
Screening of Germination and Early Seedling Growth Reveal Contrasting Cu Tolerance of Chilean Quinoa Accessions Depending on Origin

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Germination Assays
2.3. Early Growth Assays in Petri Dishes
2.4. Growth Assays in Hydroponic Pouches
2.5. Statistical Analysis
3. Results
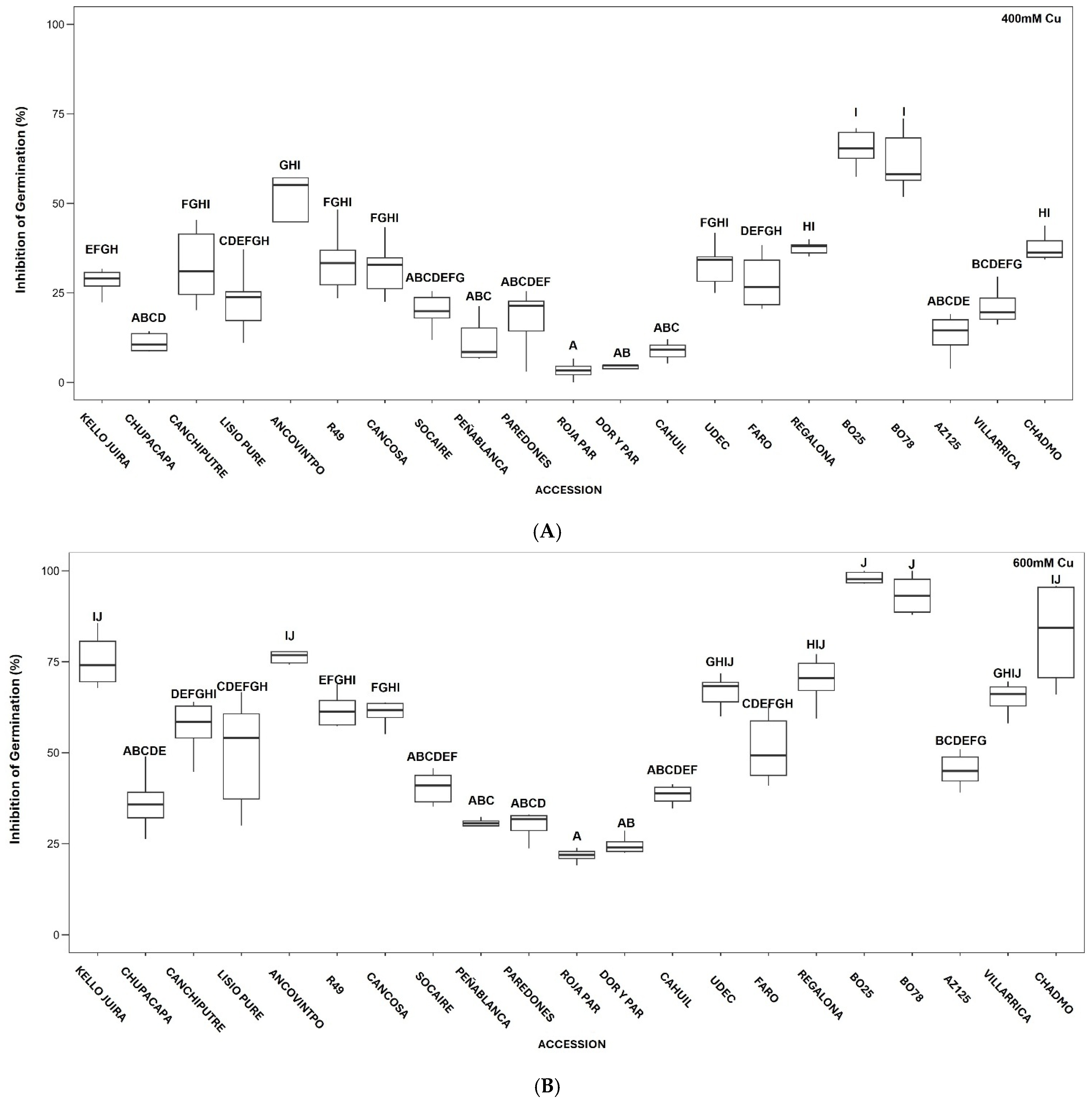
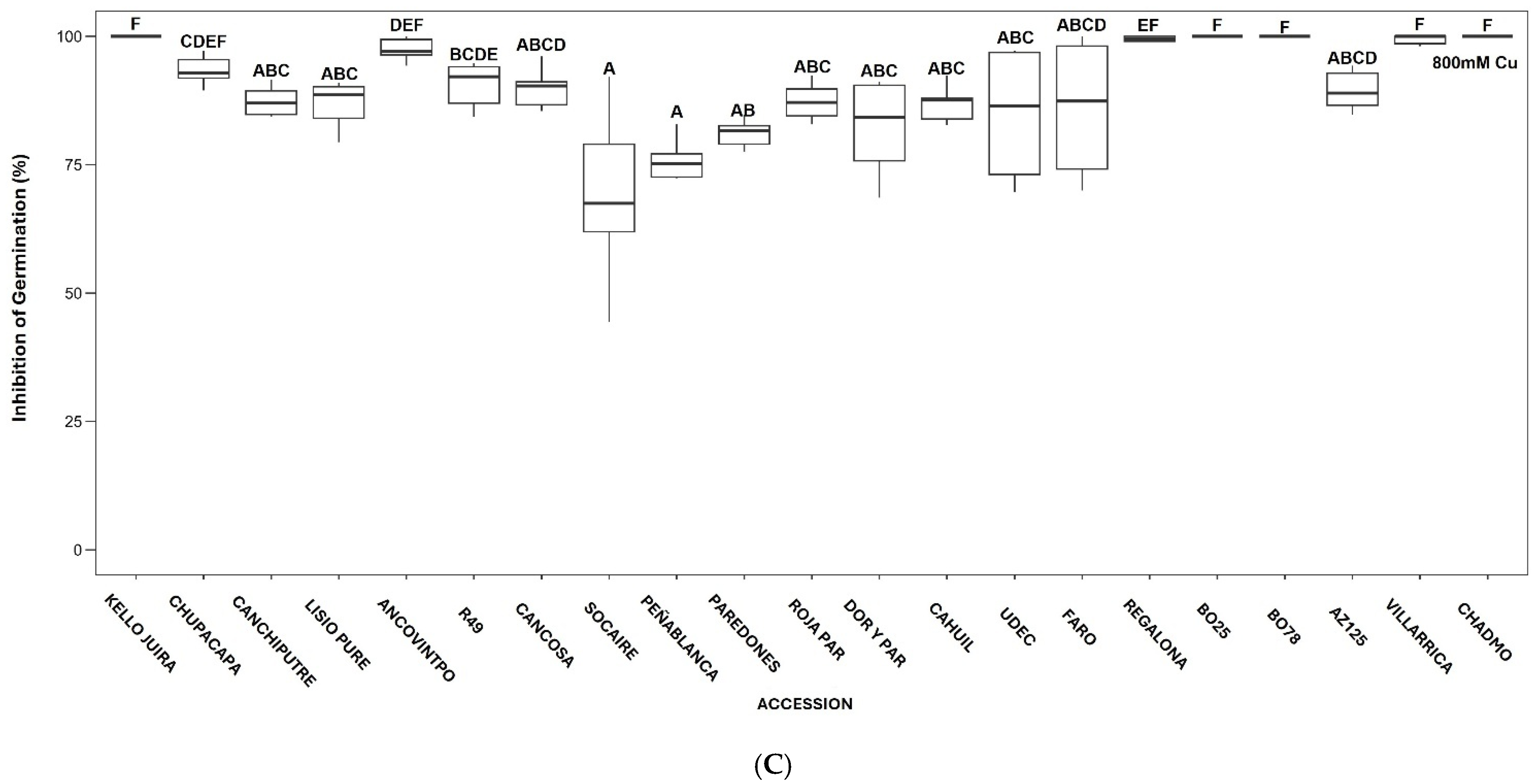
3.1. Effect of Cu on Seed Germination
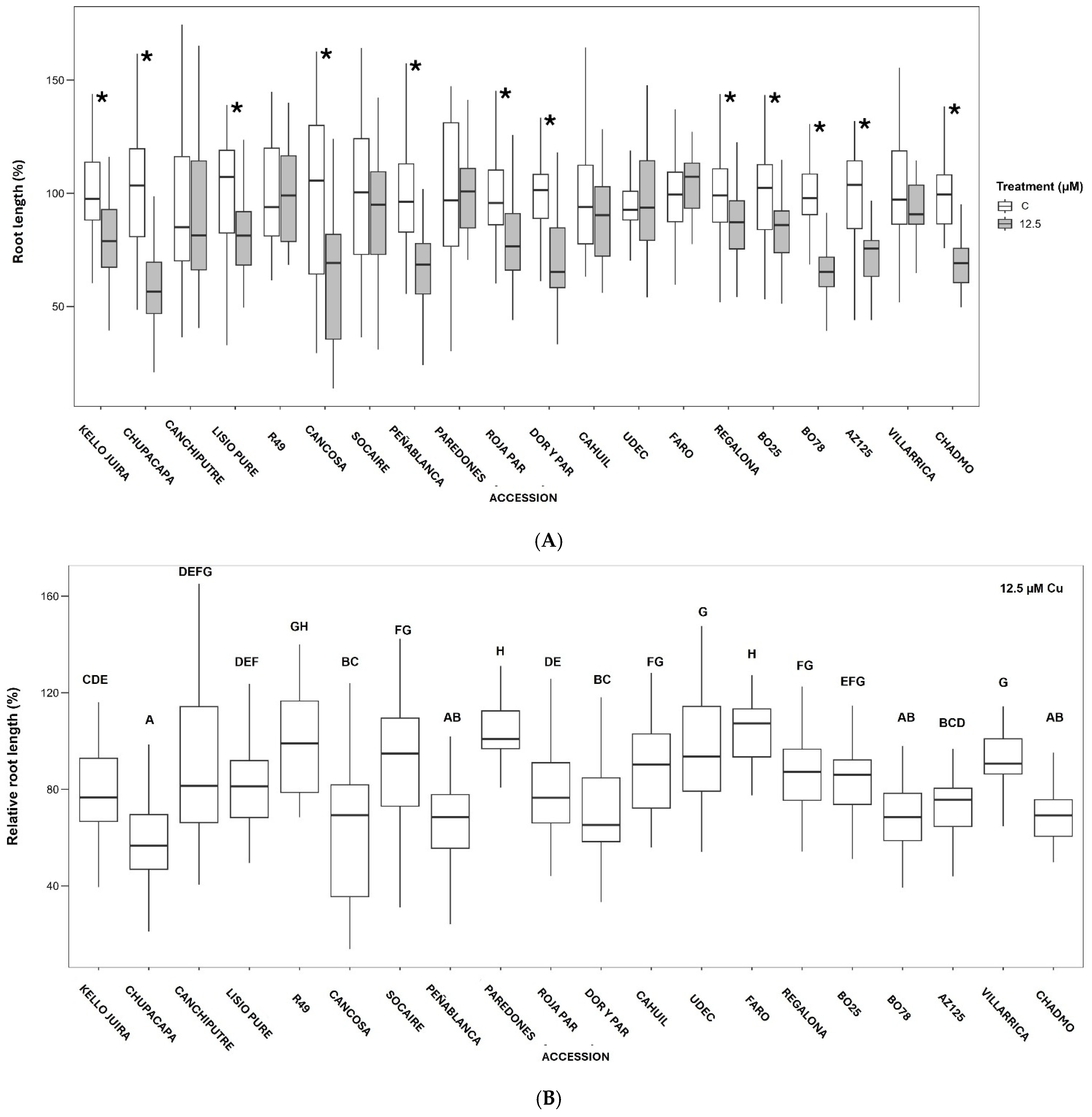
3.2. Effect of Cu on Early Seedling Growth in Petri Dishes
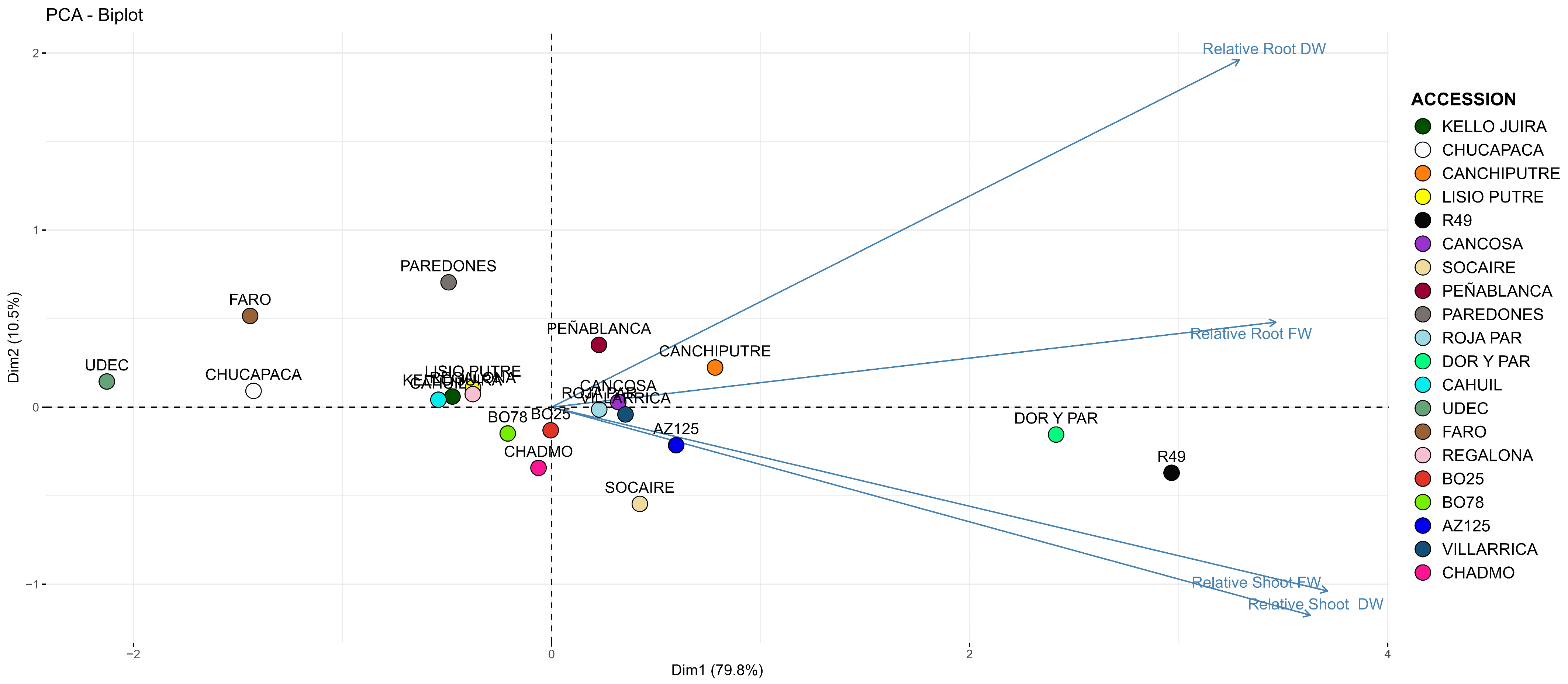
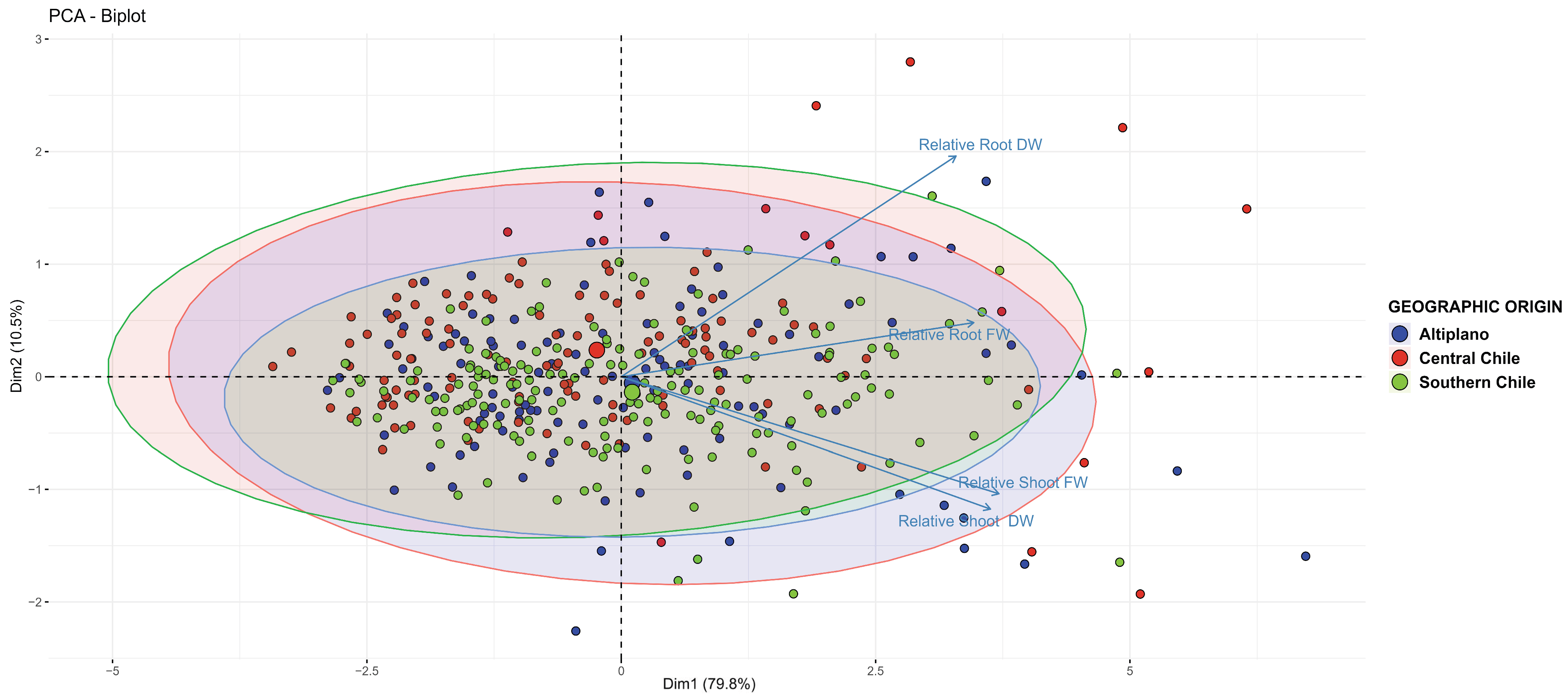
3.3. Effect of Cu on Plantlet Growth in Hydroponic Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GP | Germination Percentage |
| MGT | Mean Germination Time |
| GI | Germination Index |
| IG | Percent Inhibition of Germination |
| RRL | Relative Root Length |
| RRFW | Relative Root Fresh Weight |
| RRDW | Relative Root Dry Weight |
| RSFW | Relative Shoot Fresh Weight |
| RSDW | Relative Shoot Dry Weight |
| PCA | Principal Component Analysis |
References
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [PubMed]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in Biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [PubMed]
- Shabbir, Z.; Sardar, A.; Shabbir, A.; Abbas, G.; Shamshad, S.; Khalid, S.; Murtaza, G.; Dumat, C.; Shahid, M. Copper uptake, essentiality, toxicity, detoxification and risk assessment in soil-plant environment. Chemosphere 2020, 259, 127436. [Google Scholar] [CrossRef] [PubMed]
- Mir, A.R.; Pichtel, J.; Hayat, S. Copper: Uptake, toxicity and tolerance in plants and management of Cu-contaminated soils. Biometals 2021, 34, 737–759. [Google Scholar] [CrossRef]
- Ravet, K.; Pilon, M. Copper and iron homeostasis in plants: The challenges of oxidative stress. Antioxid. Redox Signal. 2013, 19, 919–932. [Google Scholar] [CrossRef]
- Rheman, M.; Liu, L.; Wang, Q.; Saleem, M.H.; Bashir, S.; Ullah, S.; Peng, D. Copper environmental toxicology, recent advances, and future outlook: A review. Environ. Sci. Pollut. Res. 2019, 26, 18003–18016. [Google Scholar] [CrossRef]
- Kumar, V.; Pandita, S.; Sidhu, G.P.S.; Sharma, A.; Khanna, K.; Kaur, P.; Bali, A.S.; Setia, R. Copper bioavailability, uptake, toxicity and tolerance in plants: A comprehensive review. Chemosphere 2021, 262, 127810. [Google Scholar] [CrossRef]
- Wairich, A.; De Conti, L.; Lamb, T.I.; Keil, R.; Neves, L.O.; Brunetto, G.; Sperotto, R.A.; Ricachenevsky, F.K. Throwing copper around: How Plants Control Uptake, distribution, and accumulation of copper. Agronomy 2022, 12, 994. [Google Scholar] [CrossRef]
- Chen, G.; Li, J.; Han, H.; Du, R.; Wang, X. Physiological and molecular mechanisms of plant responses to copper stress. Int. J. Mol. Sci. 2022, 23, 12950. [Google Scholar] [CrossRef]
- Adeleke, R.; Nwangburuka, C.; Oboirien, B. Origins, roles and fate of organic acids in soils: A review. S. Afr. J. Bot. 2017, 108, 393–406. [Google Scholar] [CrossRef]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-metal-induced reactive oxygen species: Phytotoxicity and physicochemical changes in plants. Rev. Environ. Contam. Toxicol. 2014, 232, 1–44. [Google Scholar] [PubMed]
- Xu, E.; Liu, Y.; Gu, D.; Zhan, X.; Li, J.; Zhou, K.; Zhang, P.; Zou, Y. Molecular mechanisms of plant responses to copper: From deficiency to stress. Int. J. Mol. Sci. 2024, 25, 6993. [Google Scholar] [PubMed]
- Miotto, A.; Ceretta, C.A.; Girotto, E.; Trentin, G.; Kaminski, J.; De Conti, L.; Moreno, T.; Elena, B.; Brunetto, G. Copper accumulation and availability in Sandy, acid, vineyard soils. Commun. Soil Sci. Plant Anal. 2017, 48, 1167–1183. [Google Scholar] [CrossRef]
- Ballabio, C.; Panagos, P.; Lugato, E.; Huang, J.-H.; Orgiazzi, A.; Jones, A.; Fernández-Ugalde, O.; Borrelli, P.; Montanarella, L. Copper distribution in European topsoils: An assessment based on LUCAS Soil Survey. Sci. Total Environ. 2018, 636, 282–298. [Google Scholar]
- Sereni, L.; Guenet, B.; Lamy, I. Mapping risks associated with soil copper contamination using availability and bio-availability proxies at the European scale. Environ. Sci. Pollut. Res. 2023, 30, 19828–19844. [Google Scholar]
- Adrees, M.; Ali, S.; Rizwan, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Zia-Ur-Rehman, M.; Irshad, M.K.; Bharwana, S.A. The effect of excess copper on growth and physiology of important food crops: A review. Environ. Sci. Pollut. Res. Int. 2015, 22, 8148–8162. [Google Scholar] [CrossRef]
- He, Z.; Shentu, J.; Yang, X.; Baligar, V.; Zhang, T.; Stoffella, P. Heavy metal contamination in soils: Sources, indicators and assessment. J. Environ. Indic. 2015, 9, 17–18. [Google Scholar]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Mantanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef]
- Fagnano, M.; Agrelli, D.; Pascale, A.; Adamo, A.; Fiorentino, N.; Rocco, C.; Pepe, O.; Ventorino, V. Copper accumulation in agricultural soils: Risks for the food chain and soil microbial populations. Sci. Total Environ. 2020, 734, 139434. [Google Scholar] [CrossRef]
- Verdejo, J.; Ginocchio, R.; Sauvé, S.; Salgado, E.; Neaman, A. Thresholds of copper phytotoxicity in field-collected agricultural soils exposed to copper mining activities in Chile. Ecotoxicol. Environ. Saft. 2015, 122, 171–177. [Google Scholar]
- Schoffer, J.T.; Aponte, H.; Neaman, A.; de la Fuente, L.M.; Arellano, E.C.; Gil, P.M.; Ginocchio, R. Copper content in soils and litter from fruit orchards in Central Chile and its relationship with soil microbial activity. Plant Soil Environ. 2022, 68, 115–128. [Google Scholar] [CrossRef]
- Casanova, M.; Salazar, O.; Seguel, O.; Luzio, W. The Soils of Chile; Hartemink, A.E., Ed.; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Angon, P.D.; Islam, M.S.; Shreejana, K.C.; Das, A.; Anjum, N.; Poudel, A.; Suchi, S.A. Sources, effects and present perspectives of heavy metals contamination: Soil, plants and human food chain. Heliyon 2024, 10, e28357. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, S.E.; Mujica, A.; Jensen, C.R. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Rev. Int. 2006, 19, 99–109. [Google Scholar]
- Vega-Gálvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martínez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef] [PubMed]
- Martínez, E.A.; Veas, E.; Jorquera, C.; San Martín, R.; Jara, P. Re-Introduction of quinoa into Arid Chile: Cultivation of two lowland races under extremely low irrigation. J. Agron. Crop Sci. 2009, 195, 1–10. [Google Scholar] [CrossRef]
- Bazile, D.; Jacobsen, S.E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Stolen, O. Quinoa—Morphology, phenology and prospects for its production in Europe. Eur. J. Agron. 1993, 2, 19–29. [Google Scholar] [CrossRef]
- Jacobsen, S.E. Adaptation of quinoa (Chenopodium quinoa) to Northern European agriculture: Studies on developmental pattern. Euphytica 1997, 96, 41–48. [Google Scholar] [CrossRef]
- Bendevis, M.A.; Sun, Y.; Rosenqvist, E.; Shabala, S.; Liu, F.; Jacobsen, S.E. Photoperiodic effects on short-pulse 14C assimilation and overall carbon and nitrogen allocation patterns in contrasting quinoa cultivars. Environ. Exp. Bot. 2014, 104, 9–15. [Google Scholar] [CrossRef]
- Bazile, D.; Bertero, D.; Nieto, C. (Eds.) Estado del Arte de la Quinua en El Mundo en 2013; FAO: Santiago de Chile, Chile; CIRAD: Montpellier, France, 2014; p. 724. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). Quinoa 2013 International Year. Available online: https://www.fao.org/quinoa-2013/en/ (accessed on 17 December 2024).
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef]
- Amjad, M.; Iqbal, M.; Abbas, G.; Farooq, A.B.U.; Naeem, M.A.; Imran, M.; Murtaza, B.; Nadeem, M.; Jacobsen, S.E. Assessment of cadmium and lead tolerance potential of quinoa (Chenopodium quinoa Willd) and its implications for phytoremediation and human health. Environ. Geochem. Health 2022, 44, 1487–1500. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Rajan, S.; Ohri, D. Genetic diversity for morphological and quality traits in quinoa (Chenopodium quinoa Willd.) germplasm. Genet. Resour. Crop Evol. 2007, 54, 167–173. [Google Scholar]
- Ellis, R.H.; Roberts, E.H. Improved equations for the prediction of seed longevity. Ann. Bot. 1981, 45, 13–30. [Google Scholar] [CrossRef]
- Brenchley, J.L.; Probert, R.J. Seed germination responses to some environmental factors in the seagrass Zostera capicorni from Eastern Australia. Aquat. Bot. 1998, 62, 177–188. [Google Scholar] [CrossRef]
- Baruah, N.; Mondal, S.C.; Farooq, M.; Gogoi, N. Influence of heavy metals on seed germination and seedling growth of wheat, pea and tomato. Water Air Soil Pollut. 2019, 230, 273. [Google Scholar]
- Sarma, B.; Devi, P.; Gogoi, N.; Devi, Y.M. Effects of cobalt induced stress in Triticum aestivum L. crop. AJAB 2014, 2, 137–147. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants Without Soil; Circular 347; University of California, College of Agriculture, Agricultural Experiment Station: Berkeley, CA, USA, 1950. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2020. Centro de Transferencia InfoStat, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, Argentina. Available online: http://www.infostat.com.ar (accessed on 17 December 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 7 January 2025).
- Kranner, I.; Colville, I. Metals and seeds: Biochemical and molecular implications and their significance for seed germination. Environ. Exp. Bot. 2011, 23, 93–105. [Google Scholar]
- Sanjosé, I.; Navarro-Roldán, F.; Infante-Izquierdo, M.D.; Martínez-Sagarra, G.; Devesa, J.A.; Polo, A.; Ramirez-Acosta, S.; Sánchez-Guillón, E.; Jiménez-Nieva, F.J.; Muñoz-Rodríguez, A.F. Accumulation and effects of heavy metals on the germination and growth of Salsoa vermiculata L. seedlings. Diversity 2021, 13, 539. [Google Scholar] [CrossRef]
- Vasilachi-Mitoresu, I.C.; Stoleru, V.; Gavrilescu, M. Integrated assessment of Pb(II) and Cu(II) metal ion phytotoxicity on Medicago sativa L., Triticum aestivum L., and Zea mays L. plants: Insights into germination inhibition, seedling development, and ecosystem health. Plants 2023, 12, 3754. [Google Scholar] [CrossRef]
- Tapia-Gatica, J.; Selles, I.; Bravo, M.A.; Tessini, C.; Barros-Parada, W.; Novoselov, A.; Neaman, A. Global issues in setting legal limits on soil metal contamination: A case study of Chile. Chemosphere 2022, 290, 133404. [Google Scholar]
- Mondaca, P.; Catrin, J.; Verdejo, J.; Sauvé, S.; Neaman, A. Advances on the determination of thresholds of Cu phytotocity in field-contaminated soils in Central Chile. Environ. Pollut. 2017, 223, 146–152. [Google Scholar]
- Bosnic, D.; Bosnic, P.; Nikolic, D.; Nikolic, M.; Samardzic, J. Silicon and iron differently alleviate copper toxicity in cucumber leaves. Plants 2019, 8, 554. [Google Scholar] [CrossRef] [PubMed]
- Ben Massoud, M.; Sakohui, L.; Chaoui, A. Effects of plant growth regulators, calcium and citric acid on copper toxicity on pea seedlings. J. Plant Nutr. 2019, 42, 1230–1242. [Google Scholar]
- De Conti, L.; Cesco, S.; Mimmo, T.; Pii, Y.; Valentinuzzi, F.; Melo, G.W.B.; Ceretta, C.A.; Trentin, E.; Marques, A.C.R.; Brunetto, G. Iron fertilization to enhance tolerance mechanisms to copper toxicity of ryegrass plants used as cover crop in vineyards. Chemosphere 2020, 243, 125298. [Google Scholar] [PubMed]
- Yadav, P.; Kaur, R.; Kohli, S.K.; Sirhindi, G.; Bhardwaj, R. Castasterone assisted accumulation of polyphenols and antioxidant to increase tolerance of B. juncea plants towards copper toxicity. Cogent. Food Agric. 2016, 2, 1276821. [Google Scholar]
- Tapia, J.; Mulet, J.; Poblete, N.; Rodríguez, C. Catastro de estudios de geoquímica Ambiental de suelos y sedimentos de Chile. In Proceedings of the XIV Congreso Geológico Chileno, La Serena, Chile, 4–8 October 2015. [Google Scholar]
- Informe País. Estado del Medio Ambiente en Chile. 2018. Universidad de Chile. Instituto de Asuntos Públicos. Centro de Análisis de Políticas Públicas. Available online: https://repositorio.uchile.cl/bitstream/handle/2250/179262/Capitulo5-Suelos.pdf?sequence=1 (accessed on 7 October 2024).
- Novello, N.; Ferfuia, C.; Pasković, I.; Fabris, A.; Baldini, M.; Schat, H.; Pošćić, F. Independent variation in copper tolerance and copper accumulation among crop species and varieties. Plant Physiol. Biochem. 2020, 156, 538–551. [Google Scholar]
- Jeon, D. Evaluating the impact of copper concentrations on apple variety performance: Insights from a callus media system. Sustainability 2024, 16, 9741. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Pérez-Romero, J.A.; Mesa-Marín, J.; López-Jurado, J.; Redondo-Gómez, S. Inter-population differences tolerance to Cu excess during the initial phases of Juncus acutus life cycle: Implications for the design of metal restoration strategies. Int. J. Phytoremediation 2019, 21, 550–555. [Google Scholar]
- Purmale-Trasune, L.; Jēkabsone, A.; Andersone, U.; Karlsons, A.; Osvalde, A.; Levins, G. Comparison of in vitro and in planta heavy metal tolerance and accumulation potential of different Armeria maritima accessions from a dry Coastal Meadow. Plants 2022, 11, 2104. [Google Scholar] [CrossRef]
- Scoccianti, V.; Bucchini, A.E.; Iacobucci, M.; Ruiz, K.B.; Biondi, S. Oxidative stress and antioxidant responses to increasing concentrations of trivalent chromium in the Andean crop species Chenopodium quinoa Willd. Ecotoxicol. Environ. Saf. 2016, 133, 25–35. [Google Scholar]
- Gao, T.; Wang, H.; Li, C.; Zuo, M.; Wang, X.; Liu, Y.; Yang, Y.; Xu, D.; Liu, Y.; Fang, X. Effects of heavy metal stress on physiology, hydraulics, and anatomy of three desert plants in the Jinchang mining area, China. Int. J. Environ. Res. Public Health 2022, 19, 15873. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Root FW | Relative Root DW | Relative Shoot FW | Relative Shoot DW | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | Mean | ± | S.E. | Mean | ± | S.E. | Mean | ± | S.E. | Mean | ± | S.E. |
| KELLO JUIRA | 39 | 3 * bcd | 73 | 7 bcdef | 54 | 5 * cd | 67 | 6 * defg | ||||
| CHUCAPACA | 32 | 5 * ab | 55 | 7 * ab | 37 | 2 * ab | 51 | 3 * abc | ||||
| CANCHI PUTRE | 61 | 5 * fg | 94 | 8 fg | 73 | 5 * ef | 79 | 6 efg | ||||
| LISIO PUTRE | 47 | 5 * cdef | 72 | 7 * bcdef | 57 | 4 * cde | 62 | 5 * bcde | ||||
| R49 | 55 | 15 bcdef | 136 | 11 h | 123 | 13 g | 131 | 11 h | ||||
| CANCOSA | 54 | 5 * defg | 82 | 6 * cdefg | 70 | 5 * de | 74 | 6 * defg | ||||
| SOCAIRE | 50 | 5 * cdef | 70 | 8 bcde | 72 | 7 * de | 92 | 10 fg | ||||
| PEÑABLANCA | 55 | 4 * efg | 88 | 7 efg | 61 | 3 * cde | 71 | 4 defg | ||||
| PAREDONES | 34 | 5 * abc | 96 | 12 defg | 49 | 5 * bc | 59 | 5 * bcd | ||||
| ROJA PAR | 53 | 7 * defg | 79 | 10 cdefg | 70 | 8 * de | 71 | 7 defg | ||||
| DOR PAR | 83 | 9 g | 108 | 10 gh | 98 | 6 fg | 114 | 9 h | ||||
| CAHUIL | 44 | 2 * bcde | 68 | 4 * bcde | 52 | 3 * cd | 64 | 3 cde | ||||
| UDEC9 | 26 | 2 * a | 43 | 4 * a | 32 | 3 * a | 30 | 5 * a | ||||
| FARO | 26 | 2 * a | 72 | 5 * bcdef | 31 | 2 * a | 50 | 3 * ab | ||||
| REGALONA | 47 | 3 * def | 71 | 5 * bcdef | 53 | 3 * cd | 67 | 4 def | ||||
| BO25 | 53 | 5 * def | 69 | 6 * bcd | 66 | 6 * cde | 70 | 6 * def | ||||
| BO78 | 42 | 3 * bcd | 70 | 4 * bcde | 60 | 4 * cde | 75 | 4 * efg | ||||
| AZ125 | 51 | 4 * def | 83 | 6 cdef | 77 | 5 e | 85 | 4 g | ||||
| VILLARRICA | 58 | 5 * efg | 77 | 6 cdef | 67 | 5 de | 77 | 5 efg | ||||
| CHADMO | 51 | 5 * def | 62 | 6 * bc | 67 | 6 * de | 72 | 7 defg | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustamante Álvarez, C.; Pinto Pizarro, I.; Zúñiga-Contreras, E.; Bascuñán-Godoy, L.; Ostria-Gallardo, E.; Coba de la Peña, T. Screening of Germination and Early Seedling Growth Reveal Contrasting Cu Tolerance of Chilean Quinoa Accessions Depending on Origin. Diversity 2025, 17, 229. https://doi.org/10.3390/d17040229
Bustamante Álvarez C, Pinto Pizarro I, Zúñiga-Contreras E, Bascuñán-Godoy L, Ostria-Gallardo E, Coba de la Peña T. Screening of Germination and Early Seedling Growth Reveal Contrasting Cu Tolerance of Chilean Quinoa Accessions Depending on Origin. Diversity. 2025; 17(4):229. https://doi.org/10.3390/d17040229
Chicago/Turabian StyleBustamante Álvarez, Catalina, Isaac Pinto Pizarro, Estrella Zúñiga-Contreras, Luisa Bascuñán-Godoy, Enrique Ostria-Gallardo, and Teodoro Coba de la Peña. 2025. "Screening of Germination and Early Seedling Growth Reveal Contrasting Cu Tolerance of Chilean Quinoa Accessions Depending on Origin" Diversity 17, no. 4: 229. https://doi.org/10.3390/d17040229
APA StyleBustamante Álvarez, C., Pinto Pizarro, I., Zúñiga-Contreras, E., Bascuñán-Godoy, L., Ostria-Gallardo, E., & Coba de la Peña, T. (2025). Screening of Germination and Early Seedling Growth Reveal Contrasting Cu Tolerance of Chilean Quinoa Accessions Depending on Origin. Diversity, 17(4), 229. https://doi.org/10.3390/d17040229