
Habitat Distribution Pattern of François’ Langur in a Human-Dominated Karst Landscape: Implications for Its Conservation

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Presence Data Collection
2.3. Anthropogenic Disturbance Distribution Data
2.4. Distribution Area Estimation
Spatial Analysis of Habitat Characteristics and Anthropogenic Disturbance
2.5. Landscape Pattern Analysis
2.6. Habitat Classification and Metric Calculation
3. Result
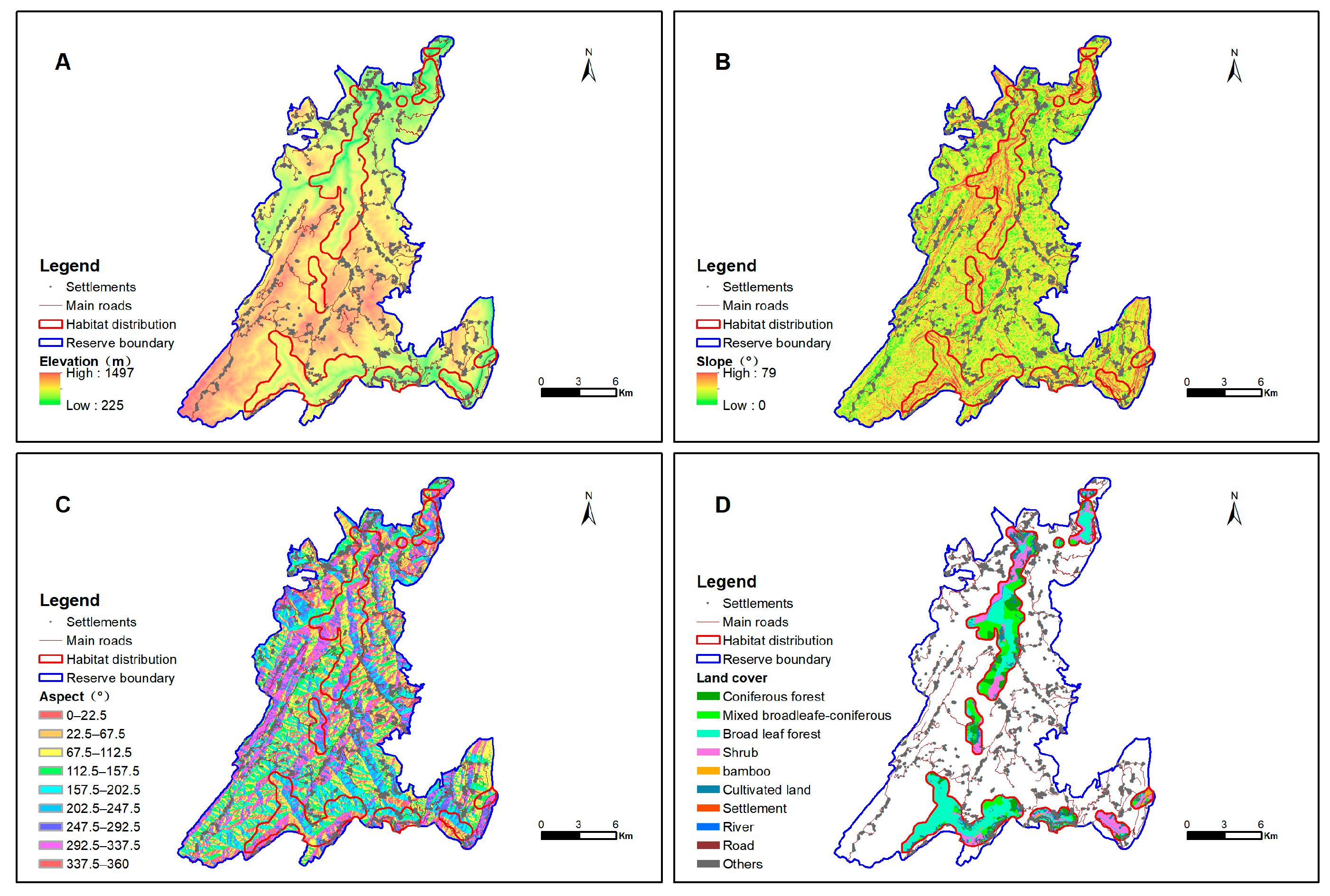
3.1. Topographic and Land Cover Differentiation in the MNNR
3.2. Habitat and Anthropogenic Disturbance Distribution Patterns
3.2.1. Altitudinal Patterns
3.2.2. Slope Gradient Distribution
3.2.3. Slope Aspect Neutrality
3.2.4. Vegetation Type and Landscape Pattern Characteristics of François’ Langur Habitat Distribution
4. Discussion
Implications for Conservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hull, V.; Zhang, J.; Huang, J.; Zhou, S.; Viña, A.; Shortridge, A.; Li, R.; Liu, D.; Xu, W.; Ouyang, Z.; et al. Habitat Use and Selection by Giant Pandas. PLoS ONE 2016, 11, e0162266. [Google Scholar] [CrossRef]
- Van Beest, F.M.; Loe, L.E.; Mysterud, A.; Milner, J.M. Comparative Space Use and Habitat Selection of Moose Around Feeding Stations. J. Wildl. Manag. 2010, 74, 219–227. [Google Scholar] [CrossRef]
- Rosenberg, D.K.; McKelvey, K.S. Estimation of Habitat Selection for Central-Place Foraging Animals. J. Wildl. Manag. 1999, 63, 1028–1038. [Google Scholar] [CrossRef]
- Bai, W.; Connor, T.; Zhang, J.; Yang, H.; Dong, X.; Gu, X.; Zhou, C. Long-Term Distribution and Habitat Changes of Protected Wildlife: Giant Pandas in Wolong Nature Reserve, China. Environ. Sci. Pollut. Res. 2018, 25, 11400–11408. [Google Scholar] [CrossRef]
- Johnson, C.J.; Nielsen, S.E.; Merrill, E.H.; McDonald, T.L.; Boyce, M.S. Resource Selection Functions Based on Use-Availability Data: Theoretical Motivation and Evaluation Methods. J. Wildl. Manag. 2006, 70, 347–357. [Google Scholar] [CrossRef]
- Gillies, C.S.; Hebblewhite, M.; Nielsen, S.E.; Krawchuk, M.A.; Aldridge, C.L.; Frair, J.L.; Saher, D.J.; Stevens, C.E.; Jerde, C.L. Application of Random Effects to the Study of Resource Selection by Animals. J. Anim. Ecol. 2006, 75, 887–898. [Google Scholar] [CrossRef]
- Rouys, S.; Theuerkauf, J.; Krasińska, M. Accuracy of Radio-Tracking to Estimate Activity and Distances Walked by European Bison in the Białowieża Forest, Poland. Acta Theriol. 2001, 46, 319–326. [Google Scholar]
- Belant, J.; Follmann, E. Sampling Considerations for American Black and Brown Bear Home Range and Habitat Use. Ursus 2002, 13, 299–315. [Google Scholar]
- Turner, L.W.; Udal, M.C.; Larson, B.T.; Shearer, S.A. Monitoring Cattle Behavior and Pasture Use with GPS and GIS. Can. J. Anim. Sci. 2000, 80, 405–413. [Google Scholar] [CrossRef]
- Liu, J.; Bhumpakphan, N. Comparison of Activity Budgets, Diet, and Habitat Utilization between Provisioned and Wild Groups of the François’ Langur (Trachypithecus francoisi) in Mayanghe National Nature Reserve, China. Folia Primatol. 2020, 91, 15–30. [Google Scholar] [CrossRef]
- Zhou, Q.; Huang, C.; Li, Y.; Cai, X. Ranging Behavior of the François’ Langur (Trachypithecus francoisi) in the Fusui Nature Reserve, China. Primates 2007, 48, 320–323. [Google Scholar] [CrossRef]
- Zeng, Y.J.; Xu, J.L.; Wang, Y.; Zhou, C.F. Habitat Association and Conservation Implications for Endangered François’ Langur (Trachypithecus francoisi). PLoS ONE 2013, 8, e75661. [Google Scholar] [CrossRef]
- Hu, G. Dietary Breadth and Resource Use of François’ Langur in a Seasonal and Disturbed Habitat. Am. J. Primatol. 2011, 73, 1176–1187. [Google Scholar] [CrossRef]
- Zhou, Q.; Wei, F.; Huang, C.; Li, M.; Ren, B.; Luo, B. Seasonal Variation in the Activity Patterns and Time Budgets of Trachypithecus francoisi in the Nonggang Nature Reserve, China. Int. J. Primatol. 2007, 28, 657–671. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Fahrig, L. Why is a Landscape Perspective Important in Studies of Primates? Am. J. Primatol. 2014, 76, 901–909. [Google Scholar] [CrossRef]
- Cruzan, M.B.; Hendrickson, E.C. Landscape Genetics of Plants: Challenges and Opportunities. Plant Commun. 2020, 1, 100100. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Broekman, M.J.E.; Hilbers, J.P.; Schipper, A.M.; Benítez-López, A.; Santini, L.; Huijbregts, M.A.J. Time-Lagged Effects of Habitat Fragmentation on Terrestrial Mammals in Madagascar. Conserv. Biol. 2022, 36, e13942. [Google Scholar] [CrossRef]
- Rybicki, J.; Abrego, N.; Ovaskainen, O. Habitat Fragmentation and Species Diversity in Competitive Communities. Ecol. Lett. 2020, 23, 506–517. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A. Comparative Evaluation of Experimental Approaches to the Study of Habitat Fragmentation Effects. Ecol. Appl. 2002, 12, 335–345. [Google Scholar] [CrossRef]
- Krishnamurthy, R. Analysis of Lion-Tailed Macaque Habitat Fragmentation Using Satellite Imagery. Curr. Sci. 1994, 66, 283–291. [Google Scholar]
- Isabirye-Basuta, G.M.; Lwanga, J.S. Primate Populations and Their Interactions with Changing Habitats. Int. J. Primatol. 2008, 29, 35–48. [Google Scholar] [CrossRef]
- Chapman, C.A.; Chapman, L.J.; Vulinec, K.; Zanne, A.; Lawes, M.J. Fragmentation and Alteration of Seed Dispersal Processes: An Initial Evaluation of Dung Beetles, Seed Fate, and Seedling Diversity. Biotropica 2003, 35, 382–393. [Google Scholar] [CrossRef]
- Chapman, C.A.; Peres, C.A. Primate Conservation in the New Millennium: The Role of Scientists. Evol. Anthropol. 2001, 10, 16–33. [Google Scholar] [CrossRef]
- Chapman, C.A.; Chapman, L.J.; Gillespie, T.R. Scale Issues in the Study of Primate Foraging: Red Colobus of Kibale National Park. Am. J. Phys. Anthropol. 2002, 117, 349–363. [Google Scholar] [CrossRef]
- Chapman, C.A.; Wasserman, M.D.; Gillespie, T.R.; Speirs, M.L.; Lawes, M.J.; Saj, T.L.; Ziegler, T.E. Do Food Availability, Parasitism, and Stress Have Synergistic Effects on Red Colobus Populations Living in Forest Fragments? Am. J. Phys. Anthropol. 2006, 131, 525–534. [Google Scholar] [CrossRef]
- Villard, M.-A.; Metzger, J.P. Beyond the Fragmentation Debate: A Conceptual Model to Predict When Habitat Configuration Really Matters. J. Appl. Ecol. 2014, 51, 309–318. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Mandujano, S.; Benítez-Malvido, J. Landscape Attributes Affecting Patch Occupancy by Howler Monkeys (Alouatta palliata mexicana) at Los Tuxtlas, Mexico. Am. J. Primatol. 2008, 70, 69–77. [Google Scholar] [CrossRef]
- De Souza, R.; Beneduzi, A.; Ambrosini, A.; Da Costa, P.B.; Meyer, J.; Vargas, L.K.; Schoenfeld, R.; Passaglia, L.M.P. The Effect of Plant Growth-Promoting Rhizobacteria on the Growth of Rice (Oryza sativa L.) Cropped in Southern Brazilian Fields. Plant Soil 2013, 366, 585–603. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; González-Perez, I.M.; Garmendia, A.; Solà, M.; Estrada, A. The Relative Impact of Forest Patch and Landscape Attributes on Black Howler Monkey Populations in the Fragmented Lacandona Rainforest, Mexico. Landsc. Ecol. 2013, 28, 1717–1727. [Google Scholar] [CrossRef]
- Thornton, D.H.; Branch, L.C.; Sunquist, M.E. The Relative Influence of Habitat Loss and Fragmentation: Do Tropical Mammals Meet the Temperate Paradigm? Ecol. Appl. 2011, 21, 2324–2333. [Google Scholar] [CrossRef]
- Ordóñez-Gómez, J.D.; Cristóbal-Azkarate, J.; Arroyo-Rodríguez, V.; Santillán-Doherty, A.M.; Valdez, R.A.; Romano, M.C. Proximal and Distal Predictors of the Spider Monkey’s Stress Levels in Fragmented Landscapes. PLoS ONE. 2016, 11, e0149671. [Google Scholar] [CrossRef]
- Han, Z.; Hu, G.; Wu, S.; Cao, C.; Dong, X. A Census and Status Review of the Endangered François’ langur in Chongqing, China. Oryx 2013, 47, 128–133. [Google Scholar] [CrossRef]
- Li, Y.; Huang, C.; Ding, P.; Tang, Z.; Wood, C. Dramatic Decline of François’ langur in Guangxi Province, China. Oryx 2007, 41, 38–43. [Google Scholar] [CrossRef]
- Wang, S.L.; Luo, Y.; Cui, G.F. Sleeping Site Selection of François’s langur (Trachypithecus francoisi) in Two Habitats in Mayanghe National Nature Reserve, Guizhou, China. Primates 2011, 52, 51–60. [Google Scholar] [CrossRef]
- Chen, T.; Huang, Z.H.; Huang, C.M.; Wei, H.; Zhou, Q.H. Positional Behaviours of François’ Langur (Trachypithecus francoisi) in the Limestone Forest of Nonggang, Guangxi, South-West China. Folia Primatol. 2020, 91, 170–187. [Google Scholar] [CrossRef]
- Pascual-Hortal, L.; Saura, S. Comparison and Development of New Graph-Based Landscape Connectivity Indices: Towards the Prioritization of Habitat Patches and Corridors for Conservation. Landsc. Ecol. 2006, 21, 959–967. [Google Scholar] [CrossRef]
- Worton, B.J. Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Hemsong, G.; Johnson, P.; South, A.; Kenward, R.; Ripley, R.; Macdonald, D. Are Kernels the Mustard? Data from Global Positioning System (GPS) Collars Suggests Problems for Kernel Home-Range Analyses with Least-Squares Cross-Validation. J. Anim. Ecol. 2005, 74, 455–463. [Google Scholar] [CrossRef]
- Horne, J.S.; Garton, E.O. Likelihood Cross-Validation Versus Least Squares Cross-Validation for Choosing the Smoothing Parameter in Kernel Home-Range Analysis. J. Wildl. Manag. 2006, 70, 641–648. [Google Scholar] [CrossRef]
- Han, J.; Fan, X.; Williams, G.M.; Zou, Q.; Dong, B. Evaluating habitat selection of François’ langur in the karst mountains of China: Implications for conservation strategies. Glob. Ecol. Conserv 2024, 56, e03330. [Google Scholar] [CrossRef]
- Grueter, C. An Observation of François’ Langurs Using Caves at Mayanghe National Nature Reserve, Guizhou, China. Zool. Res. 2006, 27, 555–558. [Google Scholar]
- Zheng, J.J.; Zhang, K.C.; Liang, J.P.; Li, Y.B.; Huang, Z.H. Food Availability, Temperature, and Day Length Drive Seasonal Variations in the Positional Behavior of White-Headed Langurs in the Limestone Forests of Southwest Guangxi, China. Ecol. Evol. 2021, 11, 14857–14872. [Google Scholar] [CrossRef]
- Zhou, Q.H.; Huang, Z.H.; Wei, H.; Huang, C.M. Variations in Diet Composition of Sympatric Trachypithecus francoisi and Macaca assamensis in the Limestone Habitats of Nonggang, China. Zool. Res. 2018, 39, 284–290. [Google Scholar] [CrossRef]
- Zhou, Q.H.; Tang, X.P.; Huang, H.L.; Huang, C.M. Factors Affecting the Ranging Behavior of White-Headed Langurs (Trachypithecus leucocephalus). Int. J. Primatol. 2011, 32, 511–523. [Google Scholar] [CrossRef]
- Zhou, Q.H.; Luo, B.; Wei, F.W.; Huang, C.M. Habitat Use and Locomotion of the François’ Langur (Trachypithecus francoisi) in Limestone Habitats of Nonggang, China. Integr. Zool. 2013, 8, 346–355. [Google Scholar] [CrossRef]
- Huang, C.; Zhou, Q.; Li, Y.; Cai, X.; Wei, F. Activity Rhythm and Diurnal Time Budget of François Langur (Trachypithecus françoisi) in Guangxi, China. Acta Theriol. Sin. 2006, 26, 380–386. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metric | Full Name | Unit | Description |
|---|---|---|---|
| CA | Class Area | m2 | Total area of a specific land cover or landscape class. |
| NP | Number of Patches | Count | Number of discrete patches of a particular class, indicating fragmentation. |
| MPS | Mean Patch Size | m2 | Average area of all patches in a given class (MPS = CA/NP). |
| PLAND | Percentage of Landscape | % | Proportion of the landscape occupied by a specific class. |
| LPI | Largest Patch Index | % | Proportion of the total landscape area comprising the largest patch. |
| ED | Edge Density | m/ha or m/km2 | Total edge length per unit area, indicating boundary complexity. |
| TE | Total Edge | m | Sum of all edge lengths in the landscape. |
| CONTAG | Contagion Index | Dimensionless (0–100) | Measures the extent of clumping or interspersion among patch types. |
| SHDI | Shannon’s Diversity Index | Dimensionless | Quantifies landscape diversity based on patch type richness and evenness. |
| SHEI | Shannon’s Evenness Index | Dimensionless (0–1) | Measures the evenness of the distribution among patch types. |
| AI | Aggregation Index | % | Indicates the degree to which patches of the same class are aggregated. |
| (A) Topographic Conditions | |||||
|---|---|---|---|---|---|
| Category | Code | Class (Unit) | Area (km2) | Proportion (%) | |
| Elevation | E1 | <500 m | 34.10 | 10.96 | |
| E2 | 500–800 m | 101.27 | 32.55 | ||
| E3 | 800–1100 m | 122.06 | 39.23 | ||
| E4 | >1100 m | 53.70 | 17.26 | ||
| Slope | S1 | <10° | 43.90 | 14.11 | |
| S2 | 10–20° | 117.12 | 37.64 | ||
| S3 | 20–30° | 92.49 | 29.73 | ||
| S4 | 30–40° | 38.92 | 12.51 | ||
| S5 | 40–50° | 14.63 | 4.70 | ||
| S6 | 50–60° | 3.52 | 1.13 | ||
| S7 | >60° | 0.56 | 0.18 | ||
| Aspect | A1–A8 | All directions | – | Evenly distributed across directions; no dominant aspect detected | |
| (B) Land Cover Composition | |||||
| Code | Land Cover Type | Area (km2) | Proportion (%) | ||
| V1 | Cultivated land | 81.47 | 26.18 | ||
| V2 | Shrubland | 52.29 | 16.81 | ||
| V3 | Others (bare land, construction) | 13.09 | 4.21 | ||
| V4 | Broadleaf forest | 73.71 | 23.69 | ||
| V5 | Mixed broadleaf–coniferous forest | 50.99 | 16.39 | ||
| V6 | Coniferous forest | 38.87 | 12.49 | ||
| V7 | Bamboo | 0.72 | 0.23 | ||
| (C)Human Disturbance Distribution by Elevation and Slope | |||||
| Type | Category | Code | Class | Value | Proportion (%) |
| Cultivated land | Slope | S1 | <10° | 21.19 km2 | 20.47 |
| Slope | S2 | 10–20° | 49.28 km2 | 47.60 | |
| Slope | S3 | 20–30° | 25.66 km2 | 24.79 | |
| Elevation | E2 | 500–800 m | 39.82 km2 | 38.46 | |
| Elevation | E3 | 800–1100 m | 35.51 km2 | 34.30 | |
| Roads | Slope | S2 | 10–20° | 176.90 km | 43.60 |
| Elevation | E2 | 500–800 m | 155.42 km | 38.31 | |
| Settlements | Slope | S2 | 10–20° | 1966 sites | 44.00 |
| Elevation | E2 | 500–800 m | 1966 sites | 44.00 | |
| Type | CA (m2) | NP | MPS | PLAND | LPI | ED | TE (m) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Used Area | Control Area | Used Area | Control Area | Used Area | Control Area | Used Area | Control Area | Used Area | Control Area | Used Area | Control Area | Used Area | Control Area | |
| Optimal | 2560.23 | 5860.71 | 80 | 315 | 32.00 | 18.61 | 37.23 | 20.43 | 20.56 | 8.98 | 28.53 | 23.00 | 196,260 | 659,880 |
| Suitable | 2579.49 | 9313.74 | 110 | 474 | 23.45 | 19.65 | 37.51 | 32.47 | 10.23 | 4.96 | 44.21 | 47.86 | 304,080 | 1,372,860 |
| Marginal | 1503.36 | 12,251.43 | 267 | 803 | 5.63 | 15.26 | 21.86 | 42.71 | 1.74 | 6.97 | 43.93 | 76.01 | 302,130 | 2,180,460 |
| Type | CONTAG | SHDI | SHEI | AI |
|---|---|---|---|---|
| Control area | 47.46 | 1.20 | 0.75 | 86.10 |
| Distribution area | 49.01 | 1.21 | 0.74 | 88.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.; Fan, X.; Wu, A.; Dong, B.; Zou, Q. Habitat Distribution Pattern of François’ Langur in a Human-Dominated Karst Landscape: Implications for Its Conservation. Diversity 2025, 17, 547. https://doi.org/10.3390/d17080547
Han J, Fan X, Wu A, Dong B, Zou Q. Habitat Distribution Pattern of François’ Langur in a Human-Dominated Karst Landscape: Implications for Its Conservation. Diversity. 2025; 17(8):547. https://doi.org/10.3390/d17080547
Chicago/Turabian StyleHan, Jialiang, Xing Fan, Ankang Wu, Bingnan Dong, and Qixian Zou. 2025. "Habitat Distribution Pattern of François’ Langur in a Human-Dominated Karst Landscape: Implications for Its Conservation" Diversity 17, no. 8: 547. https://doi.org/10.3390/d17080547
APA StyleHan, J., Fan, X., Wu, A., Dong, B., & Zou, Q. (2025). Habitat Distribution Pattern of François’ Langur in a Human-Dominated Karst Landscape: Implications for Its Conservation. Diversity, 17(8), 547. https://doi.org/10.3390/d17080547