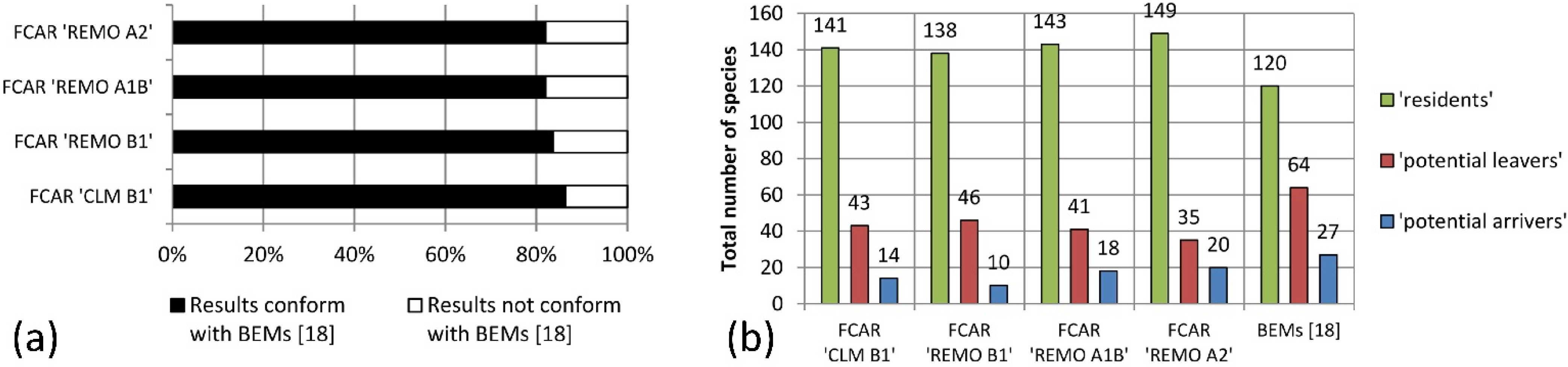
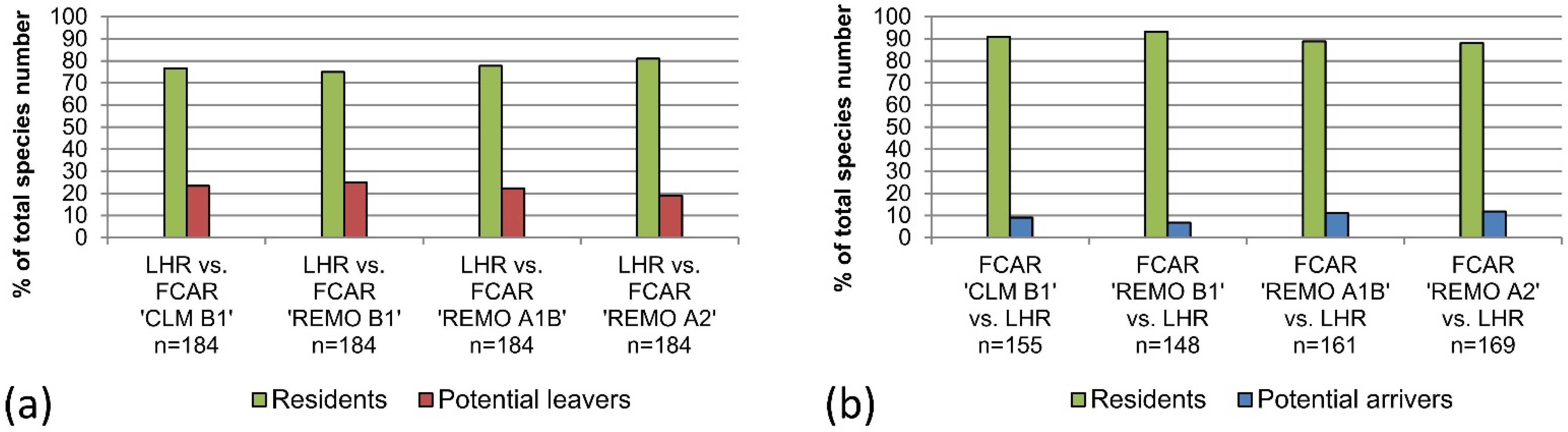
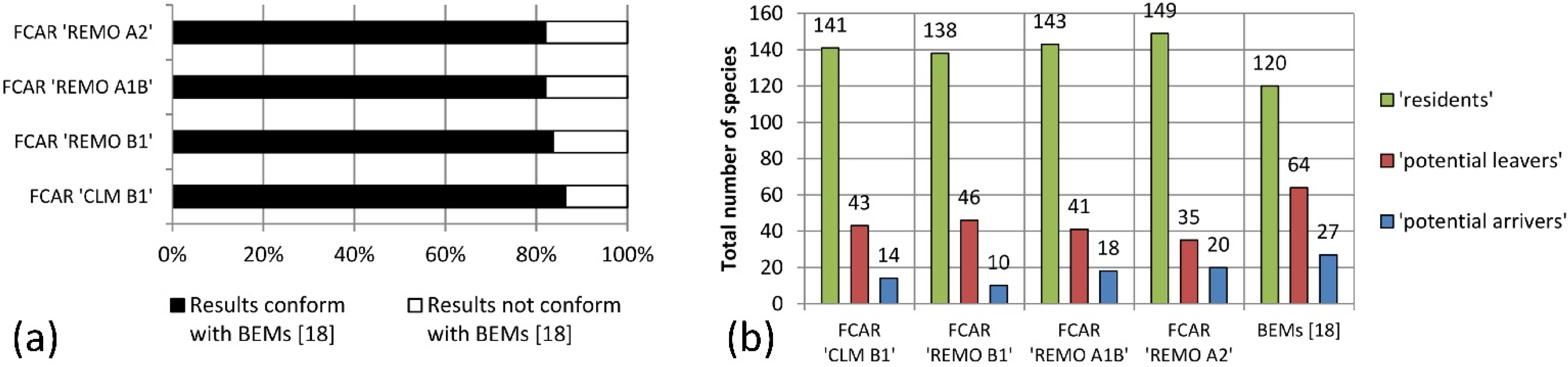
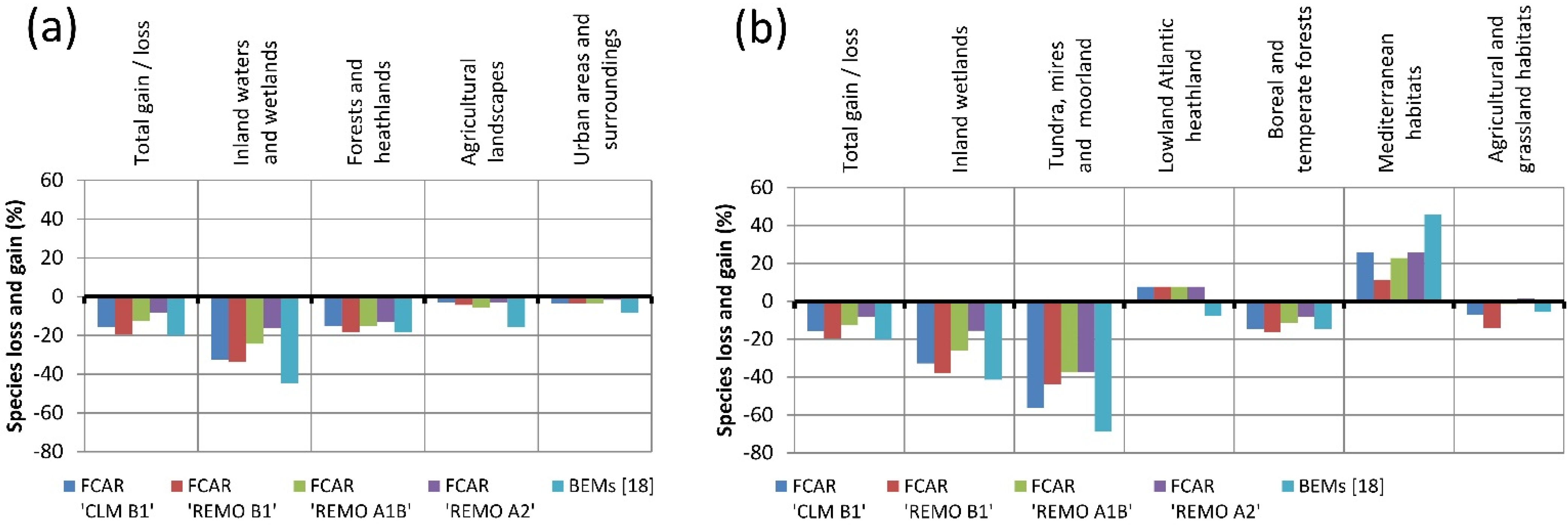
The results of this study indicate that the majority of bird species living in the LHR today would also be able to exist and co-exist in climatic conditions projected for the here considered variables for 2071–2100. However, 19%–25% of today’s LHR-species was categorized as “potential leavers” because they could not be found in the FCARs. The analysis of the data for the LHR from Huntley
et al.’s models [
18] show a higher percentage (35%) of “potential leavers.” Both our FCAR approach and the results of Huntley
et al. indicate a decrease of bird species diversity for the LHR induced by climatic changes during the 21st century, which is in accordance with other studies (e.g., [
49]). In both cases the numbers of “potential leavers” exceeds the number of “potential arrivers.” In general, the degree of conformity of our FCAR approach with the bioclimatic envelope models of Huntley
et al. is high for “potential leavers” as well as for “resident” species that have the same or a higher frequency in the FCARs as in the LHR. Interestingly, the degree of conformity is lower for species with a lower frequency of occupied EOA grid cells in the FCARs than in the LHR. Approximately half of the species that we classify as “residents” are absent from the future species pool of the LHR according to Huntley
et al. Thus, for the LHR our results show milder impacts than the bioclimatic envelope models of Huntley
et al., regarding the potential future loss of species. This is especially the case for wetland species. As we found a detectable effect between habitat availability and occurrence in the FCARs for wetland species, it is likely that many of these species can occur in future climates if sufficient habitat is available. Thus, it might be possible that the models of Huntley
et al. overestimate the impact of climate change on wetland species in the examined region. In fact, Huntley
et al. generally state that the performance of their models was lower for wetland and coastal species. On the other hand, differences between our results and those of Huntley
et al. might also be due to differences in the utilized climate models and variables. Our results, however, are based on a wider corridor of climate change models and scenarios.
With the bioclimatic envelope approach by Huntley
et al. more “potential arrivers” to the LHR can be identified than with the FCAR approach. We assume that our approach potentially underestimates the number of species that might reach the LHR during the course of climatic changes as the selected EOA grid cells for the species pool comparisons only capture part of the actual climatically analogous area. Other spots in Europe were too small to be included in the species pool comparisons, but might nonetheless function as a source for “potential arrivers.” Additionally, there are recent and historic examples for colonizers from other sources and directions, e.g., the Eurasian Penduline Tit (
Remiz pendulinus) and the Fieldfare (
Turdus pilaris) coming westwards from Siberia into Europe (for overview see [
8]). Further, it was not possible to test if some “potential arrivers” were absent in the FCARs due to habitat conditions.
Despite this underestimation, we wonder how likely it is that “potential arrivers” will actually be able to colonize the study area due to favorable climate conditions. Their colonization potential depends on many factors, including dispersal ability and the status of the source populations. While birds are generally highly mobile and some birds can shift their distribution ranges very rapidly within decades—as documented by Hengeveld [
50] for the collared dove (
Streptopelia decaocto)—most birds, in contrast, show some degree of site fidelity [
8]. In spite of the already documented range shifts (e.g., [
4]), there are also studies indicating that birds are currently lagging behind climate warming in their responses [
51]. However, the distances that species identified as “potential arrivers” would need to cross to reach the LHR differ greatly. Among the “potential arrivers,” there are species occurring in EOA grid cells adjacent to the LHR and others where the next occurrence is about 700 km away. The median distance of the closest occurrences for all “potential arrivers” is 283 km (see
supplementary material—Table S7). Thus, the assessment of the likelihood of “potential arrivers” actually reaching the LHR requires a species-specific interpretation, which is beyond the scope of this study.
Another precondition for “potential arrivers” successfully breeding in the LHR is the availability of suitable habitat. Interestingly, almost half of the “potential arrivers” are or have historically been occasional or regular breeders in Lower Saxony, the federal state the LHR is situated in (see
supplementary material—Table S7), showing that this precondition could be met.
4.2. Methodological Limitations
Our approach has some methodological limitations that need to be taken into account when interpreting the results and considering why a species might be present in the LHR, but absent in the FCARs. Some limitations are “data intrinsic.” In order to avoid false absences, we only included EOA grid cells with “good coverage,” meaning that data was provided for at least 75% of the expected breeding species. However, even though this is the atlas’s best survey category, it may be that there is simply no existing data for up to 25% of the species. Furthermore, the data we used is presence/absence data and density was not taken into account. Thus, our approach is limited to assumptions about changes in the distribution of a species because conclusions about population trends within an area are not possible. From a nature conservation point of view, it was important to us to include all species into the analyses, regardless of their frequency within the region. Furthermore, we wanted to use a European-wide data set that does not depend on the availability of regional data. Both aspects imply that the data set therefore in some cases includes occasional breeders or populations of local escapees from captivity that have native populations elsewhere in Europe. We argue that these exceptional cases are of minor importance for assessing trends within a region as we take into account a large assemblage of species.
In our study, we used the climatic variables for our projections that best reflect the distribution of the species community of the LHR. Nonetheless, the effect of other likely important climate variables could not be tested or included because the data was not available. One such climate variable is the incidence of extreme weather events, which are known to pose a risk especially to small populations [
52].
Furthermore, there might be other reasons apart from climate for a species to be absent from an area. Concerning land use, a few species that only occur in coastal habitats and not in inland wetlands, might be classified as “potential leavers” due to missing coastal/maritime habitats in the FCARs (e.g., European Herring Gull (
Larus argentatus)). However, the percentage of coastal habitats in the LHR is marginal and thus this effect is negligible for the bird community. Furthermore, peat bogs are also missing in the FCARs, but are present in the LHR. In contrast to the sea (and related maritime habitats), which does not depend on climate (
cf. [
12]), the distribution of peat bogs clearly does (
cf. [
53]). For species depending on peat bogs we therefore see climate and not land use as the main cause for their absence in the FCARs. Furthermore, we have to consider that species are absent in some cases because of special habitat requirements, which cannot be adequately reflected by the CORINE land cover data. Another source of uncertainty is that not all birds are in equilibrium with the climate within their distribution ranges. Many large raptors, for instance, have a patchy distribution in Europe due to historic or recent persecution [
8]. Therefore, when regarding “potential leavers” from a community, assessing a single species requires additional consideration. However, our approach is not designed for making predictions for single species, but for detecting trends within a region.
The method we developed can help to identify species groups that are principally able to cope with the future climatic conditions in a given study area. Nevertheless, climate is not the only factor influencing bird populations within a region and thus occurrence in projected future climates does not automatically mean that a species is not at risk of future declines. Currently, habitat loss and habitat degradation are seen as the major threats to breeding birds in Germany [
54]. Hence, species that are already endangered due to land use change have to be monitored carefully, even if they are predicted to occur in future climates. However, Lemoine
et al. [
55] estimate that climate change has already more influence on population trends of European birds than land use alterations. Additionally, Jiguet
et al. [
56] found that species with the lowest thermal maxima showed the sharpest declines in Europe recently and Gregory
et al. [
57] showed that climate change has increasingly impacted European bird populations in the past twenty years.
In our study, we include both migrating and sedentary birds. It is important to keep in mind that migrating birds might additionally be affected by impacts at stop-over and wintering sites. These impacts cannot be included in our approach.
It is necessary to interpret the results of this study cautiously with regards to its limitations. Despite these limitations, we believe the detected overall trends to be a benchmark for estimating future changes in the examined region facing climate change and for developing strategies for nature conservation as the results are based on a range of climate change projections. However, it must not be forgotten that climate change is only one threat to biodiversity and others such as habitat loss and degradation should not be neglected.
4.3. Implications for Nature Conservation
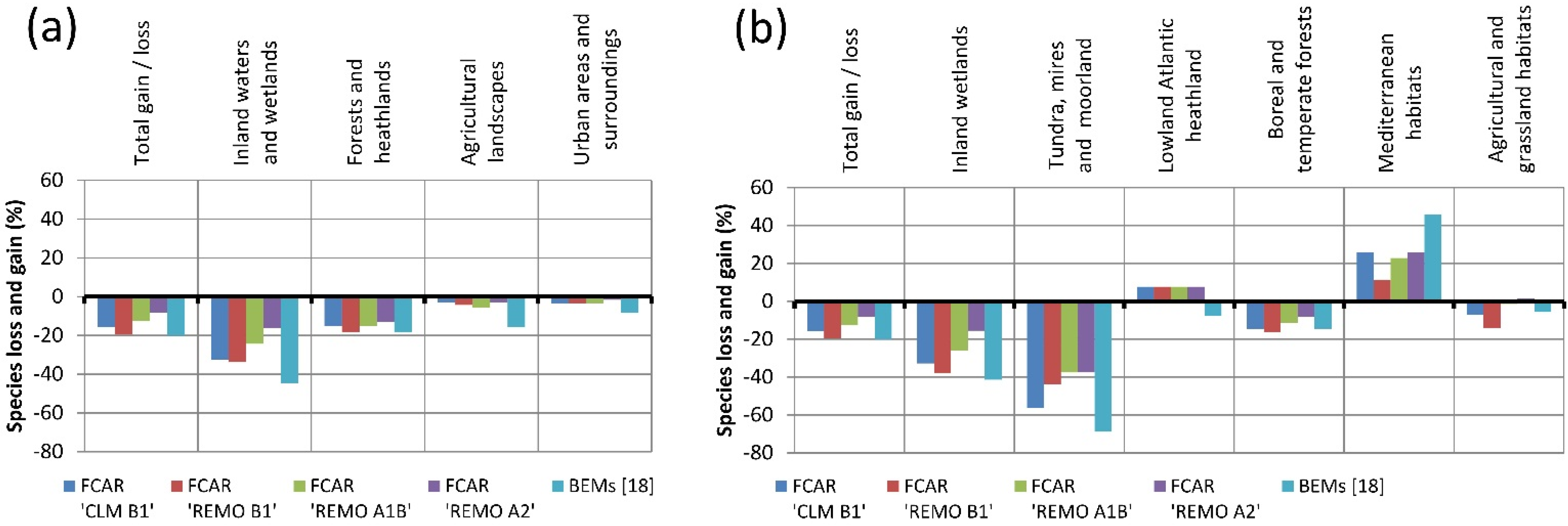
The results from our FCAR approach and the results of the bioclimatic envelope models by Huntley
et al. [
18] both point in the same direction, indicating that species of inland wetlands might be more impacted by climatic changes than other species groups in this region. However, as we include habitat data in our analyses, we found that climate is not the only important factor determining the occurrence of these species. With our approach we showed that in contrast to species of most other habitat types, the percentage of species of “inland waters and wetlands/inland wetlands” for both of the utilized habitat data sets is significantly positively correlated with the percentage of inland wetlands and inland waters in that area.
Thus, we assume that for the Lüneburg Heath region the extent to which wetland species are affected by climate change can be influenced by the availability and extent of suitable habitat for these species. This is important from a nature conservation point of view, suggesting that more effort in wetland conservation and restoration in the examined region might mitigate climate change impacts on species for these particular habitats.
For future research, testing this approach in other regions and including habitat and species data with a higher resolution is desirable.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


 )
) )
) )
)