
The Co-Occurrence of Zooplankton and Phytoplankton in Shengjin Lake, a Typical Yangtze-Connected Lake in China

Abstract
1. Introduction
2. Materials and Methods
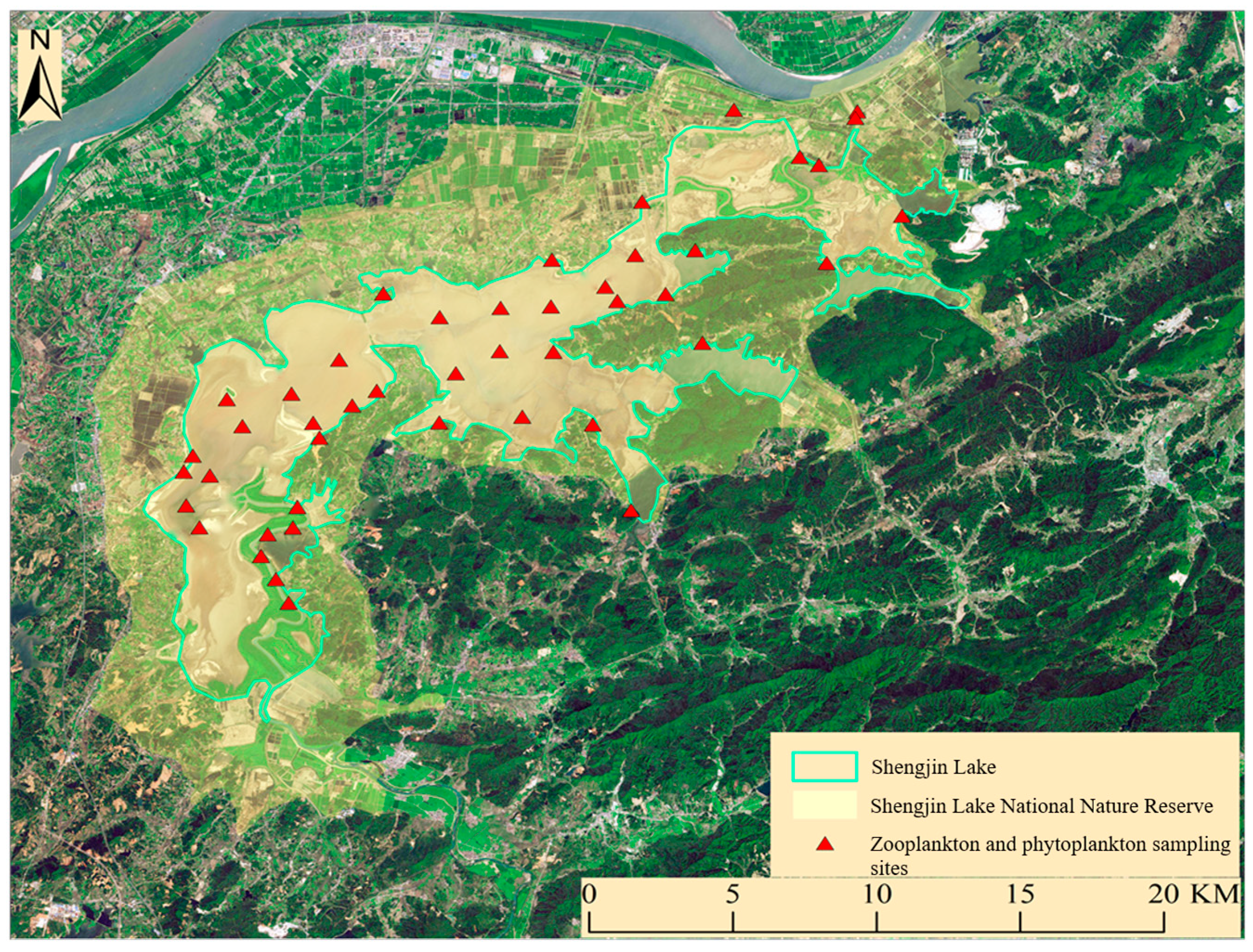
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
2.3.1. Dominant Species and Environmental Factors
2.3.2. Model Structure Setup and Fitting
2.3.3. Data Analysis Tools
3. Results
3.1. Biological and Environmental Factors
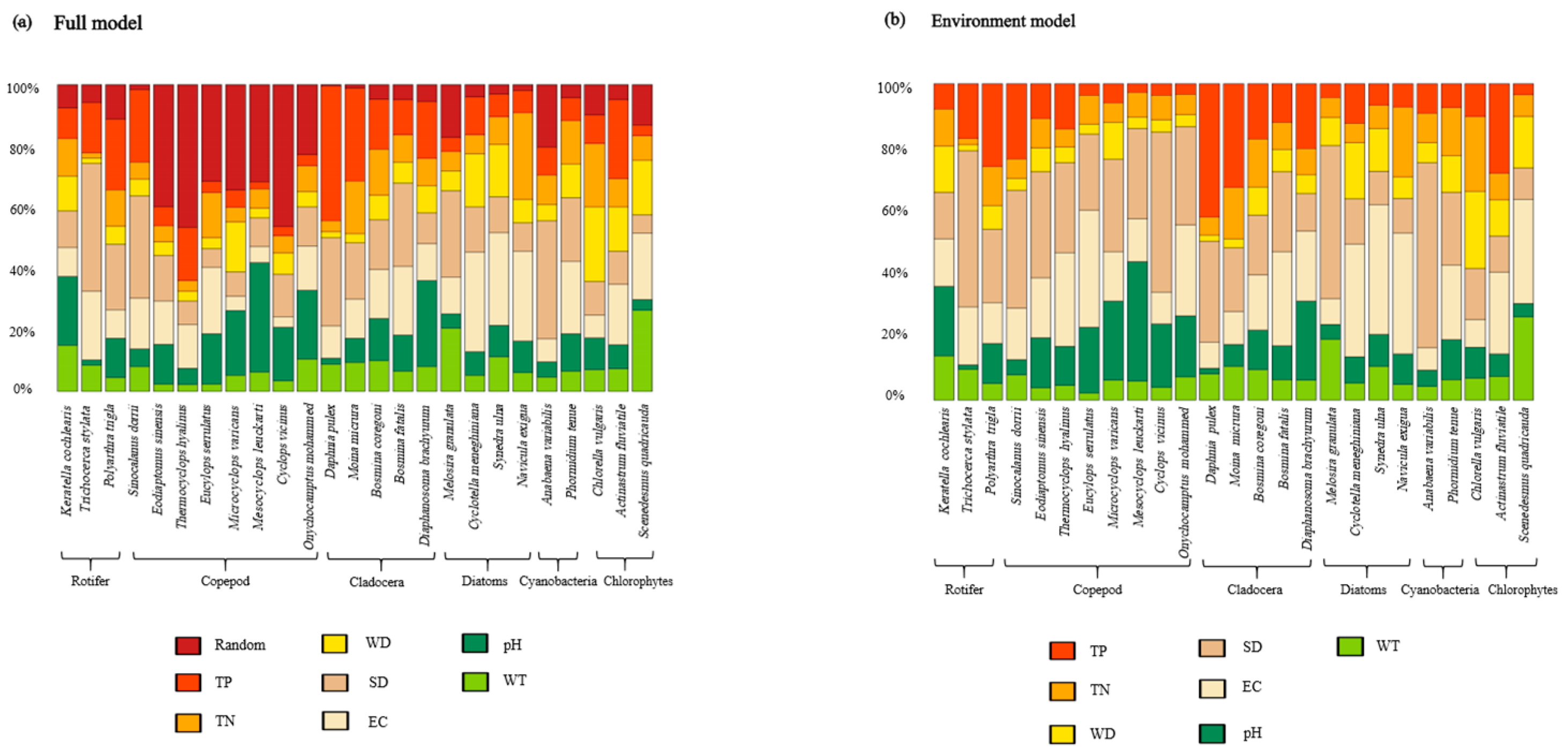
3.2. Model Evaluation
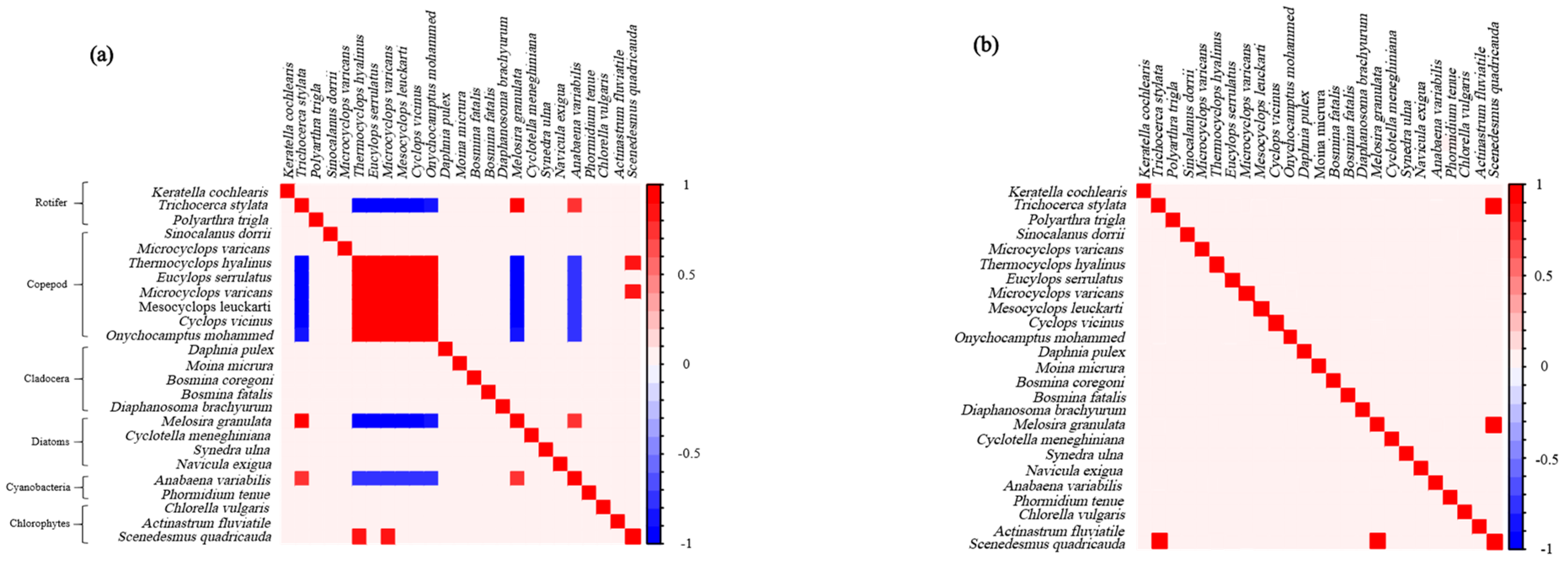
3.3. The Co-Occurrence Between Zooplankton and Phytoplankton
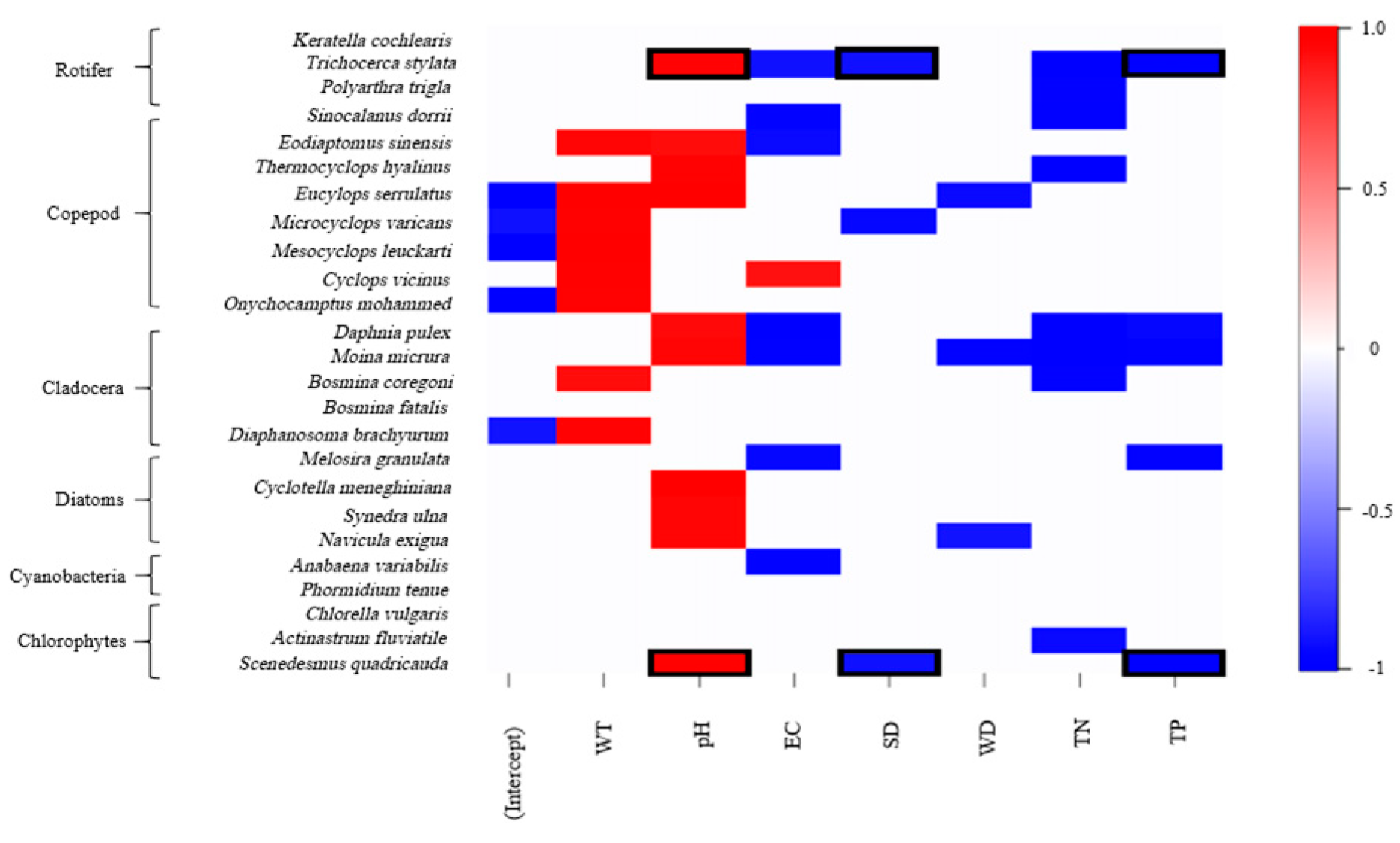
3.4. Drivers of Zooplankton and Phytoplankton Co-Occurrence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Freilich, M.A.; Wieters, E.; Broitman, B.R.; Marquet, P.A.; Navarrete, S.A. Species co-occurrence networks: Can they reveal trophic and non-trophic interactions in ecological communities? Ecology 2018, 99, 690–699. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.P.; Gonçalves, D.V.; Garcia-Raventós, A.; Lopes-Lima, M.; Varandas, S.; Froufe, E.; Teixeira, A.; Hui, F.K.C.; Filipe, A.F.; Sousa, R. Joint species distribution models unveil co-occurrences between freshwater mussels and their fish hosts. J. Biogeogr. 2023, 50, 730–742. [Google Scholar] [CrossRef]
- Aspin, T.; House, A. Alpha and beta diversity and species co-occurrence patterns in headwaters supporting rare intermittent-stream specialists. Freshw. Biol. 2022, 67, 1188–1202. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, L.; Jiang, Y.; Wang, H.; Jiguet, F. Unravelling species co-occurrence in a steppe bird community of inner mongolia: Insights for the conservation of the endangered jankowski’s bunting. Divers. Distrib. 2020, 26, 843–852. [Google Scholar] [CrossRef]
- D’Amen, M.; Mod, H.K.; Gotelli, N.J.; Guisan, A. Disentangling biotic interactions, environmental filters, and dispersal limitation as drivers of species co-occurrence. Ecography 2018, 41, 1233–1244. [Google Scholar] [CrossRef]
- Trivellone, V.; Bougeard, S.; Giavi, S.; Krebs, P.; Balseiro, D.; Dray, S.; Moretti, M. Factors shaping community assemblages and species co-occurrence of different trophic levels. Ecol. Evol. 2017, 7, 4745–4754. [Google Scholar] [CrossRef]
- Dutkiewicz, S.; Follett, C.L.; Follows, M.J.; Henderikx-Freitas, F.; Ribalet, F.; Gradoville, M.R.; Coesel, S.N.; Farnelid, H.; Finkel, Z.V.; Irwin, A.J. Multiple biotic interactions establish phytoplankton community structure across environmental gradients. Limnol. Oceanogr. 2024, 69, 1086–1100. [Google Scholar] [CrossRef]
- Inoue, K.; Stoeckl, K.; Geist, J. Joint species models reveal the effects of environment on community assemblage of freshwater mussels and fishes in european rivers. Divers. Distrib. 2017, 23, 284–296. [Google Scholar] [CrossRef]
- Ovaskainen, O.; Abrego, N. Joint Species Distribution Modelling: With Applications in R; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Pollock, L.J.; Tingley, R.; Morris, W.K.; Golding, N.; O’Hara, R.B.; Parris, K.M.; Vesk, P.A.; McCarthy, M.A. Understanding co-occurrence by modelling species simultaneously with a joint species distribution model (jsdm). Methods Ecol. Evol. 2014, 5, 397–406. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, T.; Yang, J.; Zhang, H.; Yang, L.; Li, Q.; Hou, N. Recovery capacity of constructed wetlands in response to multiple disturbances: Microbial interaction perspective. Bioresour. Technol. 2024, 408, 131155. [Google Scholar] [CrossRef]
- Reinl, K.L.; Harris, T.D.; Elfferich, I.; Coker, A.; Zhan, Q.; Domis, L.N.D.S.; Morales-Williams, A.M.; Bhattacharya, R.; Grossart, H.-P.; North, R.L.; et al. The role of organic nutrients in structuring freshwater phytoplankton communities in a rapidly changing world. Water Res. A J. Int. Water Assoc. 2022, 219, 118573. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.K.; Elmarsafy, M.; Vucic, J.M.; Teillet, M.; Pretty, T.J.; Cohen, R.S.; Huynh, M. Which physicochemical variables should zooplankton ecologists measure when they conduct field studies? J. Plankton Res. 2021, 43, 180–198. [Google Scholar] [CrossRef]
- Ebrahimzadeh, G.; Alimohammadi, M.; Kahkah, M.R.R.; Mahvi, A.H. Relationship between algae diversity and water quality–A case study: Chah niemeh reservoir southeast of Iran. J. Environ. Health Sci. Eng. 2021, 19, 437–443. [Google Scholar] [CrossRef]
- Mougi, A. Ph adaptation stabilizes bacterial communities. npj Biodiversity 2024, 3, 32. [Google Scholar] [CrossRef]
- Maraón, E.; Fernández-González, C.; Tarran, G.A. Effect of temperature, nutrients and growth rate on picophytoplankton cell size across the atlantic ocean. Sci. Rep. 2024, 14, 28034. [Google Scholar]
- Zhao, P.Z.; Ouyang, L.L.; Shen, A.L.; Wang, Y.L. The cell cycle of phytoplankton: A review. J. World Aquac. Soc. 2022, 53, 799–815. [Google Scholar] [CrossRef]
- Declerck, S.A.; de Senerpont Domis, L.N. Contribution of freshwater metazooplankton to aquatic ecosystem services: An overview. Hydrobiologia 2023, 850, 2795–2810. [Google Scholar] [CrossRef]
- Gray, D.K.; Arnott, S.E.; Shead, J.A.; Derry, A.M. The recovery of acid-damaged zooplankton communities in Canadian lakes: The relative importance of abiotic, biotic and spatial variables. Freshw. Biol. 2012, 57, 741–758. [Google Scholar] [CrossRef]
- Cavan, E.L.; Henson, S.A.; Belcher, A.; Sanders, R. Role of zooplankton in determining the efficiency of the biological carbon pump. Biogeosciences 2017, 14, 177–186. [Google Scholar] [CrossRef]
- McClain, C.R.; Barry, J.P. Habitat heterogeneity, disturbance, and productivity work in concert to regulate biodiversity in deep submarine canyons. Ecology 2010, 91, 964–976. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M.; Capps, K.A.; Allan, J.D.; Castillo, M.M.; Capps, K.A. Energy flow and nutrient cycling in aquatic communities. In Stream Ecology: Structure and Function of Running Waters; Springer: Cham, Switherland, 2021; pp. 357–381. [Google Scholar]
- Xie, C.; Dai, B.; Wu, J.; Liu, Y.; Jiang, Z. Initial recovery of fish faunas following the implementation of pen-culture and fishing bans in floodplain lakes along the yangtze river. J. Environ. Manag. 2022, 319, 115743. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lu, Y.; Wu, P.; Ma, X.; Zhou, L. Spatial-temporal analysis of landscape ecological risk in different seasons during the past 30 years in lake shengjin wetland, lower reaches of the yangtze river. J. Lake Sci. 2020, 32, 813–825. [Google Scholar]
- Rennella, A.M.; Geronazzo, M.D.; Romero, M.; Boveri, M.; Rosso, J.J. Hydrological variability, zooplankton availability and the shift between planktivore-benthivore feeding behaviour in the visual predator fish, odontesthes bonariensis. Environ. Biol. Fishes 2019, 102, 713–725. [Google Scholar] [CrossRef]
- Wentzky, V.C.; Tittel, J.; Jäger, C.G.; Bruggeman, J.; Rinke, K. Seasonal succession of functional traits in phytoplankton communities and their interaction with trophic state. J. Ecol. 2020, 108, 1649–1663. [Google Scholar] [CrossRef]
- Dakos, V.; Matthews, B.; Hendry, A.P.; Levine, J.; Loeuille, N.; Norberg, J.; Nosil, P.; Scheffer, M.; De Meester, L. Ecosystem tipping points in an evolving world. Nat. Ecol. Evol. 2019, 3, 355–362. [Google Scholar] [CrossRef]
- Song, S.; Wu, X.; Hou, J.; Peng, S.; Lin, X.; Ge, X.; Yan, D.; Lin, G. Monitoring the dynamics of aquatic vegetation in a typical shallow lake using the water bloom index algorithm—A case study in bao’an lake in the middle reaches of the yangtze river. Plants 2024, 13, 3090. [Google Scholar] [CrossRef]
- Yang, P.; Jiang, X.; Xie, Z.; Heino, J. Eutrophication is better indicated by functional traits than taxonomic composition of macroinvertebrate assemblages in floodplain lakes. Biodivers. Conserv. 2024, 33, 4257–4274. [Google Scholar] [CrossRef]
- Cranston, A.; Cooper, N.; Bro-Jørgensen, J. Using joint species distribution modelling to identify climatic and non-climatic drivers of afrotropical ungulate distributions. Ecography 2024, 2024, e07209. [Google Scholar] [CrossRef]
- Feng, S.; Wang, H.; Zhang, J.; Zhao, X.; Zhao, J.; Mao, F.; Peng, W.; Chen, Q. Effect of environmental factors on macrobenthic community structure in chishui river basin. Sustainability 2025, 17, 469. [Google Scholar] [CrossRef]
- Zhou, B.; Yuan, Y.; Jiang, L.; Sun, M.; Zhou, Z.; Wang, Y. Characteristics of plankton community structures and environmental factors in typical water bodies of eastern China. Ecol. Front. 2025, 45, 239–247. [Google Scholar] [CrossRef]
- Yuan, H.; Xiao, Y.; Xie, Y.; Luo, H.; Liu, J.; Xu, J.; Wu, X.; Chen, D.; Niu, Y. Structural characteristics of plankton community in dongting lake and its relationship with water environmental factors. Sci. Rep. 2024, 14, 28189. [Google Scholar] [CrossRef] [PubMed]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.A.; Guisan, A. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef]
- Bhandarkar, S.; Paliwal, G. Trophic status in freshwater lentic ecosystem of dhukeshwari temple pond deori with reference to zooplanktonic assemblage. Int. J. Environ. Rehabil. Conserv. 2017, 8, 145–159. [Google Scholar]
- Sommer, U.; Sommer, F. Cladocerans versus copepods: The cause of contrasting top–down controls on freshwater and marine phytoplankton. Oecologia 2006, 147, 183–194. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Deng, D.; Zhao, X.; Zhou, Z. Temporal and spatial variations in phytoplankton: Correlations with environmental factors in Shengjin lake, China. Environ. Sci. Pollut. Res. 2015, 22, 14144–14156. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, L.Z.; Lu, Y.; Wang, J. Dynamic ecosystem service value in the wetland of Shengjin lake national nature reserve. J. Anhui Agric. Univ. 2018, 45, 909–915. [Google Scholar]
- Hu, J.; Wang, Y.; Jin, Y.; Xiang, C.; Wu, L.; Xu, Y. High-resolution paleoclimatic changes recorded in lake Shengjin, lower reaches of the Yangtze river over the last millennium. Quat. Sci. 2022, 42, 421–434. [Google Scholar]
- Anhui Provincial Department of Ecology and Environment. Anhui Ecological and Environmental Statistical Bulletin 2023 [EB/OL]. (2024-12-17). Available online: https://sthjt.ah.gov.cn/public/21691/122622271.html (accessed on 9 May 2025).
- Gong, X.-H.; Ding, Q.-Q.; Jin, M.; Xue, B.; Zhang, L.; Yao, S.-C.; Wang, Z.; Lu, S.; Zhao, Z. Screening of priority pollutants and risk assessment for surface water from Shengjin lake. Huan Jing Ke Xue Huanjing Kexue 2021, 42, 4727–4738. [Google Scholar]
- Peng, L.; Dong, B.; Wang, P.; Sheng, S.; Sun, L.; Fang, L.; Li, H.; Liu, L. Research on ecological risk assessment in land use model of Shengjin lake in Anhui province, China. Environ. Geochem. Health 2019, 41, 2665–2679. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, J. Influence of the sluice on water level and area of Yangtze river-connected lakes: A case study in Shengjin lake. J. Water Resour. Water Eng 2018, 29, 47–52. [Google Scholar]
- Wang, H.-H.; Wang, J.; Cui, Y.-H. Remote sensing monitoring on spatial differentiation of suspended sediment concentration in a river-lake system based on sentinel-2 msi imaging: A case for Shengjin lake and connected Yangtze river section in Anhui province. Huan Jing Ke Xue Huanjing Kexue 2020, 41, 1207–1216. [Google Scholar] [PubMed]
- Zhang, Z.S.; Huang, X.F. Methods for Study on Freshwater Plankton; Science Press: Beijing, China, 1991. (In Chinese) [Google Scholar]
- Hu, H.; Wei, Y. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; China Science Publishing & Media: Beijing, China, 2006. [Google Scholar]
- State Environmental Protection Bureau (Sepb). Methods of Monitoring and Analysis for Water and Wastewater, 4th ed.; China Environmental Science Press: Beijing, China. (In Chinese)
- Yang, S.; Han, X.; Zhang, C.; Sun, B.; Wang, X.; Shi, X. Seasonal changes in phytoplankton biomass and dominant species in the changjiang river estuary and adjacent seas: General trends based on field survey data 1959–2009. J. Ocean Univ. China 2014, 13, 926–934. [Google Scholar] [CrossRef]
- Meng, Z.; Chen, K.; Liu, L.; Hu, F.; Zhu, Y.; Li, X.; Yang, D. Niche and interspecific association with respect to the dominant phytoplankton species in different hydrological periods of lake Wuchang, China. Front. Environ. Sci. 2022, 10, 985672. [Google Scholar] [CrossRef]
- Hou, C.W.; Sun, X.Y.; Liu, Y.L.; Zhang, C.; Zhang, W.; Zhao, J.; Dong, Z. Spatial niches of dominant zooplankton species in the Yantai offshore waters. Acta Ecol. Sin. 2020, 40, 5822–5833. [Google Scholar]
- Chi, S.; Li, M.; Zheng, J.; Chen, S.; Chen, M.; Hu, J.; Tang, J.; Hu, S.; Dong, F.; Zhao, X. Macroinvertebrate communities in the big east lake water network in relation to environmental factors. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 22. [Google Scholar] [CrossRef]
- Ovaskainen, O.; Tikhonov, G.; Norberg, A.; Guillaume Blanchet, F.; Duan, L.; Dunson, D.; Roslin, T.; Abrego, N. How to make more out of community data? A conceptual framework and its implementation as models and software. Ecol. Lett. 2017, 20, 561–576. [Google Scholar] [CrossRef]
- Warton, D.I.; Blanchet, F.G.; O’Hara, R.B.; Ovaskainen, O.; Taskinen, S.; Walker, S.C.; Hui, F.K. So many variables: Joint modeling in community ecology. Trends Ecol. Evol. 2015, 30, 766–779. [Google Scholar] [CrossRef]
- Tikhonov, G.; Opedal, Ø.H.; Abrego, N.; Lehikoinen, A.; de Jonge, M.M.; Oksanen, J.; Ovaskainen, O. Joint species distribution modelling with the R-package hmsc. Methods Ecol. Evol. 2020, 11, 442–447. [Google Scholar] [CrossRef]
- Simko, T.; Wei, T. R Package “Corrplot”: Visualization of a Correlation Matrix, Version 0.88. 2021. Available online: https://github.com/taiyun/corrplot (accessed on 11 May 2025).
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘ggplot2’. Create elegant data visualisations using the grammar of graphics. Version 2016, 2, 1–189. [Google Scholar]
- Dong, A.; Yu, X.; Yin, Y.; Zhao, K. Seasonal variation characteristics and the factors affecting plankton community structure in the Yitong river, China. Int. J. Environ. Res. Public Health 2022, 19, 17030. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Wang, G.; Liang, Y.; Li, C.; Svenning, J.C. How do rotifer communities respond to floating photovoltaic systems in the subsidence wetlands created by underground coal mining in China? J. Environ. Manag. 2023, 339, 117816. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.E.; Fischer, J.M.; Bollens, S.M.; Overholt, E.P.; Breckenridge, J.K. Toward a more comprehensive theory of zooplankton diel vertical migration: Integrating ultraviolet radiation and water transparency into the biotic paradigm. Limnol. Oceanogr. 2011, 56, 1603–1623. [Google Scholar] [CrossRef]
- Kiørboe, T. How zooplankton feed: Mechanisms, traits and trade-offs. Biol. Rev. 2011, 86, 311–339. [Google Scholar] [CrossRef] [PubMed]
- Hairston, H.N.G. Food limitation in a wild cyclopoid copepod population: Direct and indirect life history responses. Oecologia 1998, 115, 320–330. [Google Scholar]
- Gilbert, J.J. Food niches of planktonic rotifers: Diversification and implications. Limnol. Oceanogr. 2022, 67, 2218–2251. [Google Scholar] [CrossRef]
- Liu, Y.; Tackx, M.; Dauta, A.; Julien, F.; Buffan-Dubau, E. Rotifers stimulate the specific uptake rate in lotic phototrophic biofilms. Freshw. Biol. 2021, 66, 1245–1256. [Google Scholar] [CrossRef]
- Huo, X.-K.; Wang, Y.-G.; Zhou, L.-T.; Wang, S.-H.; Jiang, X.; Chen, K.; Wang, P.-F. Characterization of the ecological niche and interspecific connectivity of plankton in Baiyangdian lake by combining ecological networks. Huan Jing Ke Xue Huanjing Kexue 2024, 45, 5298–5307. [Google Scholar]
- Lampman, G.G.; Makarewicz, J.C. The phytoplankton zooplankton link in the lake ontario food web. J. Great Lakes Res. 1999, 25, 239–249. [Google Scholar] [CrossRef]
- Jakhar, P. Role of phytoplankton and zooplankton as health indicators of aquatic ecosystem: A review. Int. J. Innov. Res. Study 2013, 2, 489–500. [Google Scholar]
- Sommer, U.; Sommer, F.; Santer, B.; Jamieson, C.; Boersma, M.; Becker, C.; Hansen, T. Complementary impact of copepods and cladocerans on phytoplankton. Ecol. Lett. 2001, 4, 545–550. [Google Scholar] [CrossRef]
- De Meester, L.; Declerck, S.A.; Ger, K.A. Beyond daphnia: A plea for a more inclusive and unifying approach to freshwater zooplankton ecology. Hydrobiologia 2023, 850, 4693–4703. [Google Scholar] [CrossRef]
- Amorim, C.A.; do Nascimento Moura, A. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef] [PubMed]
- Gliwicz, Z.M. Relative significance of direct and indirect effects of predation by planktivorous fish on zooplankton. Hydrobiologia 1994, 272, 201–210. [Google Scholar] [CrossRef]
- Bai, X.; Jiang, Z.; Fang, Y.; Zhu, L.; Feng, J. Effects of environmental concentrations of total phosphorus on the plankton community structure and function in a microcosm study. Int. J. Environ. Res. Public Health 2022, 19, 8412. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zooplankton | Species | Dominant | Phytoplankton | Species | Dominant |
|---|---|---|---|---|---|
| Rotifer | Keratella cochlearis | 0.059 | Cyanobacteria | Phormidium tenue | 0.082 |
| Trichocerca stylata | 0.056 | Anabaena variabilis | 0.096 | ||
| Polyarthra trigla | 0.031 | Diatoms | Cyclotella meneghiniana | 0.043 | |
| Cladocera | Daphnia pulex | 0.028 | Navicula exigua | 0.074 | |
| Moina micrura | 0.062 | Synedra ulna | 0.033 | ||
| Bosmina coregoni | 0.055 | Melosira granulata | 0.402 | ||
| Bosmina fatalis | 0.031 | Chlorophytes | Chlorella vulgaris | 0.135 | |
| Diaphanosoma brachyurum | 0.034 | Actinastrum fluviatile | 0.021 | ||
| Copepod | Sinocalanus dorrii | 0.028 | Scenedesmus quadricauda | 0.037 | |
| Eodiaptomus sinensis | 0.026 | ||||
| Microcyclops varicans | 0.059 | ||||
| Mesocyclops leuckarti | 0.032 | ||||
| Thermocyclops hyalinus | 0.124 | ||||
| Eucylops serrulatus | 0.036 | ||||
| Cyclops vicinus | 0.022 | ||||
| Onychocamptus mohammed | 0.037 |
| Environmental Factors | Description | Range | Mean | SE |
|---|---|---|---|---|
| pH | pH | 7.44–9.03 | 8.29 | 0.15 |
| WT (°C) | Water temperature | 24.70–28.55 | 26.28 | 0.06 |
| EC (μs/cm) | Electric conductivity | 149.9–248.15 | 181.82 | 4.06 |
| SD (m) | Transparency | 0.3–0.93 | 0.49 | 2.48 |
| WD(m) | Water depth | 0.38–7.85 | 2.2 | 0.28 |
| TP (mg/L) | Total phosphorus concentration | 0.02–0.20 | 0.08 | 0.01 |
| TN (mg/L) | Total nitrogen concentration | 0.53–1.56 | 1.21 | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Liang, Y.; Wang, Y.; Xu, P.; Li, C. The Co-Occurrence of Zooplankton and Phytoplankton in Shengjin Lake, a Typical Yangtze-Connected Lake in China. Diversity 2025, 17, 382. https://doi.org/10.3390/d17060382
Shi Y, Liang Y, Wang Y, Xu P, Li C. The Co-Occurrence of Zooplankton and Phytoplankton in Shengjin Lake, a Typical Yangtze-Connected Lake in China. Diversity. 2025; 17(6):382. https://doi.org/10.3390/d17060382
Chicago/Turabian StyleShi, Yuxin, Yue Liang, Yutao Wang, Peng Xu, and Chunlin Li. 2025. "The Co-Occurrence of Zooplankton and Phytoplankton in Shengjin Lake, a Typical Yangtze-Connected Lake in China" Diversity 17, no. 6: 382. https://doi.org/10.3390/d17060382
APA StyleShi, Y., Liang, Y., Wang, Y., Xu, P., & Li, C. (2025). The Co-Occurrence of Zooplankton and Phytoplankton in Shengjin Lake, a Typical Yangtze-Connected Lake in China. Diversity, 17(6), 382. https://doi.org/10.3390/d17060382