
Differential Expression of Hsp100 Gene in Scrippsiella acuminata: Potential Involvement in Life Cycle Transition and Dormancy Maintenance


 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Scrippsiella acuminata Culture Maintenance
2.2. Full-Length cDNA Cloning of SaHsp100
2.3. Sequence Homology and Phylogenetic Analysis
2.4. Resting Cysts Preparation and Sample Treatment
2.5. Transcriptional Profiles of SaHsp100 with qPCR Detection
2.6. PRM-Based Quantification of Hsp100 Protein Levels in S. acuminata
2.6.1. Protein Extraction
2.6.2. Protein Digestion
2.6.3. Liquid Chromatography (LC)-PRM-Mass Spectrometry (MS)
2.6.4. Data Analysis
3. Results
3.1. The Full-Length cDNA Sequence of SaHsp100
3.2. Homology and Phylogenetic Analysis of SaHsp100
3.3. Differential Transcription Profiles of SaHsp100 at Different Stages of Growth and Life Cycle
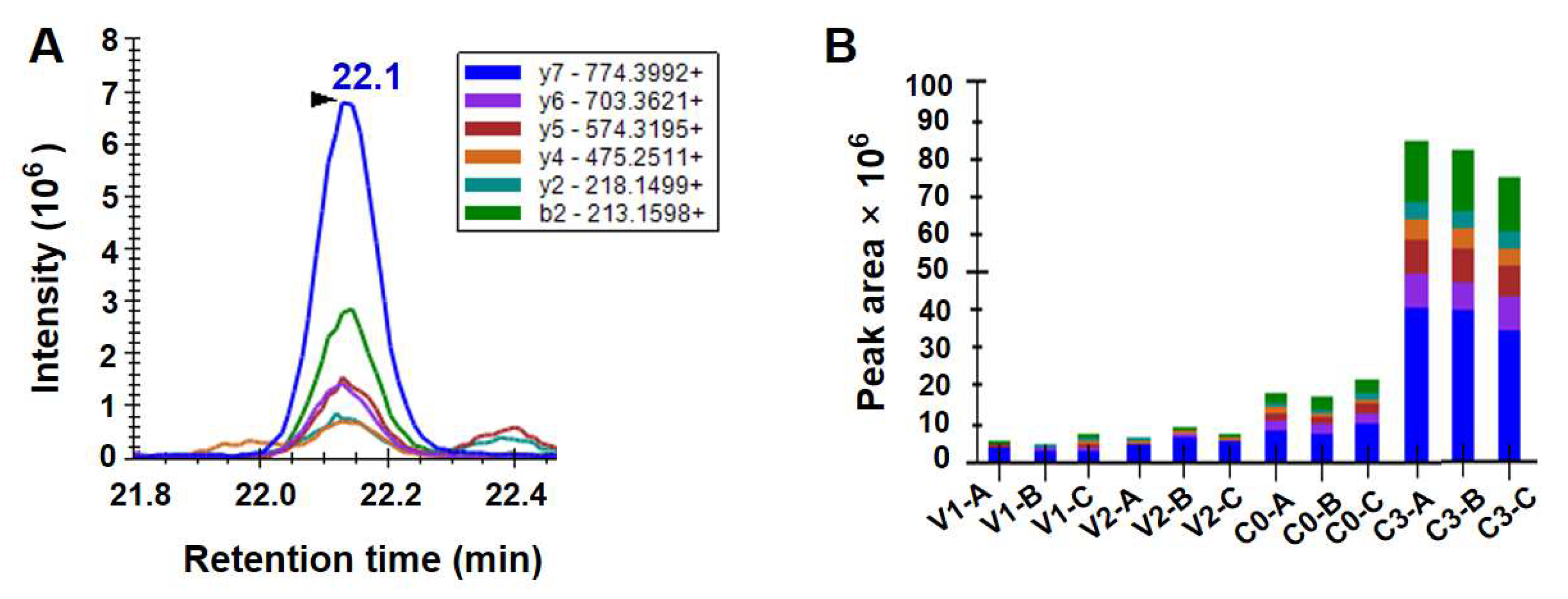
3.4. Protein Levels of Hsp100 Determined Using the Method of PRM
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HAB | Harmful algal bloom |
| Hsp100 | Heat shock protein 100 |
| PRM | Parallel reaction monitoring |
| RACE | Rapid amplification of cDNA ends |
| ORF | Open Reading Frame |
References
- Taylor, F.; Hoppenrath, M.; Saldarriaga, J.F. Dinoflagellate diversity and distribution. Biodivers. Conserv. 2008, 17, 407–418. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Jeong, H.J.; Kang, H.C.; Lim, A.S.; Jang, S.H.; Lee, K.; Lee, S.Y.; Ok, J.H.; You, J.H.; Kim, J.H.; Lee, K.H. Feeding diverse prey as an excellent strategy of mixotrophic dinoflagellates for global dominance. Sci. Adv. 2021, 7, eabe4214. [Google Scholar] [CrossRef]
- Geng, H.-X.; Kong, F.-Z.; Wang, J.-X.; Zhang, Q.-C.; Li, F.; Hong, X.; Song, M.-J.; Lian, Z.; Cai, Y.-L.; Yu, R.-C. An unusual winter bloom of dinoflagellates with notable damage to kelp cultivation around Shandong peninsula, China. Mar. Environ. Res. 2024, 201, 106687. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Anderson, D.M.; Belin, C.; Bottein, M.-Y.D.; Bresnan, E.; Chinain, M.; Enevoldsen, H.; Iwataki, M.; Karlson, B.; McKenzie, C.H. Perceived global increase in algal blooms is attributable to intensified monitoring and emerging bloom impacts. Commun. Earth Environ. 2021, 2, 117. [Google Scholar] [CrossRef]
- Bravo, I.; Figueroa, R.I. Towards an ecological understanding of dinoflagellate cyst functions. Microorganisms 2014, 2, 11–32. [Google Scholar] [CrossRef]
- Anderson, D.M.; Keafer, B.A.; Kleindinst, J.L.; McGillicuddy, D.J., Jr.; Martin, J.L.; Norton, K.; Pilskaln, C.H.; Smith, J.L.; Sherwood, C.R.; Butman, B. Alexandrium fundyense cysts in the Gulf of Maine: Long-term time series of abundance and distribution, and linkages to past and future blooms. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 103, 6–26. [Google Scholar] [CrossRef]
- Rodríguez-Villegas, C.; Figueroa, R.I.; Baldrich, Á.M.; Pérez-Santos, I.; Díaz, M.; Tomasetti, S.J.; Seguel, M.; Álvarez, G.; Salgado, P.; Díaz, P.A. Small and patchy is enough: An example about how toxic HAB events can spread through low resting cyst loads. Harmful Algae 2023, 129, 102495. [Google Scholar] [CrossRef]
- Deng, Y.; Yue, C.; Yang, H.; Li, F.; Hu, Z.; Shang, L.; Chai, Z.; Lin, S.; Tang, Y.Z. Broad active metabolic pathways, autophagy, and antagonistic hormones regulate dinoflagellate cyst dormancy in marine sediments. Sci. Adv. 2025, 11, eads7789. [Google Scholar] [CrossRef]
- Morimoto, R.I. The Heat Shock Response: Systems Biology of Proteotoxic Stress in Aging and Disease; Cold Spring Harbor Symposia on Quantitative Biology: New York, NY, USA, 2011; pp. 91–99. [Google Scholar]
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King. Saud. Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell Bio. 2013, 14, 630–642. [Google Scholar] [CrossRef]
- Kumar, R.; Khungar, L.; Shimphrui, R.; Tiwari, L.D.; Tripathi, G.; Sarkar, N.K.; Agarwal, S.-K.; Agarwal, M.; Grover, A. AtHsp101 research sets course of action for the genetic improvement of crops against heat stress. J. Plant. Biochem. Biot. 2020, 29, 715–732. [Google Scholar] [CrossRef]
- Santos, T.F.; Pereira, H.; Schüler, L.; Maia, I.B.; Jacinto, R.; Bombo, G.; Pinheiro, F.; Barreira, L.; Varela, J. Enhancement of heat tolerance by salt stress in Tetraselmis striata CTP4: Impacts on HSP gene expression, pigments, and proximal composition. J. Appl. Phycol. 2025, 37, 287–301. [Google Scholar] [CrossRef]
- Agarwal, M.; Katiyar-Agarwal, S.; Grover, A. Plant Hsp100 proteins: Structure, function and regulation. Plant Sci. 2002, 163, 397–405. [Google Scholar] [CrossRef]
- Mogk, A.; Kummer, E.; Bukau, B. Cooperation of Hsp70 and Hsp100 chaperone machines in protein disaggregation. Front. Mol. Biosci. 2015, 2, 22. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.C.; Grover, A. ClpB/Hsp100 proteins and heat stress tolerance in plants. Crit. Rev. Biotechnol. 2016, 36, 862–874. [Google Scholar] [CrossRef]
- Deng, Y.; Hu, Z.; Zhan, Z.; Ma, Z.; Tang, Y. Differential expressions of an Hsp70 gene in the dinoflagellate Akashiwo sanguinea in response to temperature stress and transition of life cycle and its implications. Harmful Algae 2015, 50, 57–64. [Google Scholar] [CrossRef]
- Deng, Y.; Li, F.; Hu, Z.; Yue, C.; Tang, Y.Z. Expression patterns of the heat shock protein 90 (Hsp90) gene suggest its possible involvement in maintaining the dormancy of dinoflagellate resting cysts. Int. J. Mol. Sci. 2021, 22, 11054. [Google Scholar] [CrossRef]
- Parkinson, J.E.; Baumgarten, S.; Michell, C.T.; Baums, I.B.; LaJeunesse, T.C.; Voolstra, C.R. Gene expression variation resolves species and individual strains among coral-associated dinoflagellates within the genus Symbiodinium. Genome Biol. Evol. 2016, 8, 665–680. [Google Scholar] [CrossRef]
- Kretschmann, J.; Elbrächter, M.; Zinssmeister, C.; Soehner, S.; Kirsch, M.; Kusber, W.-H.; Gottschling, M. Taxonomic clarification of the dinophyte Peridinium acuminatum Ehrenb.,≡ Scrippsiella acuminata, comb. nov. (Thoracosphaeraceae, Peridiniales). Phytotaxa 2015, 220, 239–256. [Google Scholar] [CrossRef]
- Deng, Y.; Hu, Z.; Shang, L.; Peng, Q.; Tang, Y.Z. Transcriptomic analyses of Scrippsiella trochoidea reveals processes regulating encystment and dormancy in the life cycle of a dinoflagellate, with a particular attention to the role of abscisic acid. Front. Microbiol. 2017, 8, 2450. [Google Scholar] [CrossRef]
- Wang, H.; Kim, H.; Ki, J.S. Transcriptome survey, molecular identification, and expression analysis of stress-responsive genes in the toxic dinoflagellate Alexandrium pacificum under algicidal agents and metal stresses. J. Appl. Phycol. 2021, 33, 3139–3151. [Google Scholar] [CrossRef]
- Wang, D.Z.; Zhang, Y.J.; Zhang, S.F.; Lin, L.; Hong, H.S. Quantitative proteomic analysis of cell cycle of the dinoflagellate Prorocentrum donghaiense (Dinophyceae). PLoS ONE 2013, 8, 63659. [Google Scholar] [CrossRef]
- Zhang, S.F.; Chen, Y.; Xie, Z.X.; Zhang, H.; Lin, L.; Wang, D.-Z. Unraveling the molecular mechanism of the response to changing ambient phosphorus in the dinoflagellate Alexandrium catenella with quantitative proteomics. J. Proteom. 2019, 196, 141–149. [Google Scholar] [CrossRef]
- Deng, Y.; Hu, Z.; Shang, L.; Chai, Z.; Tang, Y.Z. Transcriptional responses of the heat shock protein 20 (Hsp20) and 40 (Hsp40) genes to temperature stress and alteration of life cycle stages in the harmful alga Scrippsiella trochoidea (Dinophyceae). Biology 2020, 9, 408. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Hu, Z.; Chai, Z.; Tang, Y.Z. Molecular cloning of heat shock protein 60 (Hsp60) and 10 (Hsp10) genes from the cosmopolitan and harmful dinoflagellate Scrippsiella trochoidea and their differential transcriptions responding to temperature stress and alteration of life cycle. Mar. Biol. 2019, 166, 7. [Google Scholar] [CrossRef]
- Guillard, R.R. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals: Proceedings—1st Conference on Culture of Marine Invertebrate Animals Greenport; Plenum Press: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
- Zhang, H.; Hou, Y.; Miranda, L.; Campbell, D.A.; Sturm, N.R.; Gaasterland, T.; Lin, S. Spliced leader RNA trans-splicing in dinoflagellates. Proc. Natl. Acad. Sci. USA 2007, 104, 4618–4623. [Google Scholar] [CrossRef]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M. The Proteomics Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Yue, C.; Chai, Z.; Hu, Z.; Shang, L.; Deng, Y.; Tang, Y.Z. Deficiency of nitrogen but not phosphorus triggers the life cycle transition of the dinoflagellate Scrippsiella acuminata from vegetative growth to resting cyst formation. Harmful Algae 2022, 118, 102312. [Google Scholar] [CrossRef]
- Li, F.; Yue, C.; Deng, Y.; Tang, Y.Z. Characterizing the status of energetic metabolism of dinoflagellate resting cysts under mock conditions of marine sediments via physiological and transcriptional measurements. Int. J. Mol. Sci. 2022, 23, 15033. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Y.; Wang, J.; Yu, W.; Wang, W.; Ma, X. Monitoring of cell viability and proliferation in hydrogel-encapsulated system by resazurin assay. Appl. Biochem. Biotechnol. 2010, 162, 1996–2007. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Zakrajsek, B.A. Effect of experimental treatment on housekeeping gene expression: Validation by real-time, quantitative RT-PCR. J. Biochem. Biophys. Methods 2000, 46, 69–81. [Google Scholar] [CrossRef]
- Peterson, A.C.; Russell, J.D.; Bailey, D.J.; Westphall, M.S.; Coon, J.J. Parallel reaction monitoring for high resolution and high mass accuracy quantitative, targeted proteomics. Mol. Cell. Proteom. 2012, 11, 1475–1488. [Google Scholar] [CrossRef]
- Schaeffer, M.; Gateau, A.; Teixeira, D.; Michel, P.-A.; Zahn-Zabal, M.; Lane, L. The neXtProt peptide uniqueness checker: A tool for the proteomics community. Bioinformatics 2017, 33, 3471–3472. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yue, C.; Deng, Y.; Tang, Y.Z. Full-length transcriptome analysis of a bloom-forming dinoflagellate Scrippsiella acuminata (Dinophyceae). Sci. Data 2025, 12, 352. [Google Scholar] [CrossRef]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef]
- Escher, C.; Reiter, L.; MacLean, B.; Ossola, R.; Herzog, F.; Chilton, J.; MacCoss, M.J.; Rinner, O. Using i RT, a normalized retention time for more targeted measurement of peptides. Proteomics 2012, 12, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Aguilar, J.; Molloy, M.P. Label-free selected reaction monitoring enables multiplexed quantitation of S100 protein isoforms in cancer cells. J. Proteome Res. 2013, 12, 3679–3688. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Chang, C.; Vitek, O. MSstats: Protein Significance Analysis in DDA, SRM and DIA for Label-Free or Label-Based Proteomics Experiments. Available online: http://bioconductor.jp/packages/3.14/bioc/manuals/MSstats/man/MSstats.pdf (accessed on 31 May 2021).
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequences (5′→3′) | Remarks |
|---|---|---|
| Fr-F | GGTGGAGATCCCTTTCACTC | Fragment amplification |
| Fr-R | CCTCGTCATAGCCGACATAG | Fragment amplification |
| 5R-outer | CTTGCGGTACTCGTCGATGGTTGTG | 5′ RACE |
| 5R-inner | TACACCTGGCTCGCCGATGAGCACC | 5′ RACE |
| 3R-outer | GAGTTGCGGCAGGTGCAGGAGCAGA | 3′ RACE |
| 3R-inner | CCAGTGGTCACGGAGGAGGATGTAG | 3′ RACE |
| q100-F | TCCCTTTCACTCCTGCTGC | qPCR for SaHsp100 |
| q100-R | CCACCGTCTTCCTTTACCAA | qPCR for SaHsp100 |
| qUBC-F | GTCTTGACCTACTACGTGGAGC | qPCR for UBC |
| qUBC-R | CGGGCGTTGTACTGATGG | qPCR for UBC |
| anchor primer | GCTGTCAACGATACGCTACGTAACGGCATGACAGTGT | cDNA synthesis |
| Sample ID | Temperature | Oxygen | Light | Storage Period | Remarks |
|---|---|---|---|---|---|
| V1 | 21 °C | Yes | Yes | Not applicable | Vegetative cells at exponential growth stage |
| V2 | 21 °C | Yes | Yes | Not applicable | Vegetative cells at stationary growth stage |
| C0 | 21 °C | Yes | Yes | Not applicable | Newly formed resting cysts |
| C1 | 4 °C | No | No | 1 month | Resting cysts stored under conditions typically observed in marine sediments |
| C2 | 4 °C | No | No | 2 months | |
| C3 | 4 °C | No | No | 3 months |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Shang, L.; Zou, H.; Sun, C.; Hu, Z.; Tang, Y.Z.; Deng, Y. Differential Expression of Hsp100 Gene in Scrippsiella acuminata: Potential Involvement in Life Cycle Transition and Dormancy Maintenance. Diversity 2025, 17, 519. https://doi.org/10.3390/d17080519
Li F, Shang L, Zou H, Sun C, Hu Z, Tang YZ, Deng Y. Differential Expression of Hsp100 Gene in Scrippsiella acuminata: Potential Involvement in Life Cycle Transition and Dormancy Maintenance. Diversity. 2025; 17(8):519. https://doi.org/10.3390/d17080519
Chicago/Turabian StyleLi, Fengting, Lixia Shang, Hanying Zou, Chengxing Sun, Zhangxi Hu, Ying Zhong Tang, and Yunyan Deng. 2025. "Differential Expression of Hsp100 Gene in Scrippsiella acuminata: Potential Involvement in Life Cycle Transition and Dormancy Maintenance" Diversity 17, no. 8: 519. https://doi.org/10.3390/d17080519
APA StyleLi, F., Shang, L., Zou, H., Sun, C., Hu, Z., Tang, Y. Z., & Deng, Y. (2025). Differential Expression of Hsp100 Gene in Scrippsiella acuminata: Potential Involvement in Life Cycle Transition and Dormancy Maintenance. Diversity, 17(8), 519. https://doi.org/10.3390/d17080519